
Marine Fungi: Opportunities and Challenges


CESAM, Department of Biology, University of Aveiro, 3810-193 Aveiro, Portugal
*
Author to whom correspondence should be addressed.
Encyclopedia 2022, 2(1), 559-577; https://0-doi-org.brum.beds.ac.uk/10.3390/encyclopedia2010037
Submission received: 10 January 2022
/
Revised: 10 February 2022
/
Accepted: 24 February 2022
/
Published: 28 February 2022
(This article belongs to the Collection Encyclopedia of Fungi)
Definition
:Marine fungi play a crucial role in energy flow and nutrient recycling, mediating the cycling of dissolved organic matter in marine environments. However, despite being a prolific group of organisms, marine fungi have been largely neglected for a long time. Besides their importance in the marine food web, marine fungi represent an active source of natural products. Over the last years, researchers have focused on studying marine organisms to discover new metabolites with antibacterial, antiviral, and anticancer activities. Moreover, with the advances in high-throughput sequencing technologies and mass spectrometry techniques, genomic and metabolomic approaches have revealed to be of paramount importance in natural products discovery. The marine mycobiome includes many organisms still to be identified, and the ones already known are still underutilized in biotechnological applications. For this reason, it is undeniable that exploring the marine mycobiome including new habitats and substrates, even those of remote access, is fundamental for describing the true magnitude of the Earth’s mycobiome.
1. Introduction
Fungi are ubiquitous organisms widely distributed in ecosystems, including marine environments [1,2,3]. Marine environments represent the last frontier of biodiversity. Even though marine habitats account for 70% of the surface of the planet, they still remain biologically unexplored. Fungi are key players in terrestrial and marine environments representing a substantial proportion of the microbial diversity on Earth [1].
Over time, different definitions of marine fungi have been used. Marine fungi were originally defined based on their physiological characteristics, such as the requirement of more than 30% salinity to grow [4]. The first attempt to define ‘marine fungi’ and still the most quoted definition was given by Kohlmeyer and Kohlmeyer [5]. These authors restricted ‘marine fungi’ to two ecological groups: obligate and facultative marine fungi. Obligate marine fungi are those that grow and sporulate exclusively in a marine or estuarine habitat; facultative marine fungi are those from freshwater or terrestrial milieus, able to grow and sporulate in marine environments. However, this distinction is not always easy nor clear to establish, and it is somehow controversial. Later, Jones et al. [6] labelled ‘obligate’ marine fungi as those isolated from submerged substrates or sediments from marine environments. The term “marine-derived fungi” has been extensively used in the field of natural product chemistry [7]. Recently, Pang et al. [8] reviewed the use of the terms “marine fungi” and “marine-derived fungi” and proposed a wide-ranging definition. These authors replaced Kohlmeyers’ definition by a broader concept. From this point forward, marine fungi are defined as any fungus able to: (i) grow and/or sporulate (on substrata) in marine environments; (ii) form symbiotic relationships with other marine organisms; or (iii) adapt and evolve or be metabolically active in marine environments.
In the past, marine fungi were viewed as an exotic group, relatively species-poor, with low abundance. Over the last two decades the use of culture-independent approaches and, more recently, of next-generation sequencing methods have started to disclose a considerable hidden fungal diversity from a wide range of environments [9]. In this respect, only recently, marine fungi have attracted attention and are yet to find a prominent place in biotechnology [3].
Marine fungi are a biochemically diverse group of organisms representing a promising source of novel bioactive natural compounds. The secondary metabolites produced by marine fungi include terpenes, steroids, polyketides, peptides, alkaloids, and polysaccharides. These metabolites are mainly associated with antimicrobial, anticancer, antiviral, antioxidant, and anti-inflammatory activities [3,10]. Considering this wide range of activities, these metabolites hold great promise since they can be used for drug discovery and medical, pharmaceutical, agriculture and cosmetic applications [3]. Besides being a source of bioactive compounds, marine fungi are also recognized for their metabolic capacities for bioremediation. This capacity aids, for instance, in the degradation of recalcitrant cell wall compounds (marine fungi express catalases, laccases, peroxidases) and environmental contaminants (e.g., toxic pollutants and microplastics) [11].
2. Origin and Diversity of Marine Fungi
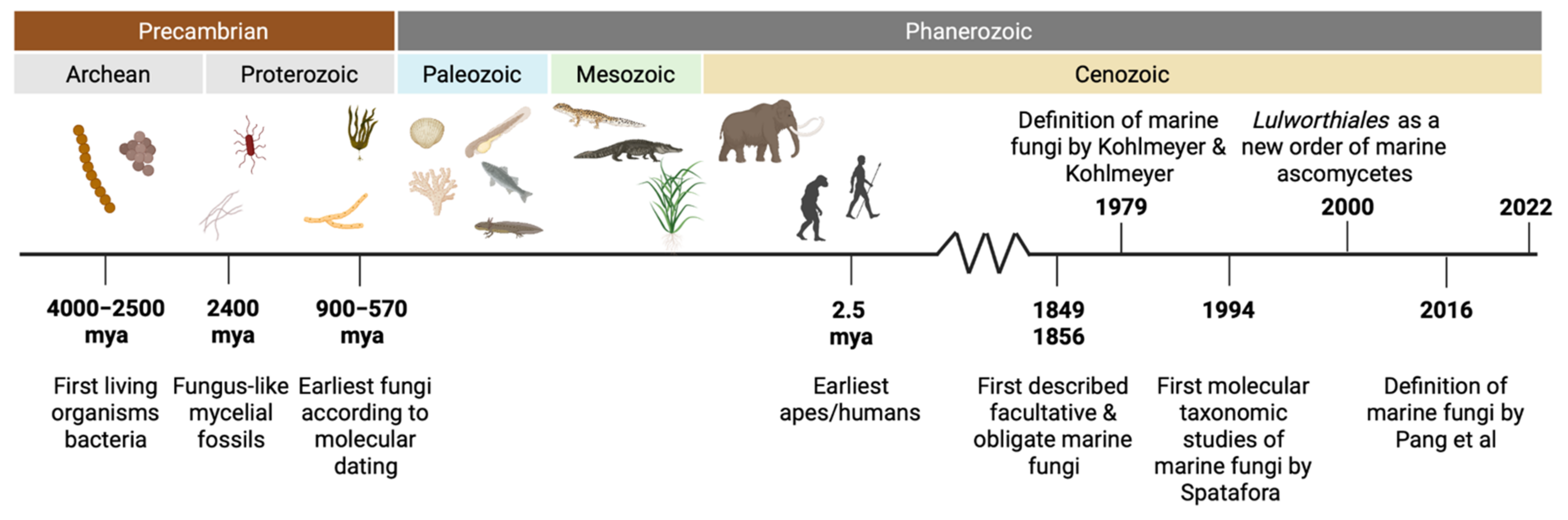
The origin of fungi remains unclear. Fungi are presumed to have evolved in the late Proterozoic, about 900–570 million years ago (mya) (Figure 1) [12,13,14,15]. However, according to the Ongeluk fossils, the fungal clade is considerably two to three times older than current age estimates [16]. The basalt from the Palaeoproterozoic Ongeluk Formation in South Africa suggests that fungi have inhabited submarine volcanoes approximately 2400 mya. Most of the species described in marine ecosystems have relations to terrestrial species. In fact, fungi transitioned several times from marine to terrestrial environments, and vice versa [17]. During evolution in the Precambrian (from the formation of Earth about 4600 mya to the beginning of the Cambrian Period, about 541 mya), cooling events and marine salinity decrease, resulted in the increase of dissolved oxygen, allowing the transition of non-marine early fungi to marine niches [18]. The existence of a terrestrial runoff or different migration trajectories from terrestrial to marine environment [19] was accompanied by several adaptations in response to these environmental changes [20]. However, the origin of marine fungi is still discussed.
2.1. Challenges and Adaptions
Fungi in the sea face substantial challenges compared to the terrestrial compartment. Figure 2 shows some of these challenges, and the molecular and morphological adaptations of fungi to thrive in the marine environment. High levels of salinity lead to osmotic and ionic stress. However, fungi can control their turgor through a conserved pathway known as the high-osmolarity-glycerol (HOG) signaling pathway. This pathway is responsible for the regulation of the salt efflux pumps and the production of osmolytes compatible with cellular functions [21]. Another evident molecular adaptation is the production of gels and slimes by lignicolous marine fungi (colonizers of driftwood or submerged wood) which may retain secreted enzymes at the substrate [21]. Additionally, a range of morphological adaptations in fungal structures and in the spore-dispersion strategies contributes for the success of marine fungi (Figure 2). One example of morphological-adapted fungi are chytrids (Chytridiomycota), also known as zoosporic true fungi, that depend on their zoospores for dispersal in water [22]. Although chytrids are one of the early diverging fungal lineages [22], and are the most common fungal group in marine habitats [23,24,25,26], there is no sufficient evidence to show that the ancestral chytrids were marine [27]. This gap is largely due to the lack of molecular data in Chytridiomycota [22].
2.2. Diversity
Recent advances in high throughput sequencing technologies stimulated a new wave of research in marine mycology, revealing an increase of fungal diversity in the marine realm. However, using culture dependent methods, the most common taxa which have been recovered are predominantly allied to the phyla Ascomycota and Basidiomycota, to which more than 90% of the total described marine species belong [22]. In marine environments, the phylum Ascomycota is represented by the subphyla Pezizomycotina, Saccharomycotina and Taphrinomycotina, while Basidiomycota is represented by Agaricomycotina, Pucciniomycotina and Ustilaginomycotina [1,28].
The subphylum Pezizomycotina, includes the classes Dothideomycetes, Eurotiomycetes and Sordariomycetes, which comprise the highest number of species described [7]. In this context, some of these classes include many marine hyphomycetes, a diverse ecological group that contains species able to alternate their life cycle between aquatic and terrestrial habitats. This ecological group is often associated with leaf litter, driftwood, water, algae, marine animals, and halophytes. Marine hyphomycetes have probably also evolved from terrestrial fungi with some morphological adaptations to colonize marine habitats. Yet, attempts to taxonomically resolve several taxa incertae sedis in these subphyla are currently ongoing. Additionally, yeast-like fungi within the phyla Ascomycota and Basidiomycota have been found in marine environments [1].
The total fungal diversity has been the subject of much discussion. It is estimated that the number of fungal species ranges from 1.5 to 5.1 million [29,30]. Marine fungi represent less than 1% of the known species [10,29,30], thus remaining a poorly characterized group. It has been estimated that the number of marine fungi range from 10,000 to 12,500 species [29], although more recently Garzoli et al. [31] suggested that up to 1 million species are yet to be discovered. The most up-to-date number of described marine fungal species until February 2021 is only 1901, distributed in 769 genera, 226 families, 88 orders, 22 classes and seven phyla into Aphelidiomycota, Ascomycota, Basidiomycota, Blastocladiomycota, Chytridiomycota, Mortierellomycota and Mucoromycota (Figure 3, www.marinefungi.org (accessed on 9 January 2022) [32]). This estimate includes yeasts and filamentous fungi and other basal fungal lineages.
Over the last two decades, attempts to understand the diversity and function of fungi in marine habitats has also been made through culture-independent approaches [23,26,33]. More recently, next-generation sequencing methods have been used to disclose a considerable number of fungal species from a wide range of marine environments [23,34]. However, from the known marine fungal species, some of them have a limited number of ITS (internal transcribed spacer region) sequences and are mostly identified based on the conserved 28S or 18S rRNA genes (large and small subunits of ribosomal RNA, respectively). The analysis of these conserved regions is not always reliable to delimitate fungal species. Therefore, multi-gene phylogenetic analyses allied with morphological characteristics are crucial for a correct species identification. Furthermore, genome sequencing approaches are critical to understand the evolution of fungi particularly those associated with early diverging lineages [22].
3. Diversity of Marine Fungi Habitats and Their Ecological Roles
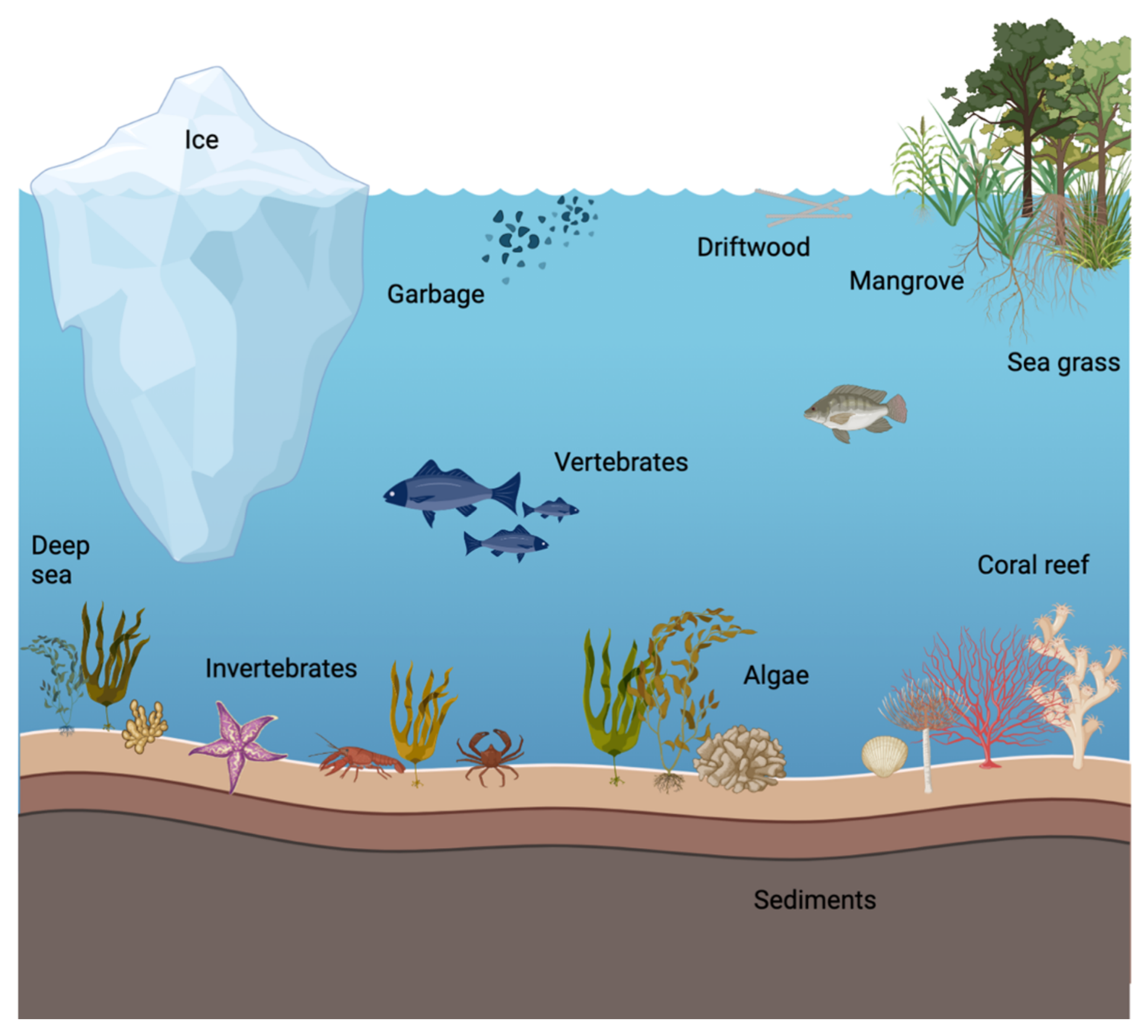
Fungi are key players in terrestrial and marine environments representing a substantial proportion of the microbial diversity on Earth [22]. However, even though marine habitats account for 70% of the surface of the planet, they remain biologically unexplored. This is especially evident when considering microbial life in these environments, most specifically its fungal component. Despite many difficulties to isolate fungal species from marine environments, researchers continue to make progresses towards uncovering marine fungal species by exploring new habitats. Marine fungi have been recovered from organic and inorganic substrata (Figure 4): mangrove plants, macroalgae, seagrass, coral reef, drift- and submerged wood, sponges, sea ice, sea garbage, and coastal and open-ocean water columns [35,36,37]. As mentioned, using high-throughput sequencing technologies, researchers have also found marine fungal species on deep-sea and benthic sediments, hydrothermal vents, oxygen-deficient environments and in global surface waters [23,26,33]. Table 1 summarizes studies (from 2015) of marine fungi diversity covering different substrata.
Although marine fungi are ubiquitous, some fungal species are found only in certain geographic regions, such as the tropics, subtropics, temperate or polar waters [149], or in specific substrates (Table 1).
To sustain the biodiversity and marine ecosystem functioning, the energy fixed by primary photosynthetic producers in the ocean is controlled to various trophic levels [22]. Microorganisms play a key role in nutrient recycling and in the regulation of energy flow in marine ecosystems [22]. From these microorganisms, marine fungi are one of the major components in marine food webs and occur as saprobes, mutualists, endobionts and parasites [22]. As saprobes, fungi can colonize and transform the organic matter originated from plants, algae, and animals into valuable nutrients. As mutualists, fungi can be associated to organisms such as sponges or protozoans, where both individuals benefit. As endobionts, fungi can provide energy to the host or even be commensals benefiting from host protection. As parasites of phytoplankton, macrophytes, algae, and animals’ marine fungi can cause leaching of dissolved organic matter and destruction of their host populations [22]. Cryptomycota and Chytridiomycota are considered as early diverging fungal lineages, displaying physiological adaptations for inhabiting aquatic ecosystems, and contributing to marine food webs [150]. For example, Senga et al. [151] showed that parasitic fungi, such as chytrids (Chytridiomycota), utilize living diatoms efficiently. Additionally, chytrids can change the quality and quantity of algal dissolved organic matter, thus influencing higher trophic levels. Therefore, by virtue of chytrids’ interactions with diverse living or dead organisms, marine fungi can transform and incorporate organic matter into the food web regulating the energy flow in marine ecosystems [1]. Similarly, members of Cryptomycota are parasitic zoosporic fungi exhibiting a similar life cycle to chytrids (e.g., free-swimming stage), and are typically found in freshwater, soil and in some marine habitats. In addition to chytrids, Cryptomycota fungi are dynamic players on microbial food webs and can parasitize many phytoplankton species such as diatoms, green algae, and dinoflagellates [152]. Despite the potential role of Chytridiomycota and Cryptomycota as mycopesticide formulations, such is still unexplored, mainly due to cultivation difficulties.
4. A Source of Potential Bioactive Compounds
The identification of novel natural products appears to be a robust approach to the approval of new drugs and therapies [34]. Therefore, there is the need to create new, safer, eco-friendly, and sustainable substances as alternatives to chemically synthetized products. Natural products are pharmacologically or biologically active chemical compounds or molecules (often below 1000 Da), found in nature and produced by a biological source [34]. Additionally, high molecular weight compounds such as proteins and polysaccharides also may show therapeutics potential. Referred to as secondary metabolites, these compounds are not directly involved in any primary function related to growth, development, and reproduction of an organism [153]. However, they are more commonly associated with organism’s survival prospects, such as, defense, competition, and communication [154].
Penicillin, the first natural product antibiotic, produced by the fungus Penicillium notatum (syn. P. rubens and previously known as P. chrysogenum) was discovered by Sir Alexander Fleming in 1928 [155]. This event fueled the exploration of microorganisms as sources for antibiotics, initiating the golden era for the discovery of other natural products from fungi. Some studies were focused on an intensive exploration of new natural products with biologically active properties, from marine sources. Initially, the study of marine natural products was limited to organisms easy to collect such as corals and sponges. For example, the toxin tedanolide was isolated from the sponge Tedania ignis, while prostaglandins were isolated from the Caribbean Sea whip Plexaura homomalla [156]. Moreover, many natural products may also be synthesized by marine microorganisms [157]. In this respect and considering that only about 5% of the world’s fungal taxa have been described, marine fungi have recently attracted attention for being a plentiful source for new chemical entities. However, marine fungi still represent an underexplored group, and researchers are yet to discover their full biotechnological potential [158,159,160,161].
In 1945, the first group of natural products from marine fungi was described as cephalosporins, a class of β-lactam antibiotics isolated from the Acremonium chrysogenum (previously known as Cephalosporium chrysogenum) [162]. Later, in 1977, Okutani identified gliotoxin as a new type of the antibiotic diketopiperazine isolated from a marine fungus Aspergillus sp. obtained from deep-sea sediments [163]. Thenceforward, the number of antibiotic compounds isolated from marine fungi was rapidly increasing [162]. More than 3500 marine fungal secondary metabolites have already been reported [164], but a high number surely remains untapped. Such may be related to the unexploited marine ecological niches and their almost ubiquitous distribution, as well as to the high number of silent biosynthetic clusters in the genomes [34,159]. Figure 5 shows some classes of metabolites produced by marine fungi and their associated biological activities.
Most of the research on secondary metabolites of marine fungi have been focused on a few genera: Penicillium, Aspergillus, Fusarium, and Cladosporium [60,159]. However, studies of natural products from marine fungi continue to increase and have been expanded to other genera [159]. These studies have demonstrated that marine fungi represent a tremendous source of secondary metabolites for drug discovery [157]. The number of studies published increases every year, with most of them coming from species associated with different substrata and new habitats. Most of these metabolites are derivatives of those found on terrestrial fungi but hold significant differences in their biological activities (sometimes more effective and more specific).
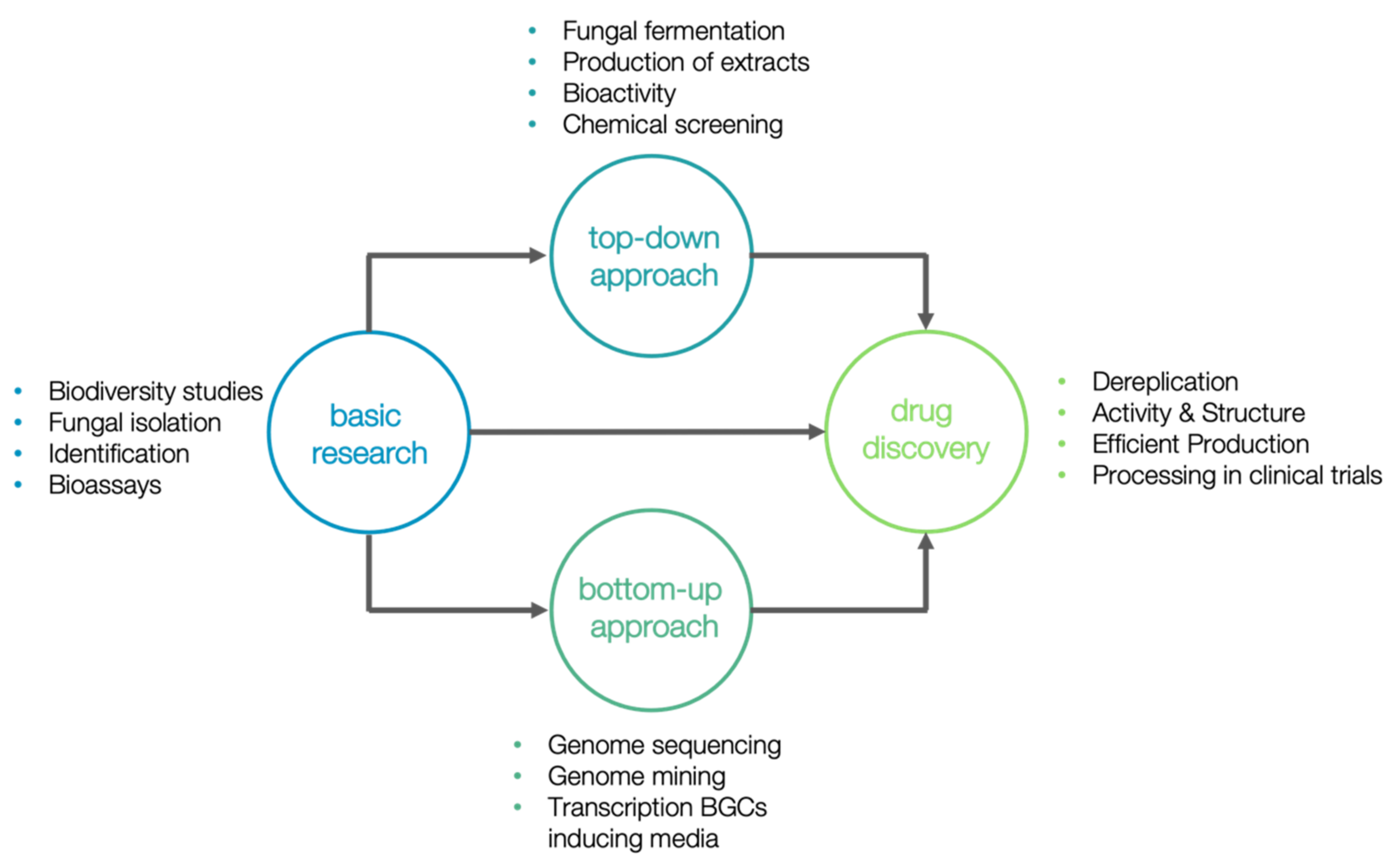
Currently, 14 marine natural products are approved as drugs and more than 30 natural products are in different phases of clinical trials [165]. Despite the large number of new natural products from marine fungi, Plinabulin (diketopiperazine class) is the only one isolated from a marine fungus (Aspergillus sp.) that is currently under clinical trials by BeyondSpring Pharmaceuticals [165]. This compound is currently in the late-stage of a world-wide phase III clinical trial for anticancer therapy of non-small cell lung cancer and for the reduction of chemotherapy-induced neutropenia [166]. Despite the research efforts towards marine fungi, most studies are focused only on the description of new natural compounds with in vitro biological activities. A simplified representation of the fungal marine natural product drug discovery pipeline is shown in Figure 6. Therefore, after the drug discovery process, efficient approaches to produce enough quantities of the interest compound for clinical trials need to be implemented. Silber et al. [162] already listed different biotechnological approaches in all stages of the discovery-to-development of antibiotics from marine fungi.
4.1. Metabolomics Approaches
Omics (genomic, transcriptomic, proteomic, and metabolomic approaches) have become popular due to the advances in high-throughput sequencing technologies, bioinformatic tools and mass spectrometry techniques. Metabolomic approaches have embraced the discovery of natural products [34]. Mass spectrometry (MS) is a highly sensitive analytical tool used for the identification and quantification of metabolites [167]. Analytical techniques (e.g., nuclear magnetic resonance (NMR)) and several types of mass analyzers are used for metabolomic studies, such as capillary electrophoresis mass spectrometry (CE-MS), gas chromatography mass spectrometry (GC-MS), liquid chromatography mass spectrometry (LC-MS), and matrix-assisted laser desorption/ionization mass spectrometry (MALDI-MS).
Mass spectrometry has a significantly higher sensitivity and resolution than NMR [168], that requires a relatively simple sample preparation step and generates large amounts of data that remain challenging to handle. Despite MS-based approaches being able to quantify the metabolic profile in different sample matrices, the metabolites’ capacity to ionize in a particular analyzer deeply influences data. MS-based approaches have become popular both in targeted and untargeted studies, due to their potential to identify an array of diverse metabolites by an organism [169,170]. On the other hand, NMR-based approaches have advantages compared to MS. NMR is quantitative, highly reproducible, and has the possibility of not using standards to elucidate the compounds [168]. NMR is also a non-destructive technique, which allows MS-based approaches to be used as complementary. The use of these two approaches is information-rich, becoming more popular for the discovery of new metabolites. Table 2 sums up the recent studies of marine fungi’s metabolome using different techniques. The most used techniques are GC-MS and LC-MS [171]. GC-MS separates a mixture of compounds according to their compound’s volatility/molecular weight, while LC-MS separates according to compound polarity or hydrophobicity for example. However, LC-MS represents the most versatile approach for profiling extracts [172].
Metabolomics has become an essential analytical tool for the drug discovery process. Nonetheless, as metabolomics of marine fungi is still in its infancy, the identification of marine natural products, remains challenging [160]. Untargeted metabolomics generate massive data sets. However, analysis is time consuming, due to the lack of user-friendly bioinformatic annotation pipeline tools and open access data repositories. Furthermore, the prediction of biological activity of the secondary metabolites identified is a challenging task—if not impossible—because small structural differences in the molecules may lead to different bioactivities [34].
Recently, Rotter et al. [173] listed an overview of databases useful for the marine biotechnology pipelines. Briefly, the major MS/MS data organization tool and open access is the Global Natural Products Social (GNPS) Molecular Networking. However, most of the specific databases that already contain some marine natural products from fungi, such as Marine Lit and Dictionary of Natural Products, are not open access. Recently, Van Santen et al. [174] developed the Natural Products Atlas, an open access database for microbial natural products discovery. In any case, at our knowledge, there is no marine fungi-specific database combining mass spectra data.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 2.
Different MS techniques used on marine fungi metabolomics.
| Technique | Species | Host/Habitat | Studies |
|---|---|---|---|
| GC-MS | Grammothele fuligo, Rigidoporus vinctus, Cystobasidium minutum, Candida railenensis, Pichia kudriavzevii, Aspergillus sp., Penicillium chrysogenum | Algae | [175,176,177] |
| GC-MS | Emericella sp., Aspergillus unguis | Sponge | [178,179] |
| GC-MS | Aspergillus versicolor | Sediments | [180] |
| GC-MS | Aspergillus flavus, Penicillium echinulatum, Microascus croci, Penicillium purpurogenum | Seaweed | [181] |
| GC-MS/LC-MS | Penicillium brevicompactum | Anemone | [182] |
| GC-MS/LC-MS | Botryosphaeria sp., Piriformospora indica | Algae | [183,184] |
| GC-MS/NMR | Aspergillus sydowii | Sponge | [185] |
| LC-MS | Trichoderma reesi, Trichoderma atroviride, Aspergillus welwitschiae | Sponge | [186,187] |
| LC-MS | Aspergillus allahabadii, Aspergillus ochraceopetaliformis, Aspergillus sp., Penicillium sp. | Sediments | [188,189,190] |
| LC-MS | A. sydowii, P. chrysogenum | Corals | [191] |
| LC-MS | Aspergillus affinis, A. sydowii | Sea water | [169,192] |
| LC-MS | Emericellopsis cladophorae | Algae | [170] |
| LC-MS | Cladosporium cladosporioides | Seaweed | [193] |
| LC-MS/NMR | Oidiodendron griseum, Aspergillus flavipes | Sediments | [194,195] |
4.2. Genomic Approaches
Marine fungal genomes’ sequencing adds more in-depth information to search for natural products from these microorganisms. The increasing number of fungal genomes sequenced has revealed that fungi contain an enormous potential for secondary metabolite production. Biosynthetic gene clusters (BGCs) can be involved in different biosynthetic pathways, resulting in a variety of possible final products [117]. However, most of these BGCs have not been linked to their corresponding natural products [34]. So far, only approximately 4000 fungal species have completed or ongoing genome projects [196], with more than 8000 genomes available. The shortage of marine fungal genomes in public databases, such as Genome–NCBI (National Center for Biotechnology Information) (https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/ (accessed on 9 January 2022) [197]) and JGI Genome Portal (Joint Genome Institute) (https://genome.jgi.doe.gov/portal/ (accessed on 9 January 2022) [198]), impairs the fast identification of BGCs, thus hampering the discovery of natural products from these microorganisms. Table 3 shows an overview of the marine fungal genomes sequenced that are deposited at Genome–NCBI and at JGI databases. Ascomycota is the phylum that contains more sequenced marine fungal genomes (21 isolates of 12 different families).
The ‘unculturable’ fungal species represent one of the major problems and challenges for marine mycology. Isolation into pure culture remains the gold standard for the identification, description, and genome sequencing of new marine fungal species. Another challenge for marine natural products identification is, as mentioned before, that BGCs may remain silent under laboratory culture conditions. To overcome this, different methods have been developed to ‘awaken’ or stimulate the expression of silent genes. Among these methods to induce the production of BGC coded compounds, we can highlight: (1) the use of epigenetic modifiers of natural or chemical elicitors (microbial cells components, microbial lysates, or non-biological origin compounds) and (2) the co-cultivation with other species [199]. These strategies are known as OSMAC (One Strain Many Compounds) approach [200]. OSMAC makes use of altering cultivation parameters or adding elicitors to activate silent and poorly expressed BGCs in microorganisms to induce the expression of more natural products [201].
5. Conclusions and Prospects
Marine fungi have been studied since the first record of a marine species. However, despite their widespread distribution and abundance, marine fungi have been largely neglected. Since isolation into pure culture remains the gold standard for discovery and description of novel taxa, the use of different methods for isolation, culture media, incubation temperatures, and damp/moist chamber incubation, may contribute for the increase of fungal diversity. Environmental alterations, such as the seasonality could also yield different diversity. However, climate changes can also have a major impact on the food web dynamics in the marine fungal communities as well as alter their structure causing unpredictable consequences. It is widely accepted that the climate is changing at a global level, and it is also essential to understand these impacts on marine fungi dynamics.
It is recognized that marine fungi represent a potential source of secondary metabolites for drug discovery. Although genome-wide sequencing allows the detection of BGCs involved in different biosynthetic pathways in marine fungi, there is still limited of genomes available. Therefore, it is crucial to broaden this field of investigation into underexplored marine taxa, to infer about the lifestyles, ecological roles, evolutionary history, and regulatory mechanisms.
Despite the research motivation, a holistic and integrated approach is necessary to ensure the success of any conservation management strategy or economic use of marine fungi. Therefore, all fungal species recorded in each study should be isolated and preserved in reference culture collections to provide the authentic biological material. This material will not only increase the accuracy of the taxonomic assignments but will also be useful for further biotechnological purposes.
Author Contributions
Conceptualization: M.F.M.G., A.C.E. and A.A.; Data curation: M.F.M.G.; Funding acquisition: A.A.; Methodology: M.F.M.G., A.C.E. and A.A.; Supervision: A.A. and A.C.E. Writing—original draft: M.F.M.G. Writing—review and editing: M.F.M.G., A.C.E. and A.A. All authors have read and agreed to the published version of the manuscript.
Funding
We acknowledge financial support of Portuguese Foundation for Science and Technology (FCT) through national funds to CESAM (UIDP/50017/2020+UIDB/50017/2020+LA/P/0094/2020) and to M. Gonçalves (SFRH/BD/129020/2017).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
Entry Link on the Encyclopedia Platform
References
- Grossart, H.P.; Van den Wyngaert, S.; Kagami, M.; Wurzbacher, C.; Cunliffe, M.; Rojas-Jimenez, K. Fungi in aquatic ecosystems. Nat. Rev. Microbiol. 2019, 17, 339–354. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jeewon, R.; Chen, Y.J.; Bhunjun, C.S.; Calabon, M.S.; Jiang, H.B.; Lin, C.G.; Norphanphoun, C.; Sysouphanthong, P.; Pem, D.; et al. The numbers of fungi: Is the descriptive curve flattening? Fungal Divers. 2020, 103, 219–271. [Google Scholar] [CrossRef]
- Tarman, K. Marine fungi as a source of natural products. Encycl. Mar. Biotechnol. 2020, 4, 2147–2160. [Google Scholar]
- Johnson, T.W.; Sparrow, F.K. Fungi in Oceans and Estuaries; J. Cramer: Weinheim, Germany, 1961; p. 685. [Google Scholar]
- Kohlmeyer, J.; Kohlmeyer, E. Marine Mycology: The Higher Fungi; Academic Press: London, UK, 1979; p. 704. [Google Scholar]
- Jones, E.B.G.; Sakayaroj, J.; Suetrong, S.; Somrithipol, S.; Pang, K.L. Classification of marine Ascomycota, anamorphic taxa and Basidiomycota. Fungal Divers. 2009, 35, 187. [Google Scholar]
- Jensen, P.R.; Fenical, W. Secondary metabolites from marine fungi. In Fungi in Marine Environments; Hyde, K.D., Ed.; Fungal Diversity Research Series; Fungal Diversity Press: Hong Kong, China, 2002; Volume 7, pp. 293–315. [Google Scholar]
- Pang, K.-L.; Overy, D.P.; Jones, E.B.G.; Calado, M.; Burgaud, G.; Walker, A.K.; Johnson, J.A.; Kerr, R.G.; Cha, H.-J.; Bills, G.F. ‘Marine fungi’ and ‘marine-derived fungi’ in natural product chemistry research: Toward a new consensual definition. Fungal Biol. Rev. 2016, 30, 163–175. [Google Scholar] [CrossRef]
- Richards, T.A.; Jones, M.D.; Leonard, G.; Bass, D. Marine fungi: Their ecology and molecular diversity. Ann. Rev. Mar. Sci. 2012, 4, 495–522. [Google Scholar] [CrossRef] [PubMed]
- Queirós, B.; Barreira, J.C.; Sarmento, A.C.; Ferreira, I.C. In search of synergistic effects in antioxidant capacity of combined edible mushrooms. Int. J. Food Sci. Nutr. 2009, 60, 160–172. [Google Scholar] [CrossRef]
- Zeghal, E.; Vaksmaa, A.; Vielfaure, H.; Boekhout, T.; Niemann, H. The potential role of marine fungi in plastic degradation—A review. Front. Mar. Sci. 2021, 8, 738877. [Google Scholar] [CrossRef]
- Remy, W.; Taylor, T.N.; Hass, H. Early Devonian fungi: A blastocladalean fungus with sexual reproduction. Am. J. Bot. 1994, 81, 690–702. [Google Scholar] [CrossRef]
- Remy, W.; Hass, H.; Kerp, H. Fossil arbuscular mycosshiza from early Debonian. Myhcologia 1995, 87, 561–573. [Google Scholar]
- Taylor, T.N.; Remy, W.; Hass, H. Fungi from the Lower Devonian Rhynie chert: Chytridiomycetes. Am. J. Bot. 1992, 79, 1233–1241. [Google Scholar] [CrossRef]
- Taylor, T.N.; Galtier, J.; Axsmith, B.J. Fungi from the Lower Carboniferous of central France. Rev. Palaeobot. Palynol. 1994, 83, 253–260. [Google Scholar] [CrossRef]
- Bengtson, S.; Rasmussen, B.; Ivarsson, M.; Muhling, J.; Broman, C.; Marone, F.; Stampanoni, M.; Bekker, A. Fungus-like mycelial fossils in 2.4-billion-year-old vesicular basalt. Nat. Ecol. Evol. 2017, 1, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Amend, A.; Burgaud, G.; Cunliffe, M.; Edgcomb, V.P.; Ettinger, C.L.; Gutierrez, M.H.; Heitman, J.; Hom, E.F.Y.; Ianiri, G.; Jones, A.C.; et al. Fungi in the marine environment: Open questions and unsolved problems. Mbio 2019, 10, e01189-18. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sarma, V.V.; Thambugala, K.M.; Huang, J.J.; Li, X.Y.; Hao, G.F. Ecology and evolution of marine fungi with their adaptation to climate change. Front. Microbiol. 2021, 12, 2527. [Google Scholar] [CrossRef]
- Raghukumar, C.; Damare, S.; Singh, P. A review on deep-sea fungi: Occurrence, diversity and adaptions. Bot. Mar. 2010, 53, 479–492. [Google Scholar] [CrossRef]
- Sakayaroj, J.; Pang, K.L.; Jones, E.B.G. Multi-gene phylogeny of the Halosphaeriaceae: Its ordinal status, relationships between genera and morphological character evolution. Fungal Divers. 2011, 46, 87–109. [Google Scholar] [CrossRef]
- Gladfelter, A.S.; James, T.Y.; Amend, A.S. Marine fungi. Curr. Biol. 2019, 29, R191–R195. [Google Scholar] [CrossRef]
- Jones, E.G.; Pang, K.-L.; Abdel-Wahab, M.A.; Scholz, B.; Hyde, K.D.; Boekhout, T.; Ebel, R.; Rateb, M.E.; Henderson, L.; Sakayaroj, J. An online resource for marine fungi. Fungal Divers. 2019, 96, 347–433. [Google Scholar] [CrossRef]
- Richards, T.A.; Leonard, G.; Mahé, F.; del Campo, J.; Romac, S.; Jones, M.D.M.; Maguire, F.; Dunthorn, M.; De Vargas, C.; Massana, R.; et al. Molecular diversity and distribution of marine fungi across 130 European environmental samples. Proc. R. Soc. B Biol. Sci. 2015, 282, 20152243. [Google Scholar] [CrossRef]
- Comeau, A.M.; Vincent, W.F.; Bernier, L.; Lovejoy, C. Novel chytrid lineages dominate fungal sequences in diverse marine and freshwater habitats. Sci. Rep. 2016, 6, 30120. [Google Scholar] [CrossRef] [PubMed]
- Hassett, B.T.; Ducluzeau, A.L.L.; Collins, R.E.; Gradinger, R. Spatial distribution of aquatic marine fungi across the western Arctic and sub-arctic. Environ. Microbiol. 2017, 19, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Picard, K.T. Coastal marine habitats harbor novel early-diverging fungal diversity. Fungal Ecol. 2017, 25, 1–13. [Google Scholar] [CrossRef]
- James, T.Y.; Kauff, F.; Schoch, C.L.; Matheny, P.B.; Hofstetter, V.; Cox, C.J.; Celio, G.; Gueidan, C.; Fraker, E.; Miadlikowska, J.; et al. Reconstructing the early evolution of Fungi using a six-gene phylogeny. Nature 2006, 443, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Sánchez-Ramírez, S.; Kõljalg, U.; Bahram, M.; Döring, M.; Schigel, D.; May, T.; Ryberg, M.; Abarenkov, K. High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 2018, 90, 135–159. [Google Scholar] [CrossRef]
- Jones, E.B.G. Are there more marine fungi to be described? Bot. Mar. 2011, 54, 343–354. [Google Scholar] [CrossRef]
- Sarma, V.V. Marine fungal diversity: Present status and future perspectives. In Microbial Diversity in Ecosystem Sustainability and Biotechnological Applications; Springer: Singapore, 2019; pp. 267–291. [Google Scholar]
- Garzoli, L.; Poli, A.; Prigione, V.; Gnavi, G.; Varese, G.C. Peacock’s tail with a fungal cocktail: First assessment of the mycobiota associated with the brown alga Padina pavonica. Fungal Ecol. 2018, 35, 87–97. [Google Scholar] [CrossRef]
- An Online Resource for Marine Fungi. Available online: https://www.marinefungi.org (accessed on 5 January 2022).
- Xu, W.; Guo, S.; Pang, K.L.; Luo, Z.H. Fungi associated with chimney and sulfide samples from a South Mid-Atlantic Ridge hydrothermal site: Distribution, diversity and abundance. Deep-Sea Res. I Oceanogr. Res. Pap. 2017, 123, 48–55. [Google Scholar] [CrossRef]
- Reich, M.; Labes, A. How to boost marine fungal research: A first step towards a multidisciplinary approach by combining molecular fungal ecology and natural products chemistry. Mar. Genom. 2017, 36, 57–75. [Google Scholar] [CrossRef]
- Gnavi, G.; Garzoli, L.; Poli, A.; Prigione, V.; Burgaud, G.; Varese, G.C. The culturable mycobiota of Flabellia petiolate: First survey of marine fungi associated to a Mediterranean green alga. PLoS ONE 2017, 12, e0175941. [Google Scholar] [CrossRef]
- Raghukumar, S. Fungi in Coastal and Oceanic Marine Ecosystems; Springer: New York, NY, USA, 2017; p. 364. [Google Scholar]
- Marchese, P.; Garzoli, L.; Young, R.; Allcock, L.; Barry, F.; Tuohy, M.; Murphy, M. Fungi populate deep-sea coral gardens as well as marine sediments in the Irish Atlantic Ocean. Environ. Microbiol. 2021, 23, 4168–4184. [Google Scholar] [CrossRef] [PubMed]
- BioRender. Available online: https://biorender.com (accessed on 25 September 2021).
- Calado, M.D.L.; Silva, J.; Alves, C.; Susano, P.; Santos, D.; Alves, J.; Martins, A.; Gaspar, H.; Pedrosa, R.; Campos, M.J. Marine endophytic fungi associated with Halopteris scoparia (Linnaeus) Sauvageau as producers of bioactive secondary metabolites with potential dermocosmetic application. PLoS ONE 2021, 16, e0250954. [Google Scholar] [CrossRef] [PubMed]
- Chin, J.M.; Puchooa, D.; Bahorun, T.; Jeewon, R. Antimicrobial properties of marine fungi from sponges and brown algae of Mauritius. Mycology 2021, 12, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Garzoli, L.; Gnavi, G.; Varese, G.C.; Picco, A.M. Mycobiota associated with the rhodophyte alien species Asparagopsis taxiformis (Delile) Trevisan de Saint-Léon in the Mediterranean Sea. Mar. Ecol. 2015, 36, 959–968. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Santos, L.; Silva, B.M.V.; Abreu, A.C.; Vicente, T.F.L.; Esteves, A.C.; Alves, A. Biodiversity of Penicillium species from marine environments in Portugal and description of Penicillium lusitanum sp. nov., a novel species isolated from sea water. Int. J. Syst. Evol. Microbiol. 2019, 69, 3014–3021. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Vicente, T.F.L.; Esteves, A.C.; Alves, A. Neptunomyces aureus gen. et sp. nov. (Didymosphaeriaceae, Pleosporales) isolated from algae in Ria de Aveiro, Portugal. MycoKeys 2019, 60, 31–44. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Vicente, T.F.L.; Esteves, A.C.; Alves, A. Novel halotolerant species of Emericellopsis and Parasarocladium associated with macroalgae in an estuarine environment. Mycologia 2020, 112, 154–171. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Esteves, A.C.; Alves, A. Revealing the hidden diversity of marine fungi in Portugal with the description of two novel species, Neoascochyta fuci sp. nov. and Paraconiothyrium salinum sp. nov. Int. J. Syst. Evol. Microbiol. 2020, 70, 5337–5354. [Google Scholar] [CrossRef]
- Hagestad, O.C.; Andersen, J.H.; Altermark, B.; Hansen, E.; Rämä, T. Cultivable marine fungi from the Arctic Archipelago of Svalbard and their antibacterial activity. Mycology 2020, 11, 230–242. [Google Scholar] [CrossRef]
- Hong, J.-H.; Jang, S.; Heo, Y.M.; Min, M.; Lee, H.; Lee, Y.M.; Lee, H.; Kim, J.-J. Investigation of marine-derived fungal diversity and their exploitable biological activities. Mar. Drugs 2015, 13, 4137–4155. [Google Scholar] [CrossRef]
- Kamat, S.; Kumari, M.; Taritla, S.; Jayabaskaran, C. Endophytic fungi of marine alga from Konkan Coast, India—A rich source of bioactive material. Front. Mar. Sci. 2020, 7, 31. [Google Scholar] [CrossRef]
- Lee, S.; Park, M.S.; Lee, H.; Kim, J.J.; Eimes, J.A.; Lim, Y.W. Fungal diversity and enzyme activity associated with the macroalgae, Agarum clathratum. Mycobiology 2019, 47, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Martorell, M.M.; Lannert, M.; Matula, C.V.; Quartino, M.L.; de Figueroa, L.I.C.; Mac Cormack, W.P.; Ruberto, L.A.M. Studies toward the comprehension of fungal-macroalgae interaction in cold marine regions from a biotechnological perspective. Fungal Biol. 2020, 125, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Pasqualetti, M.; Giovannini, V.; Barghini, P.; Gorrasi, S.; Fenice, M. Diversity and ecology of culturable marine fungi associated with Posidonia oceanica leaves and their epiphytic algae Dictyota dichotoma and Sphaerococcus coronopifolius. Fungal Ecol. 2020, 44, 100906. [Google Scholar] [CrossRef]
- Poli, A.; Bovio, E.; Ranieri, L.; Varese, G.C.; Prigione, V. News from the Sea: A new genus and seven new species in the Pleosporalean families Roussoellaceae and Thyridariaceae. Diversity 2020, 12, 144. [Google Scholar] [CrossRef]
- Poli, A.; Bovio, E.; Ranieri, L.; Varese, G.C.; Prigione, V. Fungal diversity in the Neptune forest: Comparison of the Mycobiota of Posidonia oceanica, Flabellia omycote, and Padina pavonica. Front. Microbiol. 2020, 11, 933. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, S.; Subban, K.; Chelliah, J. Diversity of marine macro-algicolous endophytic fungi and cytotoxic potential of Biscogniauxia petrensis metabolites against cancer cell lines. Front. Microbiol. 2021, 12, 1472. [Google Scholar] [CrossRef]
- Venkatachalam, A.; Govinda Rajulu, M.B.; Thirunavukkarasu, N.; Suryanarayanan, T.S. Endophytic fungi of marine algae and seagrasses: A novel source of chitin modifying enzymes. Mycosphere 2015, 6, 345–355. [Google Scholar] [CrossRef]
- Vicente, T.F.L.; Gonçalves, M.F.M.; Brandão, C.; Fidalgo, C.; Alves, A. Diversity of fungi associated with macroalgae from an estuarine environment and description of Cladosporium rubrum sp. nov. and Hypoxylon aveirense sp. nov. Int. J. Syst. Evol. Microbiol. 2021, 71, 4630. [Google Scholar] [CrossRef]
- Góes-Neto, A.; Marcelino, V.R.; Verbruggen, H.; da Silva, F.F.; Badotti, F. Biodiversity of endolithic fungi in coral skeletons and other reef substrates revealed with 18S rDNA metabarcoding. Coral Reefs 2020, 39, 229–238. [Google Scholar] [CrossRef]
- Paulino, G.V.; Félix, C.R.; Landell, M.F. Diversity of filamentous fungi associated with coral and sponges in coastal reefs of northeast Brazil. J. Basic Microbiol. 2020, 60, 103–111. [Google Scholar] [CrossRef]
- Godinho, V.M.; de Paula, M.T.R.; Silva, D.A.S.; Paresque, K.; Martins, A.P.; Colepicolo, P.; Rosa, C.A.; Rosa, L.H. Diversity and distribution of hidden cultivable fungi associated with marine animals of Antarctica. Fungal Biol. 2019, 123, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Marchese, P.; Garzoli, L.; Gnavi, G.; O’Connell, E.; Bouraoui, A.; Mehiri, M.; Murphy, J.M.; Varese, G.C. Diversity and bioactivity of fungi associated with the marine sea cucumber Holothuria poli: Disclosing the strains potential for biomedical applications. J. Appl. Microbiol. 2020, 129, 612–625. [Google Scholar] [CrossRef] [PubMed]
- Shaumi, A.; Cheang, U.C.; Yang, C.Y.; Chang, C.W.; Guo, S.Y.; Yang, C.H.; Chan, T.Y.; Pang, K.L. Culturable fungi associated with the marine shallow-water hydrothermal vent crab Xenograpsus testudinatus at Kueishan Island, Taiwan. Bot. Mar. 2021, 64, 289–300. [Google Scholar] [CrossRef]
- Abdel-Wahab, M.A.; Dayarathne, M.C.; Suetrong, S.; Guo, S.Y.; Alias, S.A.; Bahkali, A.H.; Nagahama, T.; Elgorban, A.M.; Abdel-Aziz, F.A.; Hodhod, M.S.; et al. New saprobic marine fungi and a new combination. Bot. Mar. 2017, 60, 469–488. [Google Scholar] [CrossRef]
- Abdel-Wahab, M.A.; Jones, E.B.G.; Bahkali, A.H.A.; Elgorban, A.M. Marine fungi from Red Sea mangroves in Saudi Arabia with Fulvocentrum rubrum sp. nov. (Torpedosporales, Ascomycota). Nova Hedwig. 2019, 108, 365–377. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Devadatha, B.; Sarma, V.V.; Khongphinitbunjong, K.; Chomnunti, P.; Hyde, K.D. Morpho–molecular characterization of micro fungi associated with marine based habitats. Mycosphere 2020, 11, 1–188. [Google Scholar] [CrossRef]
- Devadatha, B.; Sarma, V.V.; Ariyawansa, H.A.; Jones, E.B.G. Deniquelata vittalii sp. nov., a novel Indian saprobic marine fungus on Suaeda monoica and two new records of marine fungi from Muthupet mangroves, East coast of India. Mycosphere 2018, 9, 565–582. [Google Scholar] [CrossRef]
- Devadatha, B.; Sarma, V.V.; Jeewon, R.; Wanasinghe, D.N.; Hyde, K.D.; Jones, E.B.G. Thyridariella, a novel marine fungal genus from India: Morphological characterization and phylogeny inferred from multigene DNA sequence analyses. Mycol. Prog. 2018, 17, 791–804. [Google Scholar] [CrossRef]
- Devadatha, B.; Sarma, V.V. Pontoporeia mangrovei sp. nov, a new marine fungus from an Indian mangrove along with a new geographical and host record of Falciformispora lignatilis. Curr. Res. Environ. Appl. 2018, 8, 238–246. [Google Scholar]
- Hyde, K.D.; Tennakoon, D.S.; Jeewon, R.; Bhat, D.J.; Maharachchikumbura, S.S.N.; Rossi, W.; Leonardi, M.; Lee, H.M.; Mun, H.Y.; Houbraken, J.; et al. Fungal diversity notes 1036–1150: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers. 2019, 96, 1–242. [Google Scholar] [CrossRef]
- Manivel, G.; Raj, D.M.L.; Prathiviraj, R.; Senthilraja, P. Distribution of phylogenetic proximity upon species-rich marine Ascomycetes with reference to Pichavaram mangrove soil sediment of southern India. Gene Rep. 2020, 21, 100878. [Google Scholar] [CrossRef]
- Norphanphoun, C.; Raspé, O.; Jeewon, R.; Wen, T.C.; Hyde, K.D. Morphological and phylogenetic characterisation of novel Cytospora species associated with mangroves. MycoKeys 2018, 38, 93–120. [Google Scholar] [CrossRef] [PubMed]
- Phookamsak, R.; Hyde, K.D.; Jeewon, R.; Bhat, D.J.; Jones, E.B.G.; Maharachchikumbura, S.S.N.; Raspe’, O.; Karunarathna, S.K.; Wanasinghe, D.N.; Hongsanan, S.; et al. Fungal diversity notes 929–1035: Taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 2019, 95, 1–273. [Google Scholar] [CrossRef]
- Yuan, H.S.; Lu, X.; Dai, Y.C.; Hyde, K.D.; Kan, Y.H.; Kušan, Y.; He, S.H.; Liu, N.G.; Sarma, V.V.; Zhao, C.L.; et al. Fungal diversity notes 1277–1386: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2020, 104, 1–266. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, Z.; Ma, J.; Zhou, D.; Xu, J. Phylogenetic diversity, antimicrobial and antioxidant potential and identification of bioactive compounds from culturable endophytic fungi associated with mangrove Bruguiera sexangular (Lour.) Poir. Curr. Microbiol. 2021, 78, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Ivarsson, M.; Bengtson, S.; Skogby, H.; Lazor, P.; Broman, C.; Belivanova, V.; Marone, F. A fungal-prokaryotic consortium at the basalt-zeolite interface in subseafloor igneous crust. PLoS ONE 2015, 10, e0140106. [Google Scholar] [CrossRef]
- Ivarsson, M.; Peckmann, J.; Tehler, A.; Broman, C.; Bach, W.; Behrens, K.; Reitner, J.; Böttcher, M.E.; Norbäck-Ivarsson, L. Zygomycetes in vesicular basanites from Vesteris Seamount, Greenland Basin—A new type of cryptoendolithic fungi. PLoS ONE 2015, 10, e0133368. [Google Scholar] [CrossRef]
- Ivarsson, M.; Bengtson, S.; Neubeck, A. The igneous oceanic crust—Earth’s largest fungal habitat? Fungal Ecol. 2016, 20, 249–255. [Google Scholar] [CrossRef]
- Quemener, M.; Mara, P.; Schubotz, F.; Beaudoin, D.; Li, W.; Pachiadaki, M.; Sehein, T.R.; Sylvan, J.B.; Li, J.; Barbier, G.; et al. Meta-omics highlights the diversity, activity and adaptations of fungi in deep oceanic crust. Environ. Microbiol. 2020, 22, 3950–3967. [Google Scholar] [CrossRef]
- Kettner, M.T.; Oberbeckmann, S.; Labrenz, M.; Grossart, H. The eukaryotic life on microplastics in brackish ecosystems. Front. Mar. Sci. 2019, 10, 538. [Google Scholar] [CrossRef] [PubMed]
- Kirstein, I.V.; Wichels, A.; Krohne, G.; Gerdts, G. Mature biofilm communities on synthetic polymers in seawater—Specific or general? Mar. Environ. Res. 2018, 142, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Lacerda, A.L.D.F.; Proietti, M.C.; Secchi, E.R.; Taylor, J.D. Diverse groups of fungi are associated with plastics in the surface waters of the Western South Atlantic and the Antarctic Peninsula. Mol. Ecol. 2020, 29, 1903–1918. [Google Scholar] [CrossRef] [PubMed]
- Calado, M.D.L.; Carvalho, L.; Pang, K.L.; Barata, M. Diversity and ecological characterization of sporulating higher filamentous marine fungi associated with Spartina maritima (Curtis) Fernald in two Portuguese salt marshes. Microb. Ecol. 2015, 70, 612–633. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, M.F.M.; Aleixo, A.; Vicente, T.F.L.; Esteves, A.C.; Alves, A. Three new species of Neocamarosporium isolated from saline environments: N. aestuarinum sp. nov., N. endophyticum sp. nov. and N. halimiones sp. nov. Mycosphere 2019, 10, 608–621. [Google Scholar] [CrossRef]
- Park, J.M.; Hong, J.W.; You, Y.H.; Kim, J.G. Endophytic fungi of emersed halophytes in river deltas and tidal flats of the Korean Ramsar wetlands. J. Mar. Sci. Eng. 2021, 9, 430. [Google Scholar] [CrossRef]
- Velez, P.; Walker, A.K.; Gasca-Pineda, J.; Barrios, A.; Divanli, D.; González, M.C.; Nakagiri, A. Fine-scale temporal variation of intertidal marine fungal community structure: Insights from an impacted Baja California sandy beach in Mexico. Mar. Biodivers. 2021, 51, 6. [Google Scholar] [CrossRef]
- Walker, A.K.; Robicheau, B.M. Fungal diversity and community structure from coastal and barrier island beaches in the United States Gulf of Mexico. Sci. Rep. 2021, 11, 3889. [Google Scholar] [CrossRef]
- Abdel-Wahab, M.; Bahkali, A.H.; El-Gorban, A.M.; Jones, G. Metagenomics Study of Fungi and Fungi-like Organisms Associated with the Seagrass Halophila stipulacea (Forssk.) Asch. from Al-Leith Mangroves, Saudi Arabia; Research Square: Durham, NC, USA, 2021; pp. 1–19. [Google Scholar]
- Baral, H.O.; Rämä, T. Morphological update on Calycina marina (Pezizellaceae, Helotiales, Leotiomycetes), a new combination for Laetinaevia marina. Bot. Mar. 2015, 58, 523–534. [Google Scholar] [CrossRef]
- Ettinger, C.L.; Eisen, J.A. Characterization of the mycobiome of the seagrass, Zostera marina, reveals putative associations with marine chytrids. Front. Microbiol. 2019, 10, 2476. [Google Scholar] [CrossRef]
- Ettinger, C.L.; Eisen, J.A. Fungi, bacteria and omycote opportunistically isolated from the seagrass, Zostera marina. PLoS ONE 2020, 15, e0236135. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, C.L.; Vann, L.E.; Eisen, J.A. Global diversity and biogeography of the Zostera marina mycobiome. Appl. Environ. Microbiol. 2021, 87, e02795-20. [Google Scholar] [CrossRef]
- Kirichuk, N.N.; Pivkin, M.V. Filamentous fungi associated with the seagrass Zostera marina Linnaeus, 1753 of Rifovaya Bay (Peter the Great Bay, the Sea of Japan). Russ. J. Mar. Biol. 2015, 41, 351–355. [Google Scholar] [CrossRef]
- Kwon, S.L.; Park, M.S.; Jang, S.; Lee, Y.M.; Heo, Y.M.; Hong, J.H.; Lee, H.; Jang, Y.; Park, J.H.; Kim, C.; et al. The genus Arthrinium (Ascomycota, Sordariomycetes, Apiosporaceae) from marine habitats from Korea, with eight new species. IMA Fungus 2021, 12, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Subrmaniyan, R.; Ponnambalam, S.; Thirunavukarassu, T. Inter species variations in cultivable endophytic fungal diversity among the tropical seagrasses. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2016, 88, 849–857. [Google Scholar] [CrossRef]
- Supaphon, P.; Phongpaichit, S.; Sakayaroj, J.; Rukachaisirikul, V.; Kobmoo, N.; Spatafora, J.W. Phylogenetic community structure of fungal endobiotes in seagrass species. Bot. Mar. 2017, 60, 489–502. [Google Scholar] [CrossRef]
- Torta, L.; Piccolo, S.L.; Piazza, G.; Burruano, S.D.; Colombo, P. Lulwoana sp., a dark septate endophyte in roots of Posidonia oceanica (L.) Delile seagrass. Plant Biol. 2015, 17, 505–511. [Google Scholar] [CrossRef]
- Venkatachalam, A.; Thirunavukkarasu, N.; Suryanarayanan, T.S. Distribution and diversity of endobiontes in seagrasses. Fungal Ecol. 2015, 13, 60–65. [Google Scholar] [CrossRef]
- Vohník, M.; Borovec, O.; Kolařík, M. Communities of cultivable root mycobionts of the seagrass Posidonia oceanica in the northwest Mediterranean Sea are dominated by a hitherto undescribed pleosporalean dark septate endophyte. Microb. Ecol. 2016, 71, 442–451. [Google Scholar] [CrossRef]
- Vohník, M.; Borovec, O.; Kolaříková, Z.; Sudová, R.; Réblová, M. Extensive sampling and high-throughput sequencing reveal Posidoniomyces atricolor gen. et sp. nov. (Aigialaceae, Pleosporales) as the dominant root mycobiont of the dominant Mediterranean seagrass Posidonia oceanica. MycoKeys 2019, 55, 59. [Google Scholar] [CrossRef]
- Bovio, E.; Gnavi, G.; Prigione, V.; Spina, F.; Denaro, R.; Yakimov, M.; Calogero, R.; Crisafi, F.; Varese, G.C. The culturable mycobiota of a Mediterranean marine site after an oil spill: Isolation, identification and potential application in bioremediation. Sci. Total Environ. 2017, 576, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.; Wingfield, M.; Lombard, L.; Roets, F.; Swart, W.; Alvarado, P.; Carnegie, A.; Moreno, G.; Luangsaard, J.; Thangavel, R. Fungal Planet description sheets: 951–1041. Pers. Mol. Phylogeny Evol. Fungi 2019, 43, 223–425. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Chooi, Y.-H.; Gilchrist, C.L.M.; Lacey, E.; Pitt, J.I.; Roets, F.; Swart, W.J.; Cano-Lira, J.F.; Valenzuela-Lopez, N.; et al. Fungal Planet description sheets: 1042–1111. Persoonia 2020, 44, 301–459. [Google Scholar] [CrossRef] [PubMed]
- De Menezes, G.; Câmara, P.; Pinto, O.; Convey, P.; Carvalho-Silva, M.; Simões, J.; Rosa, C.; Rosa, L. Fungi in the Antarctic Cryosphere: Using DNA Metabarcoding to Reveal Fungal Diversity in Glacial Ice from the Antarctic Peninsula Region; Research Square: Durham, NC, USA, 2021; pp. 1–18. [Google Scholar]
- Fotedar, R.; Kolecka, A.; Boekhout, T.; Fell, J.W.; Al-Maliki, A.; Zeyara, A.; Al Marri, M. Fungal diversity of the hypersaline Inland Sea in Qatar. Bot. Mar. 2018, 61, 595–609. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Silva, B.M.V.; Esteves, A.C.; Alves, A. Verrucoconiothyrium ambiguum sp. nov., a novel species isolated from sea water, and affiliation of the genus Verrucoconiothyrium to the family Didymellaceae. Int. J. Syst. Evol. Microbiol. 2019, 69, 3769–3776. [Google Scholar] [CrossRef]
- Hassett, B.T.; Borrego, E.J.; Vonnahme, T.R.; Rama, T.; Kolomiets, M.V.; Gradinger, R. Arctic marine fungi: Biomass, functional genes, and putative ecological roles. ISME J. 2019, 13, 1484–1496. [Google Scholar] [CrossRef]
- Jeffries, T.C.; Curlevski, N.J.; Brown, M.V.; Harrison, D.P.; Doblin, M.A.; Petrou, K.; Ralph, P.J.; Seymour, J.R. Partitioning of fungal assemblages across different marine habitats. Environ. Microbiol. Rep. 2016, 8, 235–238. [Google Scholar] [CrossRef]
- Kilias, E.S.; Junges, L.; Šupraha, L.; Leonard, G.; Metfies, K.; Richards, T.A. Chytrid fungi distribution and co-occurrence with diatoms correlate with sea ice melt in the Arctic Ocean. Commun. Biol. 2020, 3, 1–13. [Google Scholar] [CrossRef]
- Li, W.; Wang, M.; Pan, H.; Burgaud, G.; Liang, S.; Guo, J.; Luo, T.; Li, Z.; Zhang, S.; Cai, L. Highlighting patterns of fungal diversity and composition shaped by ocean currents using the East China Sea as a model. Mol. Ecol. 2018, 27, 564–576. [Google Scholar] [CrossRef]
- Li, W.; Wang, M.; Burgaud, G.; Yu, H.; Cai, L. Fungal community composition and potential depth-related driving factors impacting distribution pattern and trophic modes from epi-to abyssopelagic zones of the Western Pacific Ocean. Microb. Ecol. 2019, 78, 820–831. [Google Scholar] [CrossRef]
- Wang, Y.; Sen, B.; He, Y.; Xie, N.; Wang, G. Spatiotemporal distribution and assemblages of planktonic fungi in the coastal waters of the Bohai Sea. Front. Microbiol. 2018, 9, 584. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sen, B.; He, Y.; Xie, N.; Wang, G. Impact of environmental gradients on the abundance and diversity of planktonic fungi across coastal habitats of contrasting trophic status. Sci. Total Environ. 2019, 683, 822–833. [Google Scholar] [CrossRef] [PubMed]
- Bubnova, E.N.; Konovalova, O.P. Fungi in bottom sediments of the Chukchi Sea. Russ. J. Mar. Biol. 2019, 45, 86–95. [Google Scholar] [CrossRef]
- Cecchi, G.; Cutroneo, L.; Di Piazza, S.; Capello, M.; Zotti, M. Culturable fungi from dredged and marine sediments from six ports studied in the framework of the SEDITERRA Project. J. Soils Sediments 2021, 21, 1563–1573. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.E.; Crane, C.; Barrett, S.; Cano-Lira, J.F.; Le Roux, J.J.; Thangavel, R.; Guarro, J.; et al. Fungal Planet description sheets: 469–557. Persoonia 2016, 37, 218–403. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.J.; Hardy, G.E.S.J.; Smith, D.; Summerell, B.A.; Cano-Lira, J.F.; Guarro, J.; Houbraken, J.; et al. Fungal Planet description sheets: 625–715. Persoonia 2017, 39, 270–467. [Google Scholar] [CrossRef]
- Gao, Y.; Du, X.; Xu, W.; Fan, R.; Zhang, X.; Yang, S.; Chen, X.; Lv, J.; Luo, Z. Fungal diversity in deep sea sediments from East Yap Trench and their denitrification potential. Geomicrobiol. J. 2020, 37, 848–858. [Google Scholar] [CrossRef]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From bio-chemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef]
- Luo, Y.; Wei, X.; Yang, S.; Gao, Y.H.; Luo, Z.H. Fungal diversity in deep-sea sediments from the Magellan seamounts as revealed by a metabarcoding approach targeting the ITS2 regions. Mycology 2020, 11, 214–229. [Google Scholar] [CrossRef]
- Nagano, Y.; Miura, T.; Nishi, S.; Lima, A.O.; Nakayama, C.; Pellizari, V.H.; Fujikura, K. Fungal diversity in deep-sea sediments associated with asphalt seeps at the Sao Paulo Plateau. Deep-Sea Res. Part I 2017, 146, 59–67. [Google Scholar] [CrossRef]
- Ogaki, M.B.; Coelho, L.C.; Vieira, R.; Neto, A.A.; Zani, C.L.; Alves, T.M.A.; Junior, P.A.S.; Murta, S.M.F.; Barbosa, E.C.; Oliveira, J.G.; et al. Cultivable fungi present in deep-sea sediments of Antarctica: Taxonomy, diversity, and bioprospecting of bioactive compounds. Extremophiles 2020, 24, 227–238. [Google Scholar] [CrossRef]
- Ogaki, M.B.; Pinto, O.H.B.; Vieira, R.; Neto, A.A.; Convey, P.; Carvalho-Silva, M.; Rosa, C.A.; Câmara, P.E.A.S.; Rosa, L.H. Fungi present in Antarctic deep-sea sediments assessed using DNA metabarcoding. Microb. Ecol. 2021, 82, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Park, M.S.; Oh, S.Y.; Fong, J.J.; Houbraken, J.; Lim, Y.W. The diversity and ecological roles of Penicillium in intertidal zones. Sci. Rep. 2019, 9, 13540. [Google Scholar] [CrossRef] [PubMed]
- Rédou, V.; Navarri, M.; Meslet-Cladière, L.; Barbier, G.; Burgaud, G. Species richness and adaptation of marine fungi from deep-subseafloor sediments. Appl. Environ. Microbiol. 2015, 81, 3571–3583. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Luo, Z.H.; Guo, S.; Pang, K.L. Fungal community analysis in the deep-sea sediments of the Pacific Ocean assessed by comparison of ITS, 18S and 28S ribosomal DNA regions. Deep-Sea Res. I Oceanogr. Res. Pap. 2016, 109, 51–60. [Google Scholar] [CrossRef]
- Xu, W.; Gong, L.F.; Pang, K.L.; Luo, Z.H. Fungal diversity in deep-sea sediments of a hydrothermal vent system in the southwest Indian ridge. Deep-Sea Res. I Oceanogr. Res. Pap. 2018, 131, 16–26. [Google Scholar] [CrossRef]
- Xu, W.; Gao, Y.H.; Gong, L.F.; Li, M.; Pang, K.L.; Luo, Z.H. Fungal diversity in the deep-sea hadal sediments of the Yap Trench by cultivation and high throughput sequencing methods based on ITS rRNA gene. Deep-Sea Res. I Oceanogr. Res. Pap. 2019, 145, 125–136. [Google Scholar] [CrossRef]
- Wentzel, L.C.P.; Inforsato, F.J.; Montoya, Q.V.; Rossin, B.G.; Nascimento, N.R.; Rodrigues, A.; Sette, L.D. Fungi from admiralty bay (King George Island, Antarctica) soils and marine sediments. Microb. Ecol. 2019, 77, 12–24. [Google Scholar] [CrossRef]
- Yang, S.; Xu, W.; Gao, Y.; Chen, X.; Luo, Z.H. Fungal diversity in deep-sea sediments from Magellan seamounts environment of the western Pacific revealed by high-throughput Illumina sequencing. J. Microbiol. 2020, 58, 841–852. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, N.F.; Zhang, Y.Q.; Liu, H.Y.; Yu, L.Y. Diversity and distribution of fungal communities in the marine sediments of Kongsfjorden, Svalbard (High Arctic). Sci. Rep. 2015, 5, 14524. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Wang, G.H.; Xu, X.Y.; Nong, X.H.; Wang, J.; Amin, M.; Qi, S.H. Exploring fungal diversity in deep-sea sediments from Okinawa trough using high-throughput Illumina sequencing. Deep Sea Res. Part I Oceanogr. Res. Pap. 2016, 116, 99–105. [Google Scholar] [CrossRef]
- Zhou, Y.; Gao, X.; Shi, C.; Li, M.; Jia, W.; Shao, Z.; Yan, P. Diversity and antiaflatoxigenic activities of culturable filamentous fungi from deep-sea sediments of the south Atlantic ocean. Mycobiology 2021, 49, 151–160. [Google Scholar] [CrossRef]
- Batista-García, R.A.; Sutton, T.; Jackson, S.A.; Tovar-Herrera, O.E.; Balcázar-López, E.; del Rayo Sanchez-Carbente, M.; Sánchez-Reyes, A.; Dobson, A.D.; Folch-Mallol, J.L. Characterization of lignocellulolytic activities from fungi isolated from the deep-sea sponge Stelletta normani. PLoS ONE 2017, 12, e0173750. [Google Scholar] [CrossRef] [PubMed]
- Bolaños, J.; De León, L.F.; Ochoa, E.; Darias, J.; Raja, H.A.; Shearer, C.A.; Miller, A.N.; Vanderheyden, P.; Porras-Alfaro, A.; Caballero-George, C. Phylogenetic diversity of sponge-associated fungi from the Caribbean and the Pacific of Panama and their in vitro effect on angiotensin and endothelin receptors. Mar. Biotechnol. 2015, 17, 533–564. [Google Scholar] [CrossRef] [PubMed]
- Bovio, E.; Garzoli, L.; Poli, A.; Prigione, V.; Firsova, D.; McCormack, G.P.; Varese, G.C. The culturable mycobiota associated with three Atlantic sponges, including two new species: Thelebolus balaustiformis and T. spongiae. Fungal Syst. Evol. 2018, 1, 141–167. [Google Scholar] [CrossRef] [PubMed]
- Bovio, E.; Garzoli, L.; Poli, A.; Luganini, A.; Villa, P.; Musumeci, R.; Mccormack, G.P.; Cocuzza, C.E.; Gribaudo, G.; Mehiri, M.; et al. Marine Fungi from the sponge Grantia compressa: Biodiversity, chemodiversity, and biotechnological potential. Mar. Drugs 2019, 17, 220. [Google Scholar] [CrossRef]
- Calabon, M.S.; Sadaba, R.B.; Campos, W.L. Fungal diversity of mangrove- associated sponges from New Washington, Aklan, Philippines. Mycology 2019, 10, 6–21. [Google Scholar] [CrossRef]
- Handayani, D.; Artasasta, M.A.; Safirna, N.; Ayuni, D.F.; Tallei, T.E.; Hertiani, T. Fungal isolates from marine sponge Chelonaplysilla sp.: Diversity, antimicrobial and cytotoxic activities. Biodiversitas 2020, 21, 1954–1960. [Google Scholar] [CrossRef]
- Lekshmi, N.; Umar, M.D.; Dhaneesha, M.; Rojin, J.; Ravinesh, R.; Sajeevan, T.P. Endophytic fungi isolated from the marine sponges as a source of potential bioactive compounds. J. Aquat. Biol. Fish. 2020, 8, 58–66. [Google Scholar]
- Azevedo, E.; Barata, M.; Marques, M.I.; Caeiro, M.F. Lulworthia atlantica: A new species supported by molecular phylogeny and morphological analysis. Mycologia 2017, 109, 287–295. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Roux, J.J.; Richardson, D.M. Fungal Planet description sheets: 371–399. Persoonia 2015, 35, 264–327. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Hardy, G.S.J.; Gené, J.; Guarro, J.; Baseia, I.G.; García, D.; Gusmão, L.F.P.; Souza-Motta, C.M.; et al. Fungal planet description sheets: 716–784. Persoonia 2018, 40, 240–393. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef] [PubMed]
- Garzoli, L.; Gnavi, G.; Tamma, F.; Tosi, S.; Varese, G.C.; Picco, A.M. Sink or swim: Updated knowledge on marine fungi associated with wood substrates in the Mediterranean Sea and hints about their potential to remediate hydrocarbons. Prog. Oceanogr. 2015, 137, 140–148. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Abreu, A.C.; Hilário, S.; Alves, A. Diversity of marine fungi associated with wood baits in the estuary Ria de Aveiro with descriptions of Paralulworthia halima comb. nov., Remispora submersa sp. nov. and Zalerion pseudomaritima sp. nov. Mycologia 2021, 113, 664–683. [Google Scholar] [CrossRef]
- Greco, G.; Cutroneo, L.; Di Piazza, S.; Capello, M.; Zotti, M. Trapping of marine- derived fungi on wooden baits to select species potentially usable in mycoremediation. Ital. J. Mycol. 2020, 49, 101–115. [Google Scholar]
- Kunttu, P.; Pasanen, H.; Rämä, T.; Kulju, M.; Kunttu, S.-M.; Kotiranta, H. Diversity and ecology of aphyllophoroid fungi on driftwood logs on the shores of the Baltic Sea. Nord. J. Bot. 2020, 38, e02735. [Google Scholar] [CrossRef]
- Rämä, T.; Davey, M.L.; Nordén, J.; Halvorsen, R.; Blaalid, R.; Mathiassen, G.H.; Alsos, I.G.; Kauserud, H. Fungi sailing the Arctic Ocean: Speciose communities in North Atlantic driftwood as revealed by high-throughput amplicon sequencing. Microb. Ecol. 2016, 72, 295–304. [Google Scholar] [CrossRef]
- Tibell, S.; Tibell, L.; Pang, K.L.; Calabon, M.; Jones, E.B.G. Marine fungi of the Baltic Sea. Mycology 2020, 11, 195–213. [Google Scholar] [CrossRef]
- Hyde, K.D.; Jones, E.B.G. Marine mangrove fungi. Mar. Ecol. 1988, 9, 15–33. [Google Scholar] [CrossRef]
- Rojas-Jimenez, K.; Wurzbacher, C.; Bourne, E.C.; Chiuchiolo, A.; Priscu, J.C.; Grossart, H.P. Early diverging lineages within Cryptomycota and Chytridiomycota dominate the fungal communities in ice-covered lakes of the McMurdo Dry Valleys, Antarctica. Sci. Rep. 2017, 7, 15348. [Google Scholar] [CrossRef] [PubMed]
- Senga, M.; Yabe, S.; Nakamura, T.; Kagami, M. Influence of parasitic chytrids on the quantity and quality of algal dissolved organic matter (AOM). Water Res. 2018, 145, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.D.; Richards, T.A.; Hawksworth, D.L.; Bass, D. Validation and justification of the phylum name Cryptomycota phyl. Nov. IMA Fungus 2011, 2, 173–175. [Google Scholar] [CrossRef]
- Malve, H. Exploring the ocean for new drug developments: Marine pharmacology. J. Pharm. Bioallied Sci. 2016, 8, 83–91. [Google Scholar] [CrossRef]
- Demain, A.L.; Fang, A. The natural functions of secondary metabolites. Adv. Biochem. Eng. Biotechnol. 2000, 69, 1–39. [Google Scholar] [PubMed]
- Houbraken, J.; Frisvad, J.C.; Samson, R.A. Fleming’s penicillin producing strain is not Penicillium chrysogenum but P. rubens. IMA Fungus 2011, 2, 87–95. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Moore, B.S. Lessons from the past and charting the future of marine natural products drug discovery and chemical biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef]
- Jimenez, C. Marine natural products in medicinal chemistry. ACS Med. Chem. Lett. 2018, 9, 959–961. [Google Scholar] [CrossRef]
- Bugni, T.S.; Ireland, C.M. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar] [CrossRef]
- Imhoff, J.F. Natural products from marine fungi—Still an underrepresented resource. Mar. Drugs 2016, 14, 19. [Google Scholar] [CrossRef]
- Overy, D.P.; Bayman, P.; Kerr, R.G.; Bills, G.F. An assessment of natural product discovery from marine (sensu strictu) and marine-derived fungi. Mycology 2014, 5, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Overy, D.P.; Rämä, T.; Oosterhuis, R.; Walker, A.K.; Pang, K.L. The neglected marine fungi, sensu stricto, and their isolation for natural products’ discovery. Mar. Drugs 2019, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Silber, J.; Kramer, A.; Labes, A.; Tasdemir, D. From discovery to production: Biotechnology of marine fungi for the production of new antibiotics. Mar. Drugs 2016, 14, 137. [Google Scholar] [CrossRef] [PubMed]
- Okutani, K. Gliotoxin produced by a strain of Aspergillus isolated from marine mud. Bull. Jap. Soc. Sci. Fish. 1977, 43, 995–1000. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. In Natural Product Reports; Royal Society of Chemistry: London, UK, 2019; pp. 122–173. [Google Scholar]
- Papon, N.; Copp, B.R.; Courdavault, V. Marine drugs: Biology, pipelines, current and future prospects for production. Biotechnol. Adv. 2022, 54, 107871. [Google Scholar] [CrossRef]
- Jaspars, M.; De Pascale, D.; Andersen, J.H.; Reyes, F.; Crawford, A.D.; Ianora, A. The marine biodiscovery pipeline and ocean medicines of tomorrow. J. Mar. Biol. Assoc. 2016, 96, 151–158. [Google Scholar] [CrossRef]
- Gowda, G.A.; Djukovic, D. Overview of mass spectrometry-based metabolomics: Opportunities and challenges. Methods Mol. Biol. 2014, 1198, 3–12. [Google Scholar]
- Labine, L.M.; Simpson, M.J. The use of nuclear magnetic resonance (NMR) and mass spectrometry (MS)–based metabolomics in environmental exposure assessment. Curr. Opin. Environ. Sci. Health 2020, 15, 7–15. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Hilário, S.; Tacão, M.; Van de Peer, Y.; Alves, A.; Esteves, A.C. Genome and Metabolome MS-Based Mining of a Marine Strain of Aspergillus affinis. J. Fungi 2021, 7, 1091. [Google Scholar] [CrossRef]
- Gonçalves, M.F.M.; Hilário, S.; Van de Peer, Y.; Esteves, A.C.; Alves, A. Genomic and Metabolomic Analyses of the Marine Fungus Emericellopsis cladophorae: Insights into Saltwater Adaptability Mechanisms and Its Biosynthetic Potential. J. Fungi 2022, 8, 31. [Google Scholar] [CrossRef]
- Stuart, K.A.; Welsh, K.; Walker, M.C.; Edrada-Ebel, R. Metabolomic tools used in marine natural product drug discovery. Expert Opin. Drug Discov. 2020, 15, 499–522. [Google Scholar] [CrossRef] [PubMed]
- Wolfender, J.L.; Marti, G.; Thomas, A.; Bertrand, S. Current approaches and challenges for the metabolite profiling of complex natural extracts. J. Chromatogr. A 2015, 1382, 136–164. [Google Scholar] [CrossRef] [PubMed]
- Rotter, A.; Barbier, M.; Bertoni, F.; Bones, A.M.; Cancela, M.L.; Carlsson, J.; Carvalho, M.F.; Cegłowska, M.; Chirivella-Martorell, J.; Conk Dalay, M.; et al. The essentials of marine biotechnology. Front. Mar. Sci. 2021, 8, 158. [Google Scholar] [CrossRef]
- Van Santen, J.A.; Jacob, G.; Singh, A.L.; Aniebok, V.; Balunas, M.J.; Bunsko, D.; Neto, F.C.; Castaño-Espriu, L.; Chang, C.; Clark, T.N.; et al. The natural products atlas: An open access knowledge base for microbial natural products discovery. ACS Cent. Sci. 2019, 5, 1824–1833. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, M.; Prakash, P.; Jayabaskaran, C.; Bhat, S.G. Multi-functional bioactive secondary metabolites derived from endophytic fungi of marine algal origin. Curr. Res. Microb. Sci. 2021, 2, 100037. [Google Scholar]
- Taritla, S.; Kumari, M.; Kamat, S.; Bhat, S.G.; Jayabaskaran, C. Optimization of physicochemical parameters for production of cytotoxic secondary metabolites and apoptosis induction activities in the culture extract of a marine algal–derived endophytic fungus Aspergillus sp. Front. Pharmacol. 2021, 12, 542891. [Google Scholar] [CrossRef]
- Parthasarathy, R.; Chandrika, M.; Rao, H.C.; Kamalraj, S.; Jayabaskaran, C.; Pugazhendhi, A. Molecular profiling of marine endophytic fungi from green algae: Assessment of antibacterial and anticancer activities. Process. Biochem. 2020, 96, 11–20. [Google Scholar] [CrossRef]
- Barra, L.; Barac, P.; König, G.M.; Crüsemann, M.; Dickschat, J.S. Volatiles from the fungal microbiome of the marine sponge Callyspongia cf. flammea. Org. Biomol. Chem. 2017, 15, 7411–7421. [Google Scholar] [CrossRef]
- Abd El-Hady, F.K.; Abdel-Aziz, M.S.; Shaker, K.H.; El-Shahid, Z.A.; Ibrahim, L.S. Antioxidant, acetylcholinesterase and α-Glucosidase potentials of metabolites from the marine fungus Aspergillus unguis RSPG_204 associated with the sponge (Agelas sp.). Int. J. Pharm. Sci. Rev. Res. 2015, 30, 272–278. [Google Scholar]
- Yan, M.X.; Mao, W.J.; Liu, X.; Wang, S.Y.; Xia, Z.; Cao, S.J.; Li, J.; Qin, L.; Xian, H.L. Extracellular polysaccharide with novel structure and antioxidant property produced by the deep-sea fungus Aspergillus versicolor N(2)bC. Carbohydr. Polym. 2016, 147, 272–281. [Google Scholar] [CrossRef]
- Teixeira, T.R.; Santos, G.S.; Turatti, I.C.C.; Paziani, M.H.; Kress, M.R.Z.; Colepicolo, P.; Debonsi, H.M. Characterization of the lipid profile of Antarctic brown seaweeds and their endophytic fungi by gas chromatography–mass spectrometry (GC–MS). Polar Biol. 2019, 42, 1431–1444. [Google Scholar] [CrossRef]
- Vinale, F.; Salvatore, M.M.; Nicoletti, R.; Staropoli, A.; Manganiello, G.; Venneri, T.; Borrelli, F.; DellaGreca, M.; Salvatore, F.; Andolfi, A. Identification of the main metabolites of a marine-derived strain of Penicillium brevicompactum using LC and GC MS techniques. Metabolites 2020, 10, 55. [Google Scholar] [CrossRef] [PubMed]
- De Jesus, H.C.; Jeller, A.H.; Debonsi, H.M.; Alves, P.B.; Porto, A.L. Multiple monohydroxylation products from rac-camphor by marine fungus Botryosphaeria sp. isolated from marine alga Bostrychia radicans. J. Braz. Chem. Soc. 2017, 28, 498–504. [Google Scholar] [CrossRef]
- Bhatnagar, V.S.; Bandyopadhyay, P.; Rajacharya, G.H.; Sarkar, S.; Poluri, K.M.; Kumar, S. Amelioration of biomass and lipid in marine alga by an endophytic fungus Piriformospora indica. Biotechnol. Biofuels 2019, 12, 176. [Google Scholar] [CrossRef]
- De Paula, S.F.C.; Porto, A.L.M. Cascate reactions of progesterone by mycelia and culture broth from marine-derived fungus Aspergillus sydowii CBMAI 935. Biocatal. Agric. Biotechnol. 2020, 25, 101546. [Google Scholar] [CrossRef]
- Liaw, C.C. Bioactive polypeptides from marine-derived fungi, Trichoderma spp. by LC-MS/MS and molecular networking analysis. Planta Med. 2015, 81, PM_148. [Google Scholar] [CrossRef]
- Loges, L.A.; Silva, D.B.; Paulino, G.V.; Landell, M.F.; Macedo, A.J. Polyketides from marine-derived Aspergillus welwitschiae inhibit Staphylococcus aureus virulence factors and potentiate vancomycin antibacterial activity in vivo. Microb. Pathog. 2020, 143, 104066. [Google Scholar] [CrossRef]
- Hwang, J.-Y.; Lee, J.-H.; Park, S.C.; Lee, J.; Oh, D.-C.; Oh, K.-B.; Shin, J. New peptides from the marine-derived fungi Aspergillus allahabadii and Aspergillus ochraceopetaliformis. Mar. Drugs 2019, 17, 488. [Google Scholar] [CrossRef]
- Saraiva, N.N.; Rodrigues, B.S.F.; Jimenez, P.C.; Guimaraes, L.A.; Torres, M.C.M.; Rodrigues-Filho, E.; Pfenning, L.H.; Abreu, L.M.; Mafezoli, J.; De Mattos, M.C.; et al. Cytotoxic compounds from the marine-derived fungus Aspergillus sp. revovered from the sediments of the Brazilian coast. Nat. Prod. Res. 2014, 29, 1545–1550. [Google Scholar] [CrossRef]
- Farha, A.K.; Hatha, A.M. Bioprospecting potential and secondary metabolite profile of a novel sediment-derived fungus Penicillium sp. ArCSPf from continental slope of Eastern Arabian Sea. Mycology 2019, 10, 109–117. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Amin, M.; Xu, X.Y.; Qi, S.H. Antifouling potentials and metabolite profiles of two marine-derived fungal isolates. Nat. Prod. Commun. 2018, 13, 423–426. [Google Scholar] [CrossRef]
- Wang, Y.; Mou, Y.; Dong, Y.; Wu, Y.; Liu, B.; Bai, J.; Yan, D.; Zhang, L.; Feng, D.; Pei, Y.; et al. Diphenyl ethers from a marine-derived Aspergillus sydowii. Mar. Drugs 2018, 16, 451. [Google Scholar] [CrossRef] [PubMed]
- Hulikere, M.M.; Joshi, C.G.; Ananda, D.; Poyya, J.; Nivya, T. Antiangiogenic, wound healing and antioxidant activity of Cladosporium cladosporioides (Endophytic Fungus) isolated from seaweed (Sargassum wightii). Mycology 2016, 7, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Navarri, M.; Jégou, C.; Bondon, A.; Pottier, S.; Bach, S.; Baratte, B.; Ruchaud, S.; Barbier, G.; Burgaud, G.; Fleury, Y. Bioactive metabolites from the deep subseafloor fungus Oidiodendron griseum UBOCC-A-114129. Mar. Drugs 2017, 15, 111. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Ding, W.; Ma, Z. Induced production of cytochalasans in co-culture of marine fungus Aspergillus flavipes and actinomycete Streptomyces sp. Nat. Prod. Res. 2016, 30, 1718–1723. [Google Scholar] [CrossRef]
- Hill, R.; Leitch, I.J.; Gaya, E. Targeting Ascomycota genomes: What and how big? Fungal Biol. Rev. 2021, 36, 52–59. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information. Available online: https://www.ncbi.nlm.nih.gov (accessed on 28 October 2021).
- JGI Genome Portal. Available online: https://genome.jgi.doe.gov/portal/ (accessed on 28 October 2021).
- Scharf, D.H.; Brakhage, A.A. Engineering fungal secondary metabolism: A roadmap to novel compounds. J. Biotechnol. 2013, 163, 179–183. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. Chembiochem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Pan, R.; Bai, X.; Chen, J.; Zhang, H.; Wang, H. Exploring structural diversity of microbe secondary metabolites using OSMAC strategy: A literature review. Front. Microbiol. 2019, 10, 294. [Google Scholar] [CrossRef]
Figure 1.
Timeline showing important events concerning marine fungi taxonomy. Mya: millions of years ago.
Figure 1.
Timeline showing important events concerning marine fungi taxonomy. Mya: millions of years ago.

Figure 2.
Challenges and adaptations of marine fungi to the marine environment.

Figure 3.
Fungal tree of life including the updated phylum-level classification of fungi, illustrating the morphological characteristics of the spores that are adapted for dispersal in marine habitats. Phyla which contain marine fungal lineages are depicted in blue.
Figure 3.
Fungal tree of life including the updated phylum-level classification of fungi, illustrating the morphological characteristics of the spores that are adapted for dispersal in marine habitats. Phyla which contain marine fungal lineages are depicted in blue.

Figure 4.
Diversity of marine fungal habitats. The figure was created with BioRender.com (accessed on 9 January 2022) [38].
Figure 4.
Diversity of marine fungal habitats. The figure was created with BioRender.com (accessed on 9 January 2022) [38].

Figure 5.
Diversity of metabolites’ classes produced by marine fungi and their biological activities with potential applications.
Figure 5.
Diversity of metabolites’ classes produced by marine fungi and their biological activities with potential applications.

Figure 6.
General representation of the pipeline for drug discovery in marine fungi.

Table 1.
Substrata from which marine fungi have been identified. Only research published after 2015 were included.
Table 1.
Substrata from which marine fungi have been identified. Only research published after 2015 were included.
| Substrata | Studies |
|---|---|
| Algae | [31,35,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56] |
| Corals | [37,57,58] |
| Invertebrates | [59,60,61] |
| Mangroves | [2,62,63,64,65,66,67,68,69,70,71,72,73] |
| Ocean crust | [74,75,76,77] |
| Plastic debris | [78,79,80,81,82,83,84,85,86,87,88,89] |
| Salt marsh plants | [81,82,83] |
| Sandy beaches | [84,85] |
| Seagrasses and seaweeds | [51,52,53,55,86,87,88,89,90,91,92,93,94,95,96,97,98] |
| Sea and ice water | [23,24,25,42,44,45,82,99,100,101,102,103,104,105,106,107,108,109,110,111] |
| Sediments or soil | [23,26,46,99,100,101,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131] |
| Sponges | [40,44,58,100,132,133,134,135,136,137,138] |
| Wood (drift or submerged) | [42,44,46,64,114,139,140,141,142,143,144,145,146,147,148] |
Table 3.
Overview of the marine fungal genomes sequenced.
| Family or Class | Species | Strain | Host/Substrate | NCBI Accession Number | JGI Project ID |
|---|---|---|---|---|---|
| Aspergillaceae | A. affinis | CMG 70 | Sea water | JAGXNN000000000 | - |
| Aspergillus flocculosus | CBS 112785 | Saltern | - | 1052366 | |
| Bionectriaceae | Emericellopsis maritima | TS7 | Sponge | JAFMPH000000000 | 1092660 |
| E. cladophorae | MUM 19.33 | Cladophora sp. | JAGIXG000000000 | - | |
| Chaetomiaceae | Achaetomium macrosporum | CBS 532.94 | Mangrove mud | - | 1151607 |
| Trichocladium antarcticum | CBS 123565 | Antarctic lichen | - | 1018949 | |
| Dothideomycetes | Lineolata rhizophorae | ATCC 16933 | Marine environment | JAAEJC000000000 | 1051209 |
| Halojulellaceae | Halojulella avicenniae | ATCC 66911 | Roots of Avicennia | - | 1042899 |
| Halosphaeriaceae | Corollospora maritima | CBS 119819 | Marine detritus | - | 1024020 |
| Lulworthiaceae | Lindra thalassiae | JK4322 | Thalassia sp. | - | 1070663 |
| Pleosporaceae | Decorospora gaudefroyi | CBS 332.63 | Salicornia sp. | JAADKE000000000 | 1032354 |
| Paradendryphiella salina | CBS 112865 | Fucus serratus | CAAAGB000000000 | - | |
| Sporormiaceae | Westerdykella ornata | CBS 379.55 | Mangrove mud | JAAEJA000000000 | 1019763 |
| Teratosphaeriaceae | Hortaea werneckii | MC848 | Sea water | PRJNA641248 PRJNA428320 | - |
| EXF-2788 | Hypersaline water | ||||
| MC873 | Sea water | ||||
| EXF-120 | Hypersaline water | ||||
| EXF-10513 | Deep sea water | ||||
| Torpedosporaceae | Torpedospora radiata | JK5252C | Woody debris | - | 1019473 |
| Trichocomaceae | Eurotium rubrum | CBS 135680 | Dead Sea brine | - | 1078839 |
| Penicillium antarcticum | IBT 31811 | Seaweed | MDYN00000000 | 1188177 | |
| Atheliaceae | Digitatispora marina | 008cD1.1 | Driftwood | - | 1081701 |
| Chytridiaceae | Chytridium lagenaria | Arg66 | Algae | - | 1098839 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gonçalves, M.F.M.; Esteves, A.C.; Alves, A. Marine Fungi: Opportunities and Challenges. Encyclopedia 2022, 2, 559-577. https://0-doi-org.brum.beds.ac.uk/10.3390/encyclopedia2010037
AMA Style
Gonçalves MFM, Esteves AC, Alves A. Marine Fungi: Opportunities and Challenges. Encyclopedia. 2022; 2(1):559-577. https://0-doi-org.brum.beds.ac.uk/10.3390/encyclopedia2010037
Chicago/Turabian StyleGonçalves, Micael F. M., Ana C. Esteves, and Artur Alves. 2022. "Marine Fungi: Opportunities and Challenges" Encyclopedia 2, no. 1: 559-577. https://0-doi-org.brum.beds.ac.uk/10.3390/encyclopedia2010037