
SARS–CoV–2 and Food—How Confident Are We about Them?
by
 , , , and
, , , and
Vangelis Economou
1,* ,
,
Hercules Sakkas
2 ,
,
Eugenia Bezirtzoglou
3 ,
,
Anna Papa
4 and
Nikolaos Soultos
1 1
Laboratory of Hygiene of Food of Animal Origin-Veterinary Public Health, School of Veterinary Medicine, Aristotle University of Thessaloniki, 54124 Thessaloniki, Greece
2
Microbiology Department, Faculty of Medicine, School of Health Sciences, University of Ioannina, 45110 Ioannina, Greece
3
Laboratory of Hygiene and Environmental Protection, Medical School, Democritus University of Thrace, 68100 Alexandroupolis, Greece
4
Department of Microbiology, Medical School, Aristotle University of Thessaloniki, 54124 Thessaloniki, Greece
*
Author to whom correspondence should be addressed.
Hygiene 2021, 1(3), 80-98; https://0-doi-org.brum.beds.ac.uk/10.3390/hygiene1030008
Submission received: 6 September 2021
/
Revised: 16 September 2021
/
Accepted: 21 September 2021
/
Published: 29 September 2021
(This article belongs to the Section Food Hygiene and Safety)
Abstract
:The recent COVID-19 pandemic and coronaviruses have been thrust into the lives of humans around the globe. Several concerns of the scientific community, authorities and common people have been aroused concerning the prophylaxis measures that need to be taken in order to safeguard public health. Among others, the possibility of a faecal—oral route, and consequent waterborne or foodborne transmission, have been given little attention. Ground zero was the seafood market of Huanan in Wuhan, China; therefore, it was quite logical at the time to assume a certain degree of relationship between water, seafood and SARS–CoV–2. In this manuscript, a critical review of the current literature concerning these routes of transmission is made. The main questions discussed are whether (i) SARS–CoV–2 can infect food animals, (ii) it can be detected in water, retaining its infectivity for the necessary amount of time, (iii) there is a possibility of contamination of food by SARS–CoV–2 through its various production processes and (iv) there is evidence of foodborne or waterborne transmission.
1. Introduction
The ongoing pandemic coronavirus disease 2019 (CoVID-19) is an infectious disease caused by the severe acute respiratory syndrome coronavirus 2 (SARS–CoV–2). Coronaviruses (CoVs) are notorious zoonotic pathogenic agents that have evolved for thousands of years in different mammals, including man. The origin of the recent pandemic was believed to trace back to a seafood market in Huanan, Wuhan, China [1,2]. Although two more coronavirus (CoV) pandemics, the severe acute respiratory syndrome (SARS) and the Middle East respiratory syndrome (MERS), have taken place over the last 18 years, COVID-19 has brought CoVs back to the forefront due to their high transmissibility and worldwide-spread potential [3,4]. The COVID–19 pandemic has caused a series of containment measures by countries around the world in order to intercept its rapid spread. These measures aimed mostly at the disruption of the chain of airborne transmission, considered to be the most probable transmission pathway.
The pandemic has aroused severe concerns about its human health implications. With regards to emerging pandemics, infections by CoVs are excellent paradigms of this phenomenon, as they were/are responsible for three syndromes: SARS, MERS, and the current COVID–19. The characterization of SARS–CoV–2 as zoonotic is of high importance since the complex dynamics of viruses involving animals, humans and the environment present high epidemic potential. Still, the parental origin of SARS–CoV–2 is highly disputed, since CoVs from different animals have been claimed to have this property. Zhou et al. (2020) have reported the similarity of the virus to SARS–CoV–1 and bat CoVs, implying that bats (Rhinolophus affinis) might be the reservoir host(s) of SARS–CoV–2 [5]. Other researchers have reported that pangolins (Manis javanica) harbor ancestral beta-CoVs closely related to SARS–CoV–2 [3].
It is evident that there are several gaps in the epidemiology of SARS–CoV–2 that need to be elucidated. Therefore, foodborne transmission of SARS–CoV–2 remains to be explored, and several scientists are pointing towards this direction [6,7,8,9]. The possible contamination of food by CoV has rarely been identified in field investigations, possibly due to the poor recovery efficiency of existing analytical methods [10]. However, it cannot be ruled out that food might be contaminated by CoV, although it is in debate under which circumstances contaminated food has disease potential. On the other hand, CoVs have been detected in water contaminated by faecal material [11,12]. Sewage treatment is thought to inactivate them; still, monitoring of sewage effluents has been utilized for epidemiological surveillance of the infected population since SARS–CoV–2 RNA, and possibly viable SARS–CoV–2, are present in sufficiently high concentrations, permitting their detection and quantification [13,14]. Therefore, it is expected that CoVs can end up in surface water bodies in high concentrations, contaminating water reservoirs and, possibly, aquatic animals used as food.
This article aims to provide a critical review of the possibility of SARS–CoV–2 occurrence in food with an emphasis on seafood, as a stimulus for critical thinking about this yet largely unknown virus. As reported by Ceylan et al. (2020), viral foodborne diseases can be caused by contamination of water used for food production or by animal products contaminated with zoonotic viruses [15]. Water as a vehicle can further contaminate seafood that grows in it, as well as surfaces, equipment, or hands during cleaning, or food products during washing. Therefore, the following questions are discussed:
- Can SARS–CoV–2 infect food animals? Can the virus multiply in a food animal and be detected in food products of animal origin?
- Can SARS–CoV–2 be found in water and retain its infectivity for an adequate time?
- Can food get contaminated by SARS–CoV–2 through its various production processes?
- Can SARS–CoV–2 be transmitted through water or food, and if yes, which are the possible transmission pathways related to water and food ingestion?
2. Human CoVs
CoVs are enveloped, single-stranded RNA viruses ranging from 60 to 220 nm in size. The Coronaviridae family is subdivided in four genera, alpha-CoVs, beta-CoVs, gamma-CoVs and delta-CoVs, while human CoVs mostly belong to the beta-CoVs. They are further divided in the lineages A, B, C and D [16,17]. They are responsible for mild to moderate upper-respiratory tract illnesses in humans, especially the common cold. CoVs were first discovered in animals around the 1930′s, isolated from turkeys, mice, cows, pigs, cats and dogs [3]. The first human coronavirus (HCoV) was the HCoV-229E strain B814 isolated from human patients in 1965 [18]. The main CoVs linked to human disease are HCoV-229E, HCoV-OC43, SARS–CoV–1, HCoV-NL63, HCoV-HKU1, MERS–CoV and the recent SARS–CoV–2 [3,16,19]. HCoV-229E and HCoV-OC43 are generally associated with upper respiratory tract infections, usually circulating among the global community during winter months [3,20,21]. HCoV-NL63 and HCoV-HKU1 infections also manifest as respiratory illnesses, commonly in young children, the elderly and immunocompromised patients [22]. Although a zoonotic origin has been hypothesized [23], these four CoVs are reported to be highly adapted to humans, therefore having a small chance of mutating to cause highly pathogenic disease. Only in one instance, an HCoV-NL63 subtype was associated with severe lower respiratory tract infection [3]. On the other hand, SARS–CoV–1, MERS–CoV and SARS–CoV–2 are considered zoonotic since their occurrence has been traced back to mutations in animal CoVs [3,24]. SARS–CoV–1 originated in 2002 in the province of Guangdong, China. The symptoms observed in patients infected with SARS–CoV–1 were fever, headache, chills and malaise, followed by cough, dyspnoea and respiratory distress. A considerable percentage of patients (20–30%) required intensive care hospitalization and mechanical ventilation, since the disease struck the lower respiratory tract causing diffuse alveolar damage [25]. MERS–CoV is generally localized to the Middle East area. MERS disease, along with respiratory infection that progresses to acute pneumonia, exhibited renal failure—a feature unique among CoVs. In addition, patients exhibited gastrointestinal symptoms, such as diarrhoea and vomiting. The fatality rate among the patients was rather high (34.4%), making MERS–CoV one of the most lethal viruses infecting humans [26].
Concerning SARS–CoV–2, a metanalysis involving 3600 CoVID-19 patients revealed that the patients’ median age was 41 years and that 43.5% were female; the most common symptoms were fever (83.3%), cough (60.3%) and fatigue (38.0%), while diarrhoea was less common (3.8%) [27]. Other researchers have reported higher rates of gastrointestinal symptoms, with an average rate of 7.8%, and have estimated that the median incubation period is four days [28]. Regarding laboratory findings, elevated C-reactive protein, decreased lymphocyte count and increased lactate dehydrogenase were reported in 68.6%, 57.4% and 51.6% of patients, respectively, while 80.0% of patients had lymphocytopenia on admission. Chest computed tomography revealed ground-glass opacity in 80.0% and bilateral pneumonia in 73.2% of patients. Severe cases and fatalities represented 25.6% and 3.6% of cases, respectively [27].
It is therefore evident that COVID-19 combines characteristics found in both community-acquired HCoVs, SARS–CoV–1 and MERS–CoV. The incubation and duration of the disease is similar to those reported in other diseases caused by CoVs, with symptoms ranging from asymptomatic and mild and non-specific to severe. Finally, the transmissibility of SARS–CoV–2 is rather high, similar to that of the community-acquired HCoVs; it is still under question if subsequent passages of SARS–CoV–2 lower its transmissibility, as exhibited in SARS–CoV–1 and MERS–CoV [29].
3. Zoonotic Potential of Human CoVs
CoVs have been well documented as animal pathogens. Disease caused by CoVs is usually multisystemic since they can cause (i) infection of the respiratory tract; infectious bronchitis virus in chickens (IBV) [30], pheasant coronavirus (PhCoV), porcine respiratory coronavirus (PRCoV) [31], and canine respiratory coronavirus in dogs [32], (ii) neural disease; mouse hepatitis virus [33], and (iii) gastrointestinal infection; transmissible gastroenteritis coronavirus in pigs (TGEV), bovine coronavirus (BCoV), feline enteric coronavirus (FECV), feline infectious peritonitis virus (FIPV), murine hepatitis virus (MHV) [33], canine CoVs (CCoV-I and CCoV-II), and turkey coronavirus (TCoV) [31]. In several instances, CoVs have not been linked to a specific disease, but rather have modified the outcome of other viral infections, such as an increase in susceptibility to canine parvovirus infection after infection by canine CoVs [31]. In addition, animal CoVs often exhibit different system infections, such as BCoV, which is usually associated with gastroenteritis; it can also cause respiratory disease, regardless of the age of the animal [31]. Furthermore, among animal CoVs, faecal–oral transmission is regarded as the main route of transmission for swine [34], equine [35], and canine CoVs [36].
The zoonotic potential of human CoVs in animals is summarized in Table 1. Especially for SARS–CoV–2, the virus has a 96.2% nucleotide homology with the bat CoV RaTG13 found in the horseshoe bat (Rhinolophus spp.), implying that, to a certain point, SARS–CoV–2 can be considered zoonotic, as the level of homology is too high to assign a parental relationship [5,37]. The relationship between bats and CoVs is not new, since both SARS–CoV–1 and MERS–CoV originated in and are extensively harboured by these mammals. Anderson et al. (2020) have proposed two hypotheses concerning SARS–CoV–2 evolution, either that the viruses have undergone natural selection in an animal host prior to human infection, or that the selection was performed in humans after human infection [38]. The first hypothesis is most likely since a mutation in spike glycoprotein and nucleocapsid protein makes SARS–CoV–2 distinct from bat SARS–CoV–2 like viruses [39].
CoVs are among the viruses that can easily pass the “species barrier”, enabling transmission between species. As a result, they can have a broad range of hosts, in contrast to most viruses that exert a host-specific pathogenicity [46]. The main site of SARS–CoV–2 attachment, the angiotensin-converting enzyme 2 (ACE-2) receptor, is considered to be the key feature that enables or does not enable the infection of an animal. The genetic background of this receptor seems to be highly conserved among mammal species [47]. Still, although ACE-2 homology studies showed that SARS–CoV–2 transmission from bats to humans is unlikely, the adaptation of the bat SARS-like-CoV to humans shows the virus’s ability to pass the interspecies barrier. Among animals, the human ACE-2 proteins are closer to that of cat and dog than of rabbit, mouse, rat, hamster, bat, pig, sheep, goat, and cattle ACE-2. Damas et al. [48] have studied the sequences of the ACE-2 of 410 vertebrates, in order to assess the binding ability of SARS–CoV–2 and the possibility of infection by the virus. The binding ability was reported as “very high” among 18 species (all primates), and that of 28 species was reported as “high” (mostly cetaceans, rodents, cervids, and lemuriform primates).
Concerning domestic animals, the most susceptible species are cats and ferrets with dogs having lower susceptibility [31,49]. Other animals, such as chickens, ducks, and pigs, and food animals, such as sheep, cattle, horses and rabbits, are also susceptible, with their susceptibility characterized as low [49,50,51]. It is interesting though that dogs and pigs have a high ACE-2 homology to humans—perhaps the limited expression of this protein in the respiratory tract limits their susceptibility to the virus [46]. Consequently, it is imperative to perform infectivity studies in addition to studies in silico. Infectivity studies on pigs have reported indecisive results: Pickering et al. (2021) report that eight-week-old, crossbred pigs were susceptible to SARS–CoV–2 infection after oronasal inoculation. Still, live virus was isolated from one animal, in which it was retained only in the submandibular lymph node [52]. In contrast, Meekins et al. (2020) reported that the challenge of five-week-old pigs with SARS–CoV–2 did not result in infection [50]. Ulrich et al. (2020) have inoculated six cattle with SARS–CoV–2 and kept them in proximity with three non-inoculated animals. The latter did not become infected, proposing that SARS–CoV–2 cannot be transmitted among cattle [53]. In contrast, Di Teodoro et al. (2021) reported that upon infection of cattle and sheep ex vivo organ cultures, two strains of SARS–CoV–2 used for infection could sustain viral replication in vitro, in contrast to pig tissues. In addition, they were associated with respiratory tract ACE2-expressing cells of the ruminants [54]. Concerning SARS–CoV–2-specific antibodies in animals, Deng et al. (2020) examined, among others, domestic livestock (cow, horse, rabbit, sheep, pig, chicken, duck, and goose), and all samples showed negative results [55]. In conclusion, there is controversy concerning the susceptibility of food animals to SARS–CoV–2. Still the possibility of human transmission via food animals is unlikely, since the infectivity according to the experimental infections was mild and the virus shedding quite low. Nevertheless, it cannot be ruled out at this stage.
4. Transmission Pathways Related to Ingestion
Transmission of CoVs is thought to occur directly from person to person via coughs, sneezes, and droplet inhalation, or by contact transmission with oral, nasal, and eye mucous membranes [56]. The primary mode of transmission of CoVs is thought to be through respiratory droplets, mainly due to their rapid spread during outbreaks; however, there is no direct evidence to support this [8]. Besides SARS–CoV–2, some of the other CoVs (like SARS-CoV and MERS-CoV) can be transmitted via the faecal–oral route [6,8,56,57,58] and have been traced to the human gastrointestinal tract, implying foodborne transmission [59]. Apart from symptoms in the respiratory system, symptoms from the gastrointestinal system have been extensively reported, even from the first stages of disease. Main gastrointestinal symptoms included diarrhoea (2.0–55.0%), anorexia, vomiting (1.0–12.5%) and nausea (1.0–27.5%) [1,2,60,61,62,63,64,65]. Regarding the occurrence of gastrointestinal symptoms along with respiratory manifestations, patients could be categorised into three main groups: patients with concurrent gastrointestinal symptoms, patients with onset of gastrointestinal symptoms prior to respiratory symptoms, and patients with only gastrointestinal symptoms [65]. Therefore, infection of the gastrointestinal system is certain, based on the observed clinical signs. Further, the presence of gastrointestinal symptoms is the most common characteristic of foodborne diseases. In order though to verify such a hypothesis, there are some criteria that must be met [6,59].
Several authors reported the existence of a faecal–oral route of SARS–CoV–2 transmission, based on the observed gastrointestinal symptoms and the occurrence of the ACE-2 receptor in the cells of gastrointestinal tract. In order to verify this hypothesis, there is a need for proof of (i) infection of cells of the gastrointestinal tract, (ii) shedding of infectious viruses in stool and (iii) infection by ingestion of the virus. Concerning infection of cells of the gastrointestinal tract, a perquisite for faecal shedding, entry-genes of SARS–CoV–2 have been observed in the cells of intestinal tissues [66]. Lamers et al. (2020) have reported that SARS–CoV–2 can infect human enterocytes in small intestine organoid models [45]. Zang et al. (2020) have also reported the productive infection of enterocytes in human small intestinal enteroids. Nevertheless, they have also reported that the simulated intestinal fluid inactivated the viruses, therefore predicting that active shedding does not occur [67]. ACE-2 messenger RNA is highly expressed in the gastrointestinal system [68], implying the potential of virus attachment and infection of cells of this system.
Concerning the previous SARS epidemic in 2003, Leung et al. (2003) reported that SARS–CoV–1 was more frequently isolated from the small intestine than the lung [69]. A first proof of shedding reported by several scientists, involves the high RNA level in stool [63,70,71,72]. Regarding detection of infectious virus, Wölfel et al. (2020) reported that, although they did not succeed to isolate infectious SARS–CoV–2 from stool samples taken from COVID patients, they detected cells containing subgenomic RNA, suggesting active viral replication in the intestinal cells [70]. Wang et al. (2020) observed live viruses in stool specimens of two among four patients with high viral RNA concentration in their stool [63]. Additionally, Xiao et al. (2020) reported the identification of live virions from stool, however, without providing further information [68]. Parasa et al. (2020) also reported that live virus was shed from CoVID-19 patients even days after hospitalization [28]. Oral transmission requires the presence of ACE-2 receptors in the oral cavity, the oesophagus, and the other parts of the gastrointestinal tract. It has been hypothesised that the intestine could possibly be the primary site of infection [67]. For this to happen, the virus should not be inactivated when passing through the low pH environment of the stomach. Since most CoVs share more or less similar resistance potentials, the primarily faecal–oral transmission of other animal CoVs (swine CoVs, horse CoV, and canine CoVs) suggest that CoVs can be transmitted through this route [73]. It is not yet clear whether protective conditions, such as the incorporation of the virus particles in saliva or in food can enable virus passage through the stomach acidic gastric fluid and further infect the intestinal cells. Otherwise, the presence of infectious SARS–CoV–2 in the oral cavity or the pharynx can result in infection via the respiratory system [74], as proposed for several respiratory viruses such as other CoVs, adenoviruses, and influenza viruses, which can infect via contact with mucosal membranes [75].
5. Occurrence and Survival in Water
Contamination of sewage water and the effluents of biological treatment is currently used for monitoring the prevalence of COVID-19 in the population, since it is independent of hospitalization or severity of the disease [76]. The occurrence of SARS–CoV–2 in water can result in contamination of surface water further used for irrigation or aquaculture, and even end up in potable water distribution in cases where surface water is used after treatment. Research concerning waterborne SARS–CoV–2 is under development with a limited amount of published experimental data. For the time being, the characteristics that influence the occurrence and survival of SARS–CoV–2 in water can be derived from studies on other CoVs, since the virus structure is largely common. Therefore, research on other CoVs can be extrapolated to predict the behavior of SARS–CoV–2 in water.
Wang et al. (2005) analysed 20 sewage samples from hospitals with SARS patients, and all tested negative for infectious SARS–CoV–1, while the viral genome was detected in 15% of samples after disinfection (100% of samples before disinfection) [77]. Corsi et al. (2014) examined 63 large volume (50 L) samples for several waterborne viruses, including bovine coronavirus, and all were negative [78]. Blanco et al. (2019) reported that among 21 surface water samples, only one was found positive for alphacoronavirus [79]. Field investigations for CoVs until 2019 have confirmed the scarcity of data on occurrence of CoVs in water [10]. Ahmed et al. (2020) applied two different CoV concentration methods on nine samples and tested them for CoVs by a RT–qPCR method; two different samples concentrated by different methods tested positive [80]. Medema et al. (2020) reported that CoVs were detected in 14 (77.8%) of 18 sewage samples [81], whereas Wu et al. (2020) detected SARS–CoV–2 in all 10 samples collected from a wastewater treatment plant [71]. Nemudryi et al. (2020) found positive results for SARS–CoV–2 in all seven samples of raw sewage they tested [82], whereas Wurtzer et al. (2020) reported that 100% and 75% of raw and treated wastewater samples from urban wastewater treatment plants were SARS–CoV–2 positive [83]. Although SARS–CoV–2 is regularly found in treated and untreated wastewater, there is lack of standardized concentration methods for enveloped viruses [5,84]. Concerning other types of water, CoVs have rarely been detected in field investigations, possibly due to the low recovery efficiency of analytical methods [10]. The possibility that SARS–CoV–2 can be transmitted through water has driven some countries, such as North Korea, to monitor surface waters used as sources of drinking water, even though 61 other countries are reassuring the public about the safety of drinking water [11]. The CDC also reported no SARS–CoV–2 detection in tap water [85]. The factors that can influence the survival of the virus in water include organic matter, temperature, and aerobic microorganisms. Other factors, such as predatory organisms (protozoa), nucleases and proteases, have been reported to increase the virus inactivation rate [86]. Additional factors may influence the potential of CoV transmission via water, such as the chlorination of water, which is currently known to destroy the lipoprotein of the virus [10]. Studies that report findings concerning the survival of CoVs in water are shown in Table 2 and Table 3.
Gundy et al. (2009) examined the behaviour of representative CoVs, feline infectious peritonitis virus, and human coronavirus 229E in filtered tap water, and found that CoV reduction of 99.9% can be achieved by 10 days at room temperature and by more than 100 days at 4 °C. They also reported that CoV survival in untreated tap water samples was more possible than in filtered samples, stating that the organic matter and suspended solids can provide protection for viruses. The mode of action proposed is the adsorption of the virus to these particles and the protection of CoV in water from other factors [87]. Casanova et al. (2009) examined the survival of transmissible gastroenteritis virus (TGEV) and mouse hepatitis virus (MHV); both viruses remained infectious in water and sewage for days to weeks. The time required for 99% reduction at 25 °C was 22 days for TGEV and 17 days for MHV in reagent-grade water, whereas in pasteurized sewage, 99% reduction occurred by 9 days for TGEV and 7 days for MHV. At 4 °C, there was < 1 log10 infectivity decrease for both viruses after four weeks. In addition, infectivity declined faster in wastewater, with 2 log10 reduction after nine and seven days for TGEV and MHV, respectively, in pasteurized sewage at 23–25 °C, and 2 log10 and 1 log10 reduction, respectively, after 35 days at 4 °C [88]. One of the first scientific works reporting SARS–COV–2 survival in water is that of Bivins et al. (2020), who examined the persistence of both infectious SARS–CoV–2 virus and RNA in water and wastewater; they reported that a 90% reduction of viable virus in wastewater and tap water at room temperature was achieved in 1.5 and 1.7 days, respectively, while the infectious virus persisted for the entire week of experimentation in water with a high-starting titer (105 TCID50 mL–1) [89]. Sala-Comorera et al. (2021) examined the survival of SARS–CoV–2 in surface water and seawater; the observed T90 of infectious SARS–CoV–2 at 4 °C was 3.8 and 2.2 days in river and seawater, whereas at 20 °C the T90 was 2.3 and 1.1 days, respectively, suggesting that the expected survival time of SARS–CoV–2, although low, can permit further spread through water [86].
A technical brief on water, sanitation, hygiene, and waste management for COVID-19 has been issued by the FAO (3 March 2020; amended 29 July 2020) [91]. It indicates that the existing WHO guidance should be efficient for prevention of tap water contamination by SARS–COV–2, since SARS–CoV–2 is an enveloped virus, and therefore, more susceptible than non-enveloped human enteric viruses. It also emphasizes that standard hygienic procedures, including personal protection equipment, should be used in relevant areas, such as wastewater treatment plants. Nevertheless, the brief states that at the time of publication the knowledge regarding the presence of CoVs in water is limited. As a conclusion, and according to the experimental data, it seems that contaminated water, especially when no disinfection occurs, can be a potential vehicle for human exposure— either through the respiratory tract via aerosolised contaminated water or through ingestion.
6. Food and CoVs
6.1. Other CoVs
CoVs have been shown to be resistant to environmental factors, making them quite hard to eliminate in various matrices including biological material [92]. Concerning SARS–CoV–1 and MERS, limited information on their transmission potential through the environment is available, and scientists work mainly on the survival of the virus on surfaces rather than in water. In the SARS epidemic in 2003, there was no strong evidence for food or waterborne transmission, and the WHO (2004) acknowledged that further studies were needed to define the potential role of these routes [93]. In a report by the FAO/WHO (2008) the foodborne transmission of SARS-CoV-1 is acknowledged, although it is stated that this is not the main transmission route [40]. Further, MERS–CoV has been characterized as foodborne since it could be transmitted by camel milk and its products [94]. Camels were considered as intermediate hosts for MERS–CoV between bats and humans. The virus has been detected in camel nasal swabs, faeces, and calves’ saliva suckling prior to milking. Therefore, nasal secretions, faeces, and milk can form the possible contamination routes of food or environment. In the case of MERS–CoV, the foodborne route of infection is reported as an alternative to respiratory transmission, contributing to virus transmission [95].
6.2. Is It Foodborne?
Regarding the possible SARS–CoV–2 sources of food contamination, it is evident, according to their susceptibility, that domestic animals are not likely to play such a role. It is more possible that the anthropogenic environment or the water used in various food production stages can contaminate the food or any material coming into contact with food. Food contamination by animals was reported at the start of the COVID-19 pandemic at Huanan market in the Wuhan province. In this market, several different types of food are sold, with foods of plant and animal origins being in close proximity. Except from the common meat sources, markets in China also include exotic animal products and live animals. In such markets, animal species susceptible to zoonotic CoVs can be found close to other animal species and humans [96]. Therefore, contaminated food eaten without prior treatment, such as fruits and vegetables, could provide a suitable vehicle for foodborne transmission. This hypothesis of faecal–oral transmission in certain cases was also reported by Yuen et al. (2020). Especially in the super spreading event that took place on board the Diamond Princess cruise ship, with at least 742 cases among approximately 3700 passengers, the involvement of a route different than the airborne transmission route was suspected [29]. The faecal–oral potential of SARS–CoV–2 has been shown in laboratory animals. African green monkeys challenged with SARS–CoV–2 were infected in both the respiratory and gastrointestinal tracts [97], whereas orally inoculated golden Syrian hamsters developed both respiratory and intestinal infection [98]. Both animal species are susceptible to SARS–CoV–2 with similar characteristics to human disease, with African green monkeys being primates phylogenetically close to humans. Therefore, it is an indirect evidence of the faecal–oral transmission of SARS–CoV–2 in humans.
6.3. Surfaces
Among routes of food contamination are the surfaces with which food comes in contact. The hygiene of surfaces is of paramount importance for foodborne pathogen transmission [99,100,101]. Actually, the most important foodborne viruses, like noroviruses, hepatitis A virus and sapoviruses, are mostly found on the exterior of food as a result of contact with water or surfaces, or infected handlers [100,102]. Prevention measures should be in place to hinder virus transmission by contact with contaminated surfaces, in order to avoid the indirect infection of the human oral or nasal mucosa by these known foodborne viruses. CoVs are known to be transmitted via the faecal–oral route [71,80,96,103,104], and have been traced in the human gastrointestinal tract, implying foodborne transmission [10,105]. Contamination of surfaces can occur directly by contaminated personnel neglecting the proper use of protective equipment, or by the aerosolization and subsequent airborne transport of the virus from faecally contaminated water or toilets—a route that has been demonstrated for other viruses and has been proposed for SARS–CoV–2 [96,106,107,108].
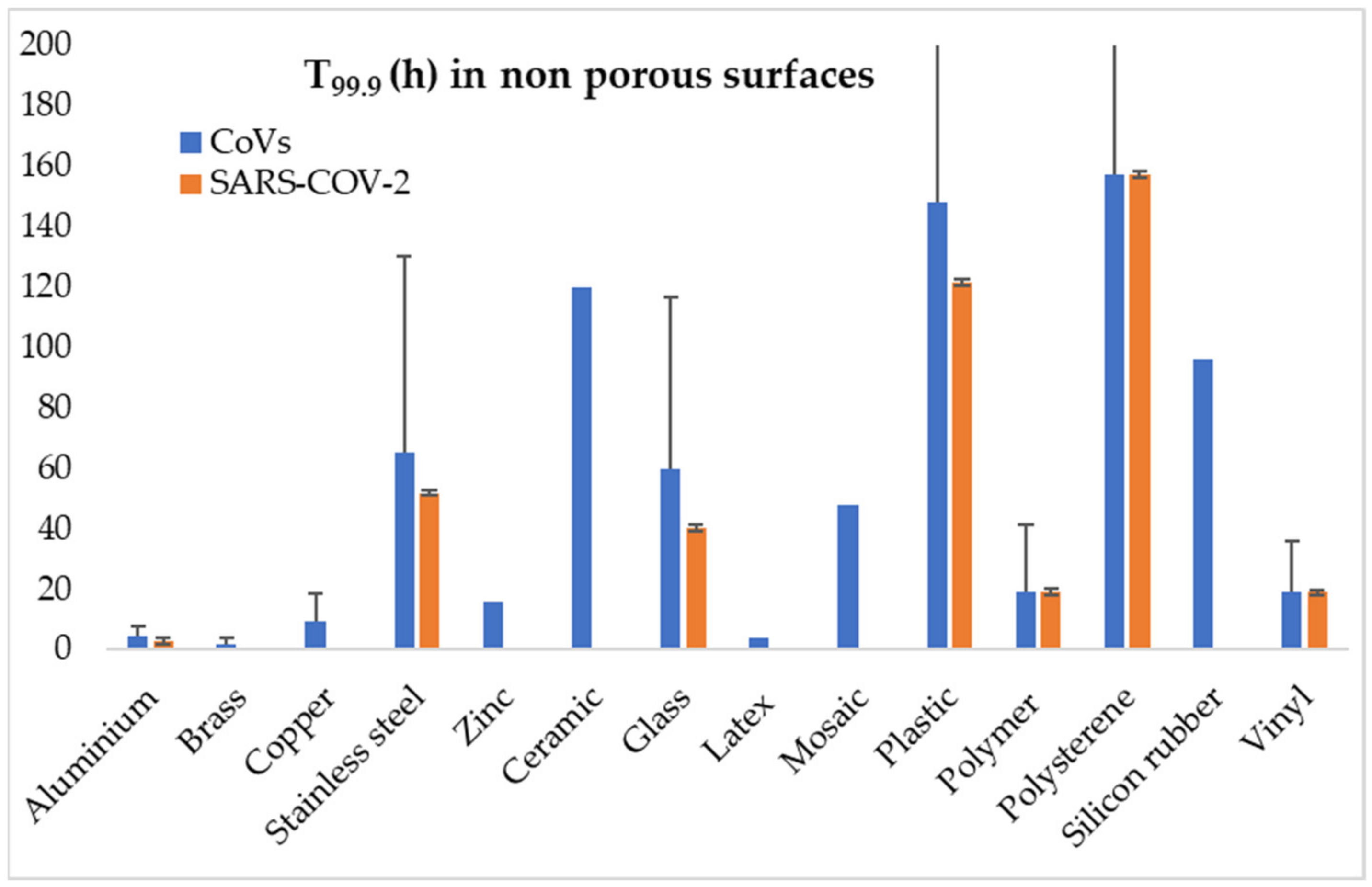
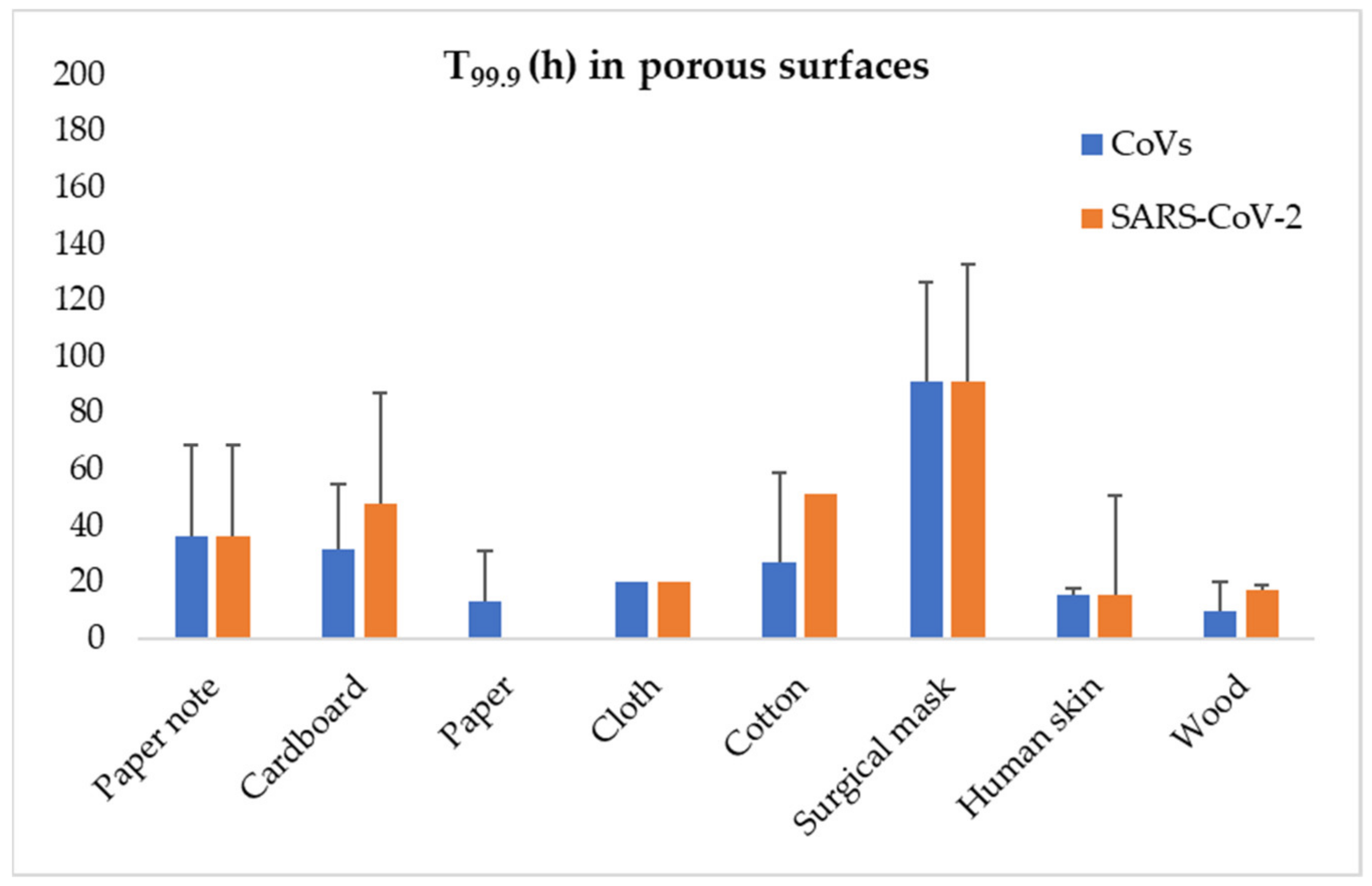
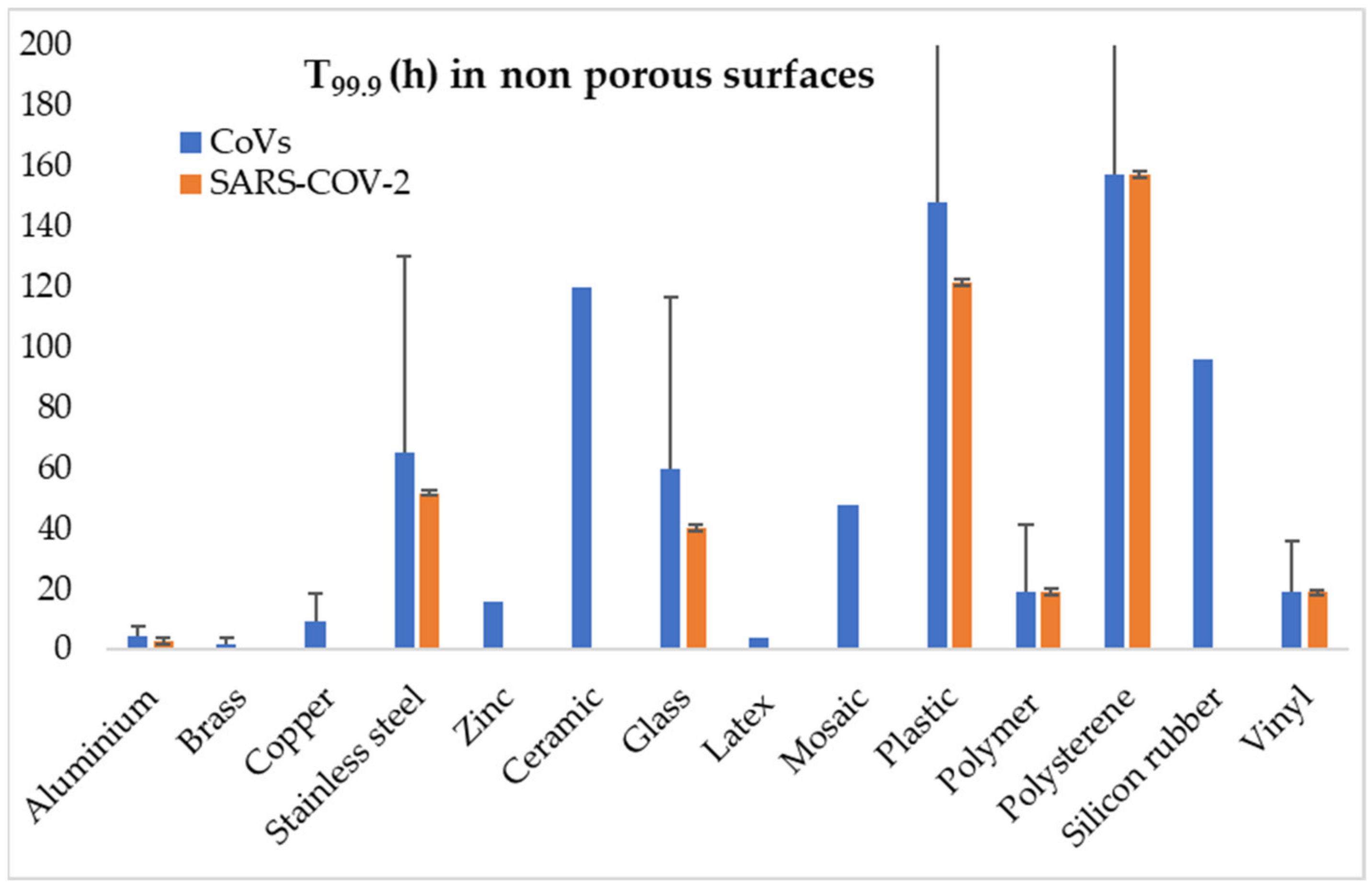
The indirect transmission of SARS–CoV–2 through contact surfaces has been given much attention, since it is of importance to several premises, including hospital settings. Surfaces though are also extremely important for the contamination of food. Previous CoVs, such as SARS–CoV–1 and MERS–CoV, have been reported to survive on surfaces for several days, with factors such as the surface type, temperature and humidity being critical for the survival of the virus [96,108]. Concerning SARS–CoV–2, Chin et al. (2020) report that its infectivity is lost after four days on plastic surfaces at room temperature and 65% relative humidity [109]. Van Doremalen et al. (2020) reported no significant difference in behaviour on surfaces between SARS–CoV–1 and SARS–CoV–2, whereas on metal surfaces the type of material used is of importance, with SARS–CoV–1 retaining its infectivity on copper for 8 h in contrast to stainless steel, where it is totally destroyed after three days; for SARS–CoV–2, they reported that the survival time was 4 h for copper, 24 h for cardboard, and 2–3 days for plastic and stainless steel [110]. Kasloff et al. (2020) examined the survival of SARS–CoV–2 on surfaces of medical importance reporting, among others, that the virus can survive for up to 14 days on stainless steel at room temperature [111]. Results of the available studies on survival of CoVs, and specifically SARS–CoV–2, on porous and non-porous surfaces are graphically presented at Figure 1 and Figure 2.
6.4. CoV Behaviour in Food Treatments
Concerning the possibility of SARS–CoV–2 survival in food, the available information is derived solely from SARS–CoV–1 and other CoVs. To our knowledge, there are currently no data regarding the survival of SARS–CoV–2 in food matrices. CoVs are known to survive in fridge temperatures, where food is usually stored. Some parameters seem of benefit to CoVs, such as the stabilizing effect of low temperature, the high relative humidity, and the protective action of organic materials. In general, thermal treatment at temperatures higher than 70 °C is reported to be sufficient for CoVs inactivation [123]. The exposure of SARS–CoV–1 to 56 °C over 30 min has been reported to reduce the virus titre to an undetectable level, except when 20% protein was added [117]. CoVs are stable at 4 °C, and also at 20 °C and 37 °C for at least 2 h, with loss of infectivity happening at high temperatures, such as 56 °C, 67 °C and 75 °C after 90, 60 and 30 min, respectively [113]. In addition, it was reported that CoVs lose their infectivity by 72 h after drying on plastic surface [117]. Concerning pH, CoVs are more stable at slightly acidic pH (6–6.5) than at alkaline pH (8) [117]. Therefore, foods that are slightly acidic and in contact with water can provide a suitable transmission vehicle for CoVs. In addition, acidification at low pH values is regarded as an efficient method to destroy the virus. Wang et al. (2005) reported that the nucleocapsid of SARS–CoV–1 starts to unfold at pH 5.0, while it unravels completely at pH 2.7 [124]. Disinfection with quaternary ammoniums or phenolic compounds, disinfectants routinely used in the food industry, should be avoided since their efficacy is questionable [92,125].
6.5. Food Type and SARS–CoV–2
Food can be contaminated by contact or by respiratory droplets at several points during the “farm-to-table” chain. This could happen during farming, processing, storage, transport, and retailing processes, where the conditions concerning contact and environmental factors are optional. The safety of different commodities is largely influenced by their characteristics. The several clusters of cases associated with abattoirs and food handling premises has aroused public opinion about the possibility of transmission through meat and meat products [126,127]. BfR (2020), upon answering the question of whether SARS–CoV–2 can be transmitted through meat products, stated that this is unlikely, especially since most domestic animals (and especially pigs and chickens) are not infected. In addition, meat and meat products are generally heat-treated prior to consumption, while pH usually drops significantly during the slaughter process and product preparation, since the pH of fermented products is usually ≤ 4.6 [128]. Therefore, meat and meat products are generally regarded as safe, with the only possibility of contamination by unclean surfaces or handlers who do not keep routine hygiene measures [129]. This is not the case for fruits, vegetables, or seafood. The consumption of fruits and vegetables without any treatment is quite common, since often they are not being washed thoroughly [100,101]. Possible water contamination by SARS–CoV–2 and subsequent contamination of food washed with it or animals living in it, makes the latter the most prominent candidate for SARS–CoV–2 transmission.
Specifically for seafood, Bondad-Reantaso et al. (2020) reported that aquatic food animals cannot be infected by SARS–CoV–2 since beta-CoVs are mammal-specific, targeting ACE-2 receptors that are not found in fishes or other seafood. Nevertheless, they agreed that the surface of fish products may potentially become contaminated with SARS–CoV–2 when handled by infected people; however, no relevant data were available at the time [9]. Since CoVs are destroyed during normal cooking, they are considered safe. This is the case with several thermolabile infectious agents in which cross-contamination can result in infection after consumption by heat-treated food. Bondad-Reantaso et al. (2020) agreed that these opinions are based on general knowledge about viruses and not by experimental data [9]. According to Yépiz-Gómez et al. (2013), emerging zoonotic viruses, such as respiratory CoVs and influenza viruses, may potentially be transmitted via contaminated foods [130]. In addition, Conway (2020) reported the detection of SARS-like CoVs sequences in the internal organs (kidneys) of carps from the Wuhan area, China, and suggested that some carp species may be open to CoV infection [131]. The detection of SARS–CoV–2 has been reported in imported frozen food [132,133]. Liu et al. (2020) reported the isolation of SARS-CoV-2 that retained its infectivity in tissue culture from the outer package of imported frozen cod [134]. Pang et al. (2020) reported that there is genetic evidence linking the COVID-19 resurgence in Beijing to cold-chain food contamination, specifically salmon [135]. Frozen foods served as a vehicle for remote transmission [136]. However, there is limited data on HCoV survival on fresh products. There is no opinion on the infection possibility of shellfish, which, as with most food animals, mammals or not, probably cannot be infected by CoVs or most other human pathogenic viruses. However, shellfish are considered potent carriers of foodborne viruses, without being infected themselves [94]. Contamination of bivalves with hepatitis A virus and noroviruses is quite common and always associated with human faecal pollution of the water in which the shellfish are kept [137]. SARS–CoV–2 is excreted by the gastrointestinal tract of patients, and contaminates sewage water and subsequent water bodies; therefore, the possibility of contamination of shellfish cannot be excluded. Consequently, it is highly probable that bivalves reared in contaminated water bodies can be contaminated with SARS–CoV–2.
Public consultations are stating that food is not a probable vehicle of SARS–CoV–2 transmission, but there is lack of experimental data supporting this opinion. CDC characterised the risk of infection by eating or handling food as very low and urged the keeping of usual hygiene procedures in food handling and preparation [85]. ANSES (2020) states that the possible contamination of foodstuffs of animal origin via an infected animal has been ruled out, since there are only limited data on animal disease by SARS–CoV–2 [138]. This report found environmental contamination by infected food handlers to be the most possible source for food contamination; therefore, strict hygienic practices should be applied. In addition, it reports that there is no evidence that food consumption can lead to infection of the digestive tract, due to the low pH of the stomach; however, the contamination of the upper respiratory tract through food consumption cannot be excluded. Moreover, the low pH in the stomach, which is a potent microbial inhibitor, can sometimes be neutralized, e.g., in cases where decrease or elimination of gastric acid secretion is provoked, such as in peptic ulcer treatment [139]. These points are in line with the reports of BfR (2020) and EFET (2020), with the first not considering the possibility of transmission through contact [128,140]. The initial statements by these organisations were supported by the knowledge of that time. Therefore, the new information produced could lead to more robust positions, possible revisions of certain aspects and better justification of the points made.
7. Conclusions
SARS–CoV–2 is a rather bizarre virus, exhibiting high transmissibility and severe disease. As already postulated by several researchers [75,96,105,136,141], the theoretical possibility of foodborne transmission is high. The information gathered since the start of the pandemic has largely augmented our knowledge, suggesting that susceptibility of food animals is limited, and even if they are infected, they are unlikely to be the source of human infection. On the contrary, the virus in contaminated water retains its infectivity for a period of time adequate for infection. However, there are no reports of direct transmission through water, while SARS–CoV–2 has not been detected in tap water. The hypothesis of indirect waterborne transmission is valid, and seafood that come into direct contact with contaminated water can harbour the virus under certain conditions. Similarly, no experimental or other data are available. Furthermore, food contamination by water or surfaces can occur, as evidenced by limited experimental data concerning the recovery of the virus from the surface of food and from food packages. The faecal–oral transmission could be possible since SARS–CoV–2 has the behaviour and resistance characteristics of viruses that are known foodborne pathogens. Although there are indications that this route of infection could occur, the question of whether SARS–CoV–2 can be transmitted through water or food remains to be answered. The even more complex question lies in the extent to which these routes contribute to the spread of COVID-19, especially compared to the established airborne route of transmission. Therefore, the only way to avoid any potential food- or water-borne transmission of SARS–CoV–2 is the strict application of hygiene measures at all stages of food production and preparation.
Author Contributions
Writing—original draft preparation, V.E. and H.S.; writing—review and editing, E.B., A.P. and N.S.; supervision, N.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Guan, W.; Ni, Z.; Hu, Y.; Liang, W.; Ou, C.; He, J.; Liu, L.; Shan, H.; Lei, C.; Hui, D.S.C.; et al. Clinical characteristics of coronavirus disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Ellenberg, R.M.; Graham, K.E.; Wigginton, K.R. Survivability, partitioning, and recovery of enveloped viruses in untreated municipal wastewater. Environ. Sci. Technol. 2016, 50, 5077–5085. [Google Scholar] [CrossRef] [PubMed]
- Goraichuk, I.V.; Arefiev, V.; Stegniy, B.T.; Gerilovych, A.P. Zoonotic and Reverse Zoonotic Transmissibility of SARS-CoV-2. Virus Res. 2021, 302, 198473. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Kingsley, D.H. Emerging foodborne and agriculture related viruses. Microbiol. Spectrum 2016, 4. [Google Scholar] [CrossRef]
- Jalava, K. First respiratory transmitted food borne outbreak? Int. J. Hyg. Environ. Health 2020, 226, 113490. [Google Scholar] [CrossRef]
- Heller, L.; Mota, C.R.; Greco, D.B. COVID-19 faecal-oral transmission: Are we asking the right questions? Sci. Tot. Environ. 2020, 729, 138919. [Google Scholar] [CrossRef]
- Bondad-Reantaso, M.G.; Mackinnon, B.; Bin, H.; Jie, H.; Tang-Nelson, K.; Surachetpong, W.; Alday-Sanz, V.; Salman, M.; Brun, E.; Karunasagar, I.; et al. Viewpoint: Sars-CoV-2 (the cause of COVID-19 in humans) is not known to infect aquatic food animals nor contaminate their products. Asian Fish. Sci. 2020, 33, 74–78. [Google Scholar] [CrossRef]
- Carducci, A.; Federigi, I.; Liu, D.; Thompson, J.R.; Verani, M. Making Waves: Coronavirus detection, presence and persistence in the water environment: State of the art and knowledge needs for public health. Water Res. 2020, 179, 115907. [Google Scholar] [CrossRef]
- La Rosa, G.; Bonadonna, L.; Lucentini, L.; Kenmoe, S.; Suffredini, E. Coronavirus in water environments: Occurrence, persistence and concentration methods—A scoping review. Water Res. 2020, 179, 115899. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Alamin, M.; Kuroda, K.; Dhangar, K.; Hata, A.; Yamaguchi, H.; Honda, R. Potential discharge, attenuation and exposure risk of SARS-CoV-2 in natural water bodies receiving treated wastewater. NPJ Clean Water 2021, 4, 8. [Google Scholar] [CrossRef]
- Hart, O.E.; Halden, R.U. Computational analysis of SARS-CoV-2/CoVID-19 surveillance by wastewater-based epidemiology locally and globally: Feasibility, economy, opportunities and challenges. Sci. Total Environ. 2020, 730, 138875. [Google Scholar] [CrossRef] [PubMed]
- Petala, M.; Dafou, D.; Kostoglou, M.; Karapantsios, T.; Kanata, E.; Chatziefstathiou, A.; Kotoulas, K.; Arsenakis, M.; Roilides, E.; Sklaviadis, T.; et al. A physicochemical model for rationalizing SARS-CoV-2 concentration in sewage. Case study: The city of Thessaloniki in Greece. Sci. Total Environ. 2021, 755 Pt 1, 142855. [Google Scholar] [CrossRef] [PubMed]
- Ceylan, Z.; Meral, R.; Cetinkaya, T. Relevance of SARS-CoV-2 in food safety and food hygiene: Potential preventive measures, suggestions and nanotechnological approaches. Virus Dis. 2020, 31, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Wong, G.; Shi, W.; Liu, J.; Lai, A.C.K.; Zhou, J.; Liu, W.; Bi, Y.; Gao, G.F. Epidemiology, Genetic Recombination, and Pathogenesis of Coronaviruses. Trends Microbiol. 2016, 24, 490–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choudhary, J.; Dheeman, S.; Sharma, V.; Katiyar, P.; Karn, S.K.; Sarangi, M.K.; Chauhan, A.K.; Chauhan, A.K.; Verma, G.; Baliyan, N. Insights of severe acute respiratory syndrome coronavirus (SARS-CoV-2) pandemic: A current review. Biol. Proc. 2021, 23, 5. [Google Scholar] [CrossRef]
- Kapikian, A.Z. The coronaviruses. Dev. Biol. Stand. 1975, 28, 42–64. [Google Scholar]
- Cui, J.; Li, F.; Shi, Z.L. Origin and evolution of pathogenic coronaviruses. Nat. Rev. Microbiol. 2019, 17, 181–192. [Google Scholar] [CrossRef] [Green Version]
- Van Der Hoek, L. Human coronaviruses: What do they cause? Antiviral Ther. 2007, 12, 651–658. [Google Scholar]
- Tyrrell, D.A.; Cohen, S.; Schlarb, J.E. Signs and symptoms in common colds. Epidemiol. Infect. 1993, 111, 143–156. [Google Scholar] [CrossRef] [Green Version]
- Corman, V.M.; Muth, D.; Niemeyer, D.; Drosten, C. Hosts and sources of endemic human coronaviruses. Adv. Virus Res. 2018, 100, 163–188. [Google Scholar] [CrossRef]
- Stout, A.E.; Millet, J.K.; Stanhope, M.J.; Whittaker, G.R. Furin cleavage sites in the spike proteins of bat and rodent coronaviruses: Implications for virus evolution and zoonotic transfer from rodent species. One Health 2021, 13, 100282. [Google Scholar] [CrossRef] [PubMed]
- Shereen, M.A.; Khan, S.; Kazmi, A.; Bashir, N.; Siddique, R. COVID-19 infection: Origin, transmission, and characteristics of human coronaviruses. J. Adv. Res. 2020, 24, 91–98. [Google Scholar] [CrossRef]
- Cheng, V.C.; Lau, S.K.; Woo, P.C.; Yuen, K.Y. Severe acute respiratory syndrome coronavirus as an agent of emerging and reemerging infection. Clin. Microbiol. Rev. 2007, 20, 660–694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hilgenfeld, R.; Peiris, M. From SARS to MERS: 10 years of research on highly pathogenic human coronaviruses. Antiviral Res. 2013, 100, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Wang, B.; Yuan, T.; Chen, X.; Ao, Y.; Fitzpatrick, T.; Li, P.; Zhou, Y.; Lin, Y.F.; Duan, Q.; et al. Clinical characteristics of coronavirus disease 2019 (CoVID-19) in China: A systematic review and meta-analysis. J. Infect. 2020, 80, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Parasa, S.; Desai, M.; Thoguluva Chandrasekar, V.; Patel, H.K.; Kennedy, K.F.; Roesch, T.; Spadaccini, M.; Colombo, M.; Gabbiadini, R.; Artifon, E.L.A.; et al. Prevalence of gastrointestinal symptoms and fecal viral shedding in patients with Coronavirus Disease 2019: A systematic review and meta-analysis. JAMA Netw. Open 2020, 3, e2011335. [Google Scholar] [CrossRef] [PubMed]
- Yuen, K.S.; Ye, Z.W.; Fung, S.Y.; Chan, C.P.; Jin, D.Y. SARS-CoV-2 and COVID-19: The most important research questions. Cell Biosci. 2020, 10, 40. [Google Scholar] [CrossRef] [Green Version]
- Bande, F.; Arshad, S.S.; Omar, A.R.; Bejo, M.H.; Abubakar, M.S.; Abba, Y. Pathogenesis and diagnostic approaches of avian infectious bronchitis. Adv. Virol. 2016, 2016, 4621659. [Google Scholar] [CrossRef] [Green Version]
- Bonilauri, P.; Rugna, G. Animal Coronaviruses and SARS-COV-2 in Animals, What Do We Actually Know? Life 2021, 11, 123. [Google Scholar] [CrossRef]
- Erles, K.; Brownlie, J. Canine respiratory coronavirus: An emerging pathogen in the canine infectious respiratory disease complex. Vet. Clin. N. Am. Small Anim. Pract. 2008, 38, 815–825. [Google Scholar] [CrossRef]
- Hierholzer, J.C.; Broderson, J.R.; Murphy, F.A. New strain of mouse hepatitis virus as the cause of lethal enteritis in infant mice. Infect. Immun. 1979, 24, 508–522. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Vlasova, A.N.; Kenney, S.P.; Saif, L.J. Emerging and re-emerging coronaviruses in pigs. Curr. Opin. Virol. 2019, 34, 39–49. [Google Scholar] [CrossRef]
- Pusterla, N.; Vin, R.; Leutenegger, C.M.; Mittel, L.D.; Divers, T.J. Enteric coronavirus infection in adult horses. Vet. J. 2018, 231, 13–18. [Google Scholar] [CrossRef]
- Decaro, N.; Buonavoglia, C. Canine coronavirus: Not only an enteric pathogen. Vet. Clin. N. Am. Small Anim. Pract. 2011, 41, 1121–1132. [Google Scholar] [CrossRef]
- Mahdy, M.A.A.; Younis, W.; Ewaida, Z. An Overview of SARS-CoV-2 and Animal Infection. Front. Vet. Sci. 2020, 7, 596391. [Google Scholar] [CrossRef] [PubMed]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The proximal origin of SARS-CoV-2. Nat Med. 2020, 26, 450–452. [Google Scholar] [CrossRef] [Green Version]
- Benvenuto, D.; Giovanetti, M.; Ciccozzi, A.; Spoto, S.; Angeletti, S.; Ciccozzi, M. The 2019-new coronavirus epidemic: Evidence for virus evolution. J. Med. Virol. 2020, 92, 455–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Food and Agriculture Organization of the United Nations; World Health Organization. Viruses in Food: Scientific Advice to Support Risk Management Activities: Meeting Report; Lawrence, T., Ed.; Microbiological Risk Assessment Series No. 13; FAO: Rome, Italy, 2008; Available online: http://www.fao.org/3/i0451e/i0451e.pdf (accessed on 27 September 2021).
- Coleman, C.M.; Frieman, M.B. Emergence of the Middle East Respiratory Syndrome Coronavirus. PLoS Pathog. 2013, 9, e1003595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- To, K.K.; Hung, I.F.; Chan, J.F.; Yuen, K.Y. From SARS coronavirus to novel animal and human coronaviruses. J. Thorac. Dis. 2020, 5 (Suppl. S2), S103–S108. [Google Scholar] [CrossRef]
- Gao, H.; Yao, H.; Yang, S.; Li, L. From SARS to MERS: Evidence and speculation. Front. Med. 2020, 10, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Shi, M.; Chommanard, C.; Queen, K.; Zhang, J.; Markotter, W.; Holmes, E.C.; Tong, S. Surveillance of Bat Coronaviruses in Kenya Identifies Relatives of Human Coronaviruses NL63 and 229E and Their Recombination History. J. Virol. 2017, 91, e01953-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamers, M.M.; Beumer, J.; van der Vaart, J.; Knoops, K.; Puschhof, J.; Breugem, T.I.; Ravelli, R.B.G.; Paul van Schayck, J.; Mykytyn, A.Z.; Duimel, H.Q.; et al. SARS-CoV-2 productively infects human gut enterocytes. Science 2020, 369, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Sharun, K.; Dhama, K.; Pawde, A.M.; Gortázar, C.; Tiwari, R.; Bonilla-Aldana, D.K.; Rodriguez-Morales, A.J.; de la Fuente, J.; Michalak, I.; Attia, Y.A. SARS-CoV-2 in animals: Potential for unknown reservoir hosts and public health implications. Vet. Q. 2021, 41, 181–201. [Google Scholar] [CrossRef]
- Sun, K.; Gu, L.; Ma, L.; Duan, Y. Atlas of ACE2 gene expression reveals novel insights into transmission of SARS-CoV-2. Heliyon 2021, 7, e05850. [Google Scholar] [CrossRef] [PubMed]
- Damas, J.; Hughes, G.M.; Keough, K.C.; Painter, C.A.; Persky, N.S.; Corbo, M.; Hiller, M.; Koepfli, K.P.; Pfenning, A.R.; Zhao, H.; et al. Broad host range of SARS-CoV-2 predicted by comparative and structural analysis of ACE2 in vertebrates. Proc. Natl. Acad. Sci. USA 2020, 117, 22311–22322. [Google Scholar] [CrossRef]
- Shi, J.; Wen, Z.; Zhong, G.; Yang, H.; Wang, C.; Huang, B.; Liu, R.; He, X.; Shuai, L.; Sun, Z.; et al. Susceptibility of ferrets, cats, dogs, and other domesticated animals to SARS-coronavirus 2. Science 2020, 368, 1016–1020. [Google Scholar] [CrossRef] [Green Version]
- Meekins, D.A.; Morozov, I.; Trujillo, J.D.; Gaudreault, N.N.; Bold, D.; Carossino, M.; Artiaga, B.L.; Indran, S.V.; Kwon, T.; Balaraman, V.; et al. Susceptibility of swine cells and domestic pigs to SARS-CoV-2. Emerg. Microbes Infect. 2020, 9, 2278–2288. [Google Scholar] [CrossRef]
- Godoy, M.G.; Kibenge, M.J.T.; Kibenge, F.S.B. SARS-CoV-2 transmission via aquatic food animal species or their products: A review. Aquaculture 2021, 536, 736460. [Google Scholar] [CrossRef]
- Pickering, B.S.; Smith, G.; Pinette, M.M.; Embury-Hyatt, C.; Moffat, E.; Marszal, P.; Lewis, C.E. Susceptibility of domestic swine to experimental infection with severe acute respiratory syndrome Coronavirus 2. Emerg. Infect. Dis. 2021, 27, 104–112. [Google Scholar] [CrossRef]
- Ulrich, L.; Wernike, K.; Hoffmann, D.; Mettenleiter, T.C.; Beer, M. Experimental infection of cattle with SARS-CoV-2. Emerg. Infect. Dis. 2020, 26, 2979–2981. [Google Scholar] [CrossRef] [PubMed]
- Di Teodoro, G.; Valleriani, F.; Puglia, I.; Monaco, F.; Di Pancrazio, C.; Luciani, M.; Krasteva, I.; Petrini, A.; Marcacci, M.; D’Alterio, N.; et al. SARS-CoV-2 replicates in respiratory ex vivo organ cultures of domestic ruminant species. Vet. Microbiol. 2021, 252, 108933. [Google Scholar] [CrossRef]
- Deng, J.; Jin, Y.; Liu, Y.; Sun, J.; Hao, L.; Bai, J.; Huang, T.; Lin, D.; Jin, Y.; Tian, K. Serological survey of SARS-CoV-2 for experimental, domestic, companion and wild animals excludes intermediate hosts of 35 different species of animals. Transbound. Emerg. Dis. 2020, 67, 1745–1749. [Google Scholar] [CrossRef] [PubMed]
- Hoseinzadeh, E.; Javan, S.; Farzadkia, M.; Mohammadi, F.; Hossini, H.; Taghavi, M. An updated min-review on environmental route of the SARS-CoV-2 transmission. Ecotoxicol. Environ. Saf. 2020, 202, 111015. [Google Scholar] [CrossRef]
- La Rosa, G.; Fratini, M.; Della Libera, S.; Iaconelli, M.; Muscillo, M. Viral infections acquired indoors through airborne, droplet or contact transmission. Ann. Ist. Super. Sanita 2013, 49, 124–132. [Google Scholar] [CrossRef]
- Khan, S.; Siddique, R.; Shereen, M.A.; Ali, A.; Liu, J.; Bai, Q.; Bashir, N.; Xue, M. Emergence of a Novel Coronavirus, Severe Acute Respiratory Syndrome Coronavirus 2: Biology and Therapeutic Options. J. Clin. Microbiol. 2020, 58, e00187-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosch, A.; Gkogka, E.; Le Guyader, F.S.; Loisy-Hamon, F.; Lee, A.; van Lieshout, L.; Marthi, B.; Myrmel, M.; Sansom, A.; Schultz, A.C.; et al. Foodborne viruses: Detection, risk assessment, and control options in food processing. Int. J. Food Microbiol. 2018, 285, 110–128. [Google Scholar] [CrossRef]
- Chen, N.; Zhou, M.; Dong, X.; Qu, J.; Gong, F.; Han, Y.; Qiu, Y.; Wang, J.; Liu, Y.; Wei, Y.; et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: A descriptive study. Lancet 2020, 395, 507–513. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Zhang, L.; Du, H.; Zhang, J.; Li, Y.Y.; Qu, J.; Zhang, W.; Wang, Y.; Bao, S.; Li, Y.; et al. SARS-CoV-2 Infection in Children. N. Engl. J. Med. 2020, 382, 1663–1665. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Zhang, X.; Xu, H. Don’t Overlook Digestive Symptoms in Patients With 2019 Novel Coronavirus Disease (COVID-19). Clin. Gastroenterol. Hepatol. 2020, 18, 1636–1637. [Google Scholar] [CrossRef]
- Wang, W.; Xu, Y.; Gao, R.; Lu, R.; Han, K.; Wu, G.; Tan, W. Detection of SARS-CoV-2 in Different Types of Clinical Specimens. JAMA 2020, 323, 1843–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heneghan, C.J.; Spencer, E.A.; Brassey, J.; Plüddemann, A.; Onakpoya, I.; Evans, D.H.; Conly, J.; Jefferson, T. SARS-CoV-2 and the role of orofecal transmission: A systematic review. F1000Research 2021, 10, 231. [Google Scholar] [CrossRef]
- Zhang, J.; Garrett, S.; Sun, J. Gastrointestinal symptoms, pathophysiology, and treatment in COVID-19. Genes Dis. 2021, 8, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Troisi, J.; Venutolo, G.; Pujolassos Tanyà, M.; Delli Carri, M.; Landolfi, A.; Fasano, A. COVID-19 and the gastrointestinal tract: Source of infection or merely a target of the inflammatory process following SARS-CoV-2 infection? World J. Gastroenterol. 2021, 27, 1406–1418. [Google Scholar] [CrossRef]
- Zang, R.; Gomez Castro, M.F.; McCune, B.T.; Zeng, Q.; Rothlauf, P.W.; Sonnek, N.M.; Liu, Z.; Brulois, K.F.; Wang, X.; Greenberg, H.B.; et al. TMPRSS2 and TMPRSS4 promote SARS-CoV-2 infection of human small intestinal enterocytes. Sci. Immunol. 2020, 5, eabc3582. [Google Scholar] [CrossRef]
- Xiao, F.; Tang, M.; Zheng, X.; Liu, Y.; Li, X.; Shan, H. Evidence for Gastrointestinal Infection of SARS-CoV-2. Gastroenterology 2020, 158, 1831–1833. [Google Scholar] [CrossRef]
- Leung, W.K.; To, K.F.; Chan, P.K.; Chan, H.L.; Wu, A.K.; Lee, N.; Yuen, K.Y.; Sung, J.J. Enteric involvement of severe acute respiratory syndrome-associated coronavirus infection. Gastroenterology 2003, 125, 1011–1017. [Google Scholar] [CrossRef] [Green Version]
- Wölfel, R.; Corman, V.M.; Guggemos, W.; Seilmaier, M.; Zange, S.; Müller, M.A.; Jones, T.C.; Vollmar, P.; Rothe, C.; Hoelscher, M.; et al. Virological assessment of hospitalized patients with COVID-2019. Nature 2020, 581, 465–469. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Guo, C.; Tang, L.; Hong, Z.; Zhou, J.; Dong, X.; Xiao, Q.; Tang, Y.; Qu, X.; Kuang, L.; et al. Prolonged presence of SARS-CoV-2 viral RNA in faecal samples. Lancet Gastroenterol. Hepatol. 2020, 5, 434–435. [Google Scholar] [CrossRef]
- Xing, Y.H.; Ni, W.; Wu, Q.; Li, W.J.; Li, G.J.; Wang, W.D.; Tong, J.N.; Song, X.F.; Wing-Kin Wong, G.; Xing, Q.S. Prolonged viral shedding in feces of pediatric patients with coronavirus disease 2019. J. Microbiol. Immunol. Infect. 2020, 53, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Blondin-Brosseau, M.; Harlow, J.; Doctor, T.; Nasheri, N. Examining the persistence of human Coronavirus 229E on fresh produce. Food Microbiol. 2021, 98, 103780. [Google Scholar] [CrossRef]
- Giacobbo, A.; Rodrigues, M.A.S.; Zoppas Ferreira, J.; Bernardes, A.M.; de Pinho, M.N. A critical review on SARS-CoV-2 infectivity in water and wastewater. What do we know? Sci. Total. Environ. 2021, 774, 145721. [Google Scholar] [CrossRef]
- O’Brien, B.; Goodridge, L.; Ronholm, J.; Nasheri, N. Exploring the potential of foodborne transmission of respiratory viruses. Food Microbiol. 2021, 95, 103709. [Google Scholar] [CrossRef]
- Mallapaty, S. How sewage could reveal true scale of coronavirus outbreak. Nature 2020, 580, 176–177. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.W.; Li, J.S.; Jin, M.; Zhen, B.; Kong, Q.X.; Song, N.; Xiao, W.J.; Yin, J.; Wei, W.; Wang, G.J.; et al. Study on the resistance of severe acute respiratory syndrome-associated coronavirus. J. Virol. Methods 2005, 126, 171–177. [Google Scholar] [CrossRef]
- Corsi, S.R.; Borchardt, M.A.; Spencer, S.K.; Hughes, P.E.; Baldwin, A.K. Human and bovine viruses in the Milwaukee River watershed: Hydrologically relevant representation and relations with environmental variables. Sci. Total Environ. 2014, 490, 849–860. [Google Scholar] [CrossRef] [Green Version]
- Blanco, A.; Abid, I.; Al-Otaibi, N.; Perez-Rodríguez, F.J.; Fuentes, C.; Guix, S.; Pintó, R.M.; Boschm, A. Glass wool concentration optimization for the detection of enveloped and non-enveloped waterborne viruses. Food Environ. Virol. 2019, 11, 184e192. [Google Scholar] [CrossRef]
- Ahmed, W.; Angel, N.; Edson, J.; Bibby, K.; Bivins, A.; O’Brien, J.W.; Choi, P.M.; Kitajima, M.; Simpson, S.L.; Li, J.; et al. First confirmed detection of SARS-CoV-2 in untreated wastewater in Australia: A proof of concept for the wastewater surveillance of CoVID-19 in the community. Sci. Total Environ. 2020, 728, 138764. [Google Scholar] [CrossRef]
- Medema, G.; Heijnen, L.; Elsinga, G.; Italiaander, R.; Brouwer, A. Presence of SARS-Coronavirus-2 RNA in Sewage and Correlation with Reported COVID-19 Prevalence in the Early Stage of the Epidemic in the Netherlands. Environ. Sci. Technol. Lett. 2020, 7, 511–516. [Google Scholar] [CrossRef]
- Nemudryi, A.; Nemudraia, A.; Wiegand, T.; Surya, K.; Buyukyoruk, M.; Cicha, C.; Vanderwood, K.K.; Wilkinson, R.; Wiedenheft, B. Temporal Detection and Phylogenetic Assessment of SARS-CoV-2 in Municipal Wastewater. Cell Rep. Med. 2020, 1, 100098. [Google Scholar] [CrossRef]
- Wurtzer, S.; Marechal, V.; Mouchel, J.M.; Moulin, L. Time course quantitative detection of SARS-CoV-2 in Parisian wastewaters correlates with COVID-19 confirmed cases. medRxiv 2020. [Google Scholar] [CrossRef]
- Bandala, E.R.; Kruger, B.R.; Cesarino, I.; Leao, A.L.; Wijesiri, B.; Goonetilleke, A. Impacts of COVID-19 pandemic on the wastewater pathway into surface water: A review. Sci. Total Environ. 2021, 774, 145586. [Google Scholar] [CrossRef]
- Center for Disease Control and Infection. Food and Coronavirus Disease 2019 (COVID-19). 2020. Available online: https://www.cdc.gov/coronavirus/2019-ncov/daily-life-coping/food-and-COVID-19.html (accessed on 15 July 2021).
- Sala-Comorera, L.; Reynolds, L.J.; Martin, N.A.; O’Sullivan, J.J.; Meijer, W.G.; Fletcher, N.F. Decay of infectious SARS-CoV-2 and surrogates in aquatic environments. Water Res. 2021, 29, 117090. [Google Scholar] [CrossRef]
- Gundy, P.; Gerba, C.; Pepper, I. Survival of Coronaviruses in Water and Wastewater. Food Environ. Virol. 2009, 1, 10–14. [Google Scholar] [CrossRef] [Green Version]
- Casanova, L.; Rutala, W.A.; Weber, D.J.; Sobsey, M.D. Survival of surrogate coronaviruses in water. Water Res. 2009, 43, 1893–1898. [Google Scholar] [CrossRef]
- Bivins, A.; Greaves, J.; Fischer, R.; Yinda, K.C.; Ahmed, W.; Kitajima, M.; Munster, V.J.; Bibby, K. Persistence of SARS-CoV-2 in Water and Wastewater. Environ. Sci. Technol. Lett. 2020, 7, 937–942. [Google Scholar] [CrossRef]
- De Oliveira, L.C.; Torres-Franco, A.F.; Lopes, B.C.; Santos, B.S.Á.D.S.; Costa, E.A.; Costa, M.S.; Reis, M.T.P.; Melo, M.C.; Polizzi, R.B.; Teixeira, M.M.; et al. Viability of SARS-CoV-2 in river water and wastewater at different temperatures and solids content. Water Res. 2021, 195, 117002. [Google Scholar] [CrossRef]
- World Health Organization; United Nations Children’s Fund (UNICEF). Water, Sanitation, Hygiene, and Waste Management for the COVID-19 Virus: Interim Guidance, 19 March 2020; World Health Organization: Geneva, Switzerland, 2020; Available online: https://apps.who.int/iris/handle/10665/331499 (accessed on 15 July 2021).
- Geller, C.; Varbanov, M.; Duval, R.E. Human coronaviruses: Insights into environmental resistance and its influence on the development of new antiseptic strategies. Viruses 2012, 4, 3044–3068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organisation. WHO Guidelines for the Global Surveillance of Severe Acute Respiratory Syndrome (SARS); WHO: Geneva, Switzerland, 2004; Available online: https://www.who.int/csr/resources/publications/WHO_CDS_CSR_ARO_2004_1.pdf?ua=1 (accessed on 15 July 2021).
- Todd, E.; Grieg, J. Viruses of foodborne origin: A review. Virus Adapt. Treat. 2015, 7, 25–45. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Li, C.; Zhao, G.; Chu, H.; Wang, D.; Yan, H.H.-N.; Poon, V.K.; Wen, L.; Wong, B.H.; Zhao, X.; et al. Human intestinal tract serves as an alternative infection route for Middle East respiratory syndrome coronavirus. Sci. Adv. 2017, 3, eaao4966. [Google Scholar] [CrossRef] [Green Version]
- Duda-Chodak, A.; Lukasiewicz, M.; Zięć, G.; Florkiewicz, A.; Filipiak-Florkiewicz, A. COVID-19 pandemic and food: Present knowledge, risks, consumers fears and safety. Trends Food Sci. Technol. 2020, 105, 145–160. [Google Scholar] [CrossRef]
- Hartman, A.L.; Nambulli, S.; McMillen, C.M.; White, A.G.; Tilston-Lunel, N.L.; Albe, J.R.; Cottle, E.; Dunn, M.D.; Frye, L.J.; Gilliland, T.H.; et al. SARS-CoV-2 infection of African green monkeys results in mild respiratory disease discernible by PET/CT imaging and shedding of infectious virus from both respiratory and gastrointestinal tracts. PLoS Pathog. 2020, 16, e1008903. [Google Scholar] [CrossRef]
- Lee, A.C.; Zhang, A.J.; Chan, J.F.; Li, C.; Fan, Z.; Liu, F.; Chenm, Y.; Liangm, R.; Sridhar, S.; Cai, J.P.; et al. Oral SARS-CoV-2 Inoculation Establishes Subclinical Respiratory Infection with Virus Shedding in Golden Syrian Hamsters. Cell Rep. Med. 2020, 1, 100121. [Google Scholar] [CrossRef]
- Todd, E.C.; Greig, J.D.; Bartleson, C.A.; Michaels, B.S. Outbreaks where food workers have been implicated in the spread of foodborne disease. Part 6. Transmission and survival of pathogens in the food processing and preparation environment. J. Food Prot. 2009, 72, 202–219. [Google Scholar] [CrossRef]
- Bachofen, C. Selected Viruses Detected on and in our Food. Curr. Clin. Micro. Rpt. 2018, 5, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Nasheri, N.; Vester, A.; Petronella, N. Foodborne viral outbreaks associated with frozen produce. Epidemiol. Infect. 2019, 147, E291. [Google Scholar] [CrossRef] [PubMed]
- Rönnqvist, M.; Maunula, L. Noroviruses on surfaces: Detection, persistence, disinfection and role in environmental transmission. Future Virol. 2016, 11, 207–217. [Google Scholar] [CrossRef]
- Lodder, W.; de Roda Husman, A. SARS-CoV-2 in wastewater: Potential health risk, but also data source. Lancet Gastroenterol. Hepatol. 2020, 1253, 533–534. [Google Scholar] [CrossRef]
- Yeo, C.; Kaushal, S.; Yeo, D. Enteric involvement of coronaviruses: Is faecal–oral transmission of SARS-CoV-2 possible? Lancet Gastroenterol. Hepatol. 2020, 5, 335–337. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Gao, J.; Yao, L.; Zeng, L.; Liu, Q.; Zhou, Q.; Zhang, H.; Lu, D.; Fu, J.; Liu, Q.S.; et al. Evidence of foodborne transmission of the coronavirus (COVID-19) through the animal products food supply chain. Environ. Sci. Technol. 2021, 55, 2713–2716. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.-D.; Wang, Z.-Y.; Zhang, S.-F.; Li, X.; Li, L.; Li, C.; Cui, Y.; Fu, R.B.; Dong, Y.Z.; Chi, X.Y.; et al. Aerosol and surface distribution of severe acute respiratory syndrome coronavirus 2 in hospital Wards, Wuhan, China, 2020. Emerg. Infect. Dis. 2020, 26, 1583–1591. [Google Scholar] [CrossRef]
- McDermott, C.V.; Alicic, R.Z.; Harden, N.; Cox, E.J.; Scanlan, J.M. Put a lid on it: Are faecal bio-aerosols a route of transmission for SARS-CoV-2? J. Hosp. Infect. 2020, 105, 397–398. [Google Scholar] [CrossRef]
- Wiktorczyk-Kapischke, N.; Grudlewska-Buda, K.; Wałecka-Zacharska, E.; Kwiecińska-Piróg, J.; Radtke, L.; Gospodarek-Komkowska, E.; Skowron, K. SARS-CoV-2 in the environment-Non-droplet spreading routes. Sci. Total Environ. 2021, 770, 145260. [Google Scholar] [CrossRef] [PubMed]
- Chin, A.W.H.; Chu, J.T.S.; Perera, M.R.A.; Hui, K.P.Y.; Yen, H.-L.; Chan, M.C.W.; Peiris, M.; Poon, L.L.M. Stability of SARS-CoV-2 in different environmental conditions. Lancet Microbe 2020, 1, e10. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Bushmaker, T.; Morris, D.H.; Holbrook, M.G.; Gamble, A.; Williamson, B.N.; Tamin, A.; Harcourt, J.L.; Thornburg, N.J.; Gerber, S.I.; et al. Aerosol and surface stability of SARS-CoV-2 as compared with SARS-CoV-1. N. Engl. J. Med. 2020, 382, 1564–1567. [Google Scholar] [CrossRef]
- Kasloff, S.B.; Leung, A.; Strong, J.E.; Funk, D.; Cutts, T. Stability of SARS-CoV-2 on critical personal protective equipment. Sci. Rep. 2021, 11, 984. [Google Scholar] [CrossRef]
- Sizun, J.; Yu, M.W.; Talbot, P. Survival of human coronaviruses 229E and OC43 in suspension and after drying on surfaces: A possible source of hospital-acquired infections. J. Hosp. Infect. 2000, 46, 55–60. [Google Scholar] [CrossRef]
- Duan, S.M.; Zhao, X.S.; Wen, R.F.; Huang, J.J.; Pi, G.H.; Zhang, S.X.; Han, J.; Bi, S.L.; Ruan, L.; Dong, X.P. Stability of SARS coronavirus in human specimens and environment and its sensitivity to heating and UV irradiation. Biomed. Environ. Sci. 2003, 16, 246–255. [Google Scholar]
- Lai, M.Y.; Cheng, P.K.; Lim, W.W. Survival of severe acute respiratory syndrome coronavirus. Clin. Infect. Dis. 2005, 41, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Hirose, R.; Ikegaya, H.; Naito, Y.; Watanabe, N.; Yoshida, T.; Bandou, R.; Daidoji, T.; Itoh, Y.; Nakaya, T. Survival of SARS-CoV-2 and influenza virus on the human skin: Importance of hand hygiene in COVID-19. Clin. Infect. Dis. 2020, ciaa1517. [Google Scholar] [CrossRef] [PubMed]
- Riddell, S.; Goldie, S.; Hill, A.; Eagles, D.; Drew, T. The effect of temperature on persistence of SARS-CoV-2 on common surfaces. Virol. J. 2020, 17, 145. [Google Scholar] [CrossRef] [PubMed]
- Rabenau, H.F.; Cinatl, J.; Morgenstern, B.; Bauer, G.; Preiser, W.; Doerr, H.W. Stability and inactivation of SARS coronavirus. Med. Microbiol. Immunol. 2005, 194, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.H.; Peiris, J.S.; Lam, S.Y.; Poon, L.L.; Yuen, K.Y.; Seto, W.H. The Effects of Temperature and Relative Humidity on the Viability of the SARS Coronavirus. Adv. Virol. 2011, 2011, 734690. [Google Scholar] [CrossRef]
- Van Doremalen, N.; Bushmaker, T.; Munster, V.J. Stability of Middle East respiratory syndrome coronavirus (MERS-CoV) under different environmental conditions. Euro Surveill. 2013, 18, 20590. [Google Scholar] [CrossRef] [Green Version]
- Warnes, S.L.; Little, Z.R.; Keevil, C.W. Human coronavirus 229E remains infectious on common touch surface materials. MBio 2015, 6, e01697-15. [Google Scholar] [CrossRef] [Green Version]
- Pastorino, B.; Touret, F.; Gilles, M.; de Lamballerie, X.; Charrel, R.N. Prolonged Infectivity of SARS-CoV-2 in Fomites. Emerg. Infect. Dis. 2020, 26, 2256–2257. [Google Scholar] [CrossRef]
- Liu, Y.; Li, T.; Deng, Y.; Liu, S.; Zhang, D.; Li, H.; Wang, X.; Jia, L.; Han, J.; Bei, Z.; et al. Stability of SARS-CoV-2 on environmental surfaces and in human excreta. J. Hosp. Infect. 2021, 107, 105–107. [Google Scholar] [CrossRef] [PubMed]
- Thippareddi, H.; Balamurugan, S.; Patel, J.; Singh, M.; Brassard, J. Coronaviruses—Potential human threat from foodborne transmission? Lebensm. Wiss. Technol. 2020, 31, 110147. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.W.; Li, J.; Guo, T.; Zhen, B.; Kong, Q.; Yi, B.; Li, Z.; Song, N.; Jin, M.; Xiao, W.; et al. Concentration and detection of SARS coronavirus in sewage from Xiao Tang Shan hospital and the 309th Hospital of the Chinese People’s Liberation Army. Water Sci. Technol. 2005, 52, 213–221. [Google Scholar] [CrossRef]
- Kampf, G.; Todt, D.; Pfaender, S.; Steinmann, E. Persistence of coronaviruses on inanimate surfaces and their inactivation with biocidal agents. J. Hosp. Infect. 2020, 104, 246–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand-Moreau, Q.A.A.; Mackenzie, G.; Bowley, J.; Straube, S.; Chan, X.H.; Zelyas, N.; Greenhalgh, T. COVID-19 in Meat and Poultry Facilities: A Rapid Review and Lay Media Analysis. 2020. Available online: https://www.cebm.net/covid-19/what-explains-the-high-rate-of-sars-cov-2-transmission-in-meat-and-poultry-facilities-2/ (accessed on 15 July 2021).
- Ursachi, C.Ș.; Munteanu, F.; Cioca, G. The safety of slaughterhouse workers during the pandemic crisis. Int. J. Environ. Res. Public Health 2021, 18, 2633. [Google Scholar] [CrossRef] [PubMed]
- Bundesinstitut für Risikobewertung (BfR). Can the New Type of Coronavirus Be Transmitted via Food and Objects? Available online: https://www.bfr.bund.de/cm/349/can-the-new-type-of-coronavirus-be-transmitted-via-food-and-objects.pdf (accessed on 15 July 2021).
- Galanakis, C.M. The Food Systems in the Era of the Coronavirus (COVID-19) Pandemic Crisis. Foods 2020, 9, 523. [Google Scholar] [CrossRef]
- Yépiz-Gómez, M.S.; Gerba, C.P.; Bright, K.R. Survival of Respiratory Viruses on Fresh Produce. Food Environ. Virol. 2013, 5, 150–156. [Google Scholar] [CrossRef]
- Conway, M. Identification of coronavirus sequences in carp cDNA from Wuhan, China. J. Med. Virol. 2020, 92, 1629–1633. [Google Scholar] [CrossRef]
- Liu, R.; Stanway, D. Traces of Coronavirus Found in Frozen Chicken Wings, Shrimp Packaging in China, Global News. Available online: https://globalnews.ca/news/7271588/coronavirus-traces-frozen-chicken-shrimp-packaging/ (accessed on 15 July 2021).
- Yusha, Z. China’s CDC Experts Investigate Xinfadi Market Three Times, Announce Groundbreaking Virus Tracing Discovery, Global Times. Available online: https://www.globaltimes.cn/content/1192146.shtml (accessed on 15 July 2021).
- Liu, P.; Yang, M.; Zhao, X.; Guo, Y.; Wang, L.; Zhang, J.; Lei, W.; Han, W.; Jiang, F.; Liu, W.J.; et al. Cold-chain transportation in the frozen food industry may have caused a recurrence of COVID-19 cases in destination: Successful isolation of SARS-CoV-2 virus from the imported frozen cod package surface. Biosaf. Health. 2020, 2, 199–201. [Google Scholar] [CrossRef]
- Pang, X.; Ren, L.; Wu, S.; Ma, W.; Yang, J.; Di, L.; Li, J.; Xiao, Y.; Kang, L.; Du, S.; et al. Cold-chain food contamination as the possible origin of Covid-19 resurgence in Beijing. Natl. Sci. Rev. 2020, 7, 1861–1864. [Google Scholar] [CrossRef]
- Han, Y.; Duan, X.; Yang, L.; Nilsson-Payant, B.; Wang, P.; Duan, F.; Yaron, T.M.; Zhang, T.; Uhl, S.; Bram, Y.; et al. Identification of SARS-CoV-2 inhibitors using lung and colonic organoids. Nature 2021, 589, 270–275. [Google Scholar] [CrossRef]
- Potasman, I.; Paz, A.; Odeh, M. Infectious Outbreaks Associated with Bivalve Shellfish Consumption: A Worldwide Perspective. Clin. Infect. Dis. 2002, 35, 921–928. [Google Scholar] [CrossRef] [Green Version]
- French Agency for Food, Environmental and Occupational Health & Safety (ANSES). Supplemented Opinion of 9 March 2020 of the French Agency for Food, Environmental and Occupational Health & Safety. 2020. Available online: https://www.anses.fr/en/system/files/SABA2020SA0037EN-1.pdf (accessed on 15 July 2021).
- Smith, J.L. The role of gastric acid in preventing foodborne disease and how bacteria overcome acid conditions. J. Food Prot. 2003, 66, 1292–1303. [Google Scholar] [CrossRef]
- Hellenic Food Authority (EFET). Guide for the Restart of Foodservice. Available online: http://efet.gr/files/pdf/other/odigos-epan-estiasis.pdf (accessed on 15 July 2021). In Greek.
- Okumus, B. Norovirus and Coronavirus Risks in Food Service Settings: A Systematic Review for Future Research. J. Culin. Sci. Technol. 2021. [Google Scholar] [CrossRef]
Figure 1.
Average time values (h) needed for 99.9% inactivation of CoVs (blue bars) and SARS-CoV-2 (orange bars) on porous surfaces (error bars depicting standard deviation). (Calculated according to results reported by Sizun et al., 2000 [112]; Duan et al., 2003 [113]; Lai et al., 2005 [114]; Chin et al., 2020 [109]; Hirose et al., 2020 [115]; Riddell et al., 2020 [116]; and Van Doremalen et al., 2020 [110]).
Figure 1.
Average time values (h) needed for 99.9% inactivation of CoVs (blue bars) and SARS-CoV-2 (orange bars) on porous surfaces (error bars depicting standard deviation). (Calculated according to results reported by Sizun et al., 2000 [112]; Duan et al., 2003 [113]; Lai et al., 2005 [114]; Chin et al., 2020 [109]; Hirose et al., 2020 [115]; Riddell et al., 2020 [116]; and Van Doremalen et al., 2020 [110]).
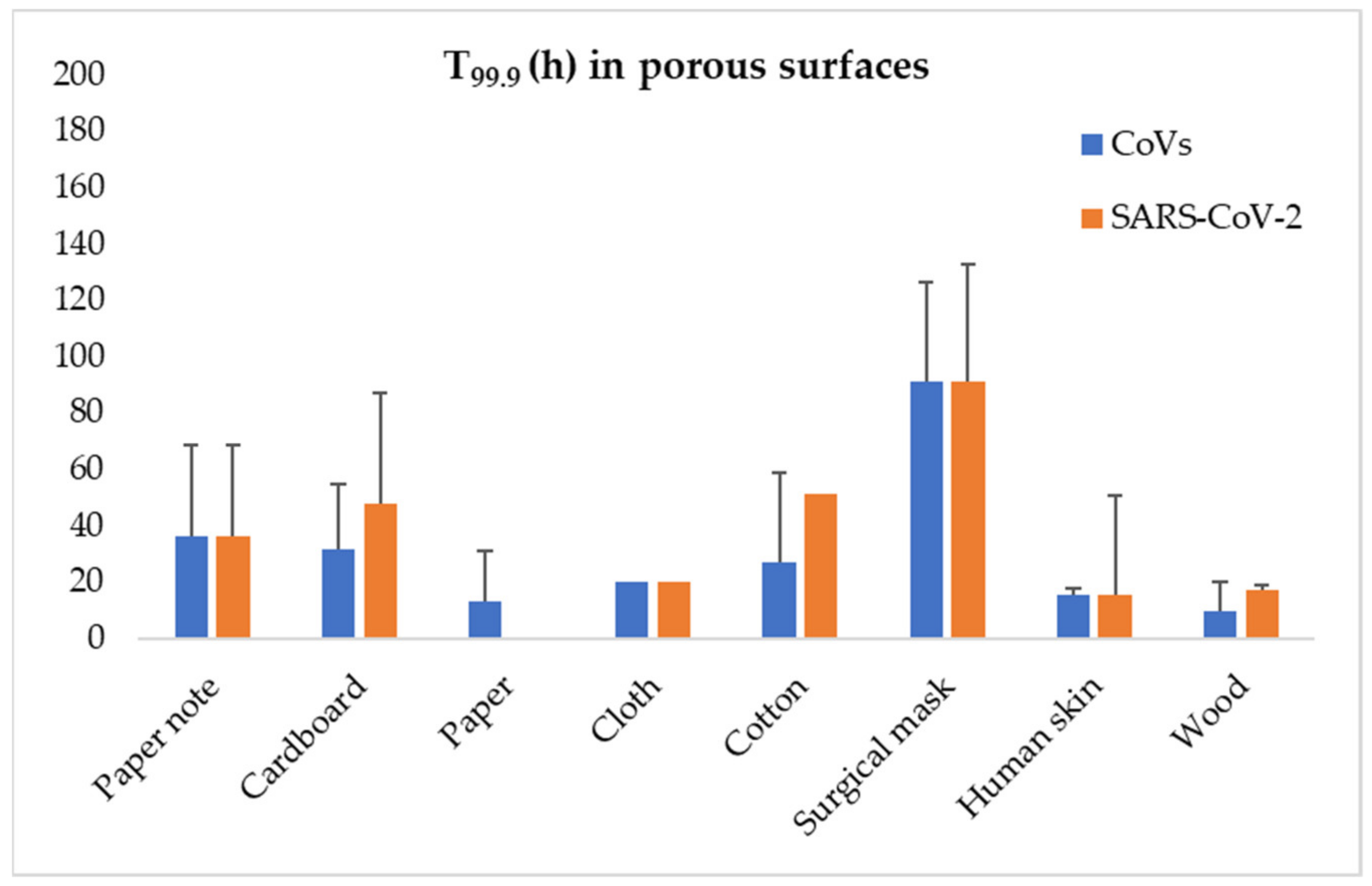
Figure 2.
Average values of time needed for 99.9% inactivation of CoVs (blue bars) and SARS-CoV-2 (orange bars) on non-porous surfaces (error bars depicting standard deviation). (Calculated according to results reported by Sizun et al., 2000 [112]; Duan et al., 2003 [113]; Rabenau et al., 2005 [117]; Chan et al., 2011 [118]; Van Doremalen et al., 2013 [119]; Warnes et al., 2015 [120]; Chin et al., 2020 [109]; Hirose et al., 2020 [115]; Pastorino et al., 2020 [121]; Riddell et al., 2020 [116]; Van Doremalen et al., 2020 [110]; and Liu et al., 2021 [122]).
Figure 2.
Average values of time needed for 99.9% inactivation of CoVs (blue bars) and SARS-CoV-2 (orange bars) on non-porous surfaces (error bars depicting standard deviation). (Calculated according to results reported by Sizun et al., 2000 [112]; Duan et al., 2003 [113]; Rabenau et al., 2005 [117]; Chan et al., 2011 [118]; Van Doremalen et al., 2013 [119]; Warnes et al., 2015 [120]; Chin et al., 2020 [109]; Hirose et al., 2020 [115]; Pastorino et al., 2020 [121]; Riddell et al., 2020 [116]; Van Doremalen et al., 2020 [110]; and Liu et al., 2021 [122]).

{kind=link}
{kind=link}
| CoV | Genus | Animal Pool | Intermediate Host | Primary Receptor | Common Transmission | Faecal Shedding | Foodborne Transmission |
|---|---|---|---|---|---|---|---|
| HCoV-229E | alpha-CoV | Bats | Camelids? | Human Aminopeptidase N (CD13) | Respiratory droplets, fomites | ||
| HCoV-OC43 | beta-CoV, lineage A | Rodents | Bovines | 9-O-Acetylated sialic acid | Respiratory droplets, fomites | ||
| SARS-CoV | beta-CoV, lineage B | Palm Civets, Bats | Palm civets | Angiotensin-converting enzyme 2 (ACE2) | Respiratory droplets, fomites, faecal—oral | Yes | Yes |
| HCoV-NL63 | alpha-CoV | Palm Civets, Bats | ? | Angiotensin-converting enzyme 2 (ACE2) | Respiratory droplets, fomites | ||
| HCoV-HKU1 | beta-CoV, lineage A | Rodents | 9-O-Acetylated sialic acid | Respiratory droplets, fomites | |||
| MERS-CoV | beta-CoV, lineage C | Bats, Camels | Dromedary camels | Dipeptidyl peptidase-4 (DPP4) | Respiratory droplets, fomites | Yes | Yes |
| SARS-CoV-2 | beta-CoV, lineage B | Bats | Pangolins? | Angiotensin-converting enzyme 2 (ACE2) | Respiratory droplets, fomites, faecal—oral | Yes | ? |
Table 2.
Coronaviruses survival in water.
| Reference | CoV | Water Characteristics | Reduction | |||||
|---|---|---|---|---|---|---|---|---|
| Type | T | pH | Turbidity | 99.9% | 99% | 90% | ||
| Casanova et al., 2009 [88] | TGEV | reagent-grade water | 25 °C | 6 | 0.1 NTU | 33 d | 22 d | 11 d |
| MHV | 26 d | 17 d | 9 d | |||||
| TGEV | 4 °C | 330 d | 220 d | 110 d | ||||
| MHV | >365 d | |||||||
| TGEV | lake water | 25 °C | 7.5 | 1.73 NTU | 13 d | |||
| MHV | 10 d | |||||||
| TGEV | 4 °C | 14 d | ||||||
| MHV | >14 d | |||||||
| Gundy et al., 2019 [87] | HCoV 229E (ATCC-740) | Tap water filtered | 23 °C | 7.8 | 10.1 d | 6.76 d | ||
| FIPV (ATCC-990) | 10.1 d | 6.76 d | ||||||
| HCoV 229E (ATCC-740) | 4 °C | 588 d | 392 d | |||||
| FIPV (ATCC-990) | 130 d | 87 d | ||||||
| HCoV 229E (ATCC-740) | Tap water unfiltered | 23 °C | 12.1 d | 8.09 d | ||||
| FIPV (ATCC-990) | 12.5 d | 8.32 d | ||||||
| Bivins et al., 2020 [89] | SARS-CoV-2 (nCoV-WA1-2020) 10 5 and 10 3 TCID50 *mL−1 | wastewater | 20 °C | 7.98 | 1.6–2.1 d | |||
| tap water | 2.0 d | |||||||
| De Oliveira et al., 2021 [90] | SARS-CoV-2 (SARS.CoV2/SP02. 2020.HIAE.Br) 2 × 10 4 PFU/mL | River water | 24 °C | 5.5 | 10 NTU | 6.4 d | 1.9 d | |
| 4 °C | 18.7 d | 7.7 d | ||||||
| Filtered river water | 24 °C | 1 NTU | 8.5 d | 3.3 d | ||||
| Sala-Comorera et al., 2021 [86] | SARS-CoV-2 3.16 × 10 4 TCID50 *mL−1 | River water | 20 °C | 11.3 d * | 7.5 d * | 3.8 d * | ||
| 4 °C | 11.2 d * | 7.5 d * | 3.7 d * | |||||
| Seawater | 20 °C | 10.5 d * | 7.0 d * | 3.5 d * | ||||
| 4 °C | 5.2 d * | 3.5 d * | 1.7 d * | |||||
* values calculated according to the reported decay rates k.
Table 3.
Coronaviruses survival in wastewater.
| Reference | CoV | Water Characteristics | Reduction | Survival | |||||
|---|---|---|---|---|---|---|---|---|---|
| Type | T | pH | Turbidity | 99.9% | 99% | 90% | |||
| Wang et al., 2005 [77] | SARS-COV-1 | Sewage | 20 °C | 2 d | |||||
| 4 °C | 14 d | ||||||||
| Casanova et al., 2009 [88] | TGEV | Pasteurised sewage | 25 °C | 7.6 | 17.6 NTU | 14 d | 9 d | 4 d | |
| MHV | 10 d | 7 d | 3 d | ||||||
| TGEV | 4 °C | 73 d | 49 d | 24 d | |||||
| MHV | 105 d | 70 d | 35 d | ||||||
| Ye et al., 2016 [3] | MHV A59 | Wastewater | 25 °C | 13 h | |||||
| 10 °C | 36 h | ||||||||
| Pasteurized wastewater | 25 °C | 19 h | |||||||
| Gundy et al., 2019 [87] | HCoV 229E (ATCC-740) | Primary effluent unfiltered | 10 °C | 3.54 d | 2.36 d | ||||
| FIPV (ATCC-990) | 2.56 d | 1.71 d | |||||||
| HCoV 229E (ATCC-740) | Primary effluent filtered | 2.35 d | 1.57 d | ||||||
| FIPV (ATCC-990) | 2.4 d | 1.6 d | |||||||
| HCoV 229E (ATCC-740) | Secondary effluent | 2.77 d | 1.85 d | ||||||
| FIPV (ATCC-990) | 2.42 d | 1.62 d | |||||||
| De Oliveira et al., 2021 [90] | SARS-CoV-2 (SARS.CoV2/SP02. 2020.HIAE.Br) 2 × 10 4 PFU/mL | Wastewater | 24 °C | 7.5 | 274 NTU | 4.0 d | 1.2 d | ||
| 4 °C | 17.5 d | 5.5 d | |||||||
| Filtered wastewater | 24 °C | 6 NTU | 4.5 d | 1.5 d | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Economou, V.; Sakkas, H.; Bezirtzoglou, E.; Papa, A.; Soultos, N. SARS–CoV–2 and Food—How Confident Are We about Them? Hygiene 2021, 1, 80-98. https://0-doi-org.brum.beds.ac.uk/10.3390/hygiene1030008
AMA Style
Economou V, Sakkas H, Bezirtzoglou E, Papa A, Soultos N. SARS–CoV–2 and Food—How Confident Are We about Them? Hygiene. 2021; 1(3):80-98. https://0-doi-org.brum.beds.ac.uk/10.3390/hygiene1030008
Chicago/Turabian StyleEconomou, Vangelis, Hercules Sakkas, Eugenia Bezirtzoglou, Anna Papa, and Nikolaos Soultos. 2021. "SARS–CoV–2 and Food—How Confident Are We about Them?" Hygiene 1, no. 3: 80-98. https://0-doi-org.brum.beds.ac.uk/10.3390/hygiene1030008