
Diversity and Adaptation of Currently Grown Wheat Landraces and Modern Germplasm in Afghanistan, Iran, and Turkey

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
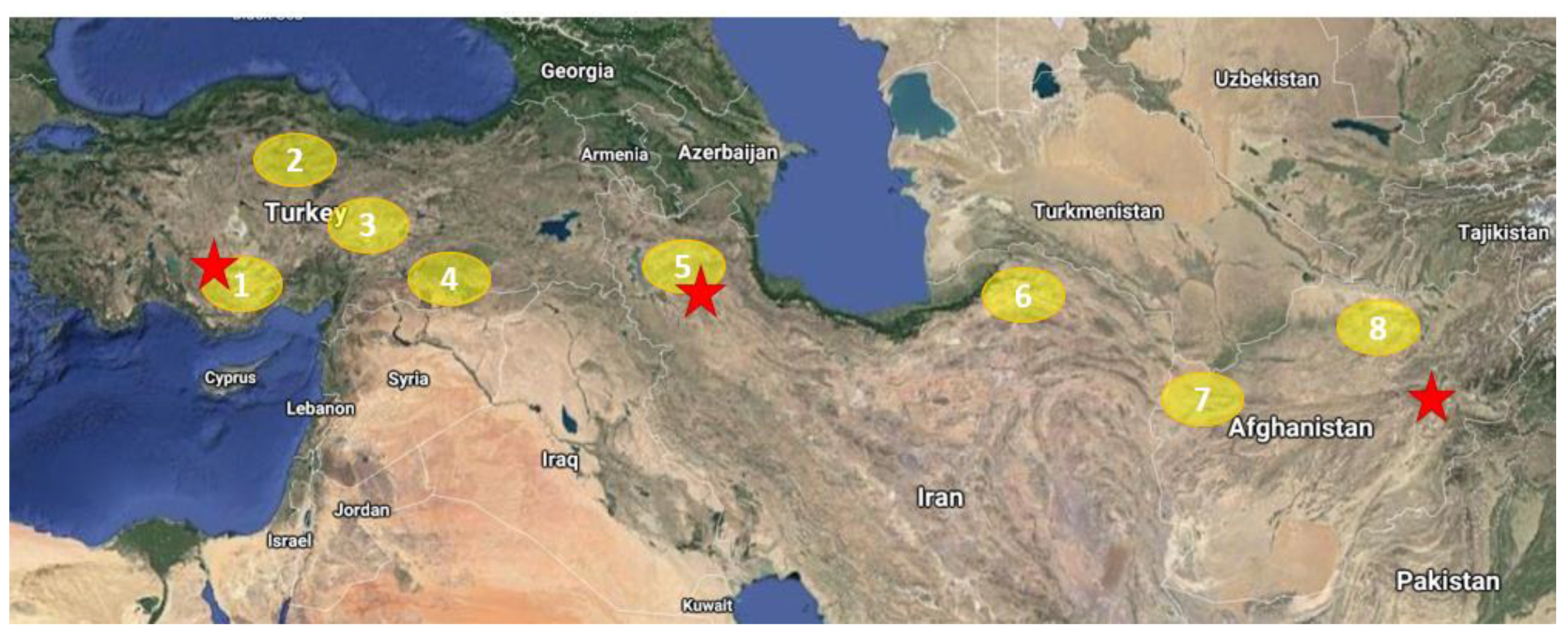
2.1. Material Used in the Study
2.2. Field Trials and Phenotyping
2.3. DNA Diversity and Molecular Markers
3. Results
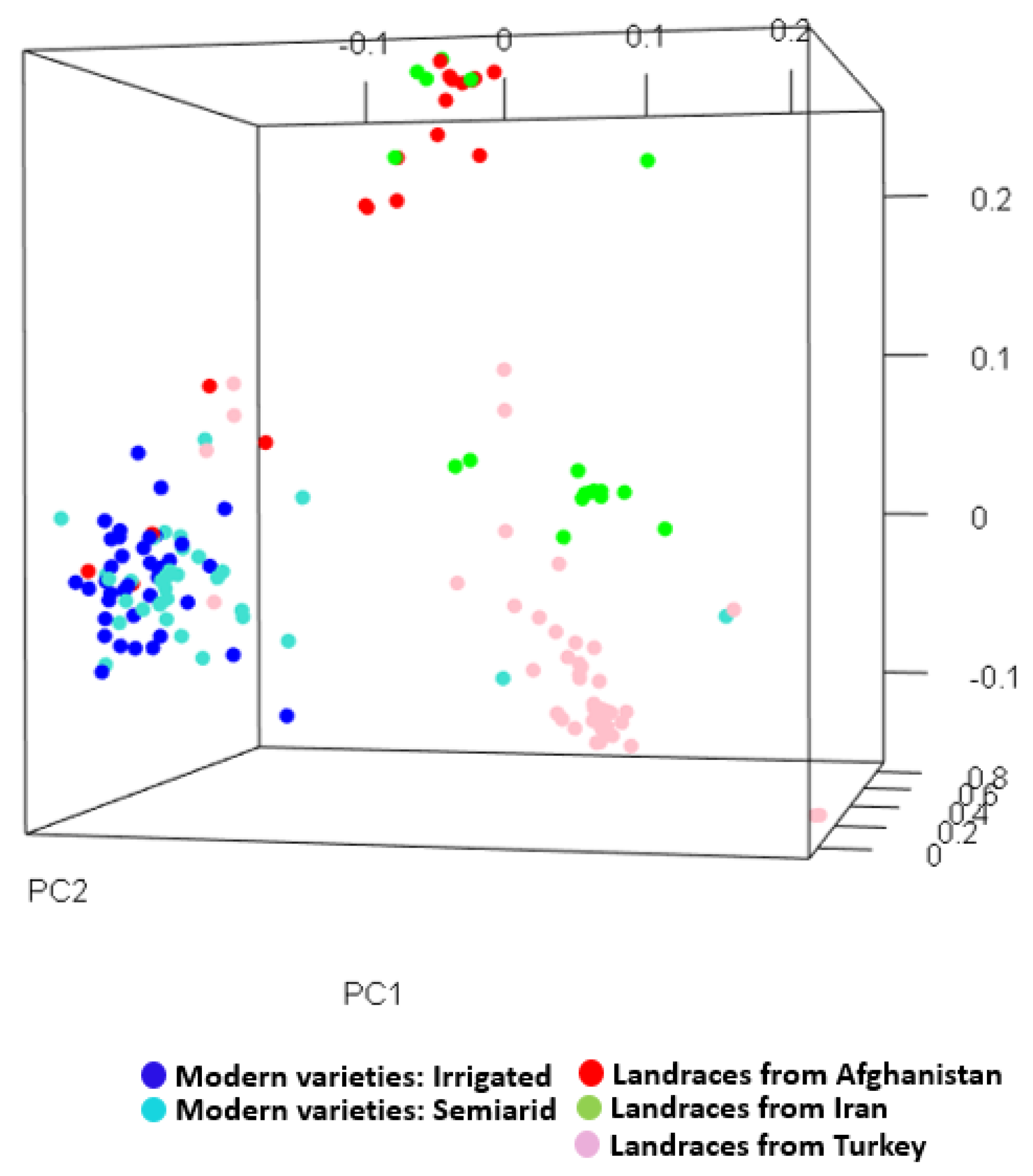
3.1. Morphological and Genetic Diversity
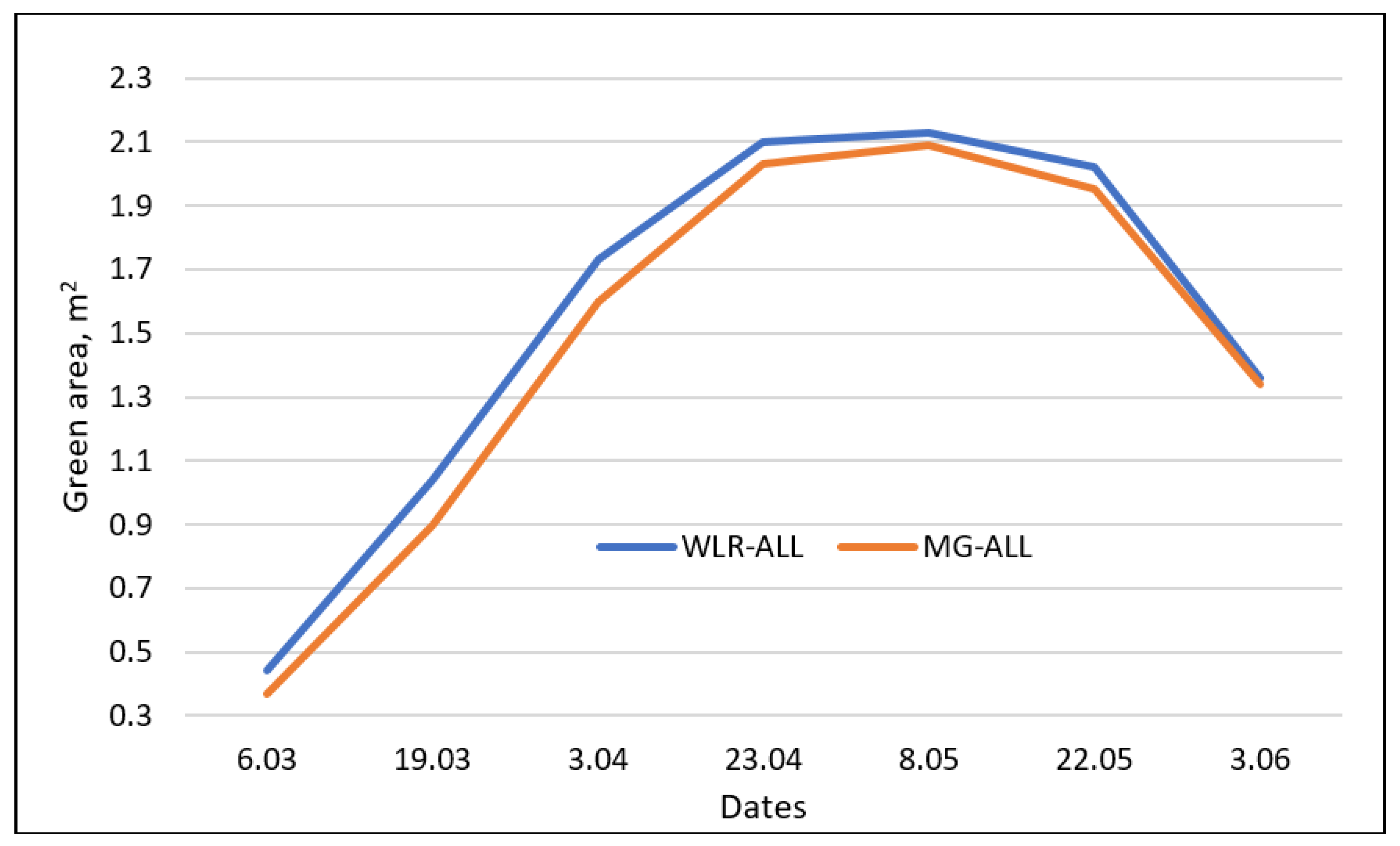
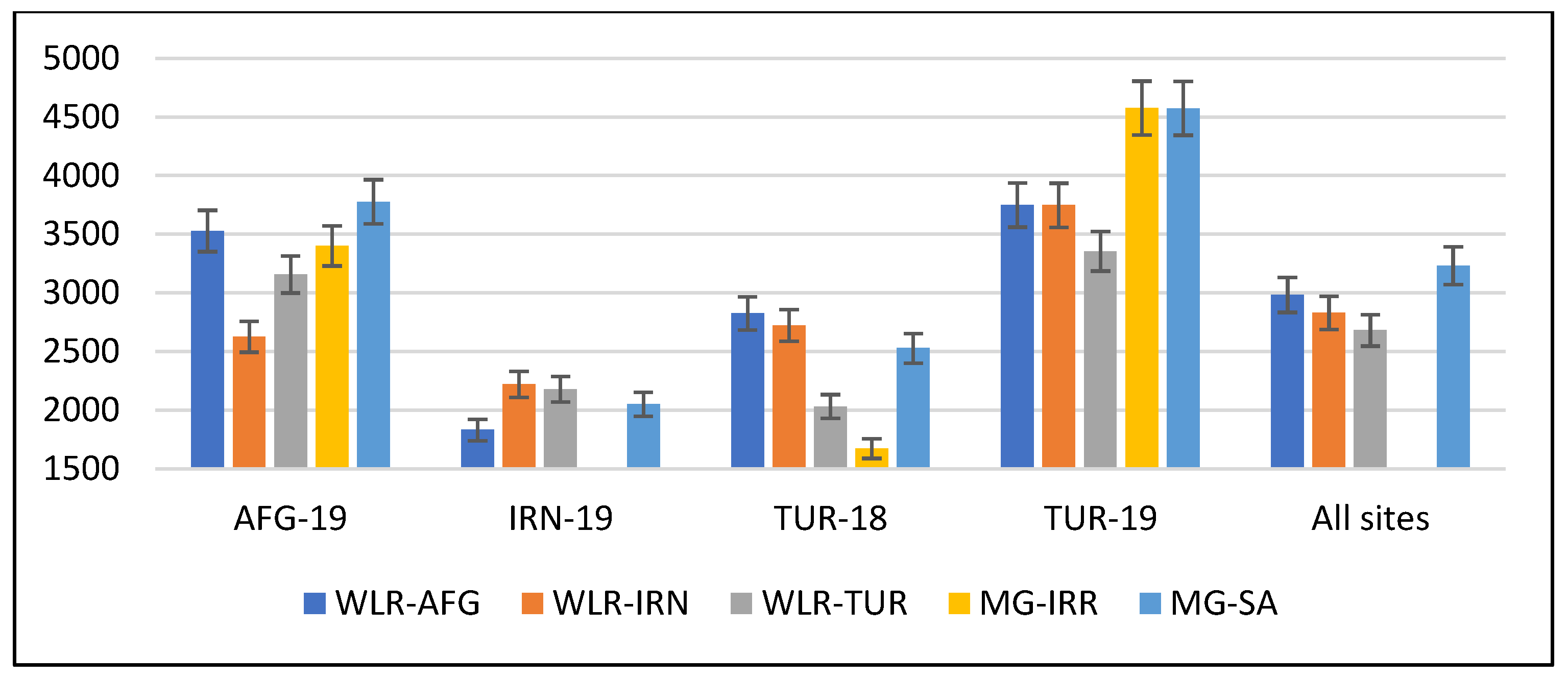
3.2. Adaptation Traits
3.3. Grain Yield and Its Components
3.4. Relationship between Grain Yield and Agronomic Traits
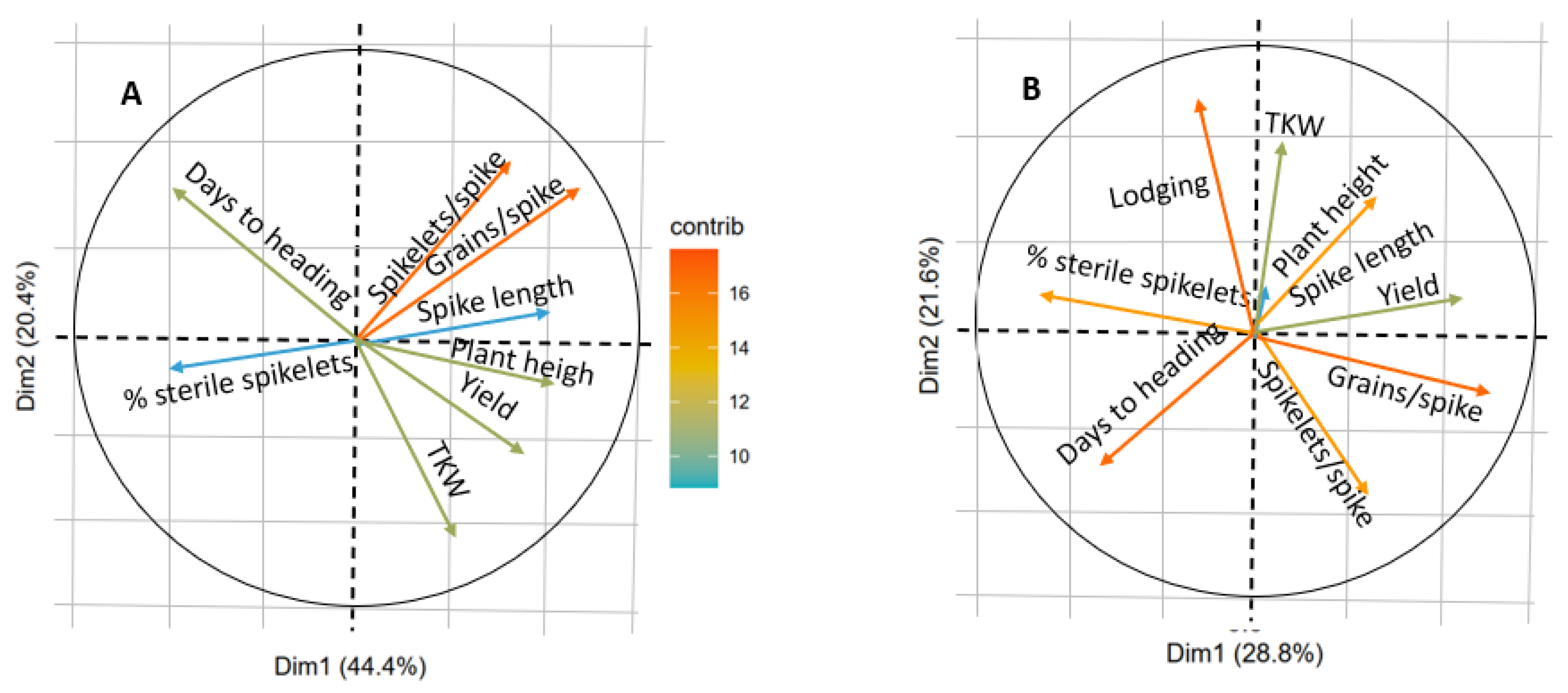
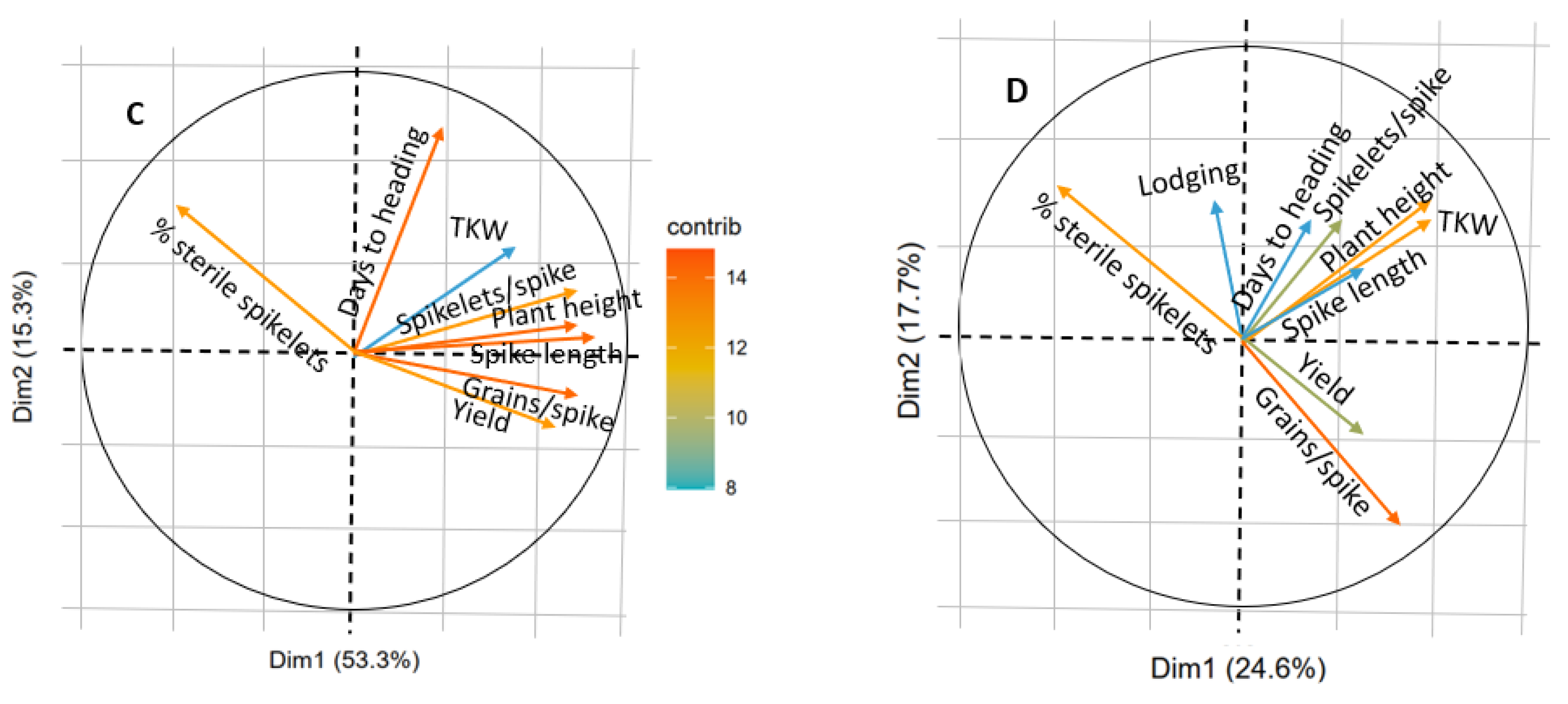
3.5. Molecular Markers Frequencies and Traits Associations
3.6. Agronomic Performance of Superior Landraces
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morgounov, A.; Ozdemir, F.; Keser, M.; Akin, B.; Payne, T.; Braun, H.-J. International Winter Wheat Improvement Program: History, activities, impact and future. Front. Agric. Sci. Eng. 2019, 6, 240–250. [Google Scholar] [CrossRef] [Green Version]
- Keser, M.; Gummadov, N.; Akin, B.; Belen, S.; Mert, Z.; Taner, S.; Topal, A.; Yazar, S.; Morgounov, A.; Sharma, R.C.; et al. Genetic gains in wheat in Turkey: Winter wheat for dryland conditions. Crop J. 2017, 5, 533–540. [Google Scholar] [CrossRef]
- Nehe, A.; Akin, B.; Sanal, T.; Kaplan, A.; Ünsal, R.; Dinçer, N.; Demir, L.; Geren, H.; Sevim, S.; Orhan, Ş.; et al. Genotype x environment interaction and genetic gain for grain yield and grain quality traits in Turkish spring wheat released between 1964 and 2010. PLoS ONE 2019, 14, e0219432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buerkert, A.; Oryakhail, M.; Filatenko, A.; Hammer, K. Cultivation and taxonomic classification of wheat landraces in the upper Panjsher valley of Afghanistan after 23 years of war. Gen. Res. Crop Evol. 2006, 53, 91–97. [Google Scholar] [CrossRef]
- Abbasabad, Z.; Mohammadi, E.; Moghaddam, A.; Kamali, M. Analysis of genetic diversity, population structure and linkage disequilibrium in Iranian wheat landraces using SSR markers. Plant Gen. Res. Charact. Util. 2017, 15, 327–334. [Google Scholar] [CrossRef]
- Husenov, B.; Muminjanov, H.; Dreisigacker, S.; Otambekova, M.; Akin, B.; Subasi, K.; Rasheed, A.; Shepelev, S.; Morgounov, A. Genetic diversity and agronomic performance of wheat landraces currently grown in Tajikistan. Crop. Sci. 2021. [Google Scholar] [CrossRef]
- Morgounov, A.; Keser, M.; Kan, M.; Küçükçongar, M.; Özdemir, F.; Gummadov, N.; Muminjanov, H.; Zuev, E.; Qualset, C.O. Wheat landraces currently grown in Turkey: Distribution, diversity, and use. Crop. Sci. 2016, 56, 3112–3124. [Google Scholar] [CrossRef] [Green Version]
- Baboev, S.; Muminjanov, H.; Turakulov, K.; Buronov, A.; Mamatkulov, I.; Koc, E.; Ozturk, I.; Dreisigacker, S.; Shepelev, S.; Morgounov, A. Diversity and sustainability of wheat landraces grown in Uzbekistan. Agron. Sustain. Dev. 2021, 41, 1–13. [Google Scholar] [CrossRef]
- Shewry, P.R. Do ancient types of wheat have health benefits compared with modern bread wheat? J. Cereal Sci. 2018, 79, 469–476. [Google Scholar] [CrossRef]
- Dwivedi, S.; Goldman, I.; Ortiz, R. Pursuing the potential of heirloom cultivars to improve adaptation, nutritional, and culinary features of food crops. Agronomy 2019, 9, 441. [Google Scholar] [CrossRef] [Green Version]
- Zuev, E.; Amri, A.; Brykova, A.; Pyukkenen, V.; Mitrofanova, O. Atlas of Bread Wheat (Triticum aestivum L.) Genetic Diversity Based on Spike and Kernel Characters; N.I. Vavilov Research Institute of Plant Industry (VIR): St. Petersburg, Russia, 2013; pp. 3–78. [Google Scholar]
- Pask, A.; Pietragalla, J.; Mullan, D.; Reynolds, M. Physiological Breeding II: A Field Guide to Wheat Phenotyping; CIMMYT: Mexico City, Mexico, 2012; pp. 1–132. [Google Scholar]
- Casadesus, J.; Villegas, D. Conventional digital cameras as a tool for assessing leaf area index and biomass for cereal breeding. J. Integr. Plant Biol. 2014, 56, 7–14. [Google Scholar] [CrossRef]
- Botstein, D.; White, R.; Skolnick, M.; Davis, R. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar] [PubMed]
- Dreisigacker, S.; Sehgal, D.; Reyes Jaimez, A.E.; Luna Garrido, B.; Muñoz Zavala, S.; Núñez Ríos, C.; Mollins, J.; Mall, S. CIMMYT Wheat Molecular Genetics: Laboratory Protocols and Applications to Wheat Breeding; CIMMYT: Mexico City, Mexico, 2016; pp. 1–154. [Google Scholar]
- Khalid, M.; Afzal, F.; Gul, A.; Amir, R.; Subhani, A.; Ahmed, Z.; Mahmood, Z.; Xia, X.; Rasheed, A.; He, Z. Molecular characterization of 87 functional genes in wheat diversity panel and their association with phenotypes under well-watered and water-limited conditions. Front. Plant Sci. 2019, 10, 717. [Google Scholar] [CrossRef]
- Rufo, R.; Alvaro, F.; Royo, C.; Soriano, J.M. From landraces to improved cultivars: Assessment of genetic diversity and population structure of Mediterranean wheat using SNP markers. PLoS ONE 2019, 14, 0219867. [Google Scholar] [CrossRef] [Green Version]
- Cavanagh, C.R.; Chao, S.; Wang, S.; Huang, B.E.; Stephen, S.; Kiani, S.; Forrest, K.; Saintenac, C.; Brown-Guedira, G.L.; Akhunova, A.; et al. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 8057–8062. [Google Scholar] [CrossRef] [Green Version]
- Sehgal, D.; Dreisigacker, S.; Belen, S.; Küçüközdemir, Ü.; Mert, Z.; Özer, E.; Morgounov, A. Mining centuries old in situ conserved Turkish wheat landraces for grain yield and stripe rust resistance genes. Front. Gen. 2016, 7, 201. [Google Scholar] [CrossRef] [Green Version]
- Rahimi, Y.; Bihamta, M.R.; Taleei, A.; Alipour, H.; Ingvarsson, P.K. Genome-wide association study of agronomic traits in bread wheat reveals novel putative alleles for future breeding programs. BMC Plant Biol. 2019, 19, 541. [Google Scholar] [CrossRef] [Green Version]
- Manickavelu, A.; Joukhadar, R.; Jighly, A.; Lan, C.; Huerta-Espino, J.; Stanikzai, A.S.; Kilian, A.; Singh, R.P.; Ban, T. Genome wide association mapping of stripe rust resistance in Afghan wheat landraces. Plant Sci. 2016, 252, 222–229. [Google Scholar] [CrossRef] [Green Version]
- Shokat, S.; Sehgal, D.; Liu, F.; Singh, S. GWAS analysis of wheat pre-breeding germplasm for terminal drought stress using next generation sequencing technology. Preprints 2020, 2020020272. [Google Scholar] [CrossRef] [Green Version]
- Buerstmayr, M.; Steiner, B.; Buerstmayr, H. Breeding for Fusarium head blight resistance in wheat—Progress and challenges. Plant Breed. 2020, 139, 429–454. [Google Scholar] [CrossRef]
- Velu, G.; Singh, R.; Crespo-Herrera, L.; Juliana, P.; Dreisigacker, S.; Valluru, R.; Stangoulis, J.; Sohu, V.; Mavi, G.S.; Mishra, V.K.; et al. Genetic dissection of grain zinc concentration in spring wheat for mainstreaming biofortification in CIMMYT wheat breeding. Sci. Rep. 2018, 8, 13526. [Google Scholar] [CrossRef]
- Parker, T.; Palkovic, A.; Brummer, C.; Gepts, P. Registration of ‘UC Rio Zape’ heirloom-like dry bean. J. Plant Reg. 2021, 15, 37–42. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trait | Number of Sites × Years | Wheat Landraces | Modern Germplasm | All | ||||
|---|---|---|---|---|---|---|---|---|
| AFG | IRN | TUR | IRR | SA | WLR | MG | ||
| Number of genotypes | 20 | 19 | 45 | 39 | 34 | 84 | 73 | |
| % of genotypes with winter growth habit | 1 | 0 | 79 | 62 | 80 | 70 | 51 | 76 |
| Days to heading from Jan. 1 | 3 | 129 ± 0.6 | 129 ± 0.4 | 135 ± 0.5 | 130 ± 0.4 | 130 ± 0.4 | 132 ± 0.4 | 130 ± 0.3 |
| Plant height, cm | 3 | 98 ± 2.2 | 91 ± 1.3 | 90 ± 0.9 | 78 ± 1.3 | 86 ± 1.9 | 92 ± 0.9 | 82 ± 1.2 |
| Stripe rust, % | 2 | 33.2 | 36.6 | 30.1 | 10.2 | 8.3 | 28.7 | 10.1 |
| Leaf rust, % | 2 | 57.0 | 61.9 | 54.2 | 26.7 | 26.3 | 56.6 | 29.2 |
| Lodging, % | 1 | 64.3 | 94.2 | 74.0 | 1.5 | 1.3 | 76.2 | 1.4 |
| Spikes/0.25 m2 | 1 | 153 ± 6.8 | 205 ± 8.9 | 174 ± 5.5 | 146 ± 4.3 | 153 ± 4.3 | 176 ± 4.3 | 149 ± 3.6 |
| Spike length, cm | 2 | 8.6 ± 0.2 | 8.2 ± 0.1 | 6.8 ± 0.2 | 8.0 ± 0.1 | 8.4 ± 0.2 | 7.5 ± 0.2 | 8.1 ± 0.1 |
| Spikelets/spike | 2 | 17.1 ± 0.4 | 14.5 ± 0.3 | 14.9 ± 0.1 | 17.1 ± 0.2 | 18.0 ± 0.5 | 15.3 ± 0.2 | 17.5 ± 0.2 |
| Sterile spikelets, % | 2 | 14.3 | 23.4 | 24.0 | 19.4 | 15.7 | 21.7 | 17.3 |
| Grains/spike | 2 | 26.2 ± 1.4 | 15.7 ± 0.6 | 15.0 ± 0.4 | 24.1 ± 0.9 | 28.6 ± 0.8 | 17.8 ± 0.7 | 26.2 ± 0.6 |
| 1000 kernel weight, g | 2 | 38.7 ± 1.0 | 42.8 ± 0.7 | 36.8 ± 0.5 | 32.8 ± 0.5 | 32.7 ± 0.7 | 38.6 ± 0.5 | 32.7 ± 0.4 |
| Grain yield, kg/ha | 3 | 3368 ± 90 | 3033 ± 65 | 2849 ± 57 | 3218 ± 68 | 3626 ± 69 | 3014 ± 66 | 3408 ± 53 |
| Entry | Local Name | Growth Habit | Days to Heading | Plant Height, cm | Stripe Rust, % | Leaf Rust, % | Grains/Spike | TKW, g | Yield, kg/ha |
|---|---|---|---|---|---|---|---|---|---|
| TUR 19 | TUR 18–19 | TUR 18–19 | TUR 18 | TUR 19 | TUR 18–19 | TUR 18–19 | AFG19 TUR 18–19 | ||
| Local Checks | |||||||||
| 85 | Bezostaya (long term IRR LC) | W | 131 | 83 | 40 | 60 | 26.1 | 35.1 | 3438 |
| 124 | Gerek (long term SA LC) | W | 131 | 82 | 40 | 40 | 19.5 | 31.7 | 3897 |
| 88 | Nacibey (IRR LC) | W | 131 | 82 | 0 | 40 | 33.4 | 35.3 | 3203 |
| 125 | Karahan (SA LC) | F | 131 | 84 | 0 | 40 | 22.8 | 31.9 | 4020 |
| Afghanistan WLR | |||||||||
| 9 | Roshan safed khosha | S | 133 | 91 | 5 | 50 | 29.0 | 34.6 | 4183 |
| 5 | Shanaze | S | 127 | 98 | 40 | 60 | 21.4 | 38.0 | 4100 |
| 7 | Safedak kalak bedon e dasa | S | 129 | 98 | 70 | 40 | 28.2 | 41.5 | 3803 |
| 1 | Kalak robat sangi | S | 129 | 99 | 60 | 60 | 26.1 | 43.2 | 3694 |
| 6 | Nesh shotor | F | 132 | 101 | 5 | 20 | 35.3 | 50.3 | 3656 |
| Iran WLR | |||||||||
| 24 | Khosheh ablaq | W | 129 | 88 | 40 | 50 | 21.1 | 41.4 | 3675 |
| 37 | Qzil khosheh | F | 132 | 90 | 30 | 40 | 22.5 | 36.7 | 3454 |
| 23 | Sardari biotype | W | 130 | 83 | 0 | 60 | 12.7 | 42.3 | 3359 |
| 30 | Sardari biotype | W | 127 | 81 | 10 | 50 | 15.5 | 43.5 | 3124 |
| 21 | Sardari biotype | W | 128 | 80 | 50 | 70 | 15.3 | 47.9 | 3067 |
| Turkey WLR | |||||||||
| 60 | Kirmizi buğday | W | 130 | 81 | 0 | 50 | 14.8 | 38.1 | 3908 |
| 50 | Şergun | W | 131 | 85 | 0 | 40 | 17.3 | 42.4 | 3890 |
| 54 | Hinta | F | 130 | 81 | 0 | 40 | 16.2 | 38.3 | 3514 |
| 62 | Akbugday | F | 135 | 79 | 80 | 40 | 17.6 | 37.1 | 3355 |
| 45 | Akbugday | F | 132 | 79 | 0 | 50 | 15.6 | 38.9 | 3196 |
| MG-IRR | |||||||||
| 112 | Grk79//Inqalab 91*2/Tukuru | S | 129 | 72 | 0 | 30 | 30.5 | 36.2 | 4171 |
| 107 | Tam200*2/Mo88//Kamb1*2/ Kukuna/3/Sw89-3218/Vorona | W | 131 | 63 | 40 | 60 | 22.4 | 32.3 | 3757 |
| 93 | Agri/Nac//Kauz/3/1d13.1/Mlt/4/ Atay/Galvez87//Shark-1 | W | 129 | 59 | 10 | 50 | 24.1 | 29.7 | 3636 |
| 114 | Mv Sed | W | 127 | 61 | 5 | 50 | 23.2 | 31.8 | 3606 |
| 95 | Mt0419/Destin//Bonito-36 | W | 131 | 70 | 0 | 40 | 26.1 | 31.5 | 3584 |
| MG-SA | |||||||||
| 128 | Nd643/2*Waxwing/4/Tam200/ Kauz/3/Agri/Bjy//Vee | F | 131 | 79 | 0 | 0 | 28.2 | 33.2 | 4335 |
| 142 | Spartanka//Pbw343*2/Kukuna | W | 133 | 78 | 0 | 20 | 31.5 | 36.6 | 4138 |
| 139 | Sultan95/Atilla//Zargana-6 | W | 136 | 98 | 0 | 70 | 29.8 | 37.2 | 4023 |
| 138 | Ks00f5-14-7/Eureka//Zargana-4 | F | 135 | 95 | 0 | 60 | 29.1 | 39.6 | 4007 |
| 133 | Vorona//Milan/Sha7/3/Mv17/4/ Atay/Galvez87//Shark-1 | W | 131 | 74 | 0 | 40 | 30.0 | 34.7 | 3919 |
| LSD 0.05 | - | 6 | 5.8 | - | - | 5.2 | 6.3 | 514 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morgounov, A.; Özdemir, F.; Keser, M.; Akin, B.; Dababat, A.A.; Dreisigacker, S.; Golkari, S.; Koc, E.; Küçükçongar, M.; Muminjanov, H.; et al. Diversity and Adaptation of Currently Grown Wheat Landraces and Modern Germplasm in Afghanistan, Iran, and Turkey. Crops 2021, 1, 54-67. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1020007
Morgounov A, Özdemir F, Keser M, Akin B, Dababat AA, Dreisigacker S, Golkari S, Koc E, Küçükçongar M, Muminjanov H, et al. Diversity and Adaptation of Currently Grown Wheat Landraces and Modern Germplasm in Afghanistan, Iran, and Turkey. Crops. 2021; 1(2):54-67. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1020007
Chicago/Turabian StyleMorgounov, Alexey, Fatih Özdemir, Mesut Keser, Beyhan Akin, Abdelfattah A. Dababat, Susanne Dreisigacker, Saber Golkari, Emrah Koc, Murat Küçükçongar, Hafiz Muminjanov, and et al. 2021. "Diversity and Adaptation of Currently Grown Wheat Landraces and Modern Germplasm in Afghanistan, Iran, and Turkey" Crops 1, no. 2: 54-67. https://0-doi-org.brum.beds.ac.uk/10.3390/crops1020007