Solid-Phase Synthesis and Evaluation of Glycopeptide Fragments from Rat Epididymal Cysteine-Rich Secretory Protein-1 (Crisp-1) ‡

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Solid-phase peptide synthesis
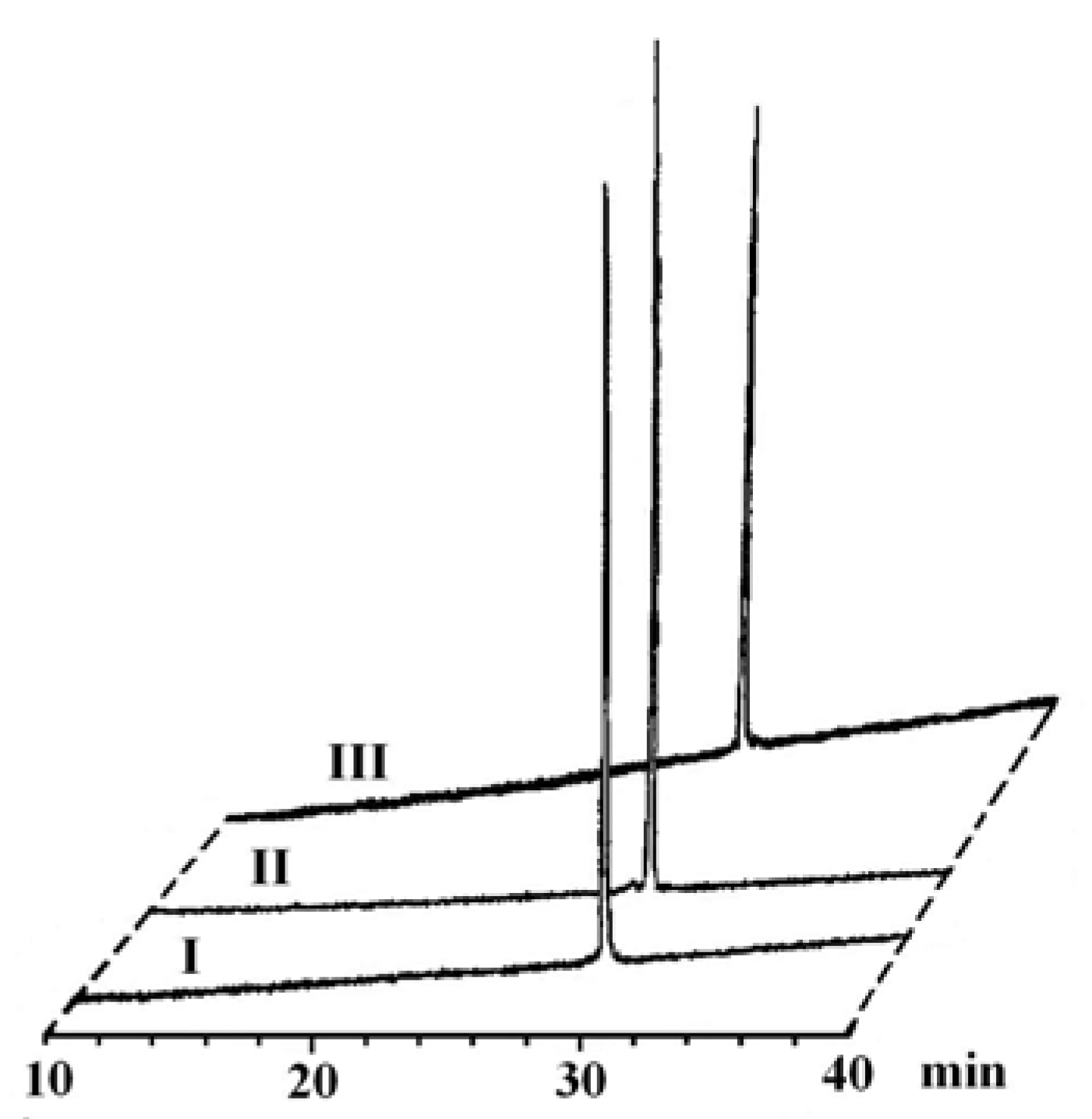
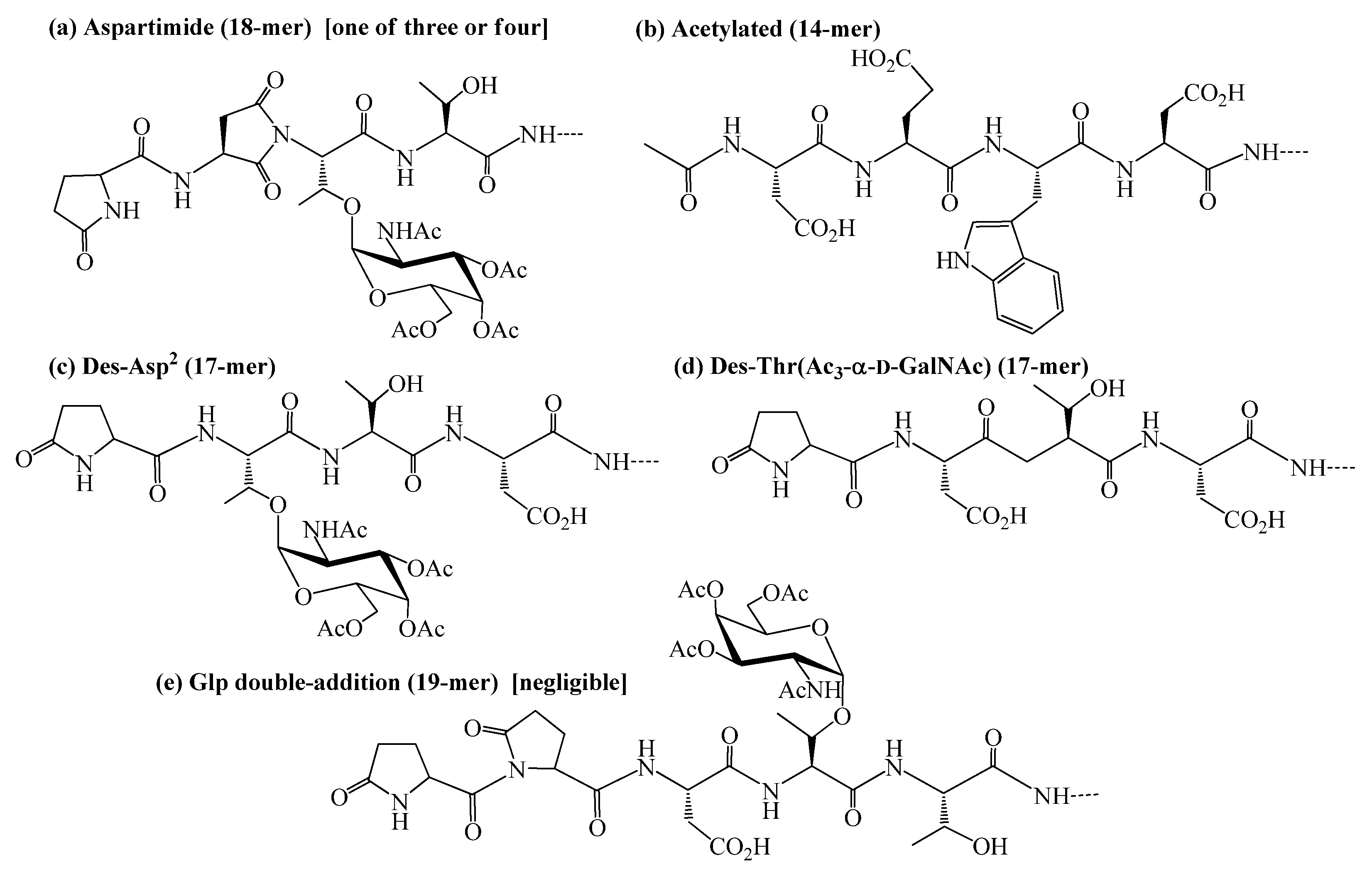
2.2. Identification and sources of glycopeptide synthesis by-products
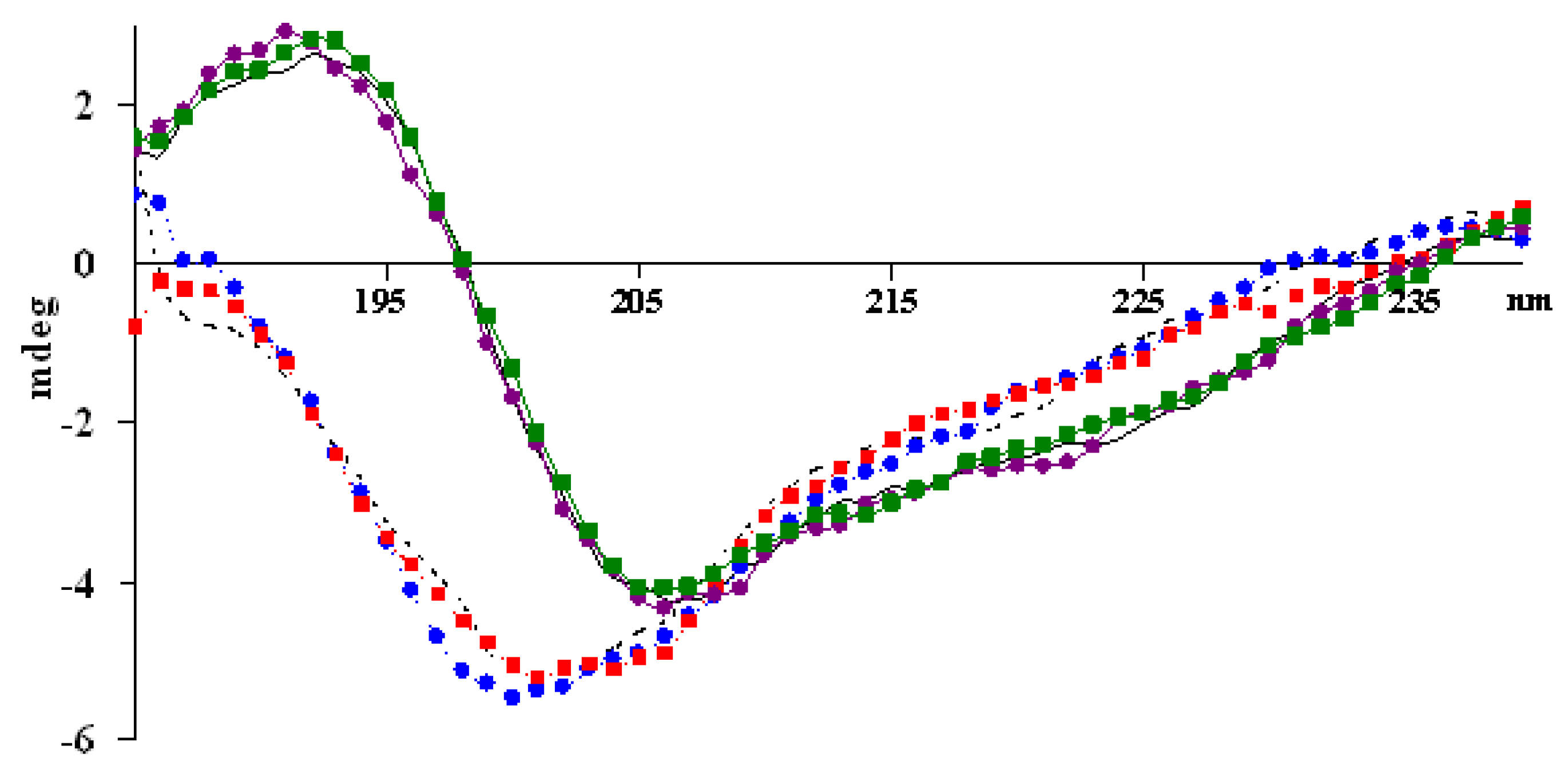
2.3. Circular dichroism spectroscopy
3. Experimental
3.1. Materials and instruments
3.2. H-Asp(OtBu)-Glu(OtBu)-Trp(Boc)-Asp(OtBu)-Arg(Pbf)-Asp(OtBu)-Leu-Glu(OtBu)-Asn(Trt)-Leu-Ser(tBu)-Thr(tBu)-Thr(tBu)-Lys(Boc)-CLEAR [‘14-mer peptide-resin’]
3.3. Glp-Asp-Thr-Thr(Ac3-α-D-GalNAc)-Asp-Glu-Trp-Asp-Arg-Asp-Leu-Glu-Asn-Leu-Ser-Thr-Thr-Lys-OH (O-acetylated I), Glp-Asp-Thr(Ac3-α-D-GalNAc)-Thr-Asp-Glu-Trp-Asp-Arg-Asp-Leu-Glu-Asn-Leu-Ser-Thr-Thr-Lys-OH (O-acetylated II), and Glp-Asp-Thr-Thr-Asp-Glu-Trp-Asp-Arg-Asp-Leu-Glu-Asn-Leu-Ser-Thr-Thr-Lys-OH (III)
3.4. Deprotection/cleavage and purification to provide glycopeptides I and II, and peptide III
3.5. Model peptide Glp-Asp-Trp-Lys
4. Conclusions
Acknowledgments
Abbreviations
References
- Yanagimachi, R. Mammalian fertilization. In The Physiology of Reproduction, 2nd ed.; Knobil, E., Neill, J.D., Eds.; Raven Press: New York, NY, USA, 1994; pp. 189–317. [Google Scholar]
- Tulsiani, D.R.; Orgebin-Crist, M.C.; Skudlarek, M.D. Role of luminal fluid glycosyltransferases and glycosidases in the modification of rat sperm plasma membrane glycoproteins during epididymal maturation. J. Reprod. Fertil. Suppl. 1998, 53, 85–97. [Google Scholar] [PubMed]
- Cooper, T.G. Interactions between epididymal secretions and spermatozoa. J. Reprod. Fertil. Suppl. 1998, 53, 119–136. [Google Scholar] [PubMed]
- Dacheux, J.L.; Gatti, J.L.; Dacheux, F. Contribution of epididymal secretory proteins for spermatozoa maturation. Microsc. Res. Tech. 2003, 61, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; James, P.S.; Howes, L.; Bruckbauer, A.; Klenerman, D. Supramolecular organization of the sperm plasma membrane during maturation and capacitation. Asian J. Androl. 2007, 9, 438–544. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Nixon, B.; Lin, M.; Koppers, A.J.; Lee, Y.H.; Baker, M.A. Proteomic changes in mammalian spermatozoa during epididymal maturation. Asian J. Androl. 2007, 9, 554–564. [Google Scholar] [CrossRef] [PubMed]
- Tanphaichitr, N.; Carmona, E.; Bou, K.M.; Xu, H.; Berger, T.; Gerton, G.L. New insights into sperm-zona pellucida interaction: Involvement of sperm lipid rafts. Front Biosci. 2007, 12, 1748–1766. [Google Scholar] [CrossRef] [PubMed]
- Van Gestel, R.A.; Brewis, I.A.; Ashton, P.R.; Brouwers, J.F.; Gadella, B.M. Multiple proteins present in purified porcine sperm apical plasma membranes interact with the zona pellucida of the oocyte. Mol. Hum. Reprod. 2007, 13, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Gadella, B.M.; Tsai, P.S.; Boerke, A.; Brewis, I.A. Sperm head membrane reorganisation during capacitation. Int. J. Dev. Biol. 2008, 52, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Hamilton, D.W. Identification of the rat epididymis-secreted 4E9 antigen as protein E. Further biochemical characterization of highly homologous epididymal secretory proteins D & E. Mol. Reprod. Dev. 1996, 43, 347–357. [Google Scholar] [PubMed]
- Xu, W.; Ensrud, K.M.; Hamilton, D.W. The 26 kD protein recognized on rat cauda epididymal sperm by monoclonal antibody 4E9 has internal peptide sequence identity with the secreted form of rat epididymal protein E. Mol. Reprod. Dev. 1997, 46, 377–382. [Google Scholar] [CrossRef]
- Roberts, K.P.; Ensrud, K.M.; Hamilton, D.W. A comparative analysis of expression and processing of the rat epididymal fluid and sperm-bound forms of proteins D and E. Biol. Reprod. 2002, 67, 525–533. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.P.; Ensrud-Bowlin, K.M.; Piehl, L.B.; Parent, K.R.; Bernhardt, M.L.; Hamilton, D.W. Association of the protein D and protein E forms of rat CRISP1 with epididymal sperm. Biol. Reprod. 2008, 79, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.P.; Wamstad, J.A.; Ensrud, K.M.; Hamilton, D.W. Inhibition of capacitation-associated tyrosine phosphorylation signaling in rat sperm by epididymal protein Crisp-1. Biol. Reprod. 2003, 69, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Brooks, D.E.; Means, A.R.; Wright, E.J.; Singh, S.P.; Tiver, K.K. Molecular cloning of the cDNA for androgen-dependent sperm-coating glycoproteins secreted by the rat epididymis. Eur. J. Biochem. 1986, 161, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.P.; Ensrud, K.M.; Wooters, J.L.; Nolan, M.A.; Johnston, D.S.; Hamilton, D.W. Epididymal secreted protein Crisp-1 and sperm function. Mol. Cell Endocrinol. 2006, 250, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Roberts, K.P.; Johnston, D.S.; Nolan, M.A.; Wooters, J.L.; Waxmonsky, N.C.; Piehl, L.B.; Ensrud-Bowlin, K.M.; Hamilton, D.W. Structure and function of epididymal protein cysteine-rich secretory protein-1. Asian J. Androl. 2007, 9, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.; Ensrud, K.M.; White, T.W.; Frethem, C.D.; Hamilton, D.W. Rat epididymis-specific sperm maturation antigens. I. Evidence that the 26 kD 4E9 antigen found on rat caudal epididymal sperm tail is derived from a protein secreted by the epididymis. Mol. Reprod. Devel. 1994, 37, 181–194. [Google Scholar] [CrossRef] [PubMed]
- Herzner, H.; Reipen, T.; Schultz, M.; Kunz, H. Synthesis of glycopeptides containing carbohydrate and peptide recognition motifs. Chem. Rev. 2000, 100, 4495–4537. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Live, D.; Barany, G. Solid-phase synthesis of mucin glycopeptides. Chimica Oggi 2004, 22, 30–34. [Google Scholar] [CrossRef]
- Buskas, T.; Ingale, S.; Boons, G. Glycopeptides as versatile tools for glycobiology. Glycobiology 2006, 16, 113R–136R. [Google Scholar] [CrossRef] [PubMed]
- Kempe, M.; Barany, G. CLEAR: A novel family of highly cross-linked polymeric supports for solid-phase peptide synthesis. J. Am. Chem. Soc. 1996, 118, 7083–7093. [Google Scholar] [CrossRef]
- Barany, G.; Merrifield, R.B. Solid-phase peptide synthesis. In The Peptides; Gross, E., Meienhofer, J., Eds.; Academic Press: New York, NY, USA, 1979; Volume 2, pp. 1–284. [Google Scholar]
- Lauer, J.L.; Fields, C.G.; Field, G.B. Sequence dependence of aspartimide formation during 9-fluorenylmethoxycarbonyl solid-phase peptide synthesis. Lett. Pept. Sci. 1995, 1, 197–205. [Google Scholar] [CrossRef]
- Martinez, J.; Bodanszky, M. Side reactions in peptide synthesis. IX. Suppression of the formation of aminosuccinyl peptides with additives. Int. J. Pept. Protein Res. 1978, 12, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Quibell, M.; Owen, D.; Packman, L.C.; Johnson, T. Suppression of piperidine-mediated side product formation for Asp(OBut)-containing peptides by the use of N-(2-hydroxy-4-methoxybenzyl)(Hmb) backbone amide protection. J. Chem. Soc. Chem. Commun. 1994, 20, 2343–2344. [Google Scholar] [CrossRef]
- Offer, J.; Quibell, M.; Johnson, T. On-resin solid-phase synthesis of asparagine N-linked glycopeptides: Use of N-(2-acetoxy-4-methoxybenzyl)(AcHmb) aspartyl amide-bond protection to prevent unwanted aspartimide formation. J. Chem. Soc. Perkin Trans. 1996, 1, 175–182. [Google Scholar] [CrossRef]
- Kelly, S.M.; Price, N.C. The use of circular dichroism in the investigation of protein structure and function. Curr. Protein Pep. Sci. 2000, 1, 349–384. [Google Scholar] [CrossRef]
- Satyanarayana, J.; Gururaja, T.L.; Narasimhamurthy, S.; Naganagowda, G.A.; Levine, M.J. Synthesis and conformational features of human salivary mucin C-terminal derived peptide epitope carrying Thomsen-Friedenreich antigen: Implications for its role in self-association. Biopolymers 2001, 58, 500–510. [Google Scholar] [CrossRef]
- Kirnarsky, L.; Prakash, O.; Vogen, S.M.; Nomoto, M.; Hollingsworth, M.A.; Sherman, S. Structural effects of O-glycosylation on a 15-residue peptide from the Mucin (MUC1) Core Protein. Biochemistry 2000, 39, 12076–12082. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.; Kahne, D. The effects of glycosylation on peptide backbone conformation. J. Am. Chem. Soc. 1993, 115, 3352–3353. [Google Scholar]
- Liang, R.; Hamilton, A.; Kahne, D. Sensitivity of glycopeptide conformation to carbohydrate chain length. J. Am. Chem. Soc. 1995, 117, 10395–10396. [Google Scholar] [CrossRef]
- Lis, H.; Sharon, N. Protein glycosylation: Structural and functional aspects. Eur. J. Biochem. 1993, 218, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Young, V.G.; Lohani, S.; Live, D.; Barany, G. Syntheses of TN building blocks Nα-(9-fluorenylmethoxycarbonyl)-O-(3,4,6-tri-O-acetyl-2-azido-2-deoxy-α-D-galactopyranosyl)-L-serine/L-threonine pentafluorophenyl esters: Comparison of protocols and elucidation of side reactions. Carbohydr. Res. 2005, 340, 1273–1285. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Barany, G.; Live, D. Parallel solid-phase synthesis of mucin-like glycopeptides. Carbohydr. Res. 2005, 340, 2111–2122. [Google Scholar] [CrossRef] [PubMed]
- Mergler, M.; Dick, F.; Sax, B.; Weiler, P.; Vorherr, T. The aspartimide problem in Fmoc-based SPPS. Part I. J. Pept. Sci. 2003, 9, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.M.; Taylor, S.D. Synthesis of disulfated peptides corresponding to the N-terminus of chemokines receptors CXCR6 (CXCR61-20) and DARC (DARC8-42) using a sulfate-protecting group strategy. J. Pept. Sci. 2010, 16, 190–199. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of glycopeptides I and II, along with peptide III, are available from Dr. Liu at his current address at The University of Georgia. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glycopeptides | Molecular Formula | Calcd. | Obsd. (m/z)a | tR (min)b | Purityc | Yieldd |
|---|---|---|---|---|---|---|
| O-Acetylated I Glycopeptide I | C103H155N25O46 C97H149N25O43 | 2478.06 2352.02 | 1240.0, 827.0 1177.0, 785.0 | 33.3 29.4 | — >98% | — 18%e |
| O-Acetylated II | C103H155N25O46 | 2478.06 | 1240.0, 827.0 | 32.9 | — | — |
| Glycopeptide II | C97H149N25O43 | 2352.02 | 1177.0, 785.0 | 29.7 | >98% | 21% |
| Peptide III | C89H136N24O38 | 2148.94 | 1075.4, 717.3 | 30.6 | >99% | 15% |
| By-products | Molecular Formula | Calcd. | Obsd. (m/z)b | tR (min)c | Amountd | |
|---|---|---|---|---|---|---|
| a | Aspartimide (18-mer) | C103H153N25O45 | 2460.05 | 1231.0, 821.4 | 31.5, 31.8, 32.0 | 8%e |
| b | Acetylated (14-mer) | C74H114N20O30 | 1762.8 | 882.4 | 30.8 | 6% |
| c | Des-Asp2 (17-mer) | C99H150N24O43 | 2363.03 | 1182.5, 788.7 | 33.3 | 3% |
| d | Des-Thr(Ac3-α-D-GalNAc) (17-mer) | C85H129N23O36 | 2047.9 | 1024.8 | 26.9 | 2% |
| e | Glp double-addition (19-mer) | C108H160N26O48 | 2589.09 | 1295.5, 864.1 | 34.3 | <1% |
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, M.; Hamilton, D.W.; Barany, G. Solid-Phase Synthesis and Evaluation of Glycopeptide Fragments from Rat Epididymal Cysteine-Rich Secretory Protein-1 (Crisp-1) ‡. Molecules 2010, 15, 6399-6410. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules15096399
Liu M, Hamilton DW, Barany G. Solid-Phase Synthesis and Evaluation of Glycopeptide Fragments from Rat Epididymal Cysteine-Rich Secretory Protein-1 (Crisp-1) ‡. Molecules. 2010; 15(9):6399-6410. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules15096399
Chicago/Turabian StyleLiu, Mian, David W. Hamilton, and George Barany. 2010. "Solid-Phase Synthesis and Evaluation of Glycopeptide Fragments from Rat Epididymal Cysteine-Rich Secretory Protein-1 (Crisp-1) ‡" Molecules 15, no. 9: 6399-6410. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules15096399