
Development of Multilayer Mesenchymal Stem Cell Cell Sheets

Emmaus Life Sciences, Inc., 21250 Hawthorne Blvd., Suite 800, Torrance, CA 90503, USA
*
Author to whom correspondence should be addressed.
Int. J. Transl. Med. 2021, 1(1), 4-24; https://0-doi-org.brum.beds.ac.uk/10.3390/ijtm1010002
Submission received: 11 March 2021
/
Revised: 28 April 2021
/
Accepted: 8 May 2021
/
Published: 24 May 2021
Abstract
:Cell and gene therapies have been developing dramatically over the past decade. To face and adapt to the development of these new therapies, the Food and Drug Administration (FDA) wrote and updated new guidelines from 2016 and keep updating them. Mesenchymal stem cells (MSCs) are the most used cells for treatment, far ahead from the induced pluripotent stem cells (iPSCs), based on registered clinical trials at clinicaltrials.gov. They are widely used because of their differentiation capacity and their anti-inflammatory properties, but some controversies still require clear answers. Additional studies are needed to determine the dosage, the number, and the route of injections (location and transplantation method), and if allogenic MSCs are safe compared to autologous MSC injection, including their long-term effect. In this review, we summarize the research our company is conducting with the adipose stromal cells in engineering cell sheets and their potential application.
1. Introduction
Different types of stem cells are used for research and for translational medicine: embryonic stem cells (ESCs) [1], mesenchymal stem cells (MSCs) [2], and induced pluripotent stem cells (iPSCs) [3]. Ethical debates about the use of ESCs make their use more difficult for human application [4], even if clinical trials have been conducted recently with ESCs [4]. The iPSCs can be engineered by the transfection of four different factors into somatic cells [3], but as for the ESCs, the iPSCs have also raised ethical issues [5,6,7], and clinical trials are conducted all over the world, but mainly in the USA, China, Japan, and France [8]. The European Medical Agency approved cell therapies in the past few years: Chondrocelect was the first approved cell therapy in 2009 [9]. This review is not focused on presenting the different types of stem cells used for cell and gene therapies, but it is focused on the MSCs and cell sheet engineering with MSCs.
MSCs are very popular cells used in the research and over sixty-eight thousand publications involving the use of MSCs were published on PubMed, as of January 2021 [10]. MSCs are pluripotent stem cells that were discovered around 30 years ago [2], and they can be isolated from bone marrow, adipose tissue, Wharton’s jelly, periosteum, villous chorion, fetus, and dental pulp [2,11,12,13,14], and there are no ethical issues. In the organism, the function of the MSC is to support the structure of the organs but also to generate cells of the specific organ when it is required. They adhere quickly to the cell culture surface, and their morphology is fibroblastic. They can be cultured easily, and their stemness is characterized by the capacity of the MSC to self-renew and maintain the stemness properties, being passaged many times without karyotype alteration [15]. However, there is always a risk that MSCs could transform into sarcoma [16], requiring a long-term follow-up on preclinical animal studies and clinical trials, up to 15 years based on the Food and Drug Administration guidelines (FDA). MSCs curative properties and advantages can be divided in three different parts:
(a) They can differentiate into different types of cells (before or after transplantation), and the self-renewal property of the MSC is very important, but it is a critical characteristic that must be understood. Even with a self-renewal capacity, the aging of the MSC could be a major problem with an increase of mutation and loss of differentiation capacity [17,18]. In 2006, Dominici et al. published a list of the minimal criteria defining the MSC: MSC must express CD73, CD90, and CD105, and must lack the expression of CD14, CD19, CD34, CD45, and HLA-DR; in addition, MSCs must differentiate into cells originated from the three embryonic stem cell germ layers (endoderm, ectoderm, and mesoderm [19]) such as adipocytes, chondrocytes, and osteoblasts, as the most used [20].
(b) MSCs have anti-inflammatory potential and immune-modulatory properties, and promote cell growth and tissue repair, through the secretion of cytokines and extracellular vesicles [21]. In addition to this, the absence of HLA class II protein is a key factor, because MSCs could be used for allogeneic graft on patients, facilitating the use of MSCs in cell therapies. The activation of the HLA Class II leads to a rejection of transplanted cells or organs [22,23,24,25]. Functional MSCs do not express or express a very low level of HLA-DR (major histocompatibility complex class II, MHC II), meaning that MSCs have a lower immunogenicity than that of other cells [26,27,28,29]. In vitro studies showed that human bone marrow stem cells (BMSCs) are not recognized by T-lymphocytes but can suppress the proliferation of the T-lymphocyte [29]. For the past 20 years, human MSCs were used in animal studies, with successes, on the basis of the low probabilities that the xenotransplantation of human cells in animals will trigger an inflammatory response and the human MSC rejection. Human MSCs (hMSCs) were injected in different animals without any adverse events reported: mouse [30], rat [31], rabbit [32], zebrafish [32], swine [33], and dog [34], and as review for xenotransplantation of hMSCs [35]. This positive characteristic can be used for gene and cell therapy preclinical tests on animals before translational application, by using the MSCs that are planned to be utilized in the clinical trials (e.g., culture media, approved cells for clinical used by Federal Agencies). The absence or low immunogenicity of MSCs will allow their mass production, a better characterization, and the decrease of cost. In the other hand, MSCs act also as immunomodulators, by reducing inflammatory activity [36,37,38], and were used as a racehorse cure with no immunoreaction [39], for the bone repair of rats [40], in a human trial for Crohn’s disease [41,42,43,44,45], and for perianal fistula [46,47], as outlined in a review publication [48].
(c) An additional positive criterion is the large-scale manufacturing of the MSCs, which will provide enough cells for cell therapies [49,50]. Typing the key words “bioprocessing, mesenchymal stem cells” in PubMed, there are only 160 publications referring to the large-scale production methodology of the mesenchymal stem cells. The bioprocessing of any stem cells must be well planned and controlled, including the determination of the donors (inclusion/exclusion criteria), the methodology of isolation, the type of culture media, and the processes for the mass production [49]. Positive and negative outcomes of stem cell therapy for animal studies and clinical trials can be related with the modification in the stem cells’ bioprocessing [50]. It is encouraged to work with MSC providers (or any other cells) that have an approved chemistry, manufacturing, and control for clinical trials, to facilitate the transition from preclinical to clinical trials; but it is also important to determine the cell culture conditions in the preclinical phase that will be used in the clinical trials to ensure that the data obtained in the preclinical studies and the methodologies will be approved by the federal agencies.
MSCs are the most used stem cells in clinical trials, and MSCs have shown promising hope for patients in need of gene and cell therapies [51], and over 1220 clinical trials have been conducted over the world (keyword: mesenchymal stem cell at clinicaltrials.gov). Most of the clinical trials reached phase I and II, and very few of them reached phase III. MSCs can be easily isolated from different tissues. Even if bone marrow stem cells are still the MSCs used most often, the invasive procedure to isolate the bone marrow makes it more difficult and more stressful for the donors [52], compared with the use of adipose stem cells that can be easily isolated from liposuction [53,54]. In addition, the number of isolated adipose stromal cells from the liposuction can be 50,000 times higher than the number of bone marrow stem cells isolated from the bone marrow [55,56,57]. For all these reasons listed above, we decided to study the potential of the adipose stromal cells, a specific mesenchymal stem cell.
2. Mesenchymal Stem Cells Therapies
Engineering of tissues and organs with mesenchymal stem cells involves not only the stem cells but it could also involve biocompatible scaffolds, important for cell signaling stimulation and for transplantation [58,59]. The simplest and most economical methodology to treat patients with MSCs is the injection of isolated MSCs. On the other hand, a more complex approach consists in growing the cells in a 3D structure, using different methodologies for support, such as scaffolds and 3D printing. The efficiency of the cell therapy is based not only on the quality and the stem cell phenotype, but it is also related with the transplantation methodology of the cells.
Our company decided to develop cell sheets using mesenchymal stem cells, to target the cells on the damaged area, in absence of a specific scaffold, for different reasons: cheaper methodology, no additional step is necessary to prepare the culture dish to engineer cell sheet, absence of scaffold will not lead to fibrosis in the empty space left during the scaffold degradation, and harvesting of cell sheet requires strong cell–cell connection and extracellular matrix.
2.1. Injection of Single Cells
The cheapest and easiest way to use cells, for cell therapy, is the injection of single isolated cells. CAR-T cells (Chimeric antigen receptor T cells) were for the first time designed in 1989, by Gross et al. [60], and the T cell chimeric antigen receptor (CAR) technology developed exponentially until the approval for cancer therapy by the FDA in 2017 [61]. CAR-T cells, after their reprogramming, are expanded in isolated cells and injected in the patients. The cells migrate to their target and fight the cancer. In 1999, for the first time, BMSCs treated with 5-azacytidine formed myotube and were injected in rats that had myocardial injury. Injected cells improved the heart function and increased the angiogenesis [62].
The cells can be injected via intravenous, intraperitoneal, intraarterial, intracerebral, intrasplenic, or intramuscular routes or into the superior mesenteric vein. Two major parameters can be controlled for the injection: the posology of MSCs (number of cells injected) and the route of injection. The number of cells injected are calculated by only the absolute value of cells (×10y), which is reported in peer review and is not precise enough because the weight of the animals differs between the studies and between the animals from the same study.
Many publications reported the curative effect of the injected cells. For example, Dos Santos Ramalho et al. reported that the intravenous or intraperitoneal injection of BMSCs did not have a different effect in repairing the spinal cord [63]. In this study, female mice weighing 20–25 g were used, and the same dose of 8 × 105 cells was injected (when the mice weight can vary by 20–25% among the mice). Another study reported the intravenous injection of xenogeneic MSCs (from human Wharton jelly) and mice MSCs. In this study, 1 million cells were injected in the tail vein, and both types of cells had similar effect on controlling the neuroinflammation [64]. Many other studies reported absolute values of the injected number of cells: intravenous injection of 107 ASCs in dogs to treat acute spinal cord injury [65] and intravenous injection of 42 × 106 cells/animal in porcine model improving heart function after an acute myocardial infarction [66]. In all these studies, the results were positive and showed an improvement of the animal health. Another way to estimate the number of MSCs to be injected, and to be reproduced by other laboratories, is to inject a number of cells per kilogram. Indeed, for animal and human studies, the weight of individuals varies, per gender and per age [67].
However, the cons for the injection of single cells are the uncontrolled migration of the cells (cf. Section 2.3 Biodistribution) and the formation of aggregates after their injection. It was reported that injected cells can aggregate rapidly with lymphocytes, which decreases their mobility and access to the needed areas. Among all the MSC properties mentioned in the paragraph 1 Introduction, MSCs tend to aggregate in vitro and in vivo. Even if the MSC attachments on a plastic surface is a criterion for MSC, it was reported for the past decade that the MSCs can form aggregates. From few thousands or more MSCs, MSCs form 3D spheroid structures [68] spontaneously. In order to create spheroids, MSCs cannot be allowed to adhere the surface, which is achieved by using different techniques such as spinning the containers. Once the MSCs form spheroids their properties change, due to a mechano-physical modification and the reorganization of the cytoskeleton [69]. It is well known that modification of the cytoskeleton can modify the expression of genes [70,71]. A comparison of the transcriptome of MSC monolayer with MSC 3D spheroids shows that the expression of 3000 genes was modified: an increase of around 1700 genes and a decrease of around 1300 genes [72]. Many publications report that the differentiation potential of stem cells increases when they form a 3D structure. Bae et al. showed that the 3D structure of MSCs improves their capacity to differentiate into adipocyte and osteoblast, with or without being treated with azacytidine [73]. Our laboratory also noticed that the multilayer cell sheets of adipose stromal cells, which is also a 3D structure, as for the spheroids, differentiate faster than isolated adipose stromal cells (data not reported). The curative properties of the MSCs are also improved. For example, it was reported that the spheroid formation increased some MSC properties, such as their anti-inflammatory activity [68]. However, in vivo studies showed that an injected single MSC forms aggregates [74] in the blood stream, potentially triggering a thromboembolism in the animal or patient. Oeller et al. injected 1.5 million of bone marrow stromal cells (BMSCs), umbilical stem cells (UCMSCs), and adipose stromal stem cells (ASCs) in Fischer rats, in the tail vein. The authors noticed that thromboembolism occurs with the three types of injected cells, but less with the BMSC group [75]. BMSCs express less tissue-factor-mediated procoagulant than the two other types of cells, explaining the difference in the thromboembolism. Other studies reported similar results [76,77]. Such a health issue was also reported in clinical trials, jeopardizing the life of patients [78,79,80], which is a major issue that needs to be overcome to increase the safety for the treated patients.
This single cell injection became popular because of the low cost and the easy protocol. It was used with MSCs, injected as isolated cells, and they migrate over the body [81]. The migration of injected MSC cannot be controlled and their survival rate is low over time. The majority of the publications reported that the injected cells migrate to the lungs [81,82], then disappear very quickly from the lungs. For example, Lee et al. showed that after the intravenous injection of 2 million human MSCs (or of MDA-MB-231 breast cancer cell line) in a mouse, the cells migrated predominantly into lungs (80% of the total cells). They were also detected, in much lower quantity in the liver, spleen, and blood [83]. By measuring the “quantity” of Alu DNA sequences of the human cells, the authors confirmed that only 0.04% of the cells were present in the lungs at 48 h, and only 0.01% at 90 h after injection. The same percentage (0.04%) was found by adding the measured Alu DNA sequence by combining six other organs and fluids (blood, brain, heart, liver, pancreas, spleen, and kidneys). The biodistribution and the detection of the cells after transplantation will be discussed in the paragraph 2.3 Biodistribution.
The survival rate of the injected cells is very low (from 0.01% to 6%), and the cells disappear in a few days or weeks [83,84,85]. Based on the survival rate of the injected cells, although that there is no real evidence, it can be expected that the biological effects of the injected cells (paracrine, cell attachment, and cell differentiation) will occur in a very short time, if the curative effect can be noticed, or will not occur at all. The low survival rate of injected MSCs could be promising for acute/local health problems, while the treatment of chronic/systemic health problems will require a longer survival rate of the cells, and better cell targeting. To improve the survival of the cells, the apoptotic pathway was regulated (e.g., pretreatment of stem cells with ROCK inhibitor (Rho Kinase inhibitor)) and/or the cell adhesion was improved (e.g., use of decellularized organs). After cell sheet harvesting, the cell–cell connection, proteins located at the membrane and the extracellular matrix (ECM) of the cell sheet were preserved. The preservation of the ECM allows the grafting of the cell sheet on the tissues and increases the survival of the cells. The development of the cell sheet technology was a breakthrough to target the cells to the damaged organs that required cell/gene therapy.
2.2. Cell Sheet Technology
To improve cell survival, cell targeting, and cell therapy, different approaches were developed, such as 3D printing, organoids development, and the cell sheet technology. We will focus on the cell sheet technology. Tissue engineering was mentioned for the first time in 1993 by R Langer and JP Vacanti [86]. Transplantation of the stem cells required the use of biodegradable materials [87]; however, after the degradation of the scaffold, the empty space left by the degradation of the scaffold leads to a fibrosis [88]. To outcome this problems, thermoresponsive surface was developed. It allows the detachment of the cells, as a whole cell sheet, without the use of enzymes such as trypsin [89]. The cell sheets can be transferred on to another surface (dishes, organs, or cell sheets), because the extracellular matrix as well as the cell–cell contact were preserved [90,91]. Cell–cell contact and communication are very important for the organs’ function [92,93]. In addition to the thermoresponsive surface, other methodologies were developed to have the cell sheets in one piece: collagenase, amniotic membrane, electroresponsive/photoresponsive/pH-responsive/magnetic-responsive surface, and mechanical harvesting [94]. Instead of injecting single isolated cells, we have focused on engineering cell sheet with adipose stromal cells for different reasons, as explained in the introduction [95].
Adipose stromal cells can be isolated from different fatty tissues. The methodology of ASC isolation is less painful for the donor (compared to the isolation of bone marrow stromal cells), well controlled, and easy to perform [96,97]. The yield in ASC isolation is superior to the number of cells isolated from the bone marrow, but the population is more heterogeneous [98]. Because of the number of cells required for a patient’s treatment (in average, 2 × 106 cells/kg [99,100]), the in vitro expansion of ASCs is necessary. For animal and clinical trials, quality and safety controls of cultured ASCs will have to be performed to ensure the quality of the cells, for the patient safety and to obtain the best outcome on the targeted disease.
We have developed multilayer cell sheets, using adipose stromal cells [95]. Using specific cell culture conditions, the undifferentiated multilayer adipose stromal cell sheets have been engineered as well as differentiated cell sheets. In general, stem cells from the targeted organ are isolated and cultured, and cell sheets are engineered. For example, chondrocyte and synovial cells can be isolated from the articulation, grown in co-culture, on thermoresponsive surface to form a cartilage cell sheet [101]. The harvested cell sheet has been transplanted to patients and improved their cartilage defects. Harvesting of a patient’s stem cells is an approach to perform autologous transplantation. Dr. Sawa’s group successfully engineered and transplanted cardiac cell sheets, to treat cardiomyopathy [102]. The cardiac functions of the patients improved over time. The cell sheets were engineered using isolated stem cells from the vastus medialis, seeded on a thermoresponsive cell culture dish. The advantage of these approaches is that the transplantation will be autologous and the risk of rejection is decreased, even if it could happen since the cells were cultured for a certain period in vitro, before transplantation; this rejection risk might be lower than that of using allogeneic transplantation, but additional studies are required.
A major concern is when the stem cells needed to engineer the cell sheets are absent. ASC cell bank can be a solution because, as part of their primary criterion, ASCs must be able to differentiate into cells from the three different types of dermis (ectoderm, endoderm, and mesoderm) and could be used to treat many diseases. Allogeneic transplantation is an alternative to autologous transplantation for different reasons. Many stem cells can be amplified in a bioreactor and stored until the cells are required for a patient’s treatment. Moreover, by building a master cell bank of the same type of cells, we will know that all the cells are identical, and there are no or few variabilities between the cells [103]. In the contrary, autologous stem cell transplantation is limited by the number of cells available, the outcome of the treatment will vary based on the patients, and it will be more expensive than allogeneic transplantation because stem cells from each patient will have to be tested. Using commercially available ASCs, we succeeded to engineer chondrocyte multilayer cell sheets, harvested and characterized after differentiation treatment, as well as osteoblast cell sheets [95]. Bone marrow stem cells were used to engineer osteoblast cells sheets, using a specific scaffold [104]. For our studies, we did not use a particular scaffold to engineer the cell sheets, but we focused on the use of specific culture media. Cell sheets were harvested mechanically, using forceps and PVDF membrane, in absence of enzymatic treatment or without using a specific surface. There are different advantages to using the cell sheet technology. For example, the extracellular matrix and the cell–cell connection are well preserved, increasing the cell adhesion for transplantation and the cell survival, respectively [105]. In addition, it is possible to compile different types of cell sheets to mimic a functional organ, improving then the curative properties of the cell sheet [106]. Another advantage is to directly target the cell sheet on the damaged area. Transplantation of the cardiac cell sheet was performed directly on the heart damaged area due to the heart failure [102]. More recently, in addition to the biological properties of the cell sheets, different physical/mechanical properties are studied that could be part of the release criteria of the cell sheet before transplantation, such as elasticity of chondrocyte or epithelial cell sheets and the strength of myocardial cell sheets [107,108].
Contrary to the injection of isolated single cells, it is not possible to precisely control the number of transplanted cells. However, while most of the injected isolated single cells disappear in a short time, leading to less efficient treatment of the disease (cf. Section 2.1 Injection of Singles Cells), all the cells from a sheet are targeted on the damaged area, and more than one cell sheet could be transplanted to increase the number of cells [102].
2.3. Biodistribution
Biodistribution after cells injection/transplantation is a major concern for the federal agencies, especially to determine if the cells could be a threat to a patient’s health on a long-term treatment. When medications are absorbed by the patients, the medications are distributed, metabolized, and excreted after a certain period of time [109]. On the contrary, stem cell treatments are expected to or could have a very long-term effect. The timeline for the pharmacokinetics (PK) of cell treatment is different from that of the drugs PK, and cells should be followed up for an extended period, estimated to a year’s level.
Different noninvasive technological approaches were developed to study the biodistribution of transplanted cell, depending on if the final goal is a short- or long-term treatment:
(a) Permanent genetic modification of the cells, which could jeopardize their natural functions as well as patient safety (wrong differentiation, tumorigenicity, ectopic proliferation, impairment of organs functions, etc.) is necessary for a long-term follow-up.
(b) Direct cell labeling was used in animal studies to track the cells after transplantation. However, the cell follow-up will be possible over a short period of time (days to weeks) for animal/human studies because the signal due to the marker used in the direct labeling will decrease by half at each cell division, which will not comply with federal agencies’ expectations. In addition, the detection of the marker will not demonstrate the cell viability.
We will not discuss the short-term follow-up in detail because the federal agencies focus on the long-term effect of cell transplantation. For short-term follow-up (days to weeks), cells could be labeled directly during the cell culture period with agents that can be detected with a noninvasive method. For example, cells could be incubated with a nontoxic superparamagnetic iron oxide particles (SPIO) dose, before their transplantation, and tracked using magnetic resonance imaging (MRI) [110]. Axolotl blastema cells, labeled with SPIO, were detected up to 84 days after transplantation in an Axolotl [110]. Other compounds for direct labeling, such as the 19F-perfluorocarbon, could be used for single cell tracking [111] but it could be more challenging when the goal is to label a multilayer cell sheet [112]. The decreasing rate of the tracker intensity signal is related with the frequency of cell division. It is expected that at each division, the signal will be decreased by half, and the signal might disappear in a short period of time because the survival rate of injected single cells is low. Another aspect of direct cell labeling is that the detection of the tracker does not reflect the cell viability and functionality, which is contrary to the genetic modification of the cells for long-term follow-up.
Even if federal agencies do not have specific guidelines for the follow-up and biodistribution on patients after cell transplantation, they strongly recommend a long-term follow-up during the animal preclinical studies (based on the animal lifespan) to determine if there is a risk or not for the patients during the clinical trials. For example, for transplantation of corneal-like cell sheets on patients with limbal stem cell deficiency, the average time of follow-up was 2 years, with a maximum of 7.5 years [113,114]. Different methodologies were developed to study cell migration and location, after their transplantation (Table 1). To follow up stem cells in small animals (i.e., rats or mice) after transplantation, genetic engineering and genetic modification of the cells with reporter genes can be used, but there are always questions and doubts about the modified stem cells safety after transplantation. In addition, the genetic modification could alter the function of the transplanted cells, and the outcome of the treatment could be different from the outcome obtained using unmodified stem cells. Small animal size allows to detect fluorescent proteins and luminescence [115,116,117]. However, for bigger animals (i.e., pig or dogs) or for human clinical trials, it will not be possible to detect the fluorescence or luminescence of the cells. An alternative was developed for a long-term follow-up in bigger animals or humans. Cells could be transfected with proteins that will modify compounds injected in the body, and that could be detected in big animals or patients. For example, the expression of thymidine kinase (HSC1-tk) in the stem cells is an approach that will phosphorylate the 18F-FEAU ([18F]fluoro-5-ethyl-1-beta-D-arabinofuranosyluracil) injected. The phosphorylated form of 18F-FEAU will be trapped in the cells and accumulate there [118]. The accumulation of 18F-FEAU in the cells will allow the long-term follow-up of the injected cells, and it is a proof that the injected cells are alive and functional. However, the expression of the herpes simplex virus thymidine kinase in cells could increase its cytotoxicity and trigger an inflammatory reaction [119]. In addition, the accumulation of the 18F-FEAU could be toxic for the cells and for the patients in the long term. Transferrin receptor was also used in stem cells to detect the accumulation of iron in the cells. Iron is an important metal for cell function, and it is transported in the cells and out of the cells when the levels of iron are too high. Overexpression of human transferrin in mouse neural stem cells showed an accumulation of extracellular iron in the cells [120]. Using magnetic resonance imaging, the authors showed that overexpressing transferrin receptor cells accumulate more iron than the nonmodified cells. The higher accumulation of iron in the cells shows their functionality. However, if the export of cellular iron is not controlled or sufficient, the high intracellular iron concentration could be harmful for the patient’s safety, because it is related with various pathologies [121].
At the best of our knowledge, there is no approved genetic cell modification to study over a long period of time the fate of the transplanted cells. Side effect of genetically modified cells will be a major concern that will require a large group of patients to determine the safety of the modifications. However, this approach has two advantages: it will be possible to have a long-term follow-up of the cells if the trackers are expressed by the cells and the cell viability could be confirmed.
Migration of injected isolated MSCs can be affected by different factors: adhesion molecules, chemokines, cytokines, and matrix metalloproteinases [142,143,144,145,146,147,148]. It is not clear, but injected MSCs must pass by the lung vascular system, and they are usually retained there for less than 24 h. It is not well reported, but the decrease of the cell number in the lung could be due to the low survival rate or by an extra migration in other organs [149]. The rate of clearance of the MSCs in the lungs was shown to be related to one factor at least: the nature of the membranous proteins. It was shown recently that the nature of the cell surface markers can influence the biodistribution of different types of MSCs. Umbilical cord MSCs are cleared much faster than the bone marrow MSCs, due to the nature of the proteins located at the cellular membrane [150]. As reported above, stem cells tend to aggregate and can clot blood vessels. In addition, MSCs express adhesion proteins that will increase their attachment to the membrane of the endothelial cells. For example, they express CD166 and CD105, which are considered as MSC markers. In addition, other adhesion molecules, such as VCAM-1 [147], ICAM-1 [148], and P-selectin [146], are expressed, and they play a role in the modulation of inflammation by the MSCs. Chemokines receptors are also important to guide MSCs migration towards damaged areas, and it could be used to determine the moment of cell injection to obtain the best curative effect from the injected MSCs [145].
For example, it was shown in many reports that injected MSCs migrate to inflammatory regions that are present in the body. However, when the inflammation is controlled and diminished, migration and differentiation of the MSCs are modified. Pretreatment of MSCs with a conditioned culture media from activated proinflammatory macrophages increased the migration and the attachment of MSCs, in vitro. However, when the MSCs are pretreated with a conditioned culture media from anti-inflammatory macrophages, MSCs have a higher capability to differentiate into osteoblasts and regenerate a bone [151]. As mentioned in the introduction, MSCs tend to migrate more and also to migrate to the inflamed area [152]. This MSC characteristic could be used if the patients need to have an inflammatory reaction decreased or to treat autoimmune reactions [153]. Many clinical trials are conducted over the world to treat inflammation with MSCs, but there is only one stem cell therapy approved in the USA, by the FDA, to treat patients having blood production problems. For the past year, recent studies have been working and testing on using MSCs for COVID-19 treatment, based on MSC/MSC extracellular vesicles anti-inflammatory properties [154,155,156]. However, in other cases, the inflammatory reaction could decrease the efficacy of the MSC treatment. For example, corneal epithelial cell sheets were developed using oral mucosal epithelial cells, and it was reported that to increase the chance of successful transplantation, it is recommended to control the inflammation on the cornea [157].
In addition, it is very important to notice that the cell culture conditions will affect the MSC phenotype [158,159], and the age of the MSC donors can also influence the migration properties of the injected MSCs [81]. Because the phenotype/behavior varies among different MSCs, it is strongly recommended by the FDA to use the MSCs for animal studies as they were manufactured as for clinical trials, to estimate their behavior and the regenerative properties during the clinical trials.
In summary, there is no existing methodology for a noninvasive long-term follow-up of transplanted cells on patients. The only alternative is to ensure that there is no difference between the transplanted patient group with the nontransplanted group on tumor formation rate, impairment of organ functions, and the adverse and severe adverse events. From necropsies or from biopsies, potential approaches to detect the transplanted cells could be as follows: usage of cells from the opposite gender, or long-term follow-up of the animals/patients to ensure that the rate of tumor formation is similar between all groups.
3. Cryopreservation of the Cell Sheets
In parallel with building organ banks [160], the cryopreservation of adult stem cells is increasingly prevalent around the world, anticipating the future needs for adult stem cells for research and also for patients’ cell therapy [161,162]. The production of organ-based stem cells must be well controlled and regulated, in GMP (Good Manufacturing Practices) facilities, based on well-controlled and defined stem cells banks. In the 21st century, cell-based therapies are growing exponentially, leading to the development of methodologies to target the cells on the damaged organs. There are different methodologies to engineer organ-based stem cells and to target them on injured organs: seeding and growing of stem cells on scaffolds [163], 3D printing [58], decellularized organs [164], and cell sheet [165]. As for the organs that must be discarded after a short period of time, those organ-based stem cells must be discarded if they are not used rapidly, but this problem could be overcome if the cell-based organs could be vitrified and stored for a long term in nitrogen liquid.
Cryopreservation was first used to freeze cells, to build cell banks for laboratory purposes but also to create banks of spermatozoids and oocytes for in vitro fecundation purposes [166]. To preserve the functionality and properties of the cells, cryoprotective agent (CPA) were used at different concentrations [167,168]. Indeed, during the freezing steps and in the absence of CPA, ice forms into the cells and damages the cells. To avoid intracellular ice formation, CPA agents such as dimethyl sulfoxide, ethylene glycol, and butanediol must be added to the freezing solutions [169,170]. This methodology is called vitrification, a process that decreases the ice formation in vivo freezing biological samples. The development of this methodology started to be used for organs banking, as the shortage of organs for transplantation is a serious problem in the medical field [171]. In the USA, the waiting list for transplant increased by 160 patients a day, but only 95 transplantations take place every day (source: https://optn.transplant.hrsa.gov/, accessed on 30 January 2014), with an average death rate of 20 patients in the waiting list per day (source: https://www.americantransplantfoundation.org/, accessed on 30 January 2014). [172]. Furthermore, there is a high chance of wasting donated organs if the patient does exist at the same time, causing unused organs to be discarded [173].
Different approaches were then developed to preserve the organ’s function before the transplantation. The first one was to preserve the organs at body temperature, using machine pumps [174]. This approach reduced the production of toxic compounds by the organ and the accumulation of reactive oxidant species that damage the cells [172,174]. The second protocol was to preserve the organs in a cold storage. Cold storage led to ischemia/reperfusion-related damages during the reperfusion of the organs. In addition, the organs can only be stored for a very short period, up to 24 h for livers and kidneys [175]. To overcome the injuries due to ischemia/reperfusion and the short preservation time of the organs, laboratories started to develop techniques to completely freeze the entire organ. To reach this goal, high concentrations of cryoprotective agent (CPA) are used, making the solutions around the cells and in the cells viscous. The first organ cryopreservation was done by Dr. Karow Jr., in 1961, when he cryopreserved a functional heart with glycerol [176]. Other organs were cryopreserved after this first experiment. For example, in 2002, Van Den Broecke et al. cryopreserved human ovaries and grafted them back into nude mice. Injection of follicle-stimulating hormone induced the release of primary and secondary follicles, indicating that the human cryopreserved ovaries were fully functional [177]. The protocols are still under development, and no standard vitrification protocol is used in the medical field. However, tremendous efforts are put into developing organs cryopreservation technology. The accumulated knowledge of organ cryopreservation was transferred recently in the cryopreservation of stem cell-based cell sheets.
Stem cell-based therapy is a new approach to treat diseases or damaged organs that can have a significant high impact on the patient’s life. Different techniques were developed to target the cells on the damaged organ’s area: simple injection, or the use of scaffolds to directly graft the cells on the organs, or by using a thermoresponsive surface to harvest cell sheets [89,178]. Cell sheets technology is an exciting approach to graft the cells directly on the damaged organs. This cell-based therapy preserves the cell–cell interaction, their physiological activities, and the extracellular matrix produced by the cell sheets [165]. The cell–cell interaction and physiological activities are sensitive cell parameters that must be preserved to maintain the cell sheet functionality. Preservation of the extracellular matrix is an important parameter to the cell sheet, not only to maintain its polarity, but also to decrease the necessity of suture to graft the cell sheet on the tissue [179].
As for the organs preserved in cold or warm ischemia conditions for transplantation, cell sheets should be used immediately to treat the patients. As for many biological living products based on cells, the shelf life of the products can be short, especially if the cells are still in proliferation. It is then necessary to deliver the final product to the patients as soon as possible, but it is not always possible. The shipping logistics can be very complex and difficult for a rapid delivery while preserving the product’s curative properties. Our laboratory did not test the shelf lifespan of the cell sheets, but it can be hypothesized that the lifespan can be short, especially if the culture media is not replaced (such as during the shipping conditions). Sipuleucel-T by Dendreon Corporation, Inc. is an FDA-approved cell-based cancer immunotherapy for prostate cancer. If Sipuleucel-T is left at room temperature for more than 3 h, Sipuleucel-T cannot be used for patient treatment and must be discarded [180]. Each cell-based therapeutic product has its own shelf lifespan, and it will define the shipping/timing for transplantation on patients. Our hypothesis is based on the nature of culture media that is used for engineering and maintaining the cell sheets. Fully prepared culture media can be stored at 4 °C, for at least 1 month, depending on the composition of the culture media. However, when the culture media is placed at 37 °C, the lifespan of the culture media decreases rapidly (L-glutamine half-life in solution at 37 °C is 6 days (source Sigma-Aldrich). Overculturing the cell sheet could lead to a change in the phenotype, a modification of their curative properties, and a higher chance of mutations that could be a threat for the patients. In addition, if the cell sheets must be transported or shipped over a long distance, the cells sheet will consume the culture media nutrients, leading to an acidification of the culture media due to the oxidative metabolism. If the transport is prolonged, the culture media cannot be replaced during the transport. The acidification and impoverishment in nutrients of the culture media could damage the cells and lead to cell death. Even if the transport of the cell sheet is well prepared, random events could delay the shipment, as occurred recently in February 2021, when more than half of the USA was hit by extremely cold weather, disturbing the normal lives of humans, including shipment, transport, power supplies, and accessibility of the surgery’s facilities for the patients and for the surgeons. Moreover, if the cell sheets are ready for transplantation but it is not possible to transplant for any reason, cryopreservation of the cell sheets is the only known approach for long-term storage and preservation. The knowledge of the cell sheet’s cryopreservation is still at the dawn of this technology, and additional work is required to improve and perfect the technology. To the best of our knowledge, there are only five publications reporting the vitrification of cell sheets [95,181,182,183,184], and very few publications are discussing the transport of the cell sheets to the clinics [185]. Among the five publications, two publications reported the preservation of the vitrified cell sheet curative properties and three of them showed the maintenance of cell sheet morphology and the expression of few proteins. However, none of these studies reported the potential cell sheet stress due to the vitrification process.
Cryopreservation of tissues results in the modification of the gene expression due to the cold storage, thawing process, and the use of cryoprotectant agents. Cryoprotectant (CPA), used in the cryopreservation of single cells, is accompanied by the modification of gene expressions. In a complex biological structure, the expression of the genes, such as fatty acid binding protein 5 (Fabp5), fructose-1,6-bisphosphate 1 (Fbp1), and T-box 20 (Tbx20), were decreased in cryopreserved mouse blastocyst [186]. The authors concluded that the slow freezing methodology, compared to the vitrification method, was inducing most of the changes in the gene expression. On the contrary, a similar study with sheep blastocyst reported that the vitrification methodology affected more genes than the slow freezing did [187]. The cryopreservation of the cells not only modifies their phenotype as mentioned above, but it has also an effect on the energy deprivation, decrease of the cellular pH, disorganization of the cytoskeleton (cytoplasmic and nuclear), which can modify the gene expression [70,188], and production of reactive oxygen species, which can induce cell injuries (lipid peroxidation, DNA damage, DNA mutations, and apoptosis) [189,190,191,192,193,194,195]. Sperm and oocytes are the most common cryopreserved cells, but even with a long experience and practice in this field, it is still very challenging to vitrify them [196]. For example, an increase of the global DNA methylation in horse semen, during their cryopreservation, could be one factor explaining the low fertility rate of insemination [191]. Modification of the DNA methylation pattern of cells and cell sheets, during the freezing/thawing steps, could affect any gene controlling tumorigenicity [197]. In porcine blastocysts, the expression of IGF2 and IGF2R were downregulated by the vitrification process, but their expression was like that of nonvitrified cells, in the presence of ethylene glycol [198]. The DNA methylation pattern of those two imprinted genes was not studied in the vitrified cell sheets, but the alteration of the gene imprinting, during the cryopreservation/thawing steps, could affect their expression and causes diseases in a long term [199,200,201,202,203]. Additional work on durable epigenetic modification should be conducted to improve the cryopreservation/thawing methodology.
In the field of cell sheet cryopreservation, very few publications and data are available [181,182,183,184,204]. Ohkawara et al. vitrified cardio-cell sheets and stored them in nitrogen liquid from 2 days to 3 months, without affecting the cell sheet morphology on a macroscopic level, and their cell therapy function, after transplantation on the heart. The improvement of the heart function was similar between the fresh and the cryopreserved cell sheets [182]. Even if there is no difference in the cell viability between fresh and vitrified cell sheets, the authors reported a significant increase of the apoptotic level in the vitrified group [182]. In addition, the vitrification process increased the expression of VEGF, HGF, and SDF-1 mRNA, and decreased the expression of mt-ND1 and mt-ATP6. The expression of those genes was measured after the thawing process. However, their expression was not analyzed a few days after their thawing, and their level of expression could have been the same as the level of expression of fresh cell sheets. Indeed, Cordeiro et al. reported that the expression of stress genes increased during the first 24 h after the cells were exposed to ethylene glycol [205]. The expression of the genes returned to partial normal level 72 h after removing the ethylene glycol [205], which could be the case after cell sheet thawing.
The preservation of the biological samples after their cryopreservation depends also on the warming/thawing methodology. During the warming process, the formation of ice crystals can damage the cell integrity, and then the function of the biological sample. The rate of temperature changes during the warming period must be optimized depending on the tissue [206,207].
The objective of the cell sheet vitrification is to build a bank of the cell sheets and transplant them, after the thawing test. Because of the low immunogenicity of the MSCs, this methodology will allow to have a large bank of cell sheets, available in a very short time for the patients, for allogeneic transplantation. Vitrified human myoblast cell sheets had a similar curative effect on the heart after their transplantation on mice. This not only means that the cell sheets maintained their curative properties, but also shows that xenogeneic transplantation with stem cells can be performed and by consequence, allogeneic transplantation is possible [182]. Because cell sheet therapies are a recent medical treatment, we do not have long-term experience on the potential harm of cryopreserved cell sheets after transplantation, and further studies are required to improve new cryopreservation and thawing protocols [208,209,210].
4. Conclusions
Engineering cell sheet with mesenchymal stem cells is a very important branch in the field of the regenerative medicine, which has been growing for the past 20 years. In addition to the use of the cell sheet for cell therapy, numerous other applications could be used (Figure 1):
Funding
This research received no external funding.
Acknowledgments
This work was supported by Emmaus Medical, Inc. (Torrance, CA, USA). Y. Niihara is the Chairman of the Board of Directors and CEO of Emmaus Medical, Inc. J. Ochiai and J. Oliva are employees of Emmaus Medical, Inc. The authors have no other relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript apart from those disclosed.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kimlin, L.; Kassis, J.; Virador, V. 3D in vitro tissue models and their potential for drug screening. Expert Opin. Drug Discov. 2013, 8, 1455–1466. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilic, D.; Ogilvie, C. Concise Review: Human Embryonic Stem Cells-What Have We Done? What Are We Doing? Where Are We Going? Stem Cells 2017, 35, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moradi, S.; Mahdizadeh, H.; Saric, T.; Kim, J.; Harati, J.; Shahsavarani, H.; Greber, B.; Moore, J.B.T. Research and therapy with induced pluripotent stem cells (iPSCs): Social, legal, and ethical considerations. Stem Cell Res. Ther. 2019, 10, 341. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.L. Some Ethical Concerns About Human Induced Pluripotent Stem Cells. Sci. Eng. Ethics 2016, 22, 1277–1284. [Google Scholar] [CrossRef]
- Zacharias, D.G.; Nelson, T.J.; Mueller, P.S.; Hook, C.C. The science and ethics of induced pluripotency: What will become of embryonic stem cells? Mayo Clin. Proc. 2011, 86, 634–640. [Google Scholar] [CrossRef] [Green Version]
- Deinsberger, J.; Reisinger, D.; Weber, B. Global trends in clinical trials involving pluripotent stem cells: A systematic multi-database analysis. NPJ Regen. Med. 2020, 5, 15. [Google Scholar] [CrossRef]
- Yu, T.T.L.; Gupta, P.; Ronfard, V.; Vertes, A.A.; Bayon, Y. Recent Progress in European Advanced Therapy Medicinal Products and Beyond. Front. Bioeng. Biotechnol. 2018, 6, 130. [Google Scholar] [CrossRef] [Green Version]
- Zomer, H.D.; Vidane, A.S.; Goncalves, N.N.; Ambrosio, C.E. Mesenchymal and induced pluripotent stem cells: General insights and clinical perspectives. Stem Cells Cloning 2015, 8, 125–134. [Google Scholar] [CrossRef]
- Miana, V.V.; Gonzalez, E.A.P. Adipose tissue stem cells in regenerative medicine. Ecancermedicalscience 2018, 12, 822. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Zhang, R.; Zou, Q.; Chen, Y.; Zhou, M.; Li, X.; Ran, R.; Chen, Q. Comparison of the Biological Characteristics of Mesenchymal Stem Cells Derived from the Human Placenta and Umbilical Cord. Sci. Rep. 2018, 8, 5014. [Google Scholar] [CrossRef]
- Charbord, P. Bone marrow mesenchymal stem cells: Historical overview and concepts. Hum. Gene Ther. 2010, 21, 1045–1056. [Google Scholar] [CrossRef] [Green Version]
- Jackson, W.M.; Nesti, L.J.; Tuan, R.S. Potential therapeutic applications of muscle-derived mesenchymal stem and progenitor cells. Expert Opin. Biol. Ther. 2010, 10, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Neri, S. Genetic Stability of Mesenchymal Stromal Cells for Regenerative Medicine Applications: A Fundamental Biosafety Aspect. Int. J. Mol. Sci. 2019, 20, 2406. [Google Scholar] [CrossRef] [Green Version]
- Hatina, J.; Kripnerova, M.; Houfkova, K.; Pesta, M.; Kuncova, J.; Sana, J.; Slaby, O.; Rodriguez, R. Sarcoma Stem Cell Heterogeneity. Adv. Exp. Med. Biol. 2019, 1123, 95–118. [Google Scholar]
- Yang, Y.K.; Ogando, C.R.; Wang See, C.; Chang, T.Y.; Barabino, G.A. Changes in phenotype and differentiation potential of human mesenchymal stem cells aging in vitro. Stem Cell Res. Ther. 2018, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Liu, C.; Xie, Z.; Song, P.; Zhao, R.C.; Guo, L.; Liu, Z.; Wu, Y. Epigenetic dysregulation in mesenchymal stem cell aging and spontaneous differentiation. PLoS ONE 2011, 6, e20526. [Google Scholar] [CrossRef] [Green Version]
- Sathananthan, H.; Selvaraj, K.; Clark, J. The fine structure of human germ layers in vivo: Clues to the early differentiation of embryonic stem cells in vitro. Reprod. Biomed. Online 2011, 23, 227–233. [Google Scholar] [CrossRef] [Green Version]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Poggi, A.; Zocchi, M.R. Immunomodulatory Properties of Mesenchymal Stromal Cells: Still Unresolved “Yin and Yang”. Curr. Stem Cell Res. Ther. 2019, 14, 344–350. [Google Scholar] [CrossRef]
- Kolb, H.J. Hematopoietic stem cell transplantation and cellular therapy. HLA 2017, 89, 267–277. [Google Scholar] [CrossRef]
- Hao, M.; Wang, R.; Wang, W. Cell Therapies in Cardiomyopathy: Current Status of Clinical Trials. Anal. Cell Pathol. 2017, 2017, 9404057. [Google Scholar] [CrossRef] [Green Version]
- Duelen, R.; Sampaolesi, M. Stem Cell Technology in Cardiac Regeneration: A Pluripotent Stem Cell Promise. EBioMedicine 2017, 16, 30–40. [Google Scholar] [CrossRef] [Green Version]
- Thakkar, U.G.; Vanikar, A.V.; Trivedi, H.L. Stem cell therapy: An emerging modality in glomerular diseases. Cytotherapy 2017, 19, 333–348. [Google Scholar] [CrossRef]
- Mohammadi, Z.; Afshari, J.T.; Keramati, M.R.; Alamdari, D.H.; Ganjibakhsh, M.; Zarmehri, A.M.; Jangjoo, A.; Sadeghian, M.H.; Ameri, M.A.; Moinzadeh, L. Differentiation of adipocytes and osteocytes from human adipose and placental mesenchymal stem cells. Iran J. Basic Med. Sci. 2015, 18, 259–266. [Google Scholar]
- Technau, A.; Froelich, K.; Hagen, R.; Kleinsasser, N. Adipose tissue-derived stem cells show both immunogenic and immunosuppressive properties after chondrogenic differentiation. Cytotherapy 2011, 13, 310–317. [Google Scholar] [CrossRef]
- Mitchell, J.B.; McIntosh, K.; Zvonic, S.; Garrett, S.; Floyd, Z.E.; Kloster, A.; Di Halvorsen, Y.; Storms, R.W.; Goh, B.; Kilroy, G.; et al. Immunophenotype of human adipose-derived cells: Temporal changes in stromal-associated and stem cell-associated markers. Stem Cells 2006, 24, 376–385. [Google Scholar] [CrossRef]
- Tse, W.T.; Pendleton, J.D.; Beyer, W.M.; Egalka, M.C.; Guinan, E.C. Suppression of allogeneic T-cell proliferation by human marrow stromal cells: Implications in transplantation. Transplantation 2003, 75, 389–397. [Google Scholar] [CrossRef]
- Ra, J.C.; Shin, I.S.; Kim, S.H.; Kang, S.K.; Kang, B.C.; Lee, H.Y.; Kim, Y.J.; Jo, J.Y.; Yoon, E.J.; Choi, H.J.; et al. Safety of intravenous infusion of human adipose tissue-derived mesenchymal stem cells in animals and humans. Stem Cells Dev. 2011, 20, 1297–1308. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, N.; Razavi, S.; Mardani, M.; Esfandiari, E.; Salehi, H.; Zarkesh Esfahani, S.H. Transplantation of human adipose-derived stem cells enhances remyelination in lysolecithin-induced focal demyelination of rat spinal cord. Mol. Biotechnol. 2014, 56, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Lasso, J.M.; Perez Cano, R.; Castro, Y.; Arenas, L.; Garcia, J.; Fernandez-Santos, M.E. Xenotransplantation of human adipose-derived stem cells in the regeneration of a rabbit peripheral nerve. J. Plast. Reconstr. Aesthet. Surg. 2015, 68, e189–e197. [Google Scholar] [CrossRef] [PubMed]
- Jun Hong, S.; Rogers, P.I.; Kihlken, J.; Warfel, J.; Bull, C.; Deuter-Reinhard, M.; Feng, D.; Xie, J.; Kyle, A.; Merfeld-Clauss, S.; et al. Intravenous xenogeneic transplantation of human adipose-derived stem cells improves left ventricular function and microvascular integrity in swine myocardial infarction model. Catheter. Cardiovasc. Interv. 2015, 86, E38–E48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelatti, M.V.; Gomes, J.P.; Vieira, N.M.; Cangussu, E.; Landini, V.; Andrade, T.; Sartori, M.; Petrus, L.; Zatz, M. Transplantation of Human Adipose Mesenchymal Stem Cells in Non-Immunosuppressed GRMD Dogs is a Safe Procedure. Stem Cell Rev. Rep. 2016, 12, 448–453. [Google Scholar] [CrossRef]
- Lin, C.S.; Lin, G.; Lue, T.F. Allogeneic and xenogeneic transplantation of adipose-derived stem cells in immunocompetent recipients without immunosuppressants. Stem Cells Dev. 2012, 21, 2770–2778. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Huang, X.; Wang, H.; Liu, X.; Zhang, T.; Wang, Y.; Hu, D. The challenges and promises of allogeneic mesenchymal stem cells for use as a cell-based therapy. Stem Cell Res. Ther. 2015, 6, 234. [Google Scholar] [CrossRef] [Green Version]
- Asano, K.; Yoshimura, S.; Nakane, A. Adipose Tissue-Derived Mesenchymal Stem Cells Attenuate Staphylococcal Enterotoxin A-Induced Toxic Shock. Infect. Immun. 2015, 83, 3490–3496. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.J.; Traktuev, D.O.; March, K.L. Therapeutic potential of adipose-derived stem cells in vascular growth and tissue repair. Curr. Opin. Organ Transplant. 2010, 15, 86–91. [Google Scholar] [CrossRef]
- Ricco, S.; Renzi, S.; Del Bue, M.; Conti, V.; Merli, E.; Ramoni, R.; Lucarelli, E.; Gnudi, G.; Ferrari, M.; Grolli, S. Allogeneic adipose tissue-derived mesenchymal stem cells in combination with platelet rich plasma are safe and effective in the therapy of superficial digital flexor tendonitis in the horse. Int. J. Immunopathol. Pharmacol. 2013, 26 (Suppl. 1), 61–68. [Google Scholar] [CrossRef] [Green Version]
- Wen, C.; Yan, H.; Fu, S.; Qian, Y.; Wang, D.; Wang, C. Allogeneic adipose-derived stem cells regenerate bone in a critical-sized ulna segmental defect. Exp. Biol. Med. 2016, 241, 1401–1409. [Google Scholar] [CrossRef] [Green Version]
- Panes, J.; Garcia-Olmo, D.; Van Assche, G.; Colombel, J.F.; Reinisch, W.; Baumgart, D.C.; Dignass, A.; Nachury, M.; Ferrante, M.; Kazemi-Shirazi, L.; et al. Expanded allogeneic adipose-derived mesenchymal stem cells (Cx601) for complex perianal fistulas in Crohn’s disease: A phase 3 randomised, double-blind controlled trial. Lancet 2016, 388, 1281–1290. [Google Scholar] [CrossRef]
- Garcia-Arranz, M.; Herreros, M.D.; Gonzalez-Gomez, C.; de la Quintana, P.; Guadalajara, H.; Georgiev-Hristov, T.; Trebol, J.; Garcia-Olmo, D. Treatment of Crohn’s-Related Rectovaginal Fistula With Allogeneic Expanded-Adipose Derived Stem Cells: A Phase I-IIa Clinical Trial. Stem Cells Transl. Med. 2016, 5, 1441–1446. [Google Scholar] [CrossRef]
- Park, K.J.; Ryoo, S.B.; Kim, J.S.; Kim, T.I.; Baik, S.H.; Kim, H.J.; Lee, K.Y.; Kim, M.; Kim, W.H. Allogeneic adipose-derived stem cells for the treatment of perianal fistula in Crohn’s disease: A pilot clinical trial. Colorectal Dis. 2016, 18, 468–476. [Google Scholar] [CrossRef]
- Molendijk, I.; Bonsing, B.A.; Roelofs, H.; Peeters, K.C.; Wasser, M.N.; Dijkstra, G.; van der Woude, C.J.; Duijvestein, M.; Veenendaal, R.A.; Zwaginga, J.J.; et al. Allogeneic Bone Marrow-Derived Mesenchymal Stromal Cells Promote Healing of Refractory Perianal Fistulas in Patients With Crohn’s Disease. Gastroenterology 2015, 149, 918–927.e6. [Google Scholar] [CrossRef] [Green Version]
- De la Portilla, F.; Alba, F.; Garcia-Olmo, D.; Herrerias, J.M.; Gonzalez, F.X.; Galindo, A. Expanded allogeneic adipose-derived stem cells (eASCs) for the treatment of complex perianal fistula in Crohn’s disease: Results from a multicenter phase I/IIa clinical trial. Int. J. Colorectal Dis. 2013, 28, 313–323. [Google Scholar] [CrossRef]
- Park, E.J.; Kang, J.; Baik, S.H. Treatment of faecal incontinence using allogeneic-adipose-derived mesenchymal stem cells: A study protocol for a pilot randomised controlled trial. BMJ Open 2016, 6, e010450. [Google Scholar] [CrossRef]
- Garcia-Olmo, D.; Herreros, D.; Pascual, I.; Pascual, J.A.; Del-Valle, E.; Zorrilla, J.; De-La-Quintana, P.; Garcia-Arranz, M.; Pascual, M. Expanded adipose-derived stem cells for the treatment of complex perianal fistula: A phase II clinical trial. Dis. Colon Rectum 2009, 52, 79–86. [Google Scholar] [CrossRef]
- Trounson, A.; McDonald, C. Stem Cell Therapies in Clinical Trials: Progress and Challenges. Cell Stem Cell 2015, 17, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Hassan, M.; Yazid, M.D.; Yunus, M.H.M.; Chowdhury, S.R.; Lokanathan, Y.; Idrus, R.B.H.; Ng, A.M.H.; Law, J.X. Large-Scale Expansion of Human Mesenchymal Stem Cells. Stem Cells Int. 2020, 2020, 9529465. [Google Scholar] [CrossRef]
- Panchalingam, K.M.; Jung, S.; Rosenberg, L.; Behie, L.A. Bioprocessing strategies for the large-scale production of human mesenchymal stem cells: A review. Stem Cell Res. Ther. 2015, 6, 225. [Google Scholar] [CrossRef] [Green Version]
- Galipeau, J.; Sensebe, L. Mesenchymal Stromal Cells: Clinical Challenges and Therapeutic Opportunities. Cell Stem Cell 2018, 22, 824–833. [Google Scholar] [CrossRef] [Green Version]
- Hernigou, P.; Homma, Y.; Flouzat Lachaniette, C.H.; Poignard, A.; Allain, J.; Chevallier, N.; Rouard, H. Benefits of small volume and small syringe for bone marrow aspirations of mesenchymal stem cells. Int. Orthop. 2013, 37, 2279–2287. [Google Scholar] [CrossRef]
- Wu, C.; Laswell, S.; Mentz, J.A.; Morales, R. Vibration Exposure Safety Guidelines for Surgeons Using Power-Assisted Liposuction (PAL). Aesthet. Surg. J. 2020. [Google Scholar] [CrossRef]
- Qu, Y.; Luan, J.; Mu, D.; Wang, Q.; Li, Z.; Liu, T.; Fu, S. Does Water-Jet Force Affect Cryopreserved Adipose-Derived Stem Cells? Evidence of Improved Cell Viability and Fat Graft Survival. Ann. Plast. Surg. 2020. [Google Scholar] [CrossRef]
- Tsuji, W.; Rubin, J.P.; Marra, K.G. Adipose-derived stem cells: Implications in tissue regeneration. World J. Stem Cells 2014, 6, 312–321. [Google Scholar] [CrossRef]
- Sachs, P.C.; Francis, M.P.; Zhao, M.; Brumelle, J.; Rao, R.R.; Elmore, L.W.; Holt, S.E. Defining essential stem cell characteristics in adipose-derived stromal cells extracted from distinct anatomical sites. Cell Tissue Res. 2012, 349, 505–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casteilla, L.; Planat-Benard, V.; Laharrague, P.; Cousin, B. Adipose-derived stromal cells: Their identity and uses in clinical trials, an update. World J. Stem Cells 2011, 3, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.S.; Yesantharao, P.; Huang, C.Y.; Mattson, G.; Boktor, J.; Fukunishi, T.; Zhang, H.; Hibino, N. 3D bioprinting using stem cells. Pediatr. Res. 2018, 83, 223–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, X.; Avci-Adali, M.; Alarcin, E.; Cheng, H.; Kashaf, S.S.; Li, Y.; Chawla, A.; Jang, H.L.; Khademhosseini, A. Development of hydrogels for regenerative engineering. Biotechnol. J. 2017, 12, 1600394. [Google Scholar] [CrossRef]
- Gross, G.; Waks, T.; Eshhar, Z. Expression of immunoglobulin-T-cell receptor chimeric molecules as functional receptors with antibody-type specificity. Proc. Natl. Acad. Sci. USA 1989, 86, 10024–10028. [Google Scholar] [CrossRef] [Green Version]
- Filley, A.C.; Henriquez, M.; Dey, M. CART Immunotherapy: Development, Success, and Translation to Malignant Gliomas and Other Solid Tumors. Front. Oncol. 2018, 8, 453. [Google Scholar] [CrossRef] [Green Version]
- Tomita, S.; Li, R.K.; Weisel, R.D.; Mickle, D.A.; Kim, E.J.; Sakai, T.; Jia, Z.Q. Autologous transplantation of bone marrow cells improves damaged heart function. Circulation 1999, 100, II-247–II-256. [Google Scholar] [CrossRef] [Green Version]
- Ramalho, B.D.S.; Almeida, F.M.; Sales, C.M.; de Lima, S.; Martinez, A.M.B. Injection of bone marrow mesenchymal stem cells by intravenous or intraperitoneal routes is a viable alternative to spinal cord injury treatment in mice. Neural Regen. Res. 2018, 13, 1046–1053. [Google Scholar]
- Lykhmus, O.; Koval, L.; Voytenko, L.; Uspenska, K.; Komisarenko, S.; Deryabina, O.; Shuvalova, N.; Kordium, V.; Ustymenko, A.; Kyryk, V.; et al. Intravenously Injected Mesenchymal Stem Cells Penetrate the Brain and Treat Inflammation-Induced Brain Damage and Memory Impairment in Mice. Front. Pharmacol. 2019, 10, 355. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.U.; Yoon, Y.; Choi, K.U.; Jo, K.R.; Kim, N.; Lee, E.; Kim, W.H.; Kweon, O.K. Therapeutic Effects of Intravenous Injection of Fresh and Frozen Thawed HO-1-Overexpressed Ad-MSCs in Dogs with Acute Spinal Cord Injury. Stem Cells Int. 2019, 2019, 8537541. [Google Scholar] [CrossRef] [Green Version]
- Lim, M.; Wang, W.; Liang, L.; Han, Z.B.; Li, Z.; Geng, J.; Zhao, M.; Jia, H.; Feng, J.; Wei, Z.; et al. Intravenous injection of allogeneic umbilical cord-derived multipotent mesenchymal stromal cells reduces the infarct area and ameliorates cardiac function in a porcine model of acute myocardial infarction. Stem Cell Res. Ther. 2018, 9, 129. [Google Scholar] [CrossRef] [Green Version]
- Kabat, M.; Bobkov, I.; Kumar, S.; Grumet, M. Trends in mesenchymal stem cell clinical trials 2004-2018: Is efficacy optimal in a narrow dose range? Stem Cells Transl. Med. 2020, 9, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Bartosh, T.J.; Ylostalo, J.H.; Mohammadipoor, A.; Bazhanov, N.; Coble, K.; Claypool, K.; Lee, R.H.; Choi, H.; Prockop, D.J. Aggregation of human mesenchymal stromal cells (MSCs) into 3D spheroids enhances their antiinflammatory properties. Proc. Natl. Acad. Sci. USA 2010, 107, 13724–13729. [Google Scholar] [CrossRef] [Green Version]
- Cesarz, Z.; Tamama, K. Spheroid Culture of Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 9176357. [Google Scholar] [CrossRef] [Green Version]
- Spichal, M.; Fabre, E. The Emerging Role of the Cytoskeleton in Chromosome Dynamics. Front. Genet. 2017, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Viita, T.; Vartiainen, M.K. From Cytoskeleton to Gene Expression: Actin in the Nucleus. Handb. Exp. Pharmacol. 2017, 235, 311–329. [Google Scholar]
- Potapova, I.A.; Gaudette, G.R.; Brink, P.R.; Robinson, R.B.; Rosen, M.R.; Cohen, I.S.; Doronin, S.V. Mesenchymal stem cells support migration, extracellular matrix invasion, proliferation, and survival of endothelial cells in vitro. Stem Cells 2007, 25, 1761–1768. [Google Scholar] [CrossRef]
- Bae, Y.J.; Kwon, Y.R.; Kim, H.J.; Lee, S.; Kim, Y.J. Enhanced differentiation of mesenchymal stromal cells by three-dimensional culture and azacitidine. Blood Res. 2017, 52, 18–24. [Google Scholar] [CrossRef]
- Bartosh, T.J.; Ylostalo, J.H. Mesenchymal Stem Cell (MSC) Aggregate Formation in vivo. Bio-Protocol 2014, 4, e1181. [Google Scholar] [CrossRef] [Green Version]
- Oeller, M.; Laner-Plamberger, S.; Hochmann, S.; Ketterl, N.; Feichtner, M.; Brachtl, G.; Hochreiter, A.; Scharler, C.; Bieler, L.; Romanelli, P.; et al. Selection of Tissue Factor-Deficient Cell Transplants as a Novel Strategy for Improving Hemocompatibility of Human Bone Marrow Stromal Cells. Theranostics 2018, 8, 1421–1434. [Google Scholar] [CrossRef] [Green Version]
- Perlee, D.; de Vos, A.F.; Scicluna, B.P.; Maag, A.; Mancheno, P.; de la Rosa, O.; Dalemans, W.; Florquin, S.; Van’t Veer, C.; Lombardo, E.; et al. Role of tissue factor in the procoagulant and antibacterial effects of human adipose-derived mesenchymal stem cells during pneumosepsis in mice. Stem Cell Res. Ther. 2019, 10, 286. [Google Scholar] [CrossRef] [Green Version]
- Tatsumi, K.; Ohashi, K.; Matsubara, Y.; Kohori, A.; Ohno, T.; Kakidachi, H.; Horii, A.; Kanegae, K.; Utoh, R.; Iwata, T.; et al. Tissue factor triggers procoagulation in transplanted mesenchymal stem cells leading to thromboembolism. Biochem. Biophys. Res. Commun. 2013, 431, 203–209. [Google Scholar] [CrossRef]
- Coppin, L.C.F.; Smets, F.; Ambroise, J.; Sokal, E.E.M.; Stephenne, X. Infusion-related thrombogenesis by liver-derived mesenchymal stem cells controlled by anticoagulant drugs in 11 patients with liver-based metabolic disorders. Stem Cell Res. Ther. 2020, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Zhang, S.; Zhou, L.; Cai, J.; Tan, J.; Gao, X.; Zeng, Z.; Li, D. Thromboembolism Induced by Umbilical Cord Mesenchymal Stem Cell Infusion: A Report of Two Cases and Literature Review. Transplant. Proc. 2017, 49, 1656–1658. [Google Scholar] [CrossRef]
- Jung, J.W.; Kwon, M.; Choi, J.C.; Shin, J.W.; Park, I.W.; Choi, B.W.; Kim, J.Y. Familial occurrence of pulmonary embolism after intravenous, adipose tissue-derived stem cell therapy. Yonsei Med. J. 2013, 54, 1293–1296. [Google Scholar] [CrossRef] [Green Version]
- Fabian, C.; Naaldijk, Y.; Leovsky, C.; Johnson, A.A.; Rudolph, L.; Jaeger, C.; Arnold, K.; Stolzing, A. Distribution pattern following systemic mesenchymal stem cell injection depends on the age of the recipient and neuronal health. Stem Cell Res. Ther. 2017, 8, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurtz, A. Mesenchymal stem cell delivery routes and fate. Int. J. Stem Cells 2008, 1, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.H.; Pulin, A.A.; Seo, M.J.; Kota, D.J.; Ylostalo, J.; Larson, B.L.; Semprun-Prieto, L.; Delafontaine, P.; Prockop, D.J. Intravenous hMSCs improve myocardial infarction in mice because cells embolized in lung are activated to secrete the anti-inflammatory protein TSG-6. Cell Stem Cell 2009, 5, 54–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gyongyosi, M.; Blanco, J.; Marian, T.; Tron, L.; Petnehazy, O.; Petrasi, Z.; Hemetsberger, R.; Rodriguez, J.; Font, G.; Pavo, I.J.; et al. Serial noninvasive in vivo positron emission tomographic tracking of percutaneously intramyocardially injected autologous porcine mesenchymal stem cells modified for transgene reporter gene expression. Circ. Cardiovasc. Imaging 2008, 1, 94–103. [Google Scholar] [CrossRef] [Green Version]
- Le Maitre, C.L.; Baird, P.; Freemont, A.J.; Hoyland, J.A. An in vitro study investigating the survival and phenotype of mesenchymal stem cells following injection into nucleus pulposus tissue. Arthritis Res. Ther. 2009, 11, R20. [Google Scholar] [CrossRef] [Green Version]
- Langer, R.; Vacanti, J.P. Tissue engineering. Science 1993, 260, 920–926. [Google Scholar] [CrossRef] [Green Version]
- Lei, M.; Wang, X. Biodegradable Polymers and Stem Cells for Bioprinting. Molecules 2016, 21, 539. [Google Scholar] [CrossRef]
- Bruzauskaite, I.; Bironaite, D.; Bagdonas, E.; Bernotiene, E. Scaffolds and cells for tissue regeneration: Different scaffold pore sizes-different cell effects. Cytotechnology 2016, 68, 355–369. [Google Scholar] [CrossRef] [Green Version]
- Yamada, N.; Okano, T.; Sakai, H.; Karikusa, F.; Sawasaki, Y.; Sakurai, Y. Thermo-responsive polymeric surfaces; control of attachment and detachment of cultured cells. Markomol. Chem. 1990, 11, 571–576. [Google Scholar]
- Ohki, T.; Yamamoto, M. Esophageal regenerative therapy using cell sheet technology. Regen. Ther. 2020, 13, 8–17. [Google Scholar] [CrossRef]
- Kushida, A.; Yamato, M.; Konno, C.; Kikuchi, A.; Sakurai, Y.; Okano, T. Decrease in culture temperature releases monolayer endothelial cell sheets together with deposited fibronectin matrix from temperature-responsive culture surfaces. J. Biomed. Mater. Res. 1999, 45, 355–362. [Google Scholar] [CrossRef]
- Delarue, C.; Contesse, V.; Lenglet, S.; Sicard, F.; Perraudin, V.; Lefebvre, H.; Kodjo, M.; Leboulenger, F.; Yon, L.; Gallo-Payet, N.; et al. Role of neurotransmitters and neuropeptides in the regulation of the adrenal cortex. Rev. Endocr. Metab. Disord. 2001, 2, 253–267. [Google Scholar] [CrossRef]
- Yasuda, K. Dominant rule of community effect in synchronized beating behavior of cardiomyocyte networks. Biophys. Rev. 2020, 12, 481–501. [Google Scholar] [CrossRef]
- Lu, Y.; Zhang, W.; Wang, J.; Yang, G.; Yin, S.; Tang, T.; Yu, C.; Jiang, X. Recent advances in cell sheet technology for bone and cartilage regeneration: From preparation to application. Int. J. Oral Sci. 2019, 11, 17. [Google Scholar] [CrossRef] [Green Version]
- Oliva, J.; Florentino, A.; Bardag-Gorce, F.; Niihara, Y. Engineering, differentiation and harvesting of human adipose-derived stem cell multilayer cell sheets. Regen. Med. 2019, 14, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Simonacci, F.; Bertozzi, N.; Grieco, M.P.; Raposio, E. From liposuction to adipose-derived stem cells: Indications and technique. Acta Biomed. 2019, 90, 197–208. [Google Scholar]
- Palumbo, P.; Lombardi, F.; Siragusa, G.; Cifone, M.G.; Cinque, B.; Giuliani, M. Methods of Isolation, Characterization and Expansion of Human Adipose-Derived Stem Cells (ASCs): An Overview. Int. J. Mol. Sci. 2018, 19, 1897. [Google Scholar] [CrossRef] [Green Version]
- Lockhart, R.A.; Aronowitz, J.A.; Dos-Anjos Vilaboa, S. Use of Freshly Isolated Human Adipose Stromal Cells for Clinical Applications. Aesthet. Surg. J. 2017, 37, S4–S8. [Google Scholar] [CrossRef] [Green Version]
- Caruso, S.R.; Orellana, M.D.; Mizukami, A.; Fernandes, T.R.; Fontes, A.M.; Suazo, C.A.; Oliveira, V.C.; Covas, D.T.; Swiech, K. Growth and functional harvesting of human mesenchymal stromal cells cultured on a microcarrier-based system. Biotechnol. Prog. 2014, 30, 889–895. [Google Scholar] [CrossRef]
- Nekanti, U.; Mohanty, L.; Venugopal, P.; Balasubramanian, S.; Totey, S.; Ta, M. Optimization and scale-up of Wharton’s jelly-derived mesenchymal stem cells for clinical applications. Stem Cell Res. 2010, 5, 244–254. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Yamato, M.; Mitani, G.; Takagaki, T.; Hamahashi, K.; Nakamura, Y.; Ishihara, M.; Matoba, R.; Kobayashi, H.; Okano, T.; et al. Combined surgery and chondrocyte cell-sheet transplantation improves clinical and structural outcomes in knee osteoarthritis. NPJ Regen. Med. 2019, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Miyagawa, S.; Domae, K.; Yoshikawa, Y.; Fukushima, S.; Nakamura, T.; Saito, A.; Sakata, Y.; Hamada, S.; Toda, K.; Pak, K.; et al. Phase I Clinical Trial of Autologous Stem Cell-Sheet Transplantation Therapy for Treating Cardiomyopathy. J. Am. Heart Assoc. 2017, 6, e003918. [Google Scholar] [CrossRef]
- Devito, L.; Petrova, A.; Miere, C.; Codognotto, S.; Blakely, N.; Lovatt, A.; Ogilvie, C.; Khalaf, Y.; Ilic, D. Cost-effective master cell bank validation of multiple clinical-grade human pluripotent stem cell lines from a single donor. Stem Cells Transl. Med. 2014, 3, 1116–1124. [Google Scholar] [CrossRef]
- Shan, X.; Hu, D. Bone engineering by cell sheet technology to repair mandibular defects. Exp. Ther. Med. 2017, 14, 5007–5011. [Google Scholar] [CrossRef] [Green Version]
- Assuncao, M.; Dehghan-Baniani, D.; Yiu, C.H.K.; Spater, T.; Beyer, S.; Blocki, A. Cell-Derived Extracellular Matrix for Tissue Engineering and Regenerative Medicine. Front. Bioeng. Biotechnol. 2020, 8, 602009. [Google Scholar] [CrossRef]
- Elloumi-Hannachi, I.; Yamato, M.; Okano, T. Cell sheet engineering: A unique nanotechnology for scaffold-free tissue reconstruction with clinical applications in regenerative medicine. J. Intern. Med. 2010, 267, 54–70. [Google Scholar] [CrossRef]
- Nakao, M.; Kim, K.; Nagase, K.; Grainger, D.W.; Kanazawa, H.; Okano, T. Phenotypic traits of mesenchymal stem cell sheets fabricated by temperature-responsive cell culture plate: Structural characteristics of MSC sheets. Stem Cell Res. Ther. 2019, 10, 353. [Google Scholar] [CrossRef]
- Kim, S.R.; Yi, H.J.; Lee, Y.N.; Park, J.Y.; Hoffman, R.M.; Okano, T.; Shim, I.K.; Kim, S.C. Engineered mesenchymal stem-cell-sheets patches prevents postoperative pancreatic leakage in a rat model. Sci. Rep. 2018, 8, 360. [Google Scholar] [CrossRef] [Green Version]
- Doogue, M.P.; Polasek, T.M. The ABCD of clinical pharmacokinetics. Ther. Adv. Drug Saf. 2013, 4, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Lauridsen, H.; Foldager, C.B.; Hansen, L.; Pedersen, M. Non-invasive cell tracking of SPIO labeled cells in an intrinsic regenerative environment: The axolotl limb. Exp. Ther. Med. 2018, 15, 3311–3319. [Google Scholar] [CrossRef]
- Wu, L.; Liu, F.; Liu, S.; Xu, X.; Liu, Z.; Sun, X. Perfluorocarbons-Based (19)F Magnetic Resonance Imaging in Biomedicine. Int. J. Nanomed. 2020, 15, 7377–7395. [Google Scholar] [CrossRef] [PubMed]
- Oliva, J.; Bardag-Gorce, F.; Wood, A.; Sota, H.; Niihara, Y. Direct labeling of 19F-perfluorocarbon onto multilayered cell sheet for MRI-based non-invasive cell tracking. Tissue Eng. Regen. Med. 2015, 12, 371–378. [Google Scholar] [CrossRef]
- Oliva, J.; Bardag-Gorce, F.; Niihara, Y. Clinical Trials of Limbal Stem Cell Deficiency Treated with Oral Mucosal Epithelial Cells. Int. J. Mol. Sci. 2020, 21, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, T.; Sotozono, C.; Bentley, A.J.; Mano, S.; Inatomi, T.; Koizumi, N.; Fullwood, N.J.; Kinoshita, S. Long-term phenotypic study after allogeneic cultivated corneal limbal epithelial transplantation for severe ocular surface diseases. Ophthalmology 2010, 117, 2247–2254.e1. [Google Scholar] [CrossRef]
- Meek, S.; Wei, J.; Oh, T.; Watson, T.; Olavarrieta, J.; Sutherland, L.; Carlson, D.F.; Salzano, A.; Chandra, T.; Joshi, A.; et al. A Stem Cell Reporter for Investigating Pluripotency and Self-Renewal in the Rat. Stem Cell Rep. 2020, 14, 154–166. [Google Scholar] [CrossRef] [Green Version]
- Jurgielewicz, P.; Harmsen, S.; Wei, E.; Bachmann, M.H.; Ting, R.; Aras, O. New imaging probes to track cell fate: Reporter genes in stem cell research. Cell Mol. Life Sci. 2017, 74, 4455–4469. [Google Scholar] [CrossRef]
- Chalfie, M. GFP: Lighting up life. Proc. Natl. Acad. Sci. USA 2009, 106, 10073–10080. [Google Scholar] [CrossRef]
- Soghomonyan, S.; Hajitou, A.; Rangel, R.; Trepel, M.; Pasqualini, R.; Arap, W.; Gelovani, J.G.; Alauddin, M.M. Molecular PET imaging of HSV1-tk reporter gene expression using [18F]FEAU. Nat. Protoc. 2007, 2, 416–423. [Google Scholar] [CrossRef] [Green Version]
- Iwasawa, C.; Tamura, R.; Sugiura, Y.; Suzuki, S.; Kuzumaki, N.; Narita, M.; Suematsu, M.; Nakamura, M.; Yoshida, K.; Toda, M.; et al. Increased Cytotoxicity of Herpes Simplex Virus Thymidine Kinase Expression in Human Induced Pluripotent Stem Cells. Int. J. Mol. Sci. 2019, 20, 810. [Google Scholar] [CrossRef] [Green Version]
- Deans, A.E.; Wadghiri, Y.Z.; Bernas, L.M.; Yu, X.; Rutt, B.K.; Turnbull, D.H. Cellular MRI contrast via coexpression of transferrin receptor and ferritin. Magn. Reson. Med. 2006, 56, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Brissot, P.; Troadec, M.B.; Loreal, O.; Brissot, E. Pathophysiology and classification of iron overload diseases; update 2018. Transfus. Clin. Biol. 2019, 26, 80–88. [Google Scholar] [CrossRef]
- Gherase, M.R.; Fleming, D.E.B. Probing Trace Elements in Human Tissues with Synchrotron Radiation. Crystals 2020, 10, 12. [Google Scholar] [CrossRef] [Green Version]
- Endo, M.; Ozawa, T. Advanced Bioluminescence System for In Vivo Imaging with Brighter and Red-Shifted Light Emission. Int. J. Mol. Sci. 2020, 21, 6538. [Google Scholar] [CrossRef]
- Perrin, J.; Capitao, M.; Mougin-Degraef, M.; Guerard, F.; Faivre-Chauvet, A.; Rbah-Vidal, L.; Gaschet, J.; Guilloux, Y.; Kraeber-Bodere, F.; Cherel, M.; et al. Cell Tracking in Cancer Immunotherapy. Front. Med. 2020, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Jokerst, J.V. Stem Cell Tracking with Nanoparticle-Based Ultrasound Contrast Agents. Methods Mol. Biol. 2020, 2126, 141–153. [Google Scholar]
- Kubelick, K.P.; Snider, E.J.; Ethier, C.R.; Emelianov, S. Development of a stem cell tracking platform for ophthalmic applications using ultrasound and photoacoustic imaging. Theranostics 2019, 9, 3812–3824. [Google Scholar] [CrossRef]
- Vandergaast, R.; Khongwichit, S.; Jiang, H.; DeGrado, T.R.; Peng, K.W.; Smith, D.R.; Russell, S.J.; Suksanpaisan, L. Enhanced noninvasive imaging of oncology models using the NIS reporter gene and bioluminescence imaging. Cancer Gene Ther. 2020, 27, 179–188. [Google Scholar] [CrossRef] [Green Version]
- Sulzer, D.; Cassidy, C.; Horga, G.; Kang, U.J.; Fahn, S.; Casella, L.; Pezzoli, G.; Langley, J.; Hu, X.P.; Zucca, F.A.; et al. Neuromelanin detection by magnetic resonance imaging (MRI) and its promise as a biomarker for Parkinson’s disease. NPJ Parkinsons Dis. 2018, 4, 11. [Google Scholar] [CrossRef]
- MacAskill, M.G.; Tavares, A.S.; Wu, J.; Lucatelli, C.; Mountford, J.C.; Baker, A.H.; Newby, D.E.; Hadoke, P.W. PET Cell Tracking Using (18)F-FLT is Not Limited by Local Reuptake of Free Radiotracer. Sci. Rep. 2017, 7, 44233. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.W.; Choi, S.I.; Lee, S.J.; Oh, Y.T.; Park, G.; Park, N.Y.; Yoon, K.A.; Kim, S.; Kim, D.; Kim, Y.H.; et al. The Effectiveness of Ferritin as a Contrast Agent for Cell Tracking MRI in Mouse Cancer Models. Yonsei Med. J. 2017, 58, 51–58. [Google Scholar] [CrossRef]
- Ansari, A.M.; Ahmed, A.K.; Matsangos, A.E.; Lay, F.; Born, L.J.; Marti, G.; Harmon, J.W.; Sun, Z. Cellular GFP Toxicity and Immunogenicity: Potential Confounders in in Vivo Cell Tracking Experiments. Stem Cell Rev. Rep. 2016, 12, 553–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crawford, A.M.; Kurecka, P.; Yim, T.K.; Kozemchak, C.; Deb, A.; Dostal, L.; Sun, C.J.; Brewe, D.L.; Barrea, R.; Penner-Hahn, J.E. Development of a single-cell X-ray fluorescence flow cytometer. J. Synchrotron Radiat. 2016, 23 Pt 4, 901–908. [Google Scholar] [CrossRef]
- Wang, Z.G.; Liu, S.L.; Hu, Y.J.; Tian, Z.Q.; Hu, B.; Zhang, Z.L.; Pang, D.W. Dissecting the Factors Affecting the Fluorescence Stability of Quantum Dots in Live Cells. ACS Appl. Mater. Interfaces 2016, 8, 8401–8408. [Google Scholar] [CrossRef] [PubMed]
- Suetsugu, A.; Jiang, P.; Yang, M.; Yamamoto, N.; Moriwaki, H.; Saji, S.; Hoffman, R.M. The Use of Living Cancer Cells Expressing Green Fluorescent Protein in the Nucleus and Red Fluorescence Protein in the Cytoplasm for Real-time Confocal Imaging of Chromosome and Cytoplasmic Dynamics During Mitosis. Anticancer Res. 2015, 35, 2553–2557. [Google Scholar] [PubMed]
- Kim, J.E.; Kalimuthu, S.; Ahn, B.C. In vivo cell tracking with bioluminescence imaging. Nucl. Med. Mol. Imaging 2015, 49, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pushie, M.J.; Pickering, I.J.; Korbas, M.; Hackett, M.J.; George, G.N. Elemental and chemically specific X-ray fluorescence imaging of biological systems. Chem. Rev. 2014, 114, 8499–8541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penheiter, A.R.; Russell, S.J.; Carlson, S.K. The sodium iodide symporter (NIS) as an imaging reporter for gene, viral, and cell-based therapies. Curr. Gene Ther. 2012, 12, 33–47. [Google Scholar] [CrossRef]
- Gildehaus, F.J.; Haasters, F.; Drosse, I.; Wagner, E.; Zach, C.; Mutschler, W.; Cumming, P.; Bartenstein, P.; Schieker, M. Impact of indium-111 oxine labelling on viability of human mesenchymal stem cells in vitro, and 3D cell-tracking using SPECT/CT in vivo. Mol. Imaging Biol. 2011, 13, 1204–1214. [Google Scholar] [CrossRef]
- Walling, M.A.; Novak, J.A.; Shepard, J.R. Quantum dots for live cell and in vivo imaging. Int. J. Mol. Sci. 2009, 10, 441–491. [Google Scholar] [CrossRef] [Green Version]
- Kummer, C.; Winkeler, A.; Dittmar, C.; Bauer, B.; Rueger, M.A.; Rueckriem, B.; Heneka, M.T.; Vollmar, S.; Wienhard, K.; Fraefel, C.; et al. Multitracer positron emission tomographic imaging of exogenous gene expression mediated by a universal herpes simplex virus 1 amplicon vector. Mol. Imaging 2007, 6, 181–192. [Google Scholar] [CrossRef]
- Hackman, T.; Doubrovin, M.; Balatoni, J.; Beresten, T.; Ponomarev, V.; Beattie, B.; Finn, R.; Bornmann, W.; Blasberg, R.; Tjuvajev, J.G. Imaging expression of cytosine deaminase-herpes virus thymidine kinase fusion gene (CD/TK) expression with [124I]FIAU and PET. Mol. Imaging 2002, 1, 36–42. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, R.C. The role of chemokines in mesenchymal stem cell homing to myocardium. Stem Cell Rev. Rep. 2012, 8, 243–250. [Google Scholar] [CrossRef]
- Ponte, A.L.; Marais, E.; Gallay, N.; Langonne, A.; Delorme, B.; Herault, O.; Charbord, P.; Domenech, J. The in vitro migration capacity of human bone marrow mesenchymal stem cells: Comparison of chemokine and growth factor chemotactic activities. Stem Cells 2007, 25, 1737–1745. [Google Scholar] [CrossRef]
- Sordi, V.; Malosio, M.L.; Marchesi, F.; Mercalli, A.; Melzi, R.; Giordano, T.; Belmonte, N.; Ferrari, G.; Leone, B.E.; Bertuzzi, F.; et al. Bone marrow mesenchymal stem cells express a restricted set of functionally active chemokine receptors capable of promoting migration to pancreatic islets. Blood 2005, 106, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Hocking, A.M. The Role of Chemokines in Mesenchymal Stem Cell Homing to Wounds. Adv. Wound Care 2015, 4, 623–630. [Google Scholar] [CrossRef]
- Lo, C.Y.; Antonopoulos, A.; Dell, A.; Haslam, S.M.; Lee, T.; Neelamegham, S. The use of surface immobilization of P-selectin glycoprotein ligand-1 on mesenchymal stem cells to facilitate selectin mediated cell tethering and rolling. Biomaterials 2013, 34, 8213–8222. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.; Zhao, X.; Zhang, L.; Zhang, J.; L’Huillier, A.; Ling, W.; Roberts, A.I.; Le, A.D.; Shi, S.; Shao, C.; et al. Inflammatory cytokine-induced intercellular adhesion molecule-1 and vascular cell adhesion molecule-1 in mesenchymal stem cells are critical for immunosuppression. J. Immunol. 2010, 184, 2321–2328. [Google Scholar] [CrossRef]
- Tang, B.; Li, X.; Liu, Y.; Chen, X.; Li, X.; Chu, Y.; Zhu, H.; Liu, W.; Xu, F.; Zhou, F.; et al. The Therapeutic Effect of ICAM-1-Overexpressing Mesenchymal Stem Cells on Acute Graft-Versus-Host Disease. Cell Physiol. Biochem. 2018, 46, 2624–2635. [Google Scholar] [CrossRef] [Green Version]
- Masterson, C.H.; Curley, G.F.; Laffey, J.G. Modulating the distribution and fate of exogenously delivered MSCs to enhance therapeutic potential: Knowns and unknowns. Intensive Care Med. Exp. 2019, 7, 41. [Google Scholar] [CrossRef]
- Nystedt, J.; Anderson, H.; Tikkanen, J.; Pietila, M.; Hirvonen, T.; Takalo, R.; Heiskanen, A.; Satomaa, T.; Natunen, S.; Lehtonen, S.; et al. Cell surface structures influence lung clearance rate of systemically infused mesenchymal stromal cells. Stem Cells 2013, 31, 317–326. [Google Scholar] [CrossRef]
- Valles, G.; Bensiamar, F.; Maestro-Paramio, L.; Garcia-Rey, E.; Vilaboa, N.; Saldana, L. Influence of inflammatory conditions provided by macrophages on osteogenic ability of mesenchymal stem cells. Stem Cell Res. Ther. 2020, 11, 57. [Google Scholar] [CrossRef] [PubMed]
- Newman, R.E.; Yoo, D.; LeRoux, M.A.; Danilkovitch-Miagkova, A. Treatment of inflammatory diseases with mesenchymal stem cells. Inflamm. Allergy Drug Targets 2009, 8, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Klinker, M.W.; Wei, C.H. Mesenchymal stem cells in the treatment of inflammatory and autoimmune diseases in experimental animal models. World J. Stem Cells 2015, 7, 556–567. [Google Scholar] [CrossRef]
- Monguio-Tortajada, M.; Bayes-Genis, A.; Rosell, A.; Roura, S. Are mesenchymal stem cells and derived extracellular vesicles valuable to halt the COVID-19 inflammatory cascade? Current evidence and future perspectives. Thorax 2020, 76, 196–200. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.J.; Harman, R.J.; Bunnell, B.A.; Schreiber, M.A.; Xiang, C.; Wang, F.S.; Santidrian, A.F.; Minev, B.R. Rationale for the clinical use of adipose-derived mesenchymal stem cells for COVID-19 patients. J. Transl. Med. 2020, 18, 203. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.S.; Khan, M.A. Mesenchymal Stem Cells: A new front emerge in COVID19 treatment: Mesenchymal Stem Cells therapy for SARS-CoV2 viral infection. Cytotherapy 2020. [Google Scholar] [CrossRef]
- Hos, D.; Saban, D.R.; Bock, F.; Regenfuss, B.; Onderka, J.; Masli, S.; Cursiefen, C. Suppression of inflammatory corneal lymphangiogenesis by application of topical corticosteroids. Arch. Ophthalmol. 2011, 129, 445–452. [Google Scholar] [CrossRef]
- Drela, K.; Stanaszek, L.; Nowakowski, A.; Kuczynska, Z.; Lukomska, B. Experimental Strategies of Mesenchymal Stem Cell Propagation: Adverse Events and Potential Risk of Functional Changes. Stem Cells Int. 2019, 2019, 7012692. [Google Scholar] [CrossRef] [Green Version]
- Sotiropoulou, P.A.; Perez, S.A.; Salagianni, M.; Baxevanis, C.N.; Papamichail, M. Characterization of the optimal culture conditions for clinical scale production of human mesenchymal stem cells. Stem Cells 2006, 24, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Dominguez, X.; Vera-Donoso, C.D.; Jimenez-Trigos, E.; Vicente, J.S.; Marco-Jimenez, F. First Steps Towards Organ Banks: Vitrification of Renal Primordial. CryoLetters 2016, 37, 47–52. [Google Scholar]
- Mahla, R.S. Stem Cells Applications in Regenerative Medicine and Disease Therapeutics. Int. J. Cell Biol. 2016, 2016, 6940283. [Google Scholar] [CrossRef] [Green Version]
- Costa, P.F.; Dias, A.F.; Reis, R.L.; Gomes, M.E. Cryopreservation of cell/scaffold tissue-engineered constructs. Tissue Eng. Part C Methods 2012, 18, 852–858. [Google Scholar] [CrossRef]
- Rahmati, M.; Pennisi, C.P.; Mobasheri, A.; Mozafari, M. Bioengineered Scaffolds for Stem Cell Applications in Tissue Engineering and Regenerative Medicine. Adv. Exp. Med. Biol. 2018, 1107, 73–89. [Google Scholar]
- Agmon, G.; Christman, K.L. Controlling stem cell behavior with decellularized extracellular matrix scaffolds. Curr. Opin. Solid State Mater. Sci. 2016, 20, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, J.; Kikuchi, A.; Aoyagi, T.; Okano, T. Cell sheet tissue engineering: Cell sheet preparation, harvesting/manipulation, and transplantation. J. Biomed. Mater. Res. A 2019, 107, 955–967. [Google Scholar] [CrossRef]
- Makar, R.S.; Toth, T.L. The evaluation of infertility. Am. J. Clin. Pathol. 2002, 117 (Suppl. 1), S95–S103. [Google Scholar] [CrossRef]
- Jang, T.H.; Park, S.C.; Yang, J.H.; Kim, J.Y.; Seok, J.H.; Park, U.S.; Choi, C.W.; Lee, S.R.; Han, J. Cryopreservation and its clinical applications. Integr. Med. Res. 2017, 6, 12–18. [Google Scholar] [CrossRef]
- Vutyavanich, T.; Lattiwongsakorn, W.; Piromlertamorn, W.; Samchimchom, S. Repeated vitrification/warming of human sperm gives better results than repeated slow programmable freezing. Asian J. Androl. 2012, 14, 850–854. [Google Scholar] [CrossRef] [Green Version]
- Best, B.P. Cryoprotectant Toxicity: Facts, Issues, and Questions. Rejuvenation Res. 2015, 18, 422–436. [Google Scholar] [CrossRef] [Green Version]
- Asghar, W.; El Assal, R.; Shafiee, H.; Anchan, R.M.; Demirci, U. Preserving human cells for regenerative, reproductive, and transfusion medicine. Biotechnol. J. 2014, 9, 895–903. [Google Scholar] [CrossRef] [Green Version]
- De Vries, R.J.; Yarmush, M.; Uygun, K. Systems engineering the organ preservation process for transplantation. Curr. Opin. Biotechnol. 2019, 58, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Israni, A.K.; Zaun, D.; Rosendale, J.D.; Snyder, J.J.; Kasiske, B.L. OPTN/SRTR 2012 Annual Data Report: Deceased organ donation. Am. J. Transplant. 2014, 14 (Suppl. 1), 167–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abouna, G.M. Organ shortage crisis: Problems and possible solutions. Transplant. Proc. 2008, 40, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Theruvath, A.J.; Ge, X.; Floerchinger, B.; Jurisch, A.; Garcia-Cardena, G.; Tullius, S.G. Machine perfusion or cold storage in organ transplantation: Indication, mechanisms, and future perspectives. Transpl. Int. 2010, 23, 561–570. [Google Scholar] [CrossRef] [PubMed]
- Mantecchini, L.; Paganelli, F.; Morabito, V.; Ricci, A.; Peritore, D.; Trapani, S.; Montemurro, A.; Rizzo, A.; Del Sordo, E.; Gaeta, A.; et al. Transportation of Organs by Air: Safety, Quality, and Sustainability Criteria. Transplant. Proc. 2016, 48, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Karow, A.M., Jr.; Webb, W.R. Cardiac storage with glycerol at zero centigrade. Arch. Surg. 1961, 83, 719–720. [Google Scholar] [CrossRef] [PubMed]
- Van den Broecke, R.; Liu, J.; Van der Elst, J.; Dhont, M. Timing of FSH-stimulation and follicular development in cryopreserved human ovarian grafts. Reprod. Biomed. Online 2002, 4, 21–26. [Google Scholar] [CrossRef]
- Carletti, E.; Motta, A.; Migliaresi, C. Scaffolds for tissue engineering and 3D cell culture. Methods Mol. Biol. 2011, 695, 17–39. [Google Scholar]
- Haraguchi, Y.; Shimizu, T.; Yamato, M.; Okano, T. Regenerative therapies using cell sheet-based tissue engineering for cardiac disease. Cardiol. Res. Pract. 2011, 2011, 845170. [Google Scholar] [CrossRef] [Green Version]
- Anassi, E.; Ndefo, U.A. Sipuleucel-T (provenge) injection: The first immunotherapy agent (vaccine) for hormone-refractory prostate cancer. Pharm. Ther. 2011, 36, 197–202. [Google Scholar]
- Hayashi, A.; Maehara, M.; Uchikura, A.; Matsunari, H.; Matsumura, K.; Hyon, S.H.; Sato, M.; Nagashima, H. Development of an efficient vitrification method for chondrocyte sheets for clinical application. Regen Ther. 2020, 14, 215–221. [Google Scholar] [CrossRef]
- Ohkawara, H.; Miyagawa, S.; Fukushima, S.; Yajima, S.; Saito, A.; Nagashima, H.; Sawa, Y. Development of a vitrification method for preserving human myoblast cell sheets for myocardial regeneration therapy. BMC Biotechnol. 2018, 18, 56. [Google Scholar] [CrossRef] [Green Version]
- Tani, Y.; Sato, M.; Maehara, M.; Nagashima, H.; Yokoyama, M.; Yokoyama, M.; Yamato, M.; Okano, T.; Mochida, J. The effects of using vitrified chondrocyte sheets on pain alleviation and articular cartilage repair. J. Tissue Eng. Regen. Med. 2017, 11, 3437–3444. [Google Scholar] [CrossRef]
- Maehara, M.; Sato, M.; Watanabe, M.; Matsunari, H.; Kokubo, M.; Kanai, T.; Sato, M.; Matsumura, K.; Hyon, S.H.; Yokoyama, M.; et al. Development of a novel vitrification method for chondrocyte sheets. BMC Biotechnol. 2013, 13, 58. [Google Scholar] [CrossRef] [Green Version]
- Oie, Y.; Nozaki, T.; Takayanagi, H.; Hara, S.; Hayashi, R.; Takeda, S.; Mori, K.; Moriya, N.; Soma, T.; Tsujikawa, M.; et al. Development of a cell sheet transportation technique for regenerative medicine. Tissue Eng. Part C Methods 2014, 20, 373–382. [Google Scholar] [CrossRef] [Green Version]
- Larman, M.G.; Katz-Jaffe, M.G.; McCallie, B.; Filipovits, J.A.; Gardner, D.K. Analysis of global gene expression following mouse blastocyst cryopreservation. Hum. Reprod. 2011, 26, 2672–2680. [Google Scholar] [CrossRef] [Green Version]
- Brair, V.L.; Maia, A.; Correia, L.F.L.; Barbosa, N.O.; Santos, J.D.R.; Brandao, F.Z.; Fonseca, J.F.; Batista, R.; Souza-Fabjan, J.M.G. Gene expression patterns of in vivo-derived sheep blastocysts is more affected by vitrification than slow freezing technique. Cryobiology 2020, 95, 110–115. [Google Scholar] [CrossRef]
- Bera, M.; Sengupta, K. Nuclear filaments: Role in chromosomal positioning and gene expression. Nucleus 2020, 11, 99–110. [Google Scholar] [CrossRef]
- Ugur, M.R.; Saber Abdelrahman, A.; Evans, H.C.; Gilmore, A.A.; Hitit, M.; Arifiantini, R.I.; Purwantara, B.; Kaya, A.; Memili, E. Advances in Cryopreservation of Bull Sperm. Front. Vet. Sci. 2019, 6, 268. [Google Scholar] [CrossRef] [Green Version]
- Golan, M.; Pribyl, J.; Pesl, M.; Jelinkova, S.; Acimovic, I.; Jaros, J.; Rotrekl, V.; Falk, M.; Sefc, L.; Skladal, P.; et al. Cryopreserved Cells Regeneration Monitored by Atomic Force Microscopy and Correlated With State of Cytoskeleton and Nuclear Membrane. IEEE Trans. Nanobiosci. 2018, 17, 485–497. [Google Scholar] [CrossRef]
- Aurich, C.; Schreiner, B.; Ille, N.; Alvarenga, M.; Scarlet, D. Cytosine methylation of sperm DNA in horse semen after cryopreservation. Theriogenology 2016, 86, 1347–1352. [Google Scholar] [CrossRef] [Green Version]
- Kopeika, J.; Thornhill, A.; Khalaf, Y. The effect of cryopreservation on the genome of gametes and embryos: Principles of cryobiology and critical appraisal of the evidence. Hum. Reprod. Update 2015, 21, 209–227. [Google Scholar] [CrossRef] [Green Version]
- Said, T.M.; Gaglani, A.; Agarwal, A. Implication of apoptosis in sperm cryoinjury. Reprod. Biomed. Online 2010, 21, 456–462. [Google Scholar] [CrossRef] [Green Version]
- Waris, G.; Ahsan, H. Reactive oxygen species: Role in the development of cancer and various chronic conditions. J. Carcinog. 2006, 5, 14. [Google Scholar] [CrossRef]
- De Leeuw, F.E.; Chen, H.C.; Colenbrander, B.; Verkleij, A.J. Cold-induced ultrastructural changes in bull and boar sperm plasma membranes. Cryobiology 1990, 27, 171–183. [Google Scholar] [CrossRef]
- Tao, Y.; Sanger, E.; Saewu, A.; Leveille, M.C. Human sperm vitrification: The state of the art. Reprod. Biol. Endocrinol. 2020, 18, 17. [Google Scholar] [CrossRef]
- Kulis, M.; Esteller, M. DNA methylation and cancer. Adv. Genet. 2010, 70, 27–56. [Google Scholar]
- Bartolac, L.K.; Lowe, J.L.; Koustas, G.; Grupen, C.G.; Sjoblom, C. Effect of different penetrating and non-penetrating cryoprotectants and media temperature on the cryosurvival of vitrified in vitro produced porcine blastocysts. Anim. Sci. J. 2018, 89, 1230–1239. [Google Scholar] [CrossRef]
- Kummerfeld, D.M.; Raabe, C.A.; Brosius, J.; Mo, D.; Skryabin, B.V.; Rozhdestvensky, T.S. A Comprehensive Review of Genetically Engineered Mouse Models for Prader-Willi Syndrome Research. Int. J. Mol. Sci. 2021, 22, 3613. [Google Scholar] [CrossRef] [PubMed]
- Madaan, M.; Mendez, M.D. Angelman Syndrome. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Wang, K.H.; Kupa, J.; Duffy, K.A.; Kalish, J.M. Diagnosis and Management of Beckwith-Wiedemann Syndrome. Front. Pediatr. 2019, 7, 562. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, A.; Adamek, A. Insulin-Like Growth Factor 2 (IGF2) Signaling in Colorectal Cancer-From Basic Research to Potential Clinical Applications. Int. J. Mol. Sci. 2019, 20, 4915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anvar, Z.; Acurzio, B.; Roma, J.; Cerrato, F.; Verde, G. Origins of DNA methylation defects in Wilms tumors. Cancer Lett. 2019, 457, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Oliva, J.; Florentino, A.; Bardag-Gorce, F.; Niihara, Y. Vitrification and storage of oral mucosa epithelial cell sheets. J. Tissue Eng. Regen. Med. 2019, 13, 1153–1163. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Bian, Y.L.; Schreurs, N.; Zhang, X.G.; Raza, S.H.A.; Fang, Q.; Wang, L.Q.; Hu, J.H. Effects of five cryoprotectants on proliferation and differentiation-related gene expression of frozen-thawed bovine calf testicular tissue. Reprod. Domest. Anim. 2018, 53, 1211–1218. [Google Scholar] [CrossRef]
- Jahan, S.; Kaushal, R.; Pasha, R.; Pineault, N. Current and Future Perspectives for the Cryopreservation of Cord Blood Stem Cells. Transfus. Med. Rev. 2021. [Google Scholar] [CrossRef]
- Fahy, G.M.; Wowk, B. Principles of Ice-Free Cryopreservation by Vitrification. Methods Mol. Biol. 2021, 2180, 27–97. [Google Scholar]
- Elliott, G.D.; Wang, S.; Fuller, B.J. Cryoprotectants: A review of the actions and applications of cryoprotective solutes that modulate cell recovery from ultra-low temperatures. Cryobiology 2017, 76, 74–91. [Google Scholar] [CrossRef]
- Storey, K.B.; Storey, J.M. Molecular Physiology of Freeze Tolerance in Vertebrates. Physiol. Rev. 2017, 97, 623–665. [Google Scholar] [CrossRef]
- Yang, J.; Gao, L.; Liu, M.; Sui, X.; Zhu, Y.; Wen, C.; Zhang, L. Advanced Biotechnology for Cell Cryopreservation. Trans. Tianjin Univ. 2020, 26, 409–423. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Kong, M.J.; Jang, H.J.; Cho, J.I.; Park, E.J.; Lee, I.K.; Frokiaer, J.; Norregaard, R.; Park, K.M.; Kwon, T.H. A nonbiodegradable scaffold-free cell sheet of genome-engineered mesenchymal stem cells inhibits development of acute kidney injury. Kidney Int. 2021, 99, 117–133. [Google Scholar] [CrossRef]
- Galvez, V.; Chacon-Solano, E.; Bonafont, J.; Mencia, A.; Di, W.L.; Murillas, R.; Llames, S.; Vicente, A.; Del Rio, M.; Carretero, M.; et al. Efficient CRISPR-Cas9-Mediated Gene Ablation in Human Keratinocytes to Recapitulate Genodermatoses: Modeling of Netherton Syndrome. Mol. Ther. Methods Clin. Dev. 2020, 18, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, R.; Zhang, Q.; Xu, Z.; Xu, F.; Li, D.; Li, Y. Chondrocyte sheet in vivo cartilage regeneration technique using miR-193b-3p to target MMP16. Aging 2019, 11, 7070–7082. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Whalley, R.D.; Ferreira, A.M.; Dalgano, K. High throughput physiological micro-models for in vitro pre-clinical drug testing: A review of engineering systems approaches. Prog. Biomed. Eng. 2020, 2, 22001. [Google Scholar] [CrossRef]
- Asadi, M.; Lotfi, H.; Salehi, R.; Mehdipour, A.; Zarghami, N.; Akbarzadeh, A.; Alizadeh, E. Hepatic cell-sheet fabrication of differentiated mesenchymal stem cells using decellularized extracellular matrix and thermoresponsive polymer. Biomed. Pharmacother. 2021, 134, 111096. [Google Scholar] [CrossRef]
- Xing, Q.; Yates, K.; Tahtinen, M.; Shearier, E.; Qian, Z.; Zhao, F. Decellularization of fibroblast cell sheets for natural extracellular matrix scaffold preparation. Tissue Eng. Part C Methods 2015, 21, 77–87. [Google Scholar] [CrossRef]
Figure 1.
(Left) Summary of the multiple applications for cell sheets. (Right) Scheme showing potential use of cell sheet. Cell sheet or stacked cell sheets (same or different type of cell sheet) can ship directly to the clinic, or they can be vitrified to build a cryobank of cell sheets, before being used for a patient’s transplantation.
Figure 1.
(Left) Summary of the multiple applications for cell sheets. (Right) Scheme showing potential use of cell sheet. Cell sheet or stacked cell sheets (same or different type of cell sheet) can ship directly to the clinic, or they can be vitrified to build a cryobank of cell sheets, before being used for a patient’s transplantation.
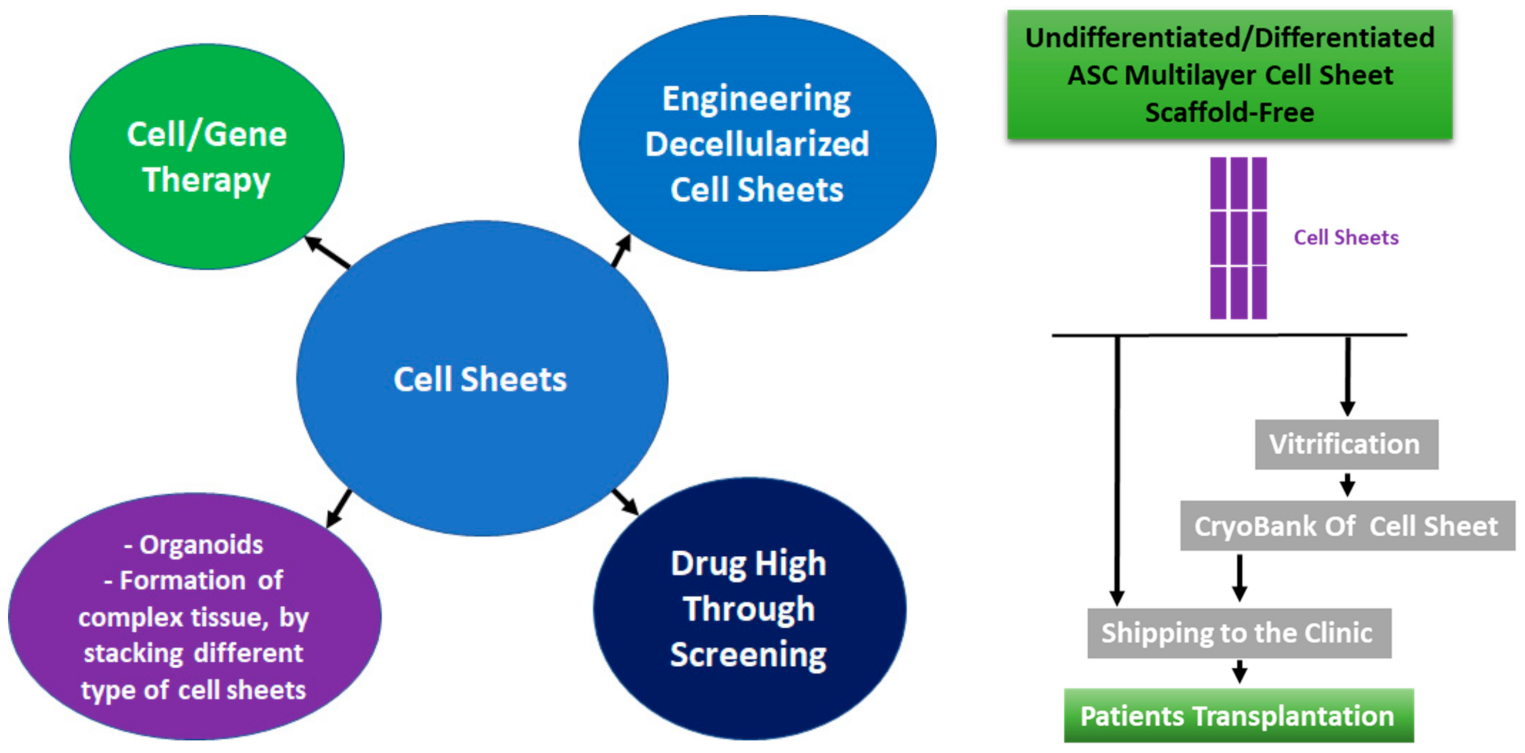

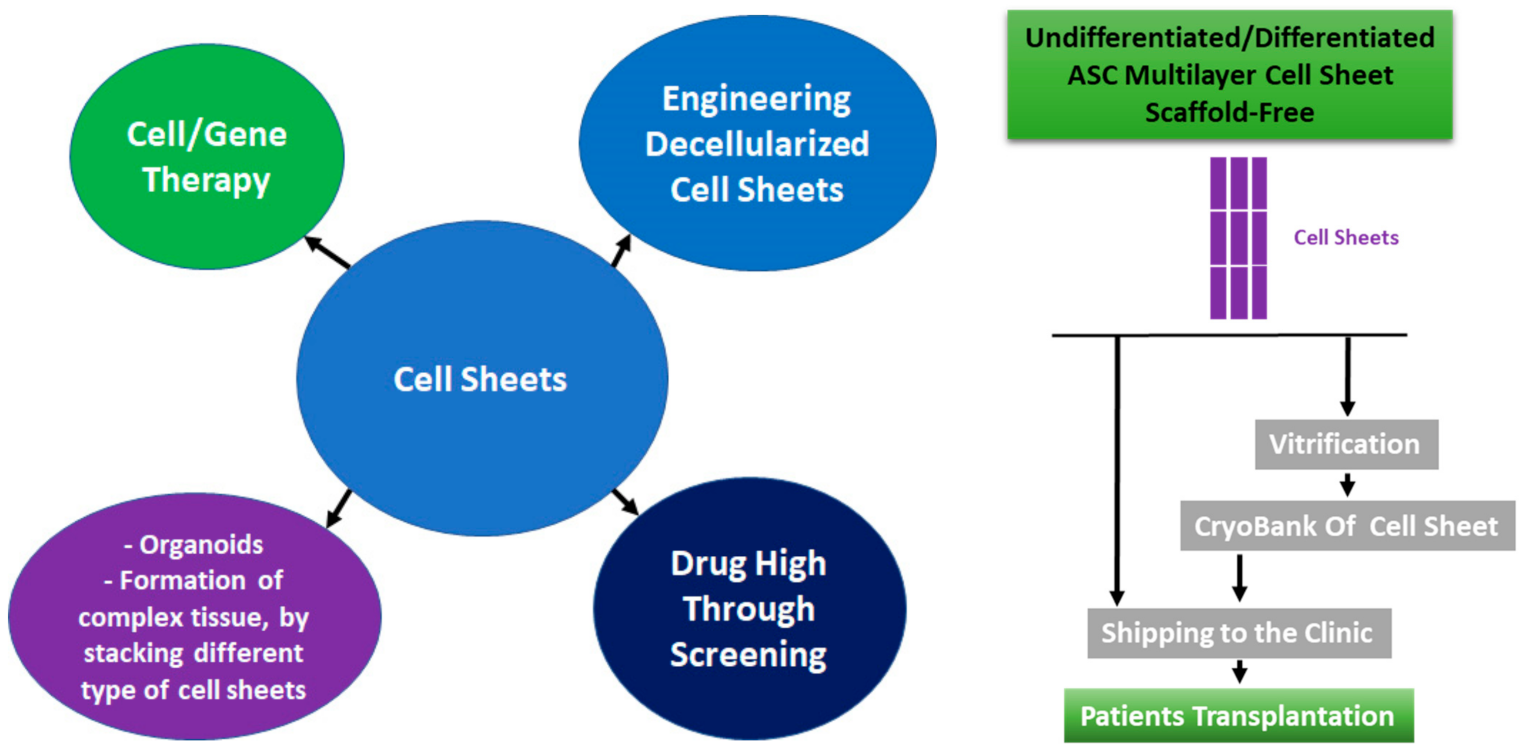{kind=link}
Table 1.
Summary of goals and methodologies for in vivo tracking [110,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141]. SPIO: superparamagnetic iron oxides; 111lnOc: (111)In-oxine; 18F-FDG: [fluorine-18]fluoro-D-glucose; 64Cu-PTSM: 64Cu-pyruvaldehyde-bis(N4-methylthiosemicarbazone); RFP: red fluorescent protein; GFP: green fluorescent protein; Fluc: firefly luciferase; Rluc: renilla luciferase; NIS: sodium iodide symporter; HSV1-TK: herpes simplex virus type 1–thymidine kinase; D2R: Dopamine type 2 receptor.
Table 1.
Summary of goals and methodologies for in vivo tracking [110,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141]. SPIO: superparamagnetic iron oxides; 111lnOc: (111)In-oxine; 18F-FDG: [fluorine-18]fluoro-D-glucose; 64Cu-PTSM: 64Cu-pyruvaldehyde-bis(N4-methylthiosemicarbazone); RFP: red fluorescent protein; GFP: green fluorescent protein; Fluc: firefly luciferase; Rluc: renilla luciferase; NIS: sodium iodide symporter; HSV1-TK: herpes simplex virus type 1–thymidine kinase; D2R: Dopamine type 2 receptor.
| Method | Labelling Method | Detection Depth of the Signal (In Vivo) | Invasive Method | Detection of Living Cells | Safe Methodology For Patients | Animal Size | Data Acquisition Time | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| Direct Cell Visualization | Ultrasound | Incubation | Whole body | No | Can’t be determinated | Yes | Any Animal | Minutes/Hours | [125,126] |
| MR Fluorescent (X-ray Fluoresence) | Incubation | Whole body | No | Can’t be determinated | Yes | Any Animal | Minutes/Hours | [122,132,135] | |
| Direct Cell Labelling | Fluorescence (e.g., quantum dots, fluorophores) | Incubation | 2–3 mm | No | Can’t be determinated | Potentially Toxic | Small Animals | Secondes/Minutes | [133,139] |
| MRI (e.g., SPIO) | Incubation | Whole body | No | Can’t be determinated | Possible Iron Toxicity Harm | Any Animal | Minutes/Hours | [110] | |
| SPECT (e.g., 111InOx) | Incubation | Whole body | No | Can’t be determinated | Potential Radioactive Harm | Any Animal | Minutes | [138] | |
| PET (e.g., 18F-FDG, 64Cu-PTSM) | Incubation | Whole body | No | Can’t be determinated | Potential Radioactive Harm | Any Animal | Minutes | [124,129] | |
| Genetic Modification of the Cells for Cell Detection | Fluorescence (RFP, GFP) | Stable Transfection | Less than 1 cm depth | No | Can’t be determinated | Potential Cell Toxicity | Small animals | Seconds/Minutes | [131,134] |
| Bioluminescence (Fluc, Rluc) | Stable Transfection + Probe injection | Less than 3 cm depth | No | Cells are alive | Yes | Small animals | Seconds/Minutes | [123,135] | |
| SPECT (NIS, somatostatin) | Stable Transfection + Probe injection | Whole body | No | Cells are alive | Potential Radioactive Harm | Any Animal | Minutes | [127,137] | |
| PET (HSV1-tk, D2R) | Stable Transfection + Probe injection | Whole Body | No | Cells are alive | Potential Radioactive Harm and Potential Immunoreactivity with TK | Any Animal | Minutes | [140,141] | |
| MRI (Transferrin, Ferritin, Tyrosine) | Stable Transfection + Probe injection | Whole body | No | Cells are alive | Possible Iron Toxicity Harm | Any Animal | Minutes/hours | [128,130] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ochiai, J.; Niihara, Y.; Oliva, J. Development of Multilayer Mesenchymal Stem Cell Cell Sheets. Int. J. Transl. Med. 2021, 1, 4-24. https://0-doi-org.brum.beds.ac.uk/10.3390/ijtm1010002
AMA Style
Ochiai J, Niihara Y, Oliva J. Development of Multilayer Mesenchymal Stem Cell Cell Sheets. International Journal of Translational Medicine. 2021; 1(1):4-24. https://0-doi-org.brum.beds.ac.uk/10.3390/ijtm1010002
Chicago/Turabian StyleOchiai, Jun, Yutaka Niihara, and Joan Oliva. 2021. "Development of Multilayer Mesenchymal Stem Cell Cell Sheets" International Journal of Translational Medicine 1, no. 1: 4-24. https://0-doi-org.brum.beds.ac.uk/10.3390/ijtm1010002