
Oxygen Is Instrumental for Biological Signaling: An Overview


Department of Applied Sciences, University of the West of England, Bristol BS16 1QY, UK
Oxygen 2021, 1(1), 3-15; https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen1010002
Submission received: 10 June 2021
/
Revised: 5 July 2021
/
Accepted: 7 July 2021
/
Published: 12 July 2021
{kind=link}
Abstract
:Control of cellular function is extremely complex, being reliant on a wide range of components. Several of these are small oxygen-based molecules. Although reactive compounds containing oxygen are usually harmful to cells when accumulated to relatively high concentrations, they are also instrumental in the control of the activity of a myriad of proteins, and control both the upregulation and downregulation of gene expression. The formation of one oxygen-based molecule, such as the superoxide anion, can lead to a cascade of downstream generation of others, such as hydrogen peroxide (H2O2) and the hydroxyl radical (∙OH), each with their own reactivity and effect. Nitrogen-based signaling molecules also contain oxygen, and include nitric oxide (NO) and peroxynitrite, both instrumental among the suite of cell signaling components. These molecules do not act alone, but form part of a complex interplay of reactions, including with several sulfur-based compounds, such as glutathione and hydrogen sulfide (H2S). Overaccumulation of oxygen-based reactive compounds may alter the redox status of the cell and lead to programmed cell death, in processes referred to as oxidative stress, or nitrosative stress (for nitrogen-based molecules). Here, an overview of the main oxygen-based molecules involved, and the ramifications of their production, is given.
1. Introduction
Oxygen-based compounds are an instrumental part of the group of small, relatively reactive molecules which control cellular activities. Traditionally such molecules have been referred to as the reactive oxygen species (ROS) and include hydrogen peroxide (H2O2), superoxide (O2∙−), and hydroxyl radicals (∙OH). However, several other reactive signaling molecules also contain oxygen, although referred to as reactive nitrogen species (RNS). These include nitric oxide (NO) and peroxynitrite (ONOO−), and therefore could be grouped together with the ROS as oxygen-based compounds. As discussed below, such compounds often work together [1].
Historically, ROS were studied in biological systems as they are produced during a pathogen challenge, and it was suggested that their reactivity was harnessed to kill the invading organism [2]. Studies concentrated on the production of ROS in phagocytic cells in animals, especially neutrophils, and the enzyme NADPH oxidase was characterized [3]. This was aided by the realization that patients with Chronic Granulomatous Disease (CDG) had an impaired ROS generation and reduced pathogen tolerance. Interestingly, CGD can be inherited in both a X-linked and autosomal fashion, enabling the different NADPH oxidase components to be discovered [4].
In 1987, it was reported that endothelial-derived relaxing factor (EDRF) was in fact the gas NO [5]. Although this was not the first work on this gas in a biological setting, for example [6], the 1987 paper did focus researchers’ efforts, and it was soon realized that other reactive compounds could partake in similar activities. NADPH oxidase homologues were reported in a range of cells suggesting a role in cell signaling reviewed in [7]. Superoxide anions were to an extent ruled out as they were charged and deemed not able to pass through membranes, although once protonated this is not the case. Focus fell on H2O2, and ever since there has been an explosion of papers on this topic reviewed [8,9]. However, the efforts did not stop there, and now there are papers showing that a range of small molecules can partake in cell signaling events in an array of organisms. These molecules include others which contain oxygen, such as ONOO− [10] and carbon monoxide (CO) [11], but also others which do not contain oxygen, including reactive sulfur species (RSS), such as hydrogen sulfide (H2S) [12], and hydrogen gas (H2) [13].
None of these molecules act in cell signaling events in isolation, and the interplay between them has been quite extensively reviewed [14,15,16]. There is a complex interplay between them which creates downstream signals. As will be discussed below, many of these molecules are in competition with each other, potentially reacting with the same amino acid groups, such as thiols. It can, therefore, be seen that oxygen-based small molecules play a key part in the regulation of cellular function in a wide range of organisms.
2. Signaling by ROS
ROS can be produced from a range of places in cells. Oxygen, a diradical itself, can scavenge “leaked” electrons from metabolic pathways, such as the electron transport chain (ETC) in mitochondria [17,18]. The one electron reduction of O2 will yield O2∙−, but this is relatively unstable and will readily dismute to H2O2 [19], especially in the presence of protons, i.e., low pH. Complexes I and III appear to be the primary sources of mitochondrial ROS [17,18]. However, other enzymes can generate ROS too. One of the main sources of cellular ROS is from the NADPH oxidases [20]. There are a family of such enzymes in animals, and homologues in plants, known as respiratory burst oxidase homologs (RBOH) [21,22]. Although originally characterized from neutrophils, it was realized that there were isoforms in a range of cells, and then further forms were found, such as the DUOX proteins [23]. Each oxidase isoform will have different roles, locations, control and kinetics, as briefly discussed below. Arabidopsis, for example, has ten RBOH isoforms [22].
Other enzymes can produce ROS too. These include the peroxidases [24] and xanthine oxidase (xanthine oxidoreductase) [25]. This latter enzyme can, when oxygen is not readily available, produce nitric oxide too [26].
Therefore, in cells ROS are going to be present, and it is now known that they have signaling roles. One of the primary times that cells produce ROS is when they are under stress, either from biotic or abiotic mechanisms [27]. This could be from a pathogen attack, or in the presence of extreme temperature, heavy metals, too much salt, or too much light. Note that not only does this apply to plants, but also to animals, however stress responses usually see a generation of other reactive signals too, such as RNS [28]. Therefore, under such conditions, ROS, RNS, along with H2S are all likely to be present together, allowing both their competition for response mechanisms, as well as their direct interactions, as discuss further below.
As well as being involved in normal physiology, and in the management of stress, ROS are also thought to be instrumental in the aging process [29,30]. This theory was mooted by Harman fifty years ago [31], but the idea still has traction [32] and shows that the production and action of these oxygen-based compounds are an integral part of life, and death, on earth.
2.1. Superoxide and Its Role
Superoxide anions (O2∙−) will be produced by the one electron reduction of molecular oxygen. The added electron leads to the molecule being both charged and a free radical, and therefore it is relatively reactive [33]. Dismutation is likely in biological systems [19] and is catalyzed by superoxide dismutases (SOD) [34], producing H2O2. However, superoxide can be measured, and early work with neutrophils assayed O2∙− by the reduction of cytochrome c in the presence and absence of SOD, with a similar technique still being employed [35].
One of the main sources of O2∙− in cells is the family of NADPH oxidases [20,36,37]. The oxidase from neutrophils was found to use NADPH as a cofactor, and on its oxidation, the electrons are sequentially passed to flavin, heme, and then oxygen. The enzyme had several subunits, including two in the membrane (gp91-phox and p22-phox) and several in the cytoplasm, which translocate to form a holistic enzyme. Among the cytoplasmic subunits is a G protein, although phosphorylation seems to also be important as part of the control mechanisms. In humans there are seven members of the NAPDH oxidase protein family. These are Nox1-5 and Duox 1-2. While most produce O2∙−, some produce H2O2. Other sources of O2∙−, as mentioned, include electron leakage from redox pathways such as the ETC [17,18].
In signaling terms, the charge on the superoxide anion was thought to limit its movement in cells and therefore its effectiveness as a cellular signal. However, it can become protonated (HO2), removing the charge and therefore trans-membrane movement is potentially possible. In this vein, signaling is known to be mediated by superoxide anions [38]. For example, O2∙− generation from mitochondrial Complex III mediates the hypoxia inducible factor (HIF)-1α signaling pathway, which is part of the hypoxic response of cells [39]. HIF acts as a key oxygen sensor in cells, ensuring optimal ATP production through a complex interplay with ROS metabolism [40].
It has to be remembered too, that O2∙− is a redox molecule. The redox mid-point potential of the O2/O2∙− couple has been estimated to be −160 mV relative to the Standard Hydrogen Electrode (SHE) [41], and this will contribute to the overall cellular redox. It has been discussed previously [42] that all the redox-active molecules will contribute to the cellular redox and downstream control of cellular activities, so O2∙− will be part of this, especially if the generation of superoxide is compartmentalized, as has been suggested for ROS signaling and other redox signaling [43,44,45].
It is very difficult to separate the signaling effects of O2∙− from that of H2O2. It is often assumed that the presence of O2∙− gives rise to H2O2 and it is the latter that has assumed the signaling role.
2.2. Hydrogen Peroxide as a Signal
The sequential oxidation of molecule oxygen produces O2∙−, then H2O2, and finally the hydroxyl radical (∙OH) before the four election reduction results in water. Therefore, once the O2∙− anion is formed, a cascade of further products is likely. As discussed below, there are side reactions likely here too. For example, hypochlorous acid can be produced in the presence of the enzyme myeloperoxidase [46]. However, when discussing ROS signaling, H2O2 always rises to prominence.
As well as arising from the dismutation of O2∙−, either spontaneously or catalyzed by SOD [34], H2O2 can be generated by enzymes such as XO and peroxidases [47]. H2O2 is not charged and can easily translocate across lipid membranes, so is not likely to be compartmentalized in organelles unless it is removed before it can diffuse. Removal will be by its interaction with antioxidant biomolecules, as well as by catalysis by enzymes such as catalase (Cat) [48]. Other systems will also be involved in H2O2 removal, including peroxiredoxin [49] and glutathione peroxidase [50].
H2O2 has been the focus of ROS signaling [8,51,52]. One of the ways in which H2O2 is known to alter cell function is by the oxidation of thiol groups in proteins [52], and such modifications can be analyzed by proteomic techniques [53,54]. The -SH group is converted to the sulfenic acid group, -SOH. This is in many ways akin to phosphorylation, and like phosphorylation, the formation of the -SOH group is likely to force a conformational change on the proteins and thus alter its activity. This is not necessarily activation. In tyrosine phosphatase, the interaction with H2O2 leads to the formation of a sulfenyl-amide intermediate and inhibition of the enzyme [55]. This means in the cell that the levels of tyrosine phosphorylation are likely to increase, with the concomitant effects that leads to.
Oxidation of the thiol can continue, with the sequential formation of the sulfinic acid group and then sulfonic acid. The latter modification is thought to be irreversible and fixes the protein in a new conformation, and probably leads to protein removal, or if the H2O2 levels are high then cell death may result.
Enzymes that are modified by H2O2 include those which are involved in metabolism, such as glyceraldehyde 3-phosphate dehydrogenase (GAPDH) [56]. Modification of this protein can control its cellular location and hence function [57], showing that it acts like a moonlighting protein. Other proteins modified include the transcription factors [51], and hence gene expression of target genes may be increased or decreased, depending on the gene involved [58]. Proteins involved here include nuclear factor kappa B (NF-κB) [59] and nuclear factor erythroid 2-related factor 2 (nrf2) [60], although others are also involved [61]. Downstream of H2O2 kinase pathways can also be affected [62], particularly mitogen-activated protein kinases (MAPKs). However, there are many more proteins which can be affected by H2O2, in a variety of cellular locations. H2O2 signaling seem to be universal across cell types and species, from plants to animals. Such studied have been reviewed by others [63,64,65].
It is not only proteins which act as targets for H2O2, lipid peroxidation is often key to the cellular effects seen [66] and is often part of the mechanism, which is harmful to the cell [67]. In fact, H2O2 is part of the programmed cell death process [68], controlling apoptosis for example [69].
As with all redox active molecules in cells, their presence and activity influence the overall redox poise of the cell [42]. H2O2 is no exception here, and it is thought to be one of the main influencers. The overaccumulation of H2O2 will lead to an oxidation of the cellular redox and this is termed oxidative stress [9,70]. It is always deemed to be detrimental and it has been argued that it can lead from normal cell function, to over proliferation and eventually cell death, either from apoptosis or necrosis [42]. One of the main buffers of redox stress is glutathione, which can be oxidized from the GSH state to the GSSG form. The ratio of GSH:GSSG is often used to calculate the intracellular redox, and the concentration of H2O2 in cells will have a direct influence on this. Other small thiol-based compounds are also involved, including cysteine (Cys), cysteinyl-glycine (Cys-Gly), and γ-glutamyl-cysteine (γ-Glu-Cys), and it has been suggested that it is all of these that need to be considered [42].
However, oxidative stress is part of balancing act. It has been argued that redox has a “Goldilocks zone”, where there are defined limits between which the redox of the cell need to be held [71]. As well as oxidative stress, the opposite is now being recognized, i.e., reductive stress [72,73], and H2O2 production and accumulation will be a major part of ensuring this balance is maintained.
2.3. Hydroxyl Radicals Can Be Signals Too
Hydroxyl radicals (∙OH) are often produced in the presence of other ROS via the Fenton reaction [74] or the Haber–Weiss reaction [75]. Transition metals are therefore important for the formation of ∙OH in cells. The formation of ∙OH has been discussed by others [76,77]. ∙OH are extremely reactive and therefore not thought to be very functional as a signal. However, there are a range of papers which show that this molecule does have a role in controlling cell function [78]. ∙OH has been shown to be involved in ion movements in plant roots [79,80] and the control of kinase pathways [81]. This radical is also involved in mitochondrial oxidative stress [82] and cytoplasmic oxidative stress [83], and to participate in the modification of proteins and lipids and polysaccharides [84,85,86].
There is no doubt that ∙OH can be detected in cells [87], and their modulation has been suggested as beneficial [88,89], not just because they do damage but because they have a positive influence.
Recently a gas, molecular hydrogen (H2), has been found to be a significant influence on cell function in plants [90] and animals [91]. This is relevant here as it has been suggested that H2 is a scavenger of ∙OH [92], and thus explains its mode of action. However, this would only be significant if ∙OH did indeed have a signaling role in cells. However, this scavenging role has been disputed [93,94].
3. Signaling by RNS
As discussed above, the role of nitrogen-based signaling molecules came into focus when EDRF was discovered to be in fact NO [5]. There was flurry of activity and the enzyme responsible for the generation of NO in animals was soon found, i.e., nitric oxide synthase (NOS) [95]. It was discovered that oxygen was used in the catalytic cycle of this enzyme. Arginine acts as a substrate, but in the presence of NADPH and oxygen this is converted to a non-released intermediate, hydroxyarginine. With a further input of oxygen and NADPH the product is citrulline, and NO could be considered as a by-product. The generation of citrulline is often used as an assay for NOS activity. Therefore, oxygen is instrumental here in the generation of NO.
It was soon found that NO was involved in wide range of functions in both plants [96] and animals [97]. Recently, we saw the fortieth anniversary of NO research in plants [98]. In 1992, NO was deemed to be the molecule of the year [99].
3.1. Nitric Oxide and Working with Other Oxygen-Based Molecules
Nitric oxide appears to be a simple molecule consisting of oxygen and nitrogen. It is a radical and a gas, so seems like an unlikely biomolecule. Perhaps this explains the interest in this molecule when it came to prominence in 1987 [5]. However, even though it is often assumed that it is a radical, it can lose or gain electrons and therefore can have other chemical characteristics, a facet often overlooked when NO donor molecules are used.
NO has been found to be involved in the mediation of a wide range of biological functions, from controlled blood flow in humans [100], to controlling stomatal apertures in plants [101]. In animals, the main source is NOS. In humans, there are three isoforms of this enzyme: eNOS, iNOS, and nNOS [102]. However, the existence of such an enzyme in plants has been hotly contested and it is unlikely to exist, at least in the form that would be easily recognizable [96]. It is more likely that in plants the main source of NO is the enzyme nitrate reductase (NR) [103], although as mentioned above there are other sources of NO in biological systems.
In animals, one of the main signaling targets of NO is the enzyme soluble guanylyl cyclase (sGC) [104]. This enzyme contains a heme group which is the direct interaction with NO, and this activates the enzyme so increasing the cellular accumulation of cGMP. This molecule can then control a range of mediators including kinases and phosphodiesterases. However, the use of this pathway in plants has recently been disputed [105].
A universal mechanism of NO signaling is the modification of protein thiol groups, in what has been dubbed S-nitrosylation [106]. However, this terminology is technically incorrect, and this modification should preferably be called S-nitrosation [107]. Either way, this is the formation of the -SNO group, and like the formation of -SOH by H2O2, this formation of -SNO causes a conformational change on the protein and therefore a modulation of its activity or function. As this is a reversable reaction it is again akin to phosphorylation. However, the thiols are also able to be oxidized, as discussed, so there is likely to be competition for the thiol between the oxidation by ROS and nitrosation by NO. Furthermore, the same thiols may be under attack by H2S, in S-sulfhydration [108,109], as well as being able to be glutathionylated [110]. Which thiol modification actually results depends on the environment of the thiol and the relative concentrations of the molecule trying to attack it. As many of these reactions are reversible, the whole system is likely to be very dynamic, allowing different modifications happening with time and in different locations.
Proteins can also be nitrated on tyrosine. Therefore, NO can mediate the modification of polypeptides in more than one manner [111], and such changes are not mutually exclusive.
Last, NO can partake in some direct reactions with other important redox molecules. One of the most significant is the generation of S-nitrosoglutathione (GSNO). This not only removes glutathione from its important role as a redox mediator, especially in ROS metabolism, as discussed above [42], but it also creates a new signaling molecule. It has been suggested that GSNO is a buffer for NO, GSNO formation being reversed by S-nitrosoglutathione reductase (GSNOR) [112], but it may also be able to be moved around an organism in the vasculature [113], so allowing long-range NO signaling. NO can also react with H2S in the formation of nitrosothiol, which can act as a signal as well [114]. Furthermore, such a reaction lowers the accumulation of both NO and H2S, so may have significance to other signaling pathways.
As can be seen, NO is an immensely important signaling molecule which can alter the activity of enzymes in a variety of ways: by targeting transitions metals, such as in heme; S-nitrosation; and nitration. NO can also alter other pathways, either directly by reacting with ROS (see below), glutathione, or H2S, or by inducing antioxidant activity [115].
3.2. Peroxynitrite, as a Signal
The reaction of ROS, particularly O2∙−, and NO will yield peroxynitrite (ONOO−). This is relatively reactive compound, but it is known to partake in signaling in cells [10].
Peroxynitrite is produced during the hypersensitive response of plants, which is a result of pathogen challenge. The effects of peroxynitrite accumulation are mediated by tyrosine nitration of proteins [116]. Peroxynitrite can also react with amino acids, such as cysteine, methionine, and tryptophan [117]. As well as amino acids, RNA nitration is also possible [118]. Through nitration reactions, as well as oxidation effects, peroxynitrite can alter the phosphorylation levels in cells, by affecting both kinases and phosphatase activities [119], which would have significant consequences for signal transduction pathways. A profound effect of peroxynitrite can be seen in its control of the intrinsic apoptosis pathway, mediated by MAPK and Akt signaling [120], which would lead to cell death.
Therefore, peroxynitrite may have effects on signaling, but may be a significant mediator of NO signaling pathways, especially if ROS are accumulating spatially and temporally together with NO.
4. The Signaling of Carbon Monoxide
The last oxygen containing small signaling molecule considered here is carbon monoxide (CO) [11]. Unlike the ROS and RNS compounds, CO appears to have a more independent mode of action. Many of the effects of CO are mediated through the action of heme oxygenase [121,122]. This enzyme degrades heme to produce biliverdin, ferrous ions, and CO.
In a similar manner to ROS and RNS, CO is inherently toxic [123]. It can inhibit the activity of Complex IV of the mitochondrial ETC, for example. Even so, as it can inherently interact with metal containing proteins, it is known to modulate the activities of several enzymes, and this can lead to changes to the accumulation of ROS and NO. It can also alter cGMP levels, an instrumental intracellular signaling molecule. Furthermore, CO effects can be mediated by MAPK pathways and by changes in the activity of ion channels [124]. One of the mechanisms of action of H2 is thought to be mediated by heme oxygenase [125], which would then impinge on CO signaling.
It can be seen therefore, that CO, another oxygen-containing gaseous signal, has important effects in cells, and has even been suggested, despite its toxicity, to be a therapeutic agent [126].
5. Discussion and Conclusions
Evolution would have started in the absence of oxygen, but as the atmospheric oxygen concentration increased, this relatively reactive di-radical, and products which could be generated, had to be tolerated. Many of the compounds to which cells became exposed would have been toxic, including ROS, RNS, and H2S. Instead of simply managing the presence of these molecules, cells adapted to adopt these compounds as signal molecules, and many are now instrumental in the control of cellular activity [127]. Interestingly, they are often involved in stress responses, being produced by cells deliberately. Furthermore, such generation of these molecules is often spatial and temporally the same. This can lead to competitions and interactions between them, making the downstream effects often difficult to unravel.
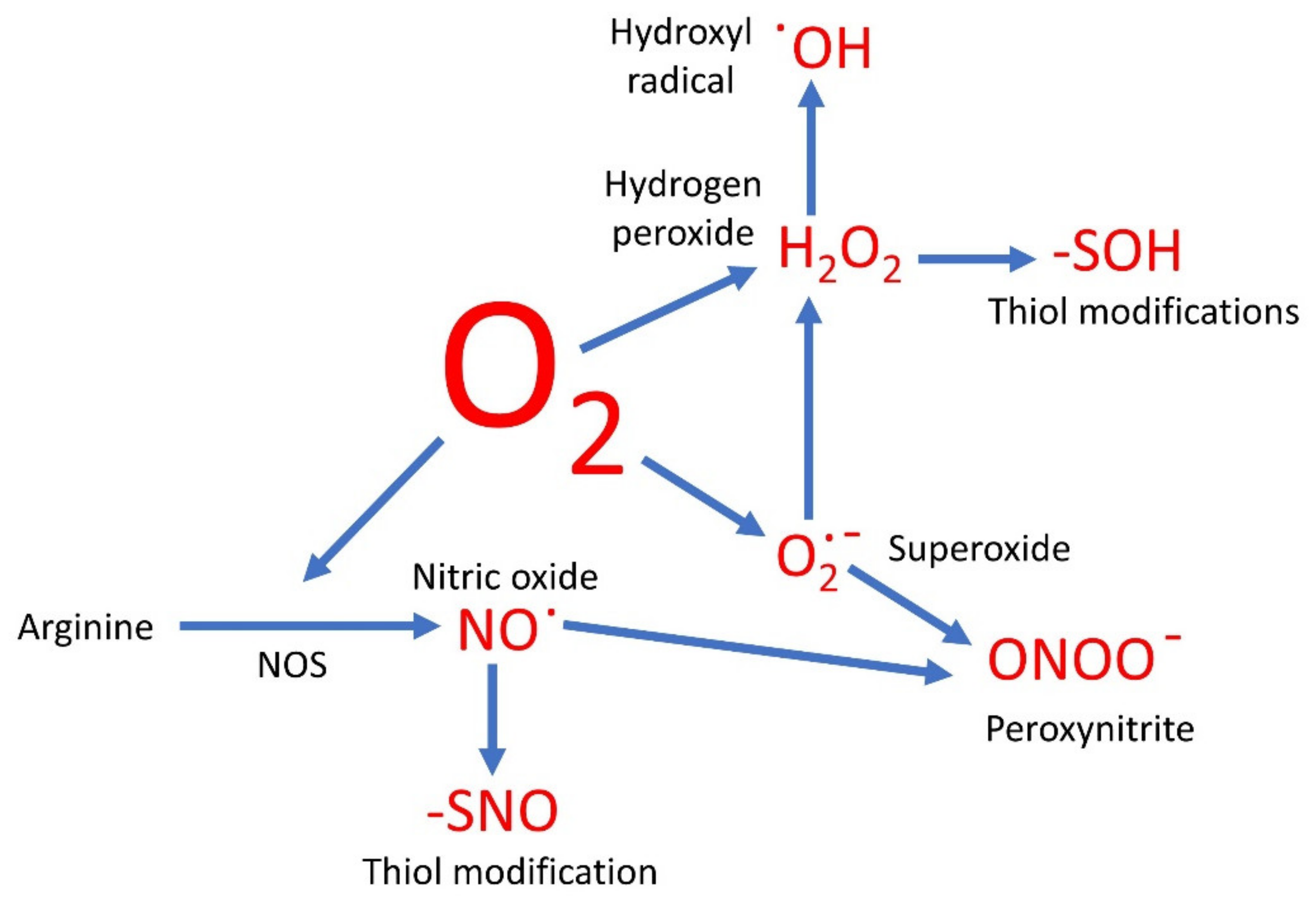
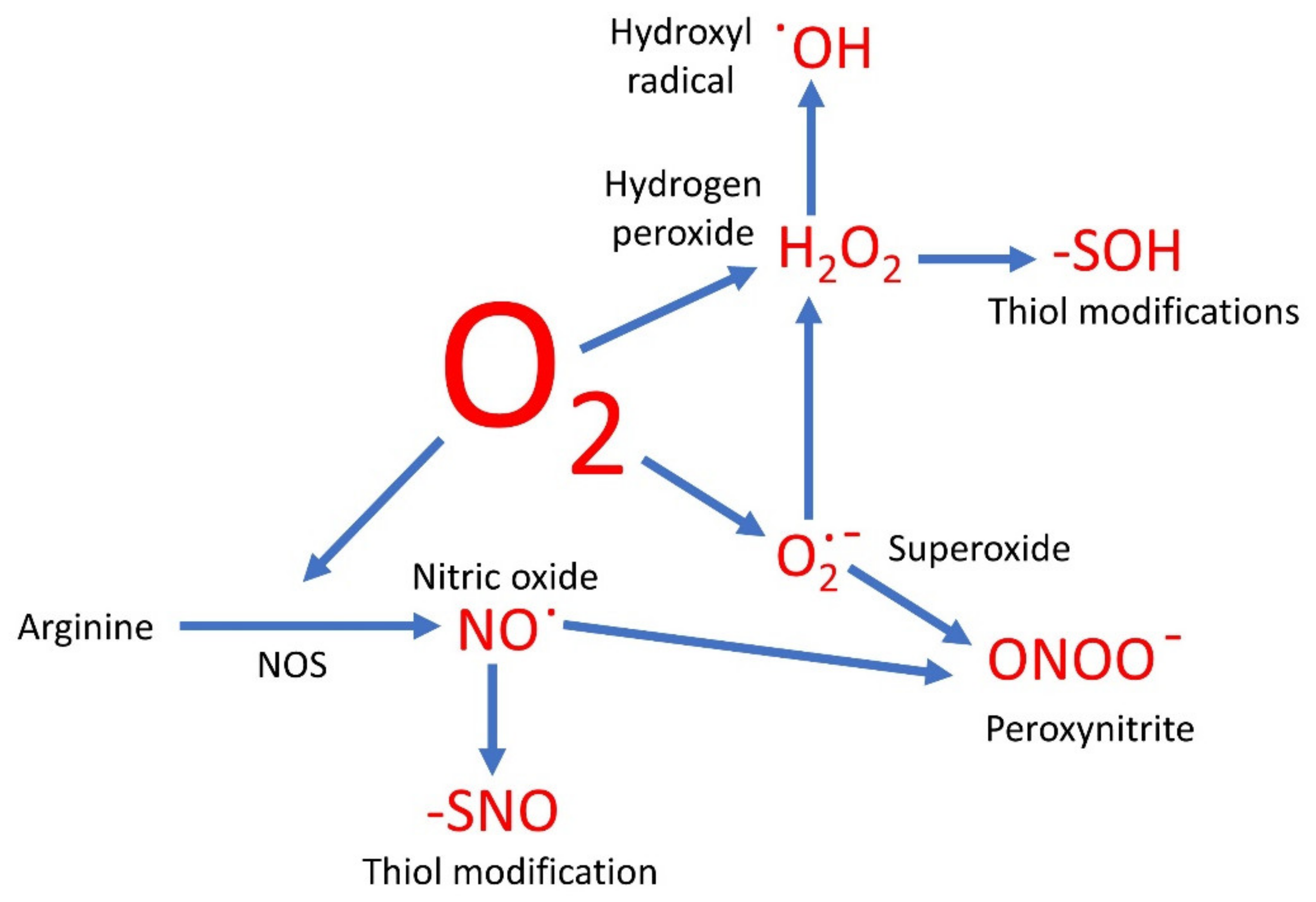
Evolution has therefore resulted in the use of a range of cell signaling molecules which are both instrumental to cellular control and contain oxygen (Figure 1). These may be reduced states of molecular oxygen, or have oxygen covalently bonded to nitrogen (NO) or carbon (CO). Each of these has potentially different roles in the cell, but they rarely work in isolation.
The accumulation of ROS and RNS has been implicated in normal and dysfunctional cellular function. ROS is seen as instrumental in hypoxia for example [39,40]. Cell growth, proliferation [128], and death [129] are also mediated by oxygen-based signaling molecules. This has implications for cancer and therapy, where many ROS- and RNS-modified proteins are being identified [130]. As well as direct control of proteins, ROS has been shown to control gene expression [131] and has been implicated in a range of diseases, including diabetes [132], neurodegenerative, and inflammatory diseases [133,134].
Overaccumulation of ROS leads to what is referred to as oxidative stress [8,9]. In this condition, the redox of the inside of the cell is pushed to an oxidized state, and this can lead to the onset of apoptosis (programmed cell death) or even necrosis [42]. Many biomolecules are damaged by ROS, including proteins, lipids, and nucleotides [135]. In a similar manner, overaccumulation of RNS can lead to nitrosative stress, with similar consequences. It is now thought that these two cellular conditions need to be considered together, in what has been referred to as nitro-oxidative stress [136].
As discussed above, there are many enzymes which can produce ROS and RNS, as well as non-enzymatic sources. It would be likely therefore, that there would be localized nitro-oxidative stress in cells, as the production of these molecules is likely to be in a diffusion gradient away from their site of generation. Such effects of signaling gradients are well known for other signals such as cAMP [137] and calcium ions (Ca2+) [138]. The cell will have numerous mechanisms to keep the levels of ROS and RNS in check. These include the presence of scavenging molecules such as glutathione. The levels and redox states of glutathione (i.e., GSH and GSSG) are used to measure the redox state of the cell, to give an estimation of oxidative stress [42]. However, there are many other small scavenging molecules, including ascorbate and α-tocopherol. Many of these are obtained by organisms in their diet. Enzymes too are present to remove harmful redox molecules. SOD will remove superoxide anions to produce H2O2. Catalase (Cat) will remove H2O2.
Some of the products of what appears to be scavenging have a useful signaling role. The production of H2O2, for example, may be important, allowing signaling that O2∙− may not be able to mediate. The reaction of glutathione with NO can lead to S-nitrosoglutathione (GSNO) [112], which may be able to move around an organism giving long-distance signaling which NO would not be able to partake in owing to its reactivity [113].
Cells, however, need to control the accumulation of these reactive molecules but still allow their concentrations to transiently rise to a level which allows them to signal to the next component in the cell’s signal transduction pathway. To do this, compartmentalization is almost certainly the key [44]. Enzymes such as SOD are compartmentalized. There are specific SODs in the mitochondria which contain manganese as their prosthetic groups, while the SOD of the cytoplasm contains copper and zinc. These will have different kinetics as well as location. Compartmentalization is known in other signaling arenas, such as those involving cAMP [137] and Ca2+ [138], and it is now becoming more recognized in signaling involving ROS, RNS, H2S, and CO [43,44,45].
Oxidative stress is on the spectrum of the redox scale, but there is now a recognition that cells can also undergo reductive stress [72,73]. This highlights how the generation and removal of reactive oxygen-containing small molecules is a balance. Like all signaling, there will be a point at which signaling is stopped, and threshold levels above which signaling proceeds. In redox, the notion of a “Goldilocks zone” has been mooted [71] and should be considered more when this type of signaling is researched and discussed.
In conclusion, there are a range of oxygen-based small, and often relatively reactive, molecules which are instrumental to signaling in cells. This applies across the kingdoms of organisms, from prokaryotes [139], through plants and animals to humans. These reactive molecules have a complex interplay which can lead to a range of responses. Metabolic enzymes, such as GAPDH [57], as well as gene expression [51] may be controlled by these molecules. The production of ROS, RSS, and RNS was not only tolerated by organisms during the early stages of evolution, but they have since been adopted as instrumental signaling components [127]. A better understanding of how the balance and compartmentalization of these molecules is achieved in cells, along with the pathologies and diseases in which they are involved, will allow such metabolism to be better controlled, with the concomitant benefits that will bring. There is still much research to carry out to measure the spatial and temporal accumulation of these molecules, and it is becoming more apparent that they should not be studied in isolation, but a holistic view of oxygen-based signaling molecules should, be taken.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The author declares no conflict of interest.
References
- Lindermayr, C. Crosstalk between reactive oxygen species and nitric oxide in plants: Key role of S-nitrosoglutathione reductase. Free Radic. Biol. Med. 2018, 122, 110–115. [Google Scholar] [CrossRef]
- Beckmann, R.; Flohé, L. The pathogenic role of superoxide radicals in inflammation: Efficacy of exogenous superoxide dismutase. Bull. Eur. Physiopathol. Respir. 1981, 17, 275–286. [Google Scholar]
- Hohn, D.C.; Lehrer, R.I. NADPH oxidase deficiency in X-linked chronic granulomatous disease. J. Clin. Investig. 1975, 55, 707–713. [Google Scholar] [CrossRef] [Green Version]
- Arnold, D.E.; Heimall, J.R. A review of Chronic Granulomatous Disease. Adv. Ther. 2017, 34, 2543–2557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, R.M.; Ferrige, A.G.; Moncada, S. Nitric oxide release accounts for the biological activity of endothelium-derived relaxing factor. Nature 1987, 327, 524–526. [Google Scholar] [CrossRef] [PubMed]
- Anson, M.L.; Mirsky, A.E. On the combination of nitric oxide with haemoglobin. J. Physiol. 1925, 60, 100–102. [Google Scholar] [CrossRef] [Green Version]
- Brandes, R.P.; Weissmann, N.; Schröder, K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014, 76, 208–226. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Hydrogen peroxide as a central redox signaling molecule in physiological oxidative stress: Oxidative eustress. Redox Biol. 2017, 11, 613–619. [Google Scholar] [CrossRef] [PubMed]
- El-Bahr, S.M. Biochemistry of free radicals and oxidative stress. Sci. Int. 2013, 1, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Speckmann, B.; Steinbrenner, H.; Grune, T.; Klotz, L.O. Peroxynitrite: From interception to signaling. Arch. Biochem. Biophys. 2016, 595, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.; Sethi, J.; Choi, A.M. Carbon monoxide-dependent signaling. Crit. Care Med. 2002, 30 Suppl. 1, S12–S17. [Google Scholar] [CrossRef] [PubMed]
- Olas, B. Hydrogen sulfide in signaling pathways. Clin. Chim. Acta. 2015, 439, 212–218. [Google Scholar] [CrossRef]
- Zeng, J.; Zhang, M.; Sun, X. Molecular hydrogen is involved in phytohormone signaling and stress responses in plants. PLoS ONE 2013, 8, e71038. [Google Scholar] [CrossRef] [PubMed]
- Hancock, J.T.; Whiteman, M. Hydrogen sulfide and cell signaling: Team player or referee? Plant Physiol. Biochem. 2014, 78, 37–42. [Google Scholar] [CrossRef]
- Corpas, F.J.; González-Gordo, S.; Cañas, A.; Palma, J.M. Nitric oxide and hydrogen sulfide in plants: Which comes first? J. Exp. Bot. 2019, 70, 4391–4404. [Google Scholar] [CrossRef]
- Monteiro, H.P.; Rodrigues, E.G.; Amorim Reis, A.K.C.; Longo, L.S., Jr.; Ogata, F.T.; Moretti, A.I.S.; da Costa, P.E.; Teodoro, A.C.S.; Toledo, M.S.; Stern, A. Nitric oxide and interactions with reactive oxygen species in the development of melanoma, breast, and colon cancer: A redox signaling perspective. Nitric Oxide 2019, 89, 1–13. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Brand, M.D. Mitochondrial generation of superoxide and hydrogen peroxide as the source of mitochondrial redox signaling. Free Radic. Biol. Med. 2016, 100, 14–31. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J.; Hampton, M.B. Reactive oxygen species and neutrophil function. Annu. Rev. Biochem. 2016, 85, 765–792. [Google Scholar] [CrossRef]
- Schröder, K. NADPH oxidases: Current aspects and tools. Redox Biol. 2020. [Google Scholar] [CrossRef]
- Qu, Y.; Yan, M.; Zhang, Q. Functional regulation of plant NADPH oxidase and its role in signaling. Plant Signal. Behav. 2017, 12, e1356970. [Google Scholar] [CrossRef] [Green Version]
- Chapman, J.M.; Muhlemann, J.K.; Gayomba, S.R.; Muday, G.K. RBOH-dependent ROS synthesis and ROS scavenging by plant specialized metabolites to modulate plant development and stress responses. Chem. Res. Toxicol. 2019, 32, 370–396. [Google Scholar] [CrossRef]
- Lambeth, J.D. Nox/Duox family of nicotinamide adenine dinucleotide (phosphate) oxidases. Curr. Opin. Hematol. 2002, 9, 11–17. [Google Scholar] [CrossRef]
- Kámán-Tóth, E.; Dankó, T.; Gullner, G.; Bozsó, Z.; Palkovics, L.; Pogány, M. Contribution of cell wall peroxidase- and NADPH oxidase-derived reactive oxygen species to Alternaria brassicicola-induced oxidative burst in Arabidopsis. Mol. Plant Pathol. 2019, 20, 485–499. [Google Scholar] [CrossRef] [Green Version]
- Colton, C.; Yao, J.; Grossman, Y.; Gilbert, D. The effect of xanthine/xanthine oxidase generated reactive oxygen species on synaptic transmission. Free Radic. Res. Commun. 1991, 14, 385–393. [Google Scholar] [CrossRef]
- Millar, T.M.; Stevens, C.R.; Benjamin, N.; Eisenthal, R.; Harrison, R.; Blake, D.R. Xanthine oxidoreductase catalyses the reduction of nitrates and nitrite to nitric oxide under hypoxic conditions. FEBS Lett. 1998, 427, 225–228. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Arasimowicz, M.; Floryszak-Wieczorek, J. Nitric oxide as a bioactive signalling molecule in plant stress responses. Plant Sci. 2007, 172, 876–887. [Google Scholar] [CrossRef]
- Liochev, S.I. Reactive oxygen species and the free radical theory of aging. Free Radic. Biol. Med. 2013, 60, 1–4. [Google Scholar] [CrossRef]
- Lapointe, J.; Hekimi, S. When a theory of aging ages badly. Cell. Mol. Life Sci. 2010, 67, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harman, D. The biologic clock: The mitochondria? J. Am. Geriatr. Soc. 1972, 20, 145–147. [Google Scholar] [CrossRef]
- Jakubczyk, K.; Dec, K.; Kałduńska, J.; Kawczuga, D.; Kochman, J.; Janda, K. Reactive oxygen species-sources, functions, oxidative damage. Pol. Merkur. Lek. Organ Pol. Tow. Lek. 2020, 48, 124–127. [Google Scholar]
- Ma, J.; Zhou, H.; Yan, S.; Song, W. Kinetics studies and mechanistic considerations on the reactions of superoxide radical ions with dissolved organic matter. Water Res. 2019, 149, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef] [PubMed]
- Wegerich, F.; Turano, P.; Allegrozzi, M.; Möhwald, H.; Lisdat, F. Cytochrome C mutants for superoxide biosensors. Anal. Chem. 2009, 81, 2976–2984. [Google Scholar] [CrossRef]
- Miller, G.; Schlauch, K.; Tam, R.; Cortes, D.; Torres, M.A.; Shulaev, V.; Dangl, J.L.; Mittler, R. The plant NADPH oxidase RBOHD mediates rapid systemic signaling in response to diverse stimuli. Sci. Signal. 2009, 2, ra45. [Google Scholar] [CrossRef] [Green Version]
- Ewald, C.Y. Redox signaling of NADPH oxidases regulates oxidative stress responses, immunity and aging. Antioxidants 2018, 7, 130. [Google Scholar] [CrossRef] [Green Version]
- Buetler, T.M.; Krauskopf, A.; Ruegg, U.T. Role of superoxide as a signaling molecule. News Physiol. Sci. 2004, 19, 120–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandel, N.S.; McClintock, D.S.; Feliciano, C.E.; Wood, T.M.; Melendez, J.A.; Rodriguez, A.M.; Schumacker, P.T. Reactive oxygen species generated at mitochondrial complex III stabilize hypoxia-inducible factor-1 alpha during hypoxia: A mechanism of O2 sensing. J. Biol. Chem. 2000, 275, 25130–25138. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Lai, U.H.; Zhu, L.; Singh, A.; Ahmed, M.; Forsyth, N.R. Reactive oxygen species formation in the brain at different oxygen levels: The role of hypoxia inducible factors. Front. Cell Dev. Biol. 2018, 6, 132. [Google Scholar] [CrossRef] [Green Version]
- Wood, P.M. The potential diagram for oxygen at pH 7. Biochem. J. 1988, 253, 287–289. [Google Scholar] [CrossRef]
- Schafer, F.Q.; Buettner, G.R. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic. Biol. Med. 2001, 30, 1191–1212. [Google Scholar] [CrossRef]
- Castro, B.; Citterico, M.; Kimura, S.; Stevens, D.M.; Wrzaczek, M.; Coaker, G. Stress-induced reactive oxygen species compartmentalization, perception and signalling. Nat. Plants 2021, 7, 1–10. [Google Scholar] [CrossRef]
- Hancock, J.T. Considerations of the importance of redox state for reactive nitrogen species action. J. Exp. Bot. 2019, 70, 4323–4331. [Google Scholar] [CrossRef] [PubMed]
- Wojtovich, A.P.; Berry, B.J.; Galkin, A. Redox signaling through compartmentalization of reactive oxygen species: Implications for health and disease. Antioxid. Redox Signal. 2019, 31, 591–593. [Google Scholar] [CrossRef] [PubMed]
- Aratani, Y. Myeloperoxidase: Its role for host defense, inflammation, and neutrophil function. Arch. Biochem. Biophys. 2018, 640, 47–52. [Google Scholar] [CrossRef]
- Smirnoff, N.; Arnaud, D. Hydrogen peroxide metabolism and functions in plants. New Phytol. 2019, 221, 1197–1214. [Google Scholar] [CrossRef] [PubMed]
- Glorieux, C.; Calderon, P.B. Catalase, a remarkable enzyme: Targeting the oldest antioxidant enzyme to find a new cancer treatment approach. Biol. Chem. 2017, 398, 1095–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhee, S.G. Overview on peroxiredoxin. Mol. Cells 2016, 39, 1. [Google Scholar]
- Brigelius-Flohé, R.; Flohé, L. Regulatory phenomena in the glutathione peroxidase superfamily. Antioxid. Redox Signal. 2020, 33, 498–516. [Google Scholar] [CrossRef]
- Marinho, H.S.; Real, C.; Cyrne, L.; Soares, H.; Antunes, F. Hydrogen peroxide sensing, signaling and regulation of transcription factors. Redox Biol. 2014, 2, 535–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winterbourn, C.C. Hydrogen peroxide reactivity and specificity in thiol-based cell signalling. Biochem. Soc. Trans. 2020, 48, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Baty, J.W.; Hampton, M.B.; Winterbourn, C.C. Proteomic detection of hydrogen peroxide-sensitive thiol proteins in Jurkat cells. Biochem. J. 2005, 389, 785–795. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, C.L.; Davies, M.J. Detection, identification, and quantification of oxidative protein modifications. J. Biol. Chem. 2019, 294, 19683–19708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmeen, A.; Andersen, J.N.; Myers, M.P.; Meng, T.C.; Hinks, J.A.; Tonks, N.K.; Barford, D. Redox regulation of protein tyrosine phosphatase 1B involves a sulphenyl-amide intermediate. Nature 2003, 423, 769–773. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Lee, S.; Park, J.B.; Lee, S.D.; Kim, J.H.; Ha, S.H.; Hasumi, K.; Endo, A.; Suh, P.G.; Ryu, S.H. Hydrogen peroxide induces association between glyceraldehyde 3-phosphate dehydrogenase and phospholipase D2 to facilitate phospholipase D2 activation in PC12 cells. J. Neurochem. 2003, 85, 1228–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildebrandt, T.; Knuesting, J.; Berndt, C.; Morgan, B.; Scheibe, R. Cytosolic thiol switches regulating basic cellular functions: GAPDH as an information hub? Biol. Chem. 2015, 396, 523–537. [Google Scholar] [CrossRef]
- Desikan, R.; A.-H.-Mackerness, S.; Hancock, J.T.; Neill, S.J. Regulation of the Arabidopsis transcriptome by oxidative stress. Plant Physiol. 2001, 127, 159–172. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.C.; Arany, P.R.; Huang, Y.Y.; Tomkinson, E.M.; Sharma, S.K.; Kharkwal, G.B.; Saleem, T.; Mooney, D.; Yull, F.E.; Blackwell, T.S.; et al. Low-level laser therapy activates NF-kB via generation of reactive oxygen species in mouse embryonic fibroblasts. PLoS ONE 2011, 6, e22453. [Google Scholar]
- Ma, Q. Role of nrf2 in oxidative stress and toxicity. Annu. Rev. Pharmacol. Toxicol. 2013, 53, 401–426. [Google Scholar] [CrossRef] [Green Version]
- Kohlgrüber, S.; Upadhye, A.; Dyballa-Rukes, N.; McNamara, C.A.; Altschmied, J. Regulation of transcription factors by reactive oxygen species and nitric oxide in vascular physiology and pathology. Antioxid. Redox Signal. 2017, 26, 679–699. [Google Scholar] [CrossRef] [PubMed]
- Burgoyne, J.R.; Oka, S.; Ale-Agha, N.; Eaton, P. Hydrogen peroxide sensing and signaling by protein kinases in the cardiovascular system. Antioxid. Redox Signal. 2013, 18, 1042–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roscoe, J.M.; Sevier, C.S. Pathways for sensing and responding to hydrogen peroxide at the endoplasmic reticulum. Cells 2020, 9, 2314. [Google Scholar] [CrossRef] [PubMed]
- Veal, E.A.; Day, A.M.; Morgan, B.A. Hydrogen peroxide sensing and signaling. Mol. Cell. 2007, 26, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Bae, S.; Choi, K.H.; An, S. Hydrogen peroxide controls Akt activity via ubiquitination/degradation pathways. Oncol. Rep. 2011, 26, 1561–1566. [Google Scholar]
- Gaschler, M.M.; Stockwell, B.R. Lipid peroxidation in cell death. Biochem. Biophys. Res. Commun. 2017, 482, 419–425. [Google Scholar] [CrossRef]
- Watt, B.E.; Proudfoot, A.T.; Vale, J.A. Hydrogen peroxide poisoning. Toxicol. Rev. 2004, 23, 51–57. [Google Scholar] [CrossRef]
- Gechev, T.S.; Hille, J. Hydrogen peroxide as a signal controlling plant programmed cell death. J. Cell Biol. 2005, 168, 17–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Zhang, Y.N.; Jia, Q.Q.; Ji, A.; Shao, S.X.; Zhang, L.; Gong, M.; Yin, Q.; Huang, X.L. MicroRNA-214 protects L6 skeletal myoblasts against hydrogen peroxide-induced apoptosis. Free Radic. Res. 2020, 54, 162–172. [Google Scholar] [CrossRef]
- Sies, H. Role of metabolic H2O2 generation: Redox signaling and oxidative stress. J. Biol. Chem. 2014, 289, 8735–8741. [Google Scholar] [CrossRef] [Green Version]
- Alleman, R.J.; Katunga, L.A.; Nelson, M.A.; Brown, D.A.; Anderson, E.J. The “Goldilocks Zone” from a redox perspective-Adaptive vs. deleterious responses to oxidative stress in striated muscle. Front. Physiol. 2014, 5, 358. [Google Scholar] [CrossRef] [Green Version]
- Manford, A.G.; Rodríguez-Pérez, F.; Shih, K.Y.; Shi, Z.; Berdan, C.A.; Choe, M.; Titov, D.V.; Nomura, D.K.; Rape, M. A cellular mechanism to detect and alleviate reductive stress. Cell 2020, 183, 46–61. [Google Scholar] [CrossRef]
- Hancock, J.T.; Veal, D. Nitric oxide, other reactive signalling compounds, redox, and reductive stress. J. Exp. Bot. 2021, 72, 819–829. [Google Scholar] [CrossRef] [PubMed]
- Fischbacher, A.; von Sonntag, C.; Schmidt, T.C. Hydroxyl radical yields in the Fenton process under various pH, ligand concentrations and hydrogen peroxide/Fe(II) ratios. Chemosphere 2017, 182, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Kehrer, J.P. The Haber-Weiss reaction and mechanisms of toxicity. Toxicology 2000, 149, 43–50. [Google Scholar] [CrossRef]
- Fong, K.L.; McCay, P.B.; Poyer, J.L. Evidence for superoxide-dependent reduction of Fe3+ and its role in enzyme-generated hydroxyl radical formation. Chem. Biol. Interact. 1976, 15, 77–89. [Google Scholar] [CrossRef]
- Halliwell, B. Superoxide-dependent formation of hydroxyl radicals in the presence of iron chelates: Is it a mechanism for hydroxyl radical production in biochemical systems? FEBS Lett. 1978, 92, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.L.; Wilkins, K.A.; Swarbreck, S.M.; Anderson, A.A.; Habib, N.; Smith, A.G.; McAinsh, M.; Davies, J.M. The hydroxyl radical in plants: From seed to seed. J. Exp. Bot. 2015, 66, 37–46. [Google Scholar] [CrossRef]
- Pottosin, I.; Zepeda-Jazo, I.; Bose, J.; Shabala, S. An anion conductance, the essential component of the hydroxyl-radical-induced ion current in plant roots. Int. J. Mol. Sci. 2018, 19, 897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demidchik, V.; Cuin, T.A.; Svistunenko, D.; Smith, S.J.; Miller, A.J.; Shabala, S.; Sokolik, A.; Yurin, V. Arabidopsis root K+-efflux conductance activated by hydroxyl radicals: Single-channel properties, genetic basis and involvement in stress-induced cell death. J. Cell Sci. 2010, 123, 1468–1479. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.J.; Lin, K.H.; Hsu, M.J.; Chou, D.S.; Hsiao, G.; Sheu, J.R. Suppression of NF-κB signaling by andrographolide with a novel mechanism in human platelets: Regulatory roles of the p38 MAPK-hydroxyl radical-ERK2 cascade. Biochem. Pharmacol. 2012, 84, 914–924. [Google Scholar] [CrossRef]
- Chen, W.; Ding, S.; Wu, J.; Shi, G.; Zhu, A. In situ detection of hydroxyl radicals in mitochondrial oxidative stress with a nanopipette electrode. Chem. Commun. 2020, 56, 13225–13228. [Google Scholar] [CrossRef]
- Sakai, T.; Imai, J.; Ito, T.; Takagaki, H.; Ui, M.; Hatta, S. The novel antioxidant TA293 reveals the role of cytoplasmic hydroxyl radicals in oxidative stress-induced senescence and inflammation. Biochem. Biophys. Res. Commun. 2017, 482, 1183–1189. [Google Scholar] [CrossRef]
- Xu, G.; Chance, M.R. Hydroxyl radical-mediated modification of proteins as probes for structural proteomics. Chem. Rev. 2007, 107, 3514–3543. [Google Scholar] [CrossRef]
- Tejero, I.; Gonzalez-Lafont, A.; Lluch, J.M.; Eriksson, L.A. Theoretical modeling of hydroxyl-radical-induced lipid peroxidation reactions. J. Phys. Chem. B 2007, 111, 5684–5693. [Google Scholar] [CrossRef]
- Gilbert, B.C.; King, D.M.; Thomas, C.B. The oxidation of some polysaccharides by the hydroxyl radical: An e.s.r. investigation. Carbohydr. Res. 1984, 125, 217–235. [Google Scholar] [CrossRef]
- Kumar, A.; Prasad, A.; Sedlářová, M.; Pospíšil, P. Data on detection of singlet oxygen, hydroxyl radical and organic radical in Arabidopsis thaliana. Data Brief. 2018, 21, 2246–2252. [Google Scholar] [CrossRef]
- Shen, B.; Jensen, R.G.; Bohnert, H.J. Mannitol protects against oxidation by hydroxyl radicals. Plant Physiol. 1997, 115, 527–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipinski, B. Hydroxyl radical and its scavengers in health and disease. Oxid. Med. Cell Longev. 2011, 2011, 809696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohta, S. Molecular hydrogen as a novel antioxidant: Overview of the advantages of hydrogen for medical applications. Methods Enzymol. 2015, 555, 289–317. [Google Scholar]
- Iida, A.; Nosaka, N.; Yumoto, T.; Knaup, E.; Naito, H.; Nishiyama, C.; Yamakawa, Y.; Tsukahara, K.; Terado, M.; Sato, K.; et al. The clinical application of hydrogen as a medical treatment. Acta Med. Okayama 2016, 70, 331–337. [Google Scholar] [PubMed]
- Ohsawa, I.; Ishikawa, M.; Takahashi, K.; Watanabe, M.; Nishimaki, K.; Yamagata, K.; Katsura, K.; Katayama, Y.; Asoh, S.; Ohta, S. Hydrogen acts as a therapeutic antioxidant by selectively reducing cytotoxic oxygen radicals. Nat. Med. 2007, 13, 688. [Google Scholar] [CrossRef]
- Penders, J.; Kissner, R.; Koppenol, W.H. ONOOH does not react with H2: Potential beneficial effects of H2 as an antioxidant by selective reaction with hydroxyl radicals and peroxynitrite. Free Radic. Biol. Med. 2014, 75, 191–194. [Google Scholar] [CrossRef]
- Hancock, J.T.; Russell, G. Downstream signalling from molecular hydrogen. Plants 2021, 10, 367. [Google Scholar] [CrossRef] [PubMed]
- Bredt, D.S.; Snyder, S.H. Isolation of nitric oxide synthetase, a calmodulin-requiring enzyme. Proc. Natl. Acad. Sci. USA 1990, 87, 682–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Astier, J.; Gross, I.; Durner, J. Nitric oxide production in plants: An update. J. Exp. Bot. 2018, 69, 3401–3411. [Google Scholar] [CrossRef] [PubMed]
- Tenopoulou, M.; Doulias, P.T. Endothelial nitric oxide synthase-derived nitric oxide in the regulation of metabolism. F1000Research 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Kolbert, Z.; Barroso, J.B.; Brouquisse, R.; Corpas, F.J.; Gupta, K.J.; Lindermayr, C.; Loake, G.J.; Palma, M.; Petřivalský, M.; Wendehenne, D.; et al. A forty year journey: The generation and roles of NO in plants. Nitric Oxide 2019, 93, 53–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koshland, D.E. The molecular of the year (Editorial). Science 1992, 258, 1861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toda, N.; Ayajiki, K.; Okamura, T. Cerebral blood flow regulation by nitric oxide: Recent advances. Pharmacol. Rev. 2009, 61, 62–97. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.R.; Yue, C.M.; Hao, F.S. Update on roles of nitric oxide in regulating stomatal closure. Plant Signal. Behav. 2019, 14, 1649569. [Google Scholar] [CrossRef] [PubMed]
- Stuehr, D.J.; Vasquez-Vivar, J. Nitric oxide synthases-from genes to function. Nitric Oxide 2017, 63, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chamizo-Ampudia, A.; Sanz-Luque, E.; Llamas, A.; Galvan, A.; Fernandez, E. Nitrate reductase regulates plant nitric oxide homeostasis. Trends Plant Sci. 2017, 22, 163–174. [Google Scholar] [CrossRef]
- Xiao, S.; Li, Q.; Hu, L.; Yu, Z.; Yang, J.; Chang, Q.; Chen, Z.; Hu, G. Soluble guanylate cyclase stimulators and activators: Where are we and where to go? Mini Rev. Med. Chem. 2019, 19, 1544–1557. [Google Scholar] [CrossRef] [PubMed]
- Astier, J.; Mounier, A.; Santolini, J.; Jeandroz, S.; Wendehenne, D. The evolution of nitric oxide signalling diverges between animal and green lineages. J. Exp. Bot. 2019, 70, 4355–4364. [Google Scholar] [CrossRef]
- Feng, J.; Chen, L.; Zuo, J. Protein S-nitrosylation in plants: Current progresses and challenges. J. Integr. Plant Biol. 2019, 61, 1206–1223. [Google Scholar] [CrossRef]
- Gupta, K.J.; Hancock, J.T.; Petrivalsky, M.; Kolbert, Z.; Lindermayr, C.; Durner, J.; Barroso, J.B.; Palma, J.M.; Brouquisse, R.; Wendehenne, D.; et al. Recommendations on terminology and experimental best practice associated with plant nitric oxide research. New Phytol. 2020, 225, 1828–1834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustafa, A.K.; Gadalla, M.M.; Sen, N.; Kim, S.; Mu, W.; Gazi, S.K.; Barrow, R.K.; Yang, G.; Wang, R.; Snyder, S.H. H2S signals through protein S-sulfhydration. Sci. Signal. 2009, 2, ra72. [Google Scholar] [CrossRef] [Green Version]
- Aroca, Á.; Serna, A.; Gotor, C.; Romero, L.C. S-sulfhydration: A cysteine posttranslational modification in plant systems. Plant Physiol. 2015, 168, 334–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Checconi, P.; Limongi, D.; Baldelli, S.; Ciriolo, M.R.; Nencioni, L.; Palamara, A.T. Role of glutathionylation in infection and inflammation. Nutrients 2019, 11, 1952. [Google Scholar] [CrossRef] [Green Version]
- Kolbert, Z.; Feigl, G.; Bordé, Á.; Molnár, Á.; Erdei, L. Protein tyrosine nitration in plants: Present knowledge, computational prediction and future perspectives. Plant Physiol. Biochem. 2017, 113, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Ventimiglia, L.; Mutus, B. The physiological implications of S-nitrosoglutathione reductase (GSNOR) activity mediating NO signalling in plant root structures. Antioxidants 2020, 9, 1206. [Google Scholar] [CrossRef]
- Rassaf, T.; Preik, M.; Kleinbongard, P.; Lauer, T.; Heiss, C.; Strauer, B.E.; Feelisch, M.; Kelm, M. Evidence for in vivo transport of bioactive nitric oxide in human plasma. J. Clin. Investig. 2002, 109, 1241–1248. [Google Scholar] [CrossRef] [PubMed]
- Whiteman, M.; Li, L.; Kostetski, I.; Chu, S.H.; Siau, J.L.; Bhatia, M.; Moore, P.K. Evidence for the formation of a novel nitrosothiol from the gaseous mediators nitric oxide and hydrogen sulphide. Biochem. Biophys. Res. Commun. 2006, 343, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Sun, L.; Liu, M.; Zhou, J. Effect of nitric oxide on reactive oxygen species and antioxidant enzymes in kiwifruit during storage. J. Sci. Food Agric. 2008, 88, 2324–2331. [Google Scholar] [CrossRef]
- Vandelle, E.; Delledonne, M. Peroxynitrite formation and function in plants. Plant Sci. 2011, 181, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, B.; Radi, R. Peroxynitrite reactivity with amino acids and proteins. Amino Acids 2003, 25, 295–311. [Google Scholar] [CrossRef]
- Staszek, P.; Gniazdowska, A. Peroxynitrite induced signaling pathways in plant response to non-proteinogenic amino acids. Planta 2020, 252, 5. [Google Scholar] [CrossRef] [PubMed]
- Klotz, L.O.; Schroeder, P.; Sies, H. Peroxynitrite signaling: Receptor tyrosine kinases and activation of stress-responsive pathways. Free Radic. Biol. Med. 2002, 33, 737–743. [Google Scholar] [CrossRef]
- Shacka, J.J.; Sahawneh, M.A.; Gonzalez, J.D.; Ye, Y.Z.; D’alessandro, T.L.; Estevez, A.G. Two distinct signaling pathways regulate peroxynitrite-induced apoptosis in PC12 cells. Cell Death Differ. 2006, 13, 1506–1514. [Google Scholar] [CrossRef] [Green Version]
- Wilks, A. Heme oxygenase: Evolution, structure, and mechanism. Antioxid. Redox Signal. 2002, 4, 603–614. [Google Scholar] [CrossRef]
- Wegiel, B.; Nemeth, Z.; Correa-Costa, M.; Bulmer, A.C.; Otterbein, L.E. Heme oxygenase-1: A metabolic nike. Antioxid. Redox Signal. 2014, 20, 1709–1722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldbaum, L.R.; Ramirez, R.G.; Absalon, K.B. What is the mechanism of carbon monoxide toxicity? Aviat. Space Environ. Med. 1975, 46, 1289–1291. [Google Scholar]
- Peers, C.; Boyle, J.P.; Scragg, J.L.; Dallas, M.L.; Al-Owais, M.M.; Hettiarachichi, N.T.; Elies, J.; Johnson, E.; Gamper, N.; Steele, D.S. Diverse mechanisms underlying the regulation of ion channels by carbon monoxide. Br. J. Pharmacol. 2015, 172, 1546–1556. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Zhang, W.; Qi, F.; Cui, W.; Xie, Y.; Shen, W. Hydrogen-rich water regulates cucumber adventitious root development in a heme oxygenase-1/carbon monoxide-dependent manner. J. Plant Physiol. 2014, 171, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kourti, M.; Jiang, W.G.; Cai, J. Aspects of carbon monoxide in form of CO-releasing molecules used in cancer treatment: More light on the way. Oxidative Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef]
- Hancock, J.T. Harnessing evolutionary toxins for signaling: Reactive oxygen species, nitric oxide and hydrogen sulfide in plant cell regulation. Front. Plant Sci. 2017, 8, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dizdaroglu, M.; Jaruga, P. Mechanisms of free radical-induced damage to DNA. Free Radic. Res. 2012, 46, 382–419. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, H. Reactive oxygen species promote vascular smooth muscle cell proliferation. Circ. Res. 2013, 113, 1040–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef] [PubMed]
- Moldogazieva, N.T.; Lutsenko, S.V.; Terentiev, A.A. Reactive oxygen and nitrogen species–induced protein modifications: Implication in carcinogenesis and anticancer therapy. Cancer Res. 2018, 78, 6040–6047. [Google Scholar] [CrossRef] [Green Version]
- Turpaev, K.T. Reactive oxygen species and regulation of gene expression. Biochemistry 2002, 67, 281–292. [Google Scholar] [CrossRef]
- Volpe, C.M.O.; Villar-Delfino, P.H.; Dos Anjos, P.M.F.; Nogueira-Machado, J.A. Cellular death, reactive oxygen species (ROS) and diabetic complications. Cell Death Dis. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Collin, F. Chemical basis of reactive oxygen species reactivity and involvement in neurodegenerative diseases. Int. J. Mol. Sci. 2019, 20, 2407. [Google Scholar] [CrossRef] [Green Version]
- Forrester, S.J.; Kikuchi, D.S.; Hernandes, M.S.; Xu, Q.; Griendling, K.K. Reactive oxygen species in metabolic and inflammatory signaling. Circ. Res. 2018, 122, 877–902. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B. Nitro-oxidative stress vs. oxidative or nitrosative stress in higher plants. New Phytol. 2013, 199, 633–635. [Google Scholar] [CrossRef] [PubMed]
- Grisan, F.; Iannucci, L.F.; Surdo, N.C.; Gerbino, A.; Zanin, S.; Di Benedetto, G.; Pozzan, T.; Lefkimmiatis, K. PKA compartmentalization links cAMP signaling and autophagy. Cell Death Differ. 2021, 1–14. [Google Scholar] [CrossRef]
- Petersen, O.H. Calcium signal compartmentalization. Biol. Res. 2002, 35, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Hillion, M.; Antelmann, H. Thiol-based redox switches in prokaryotes. Biol. Chem. 2015, 396, 415–444. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
A simplified overview of oxygen-based molecules in signaling. Oxygen and oxygen-based signaling molecules are shown in red.
Figure 1.
A simplified overview of oxygen-based molecules in signaling. Oxygen and oxygen-based signaling molecules are shown in red.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hancock, J.T. Oxygen Is Instrumental for Biological Signaling: An Overview. Oxygen 2021, 1, 3-15. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen1010002
AMA Style
Hancock JT. Oxygen Is Instrumental for Biological Signaling: An Overview. Oxygen. 2021; 1(1):3-15. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen1010002
Chicago/Turabian StyleHancock, John T. 2021. "Oxygen Is Instrumental for Biological Signaling: An Overview" Oxygen 1, no. 1: 3-15. https://0-doi-org.brum.beds.ac.uk/10.3390/oxygen1010002