
Peruvoside, a Cardiac Glycoside, Induces Primitive Myeloid Leukemia Cell Death


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
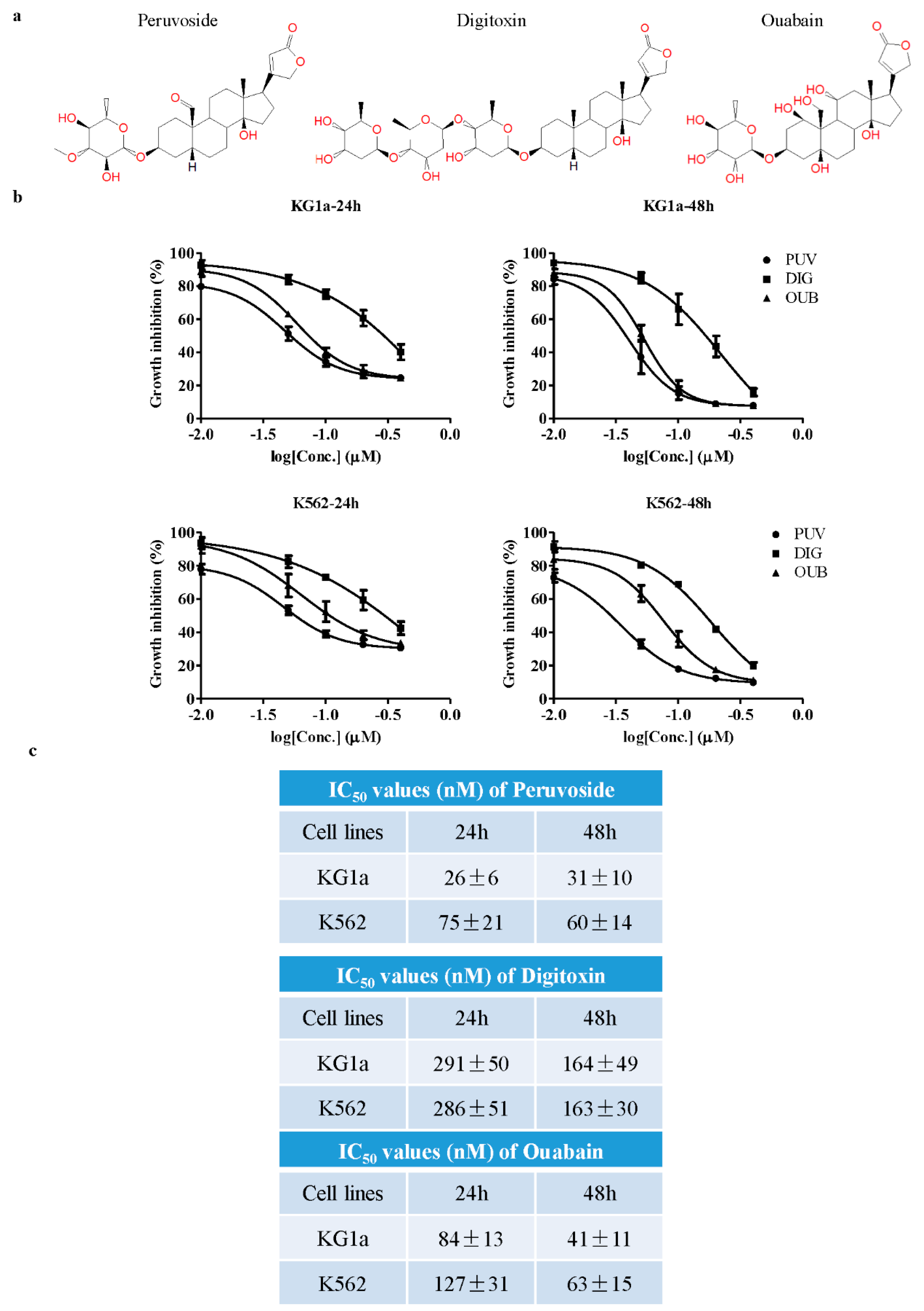
2.1. Cytotoxicity of Peruvoside on Primitive Leukemia Cells
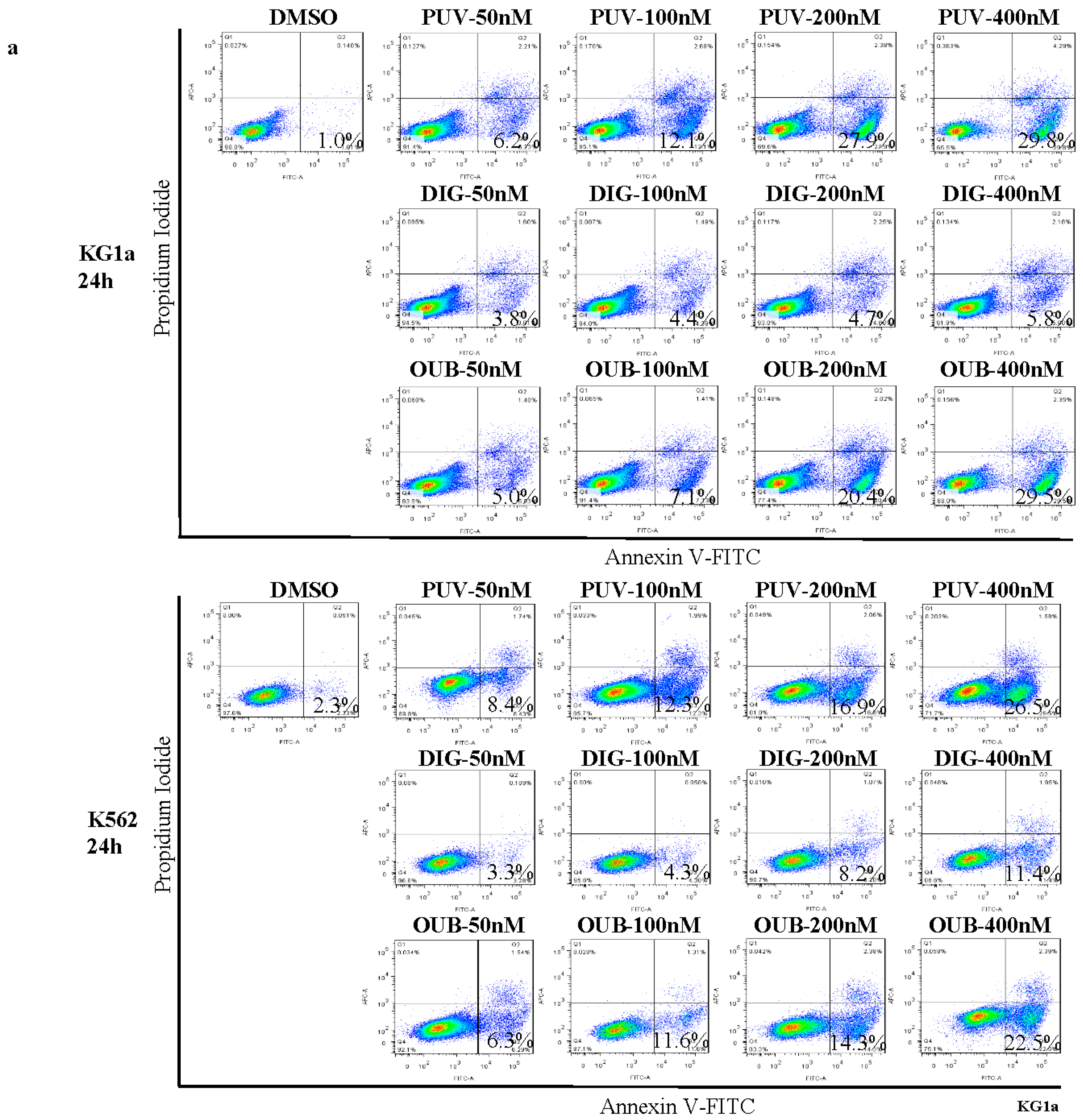
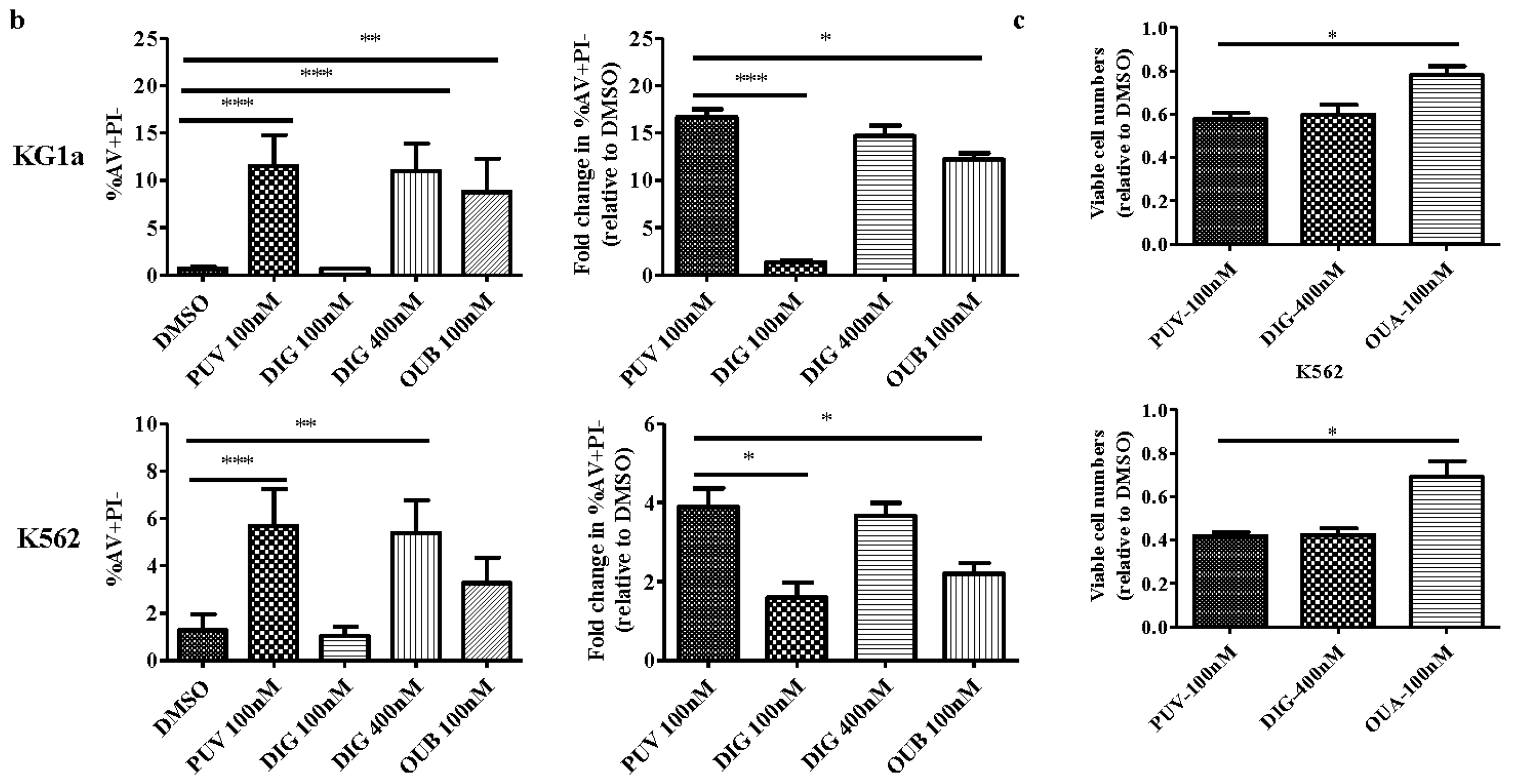
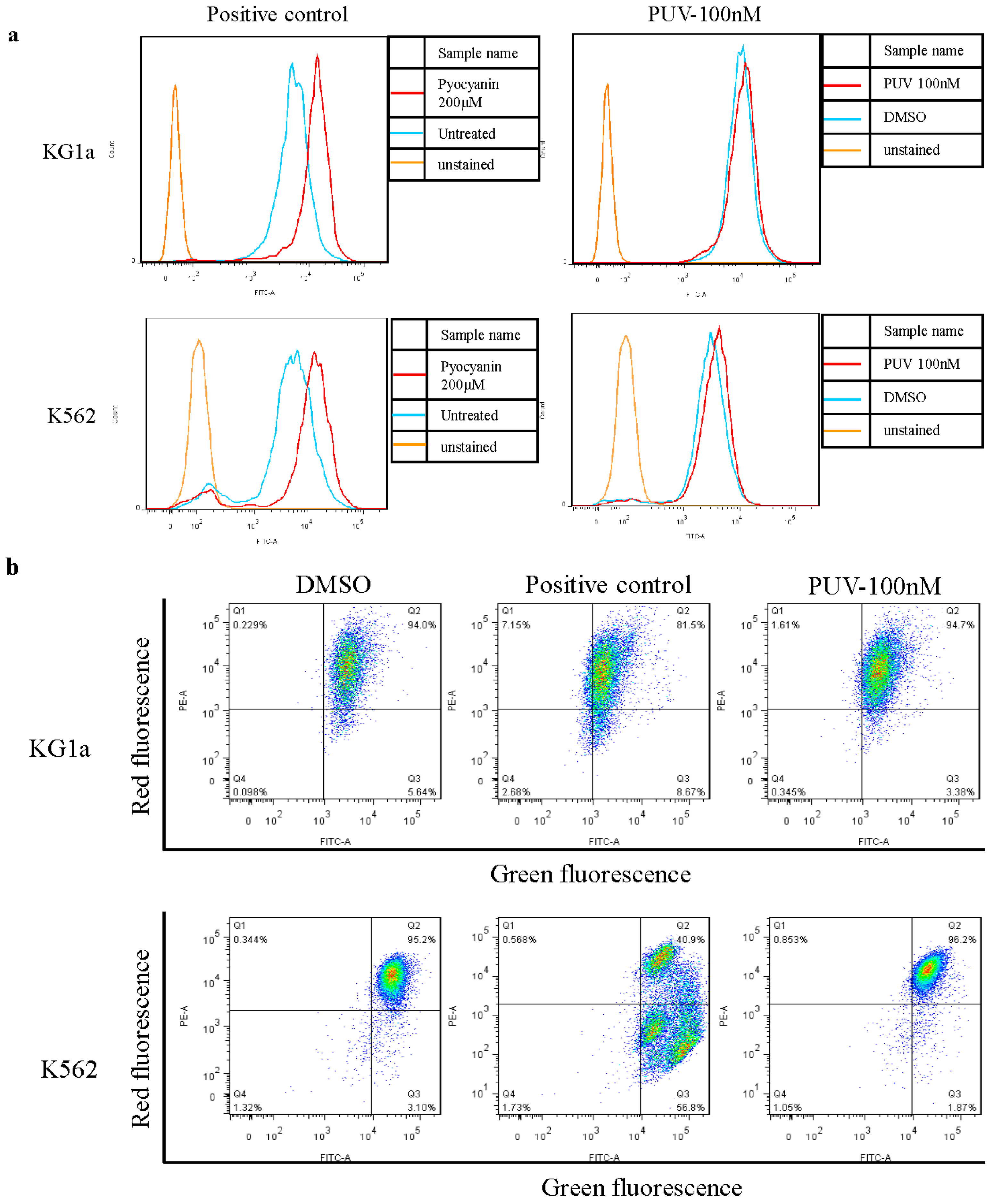
2.2. Peruvoside Induces Apoptosis in Human Leukemia Cells
2.3. Peruvoside Does Not Show Obvious Cytotoxicity on Normal Blood Cells
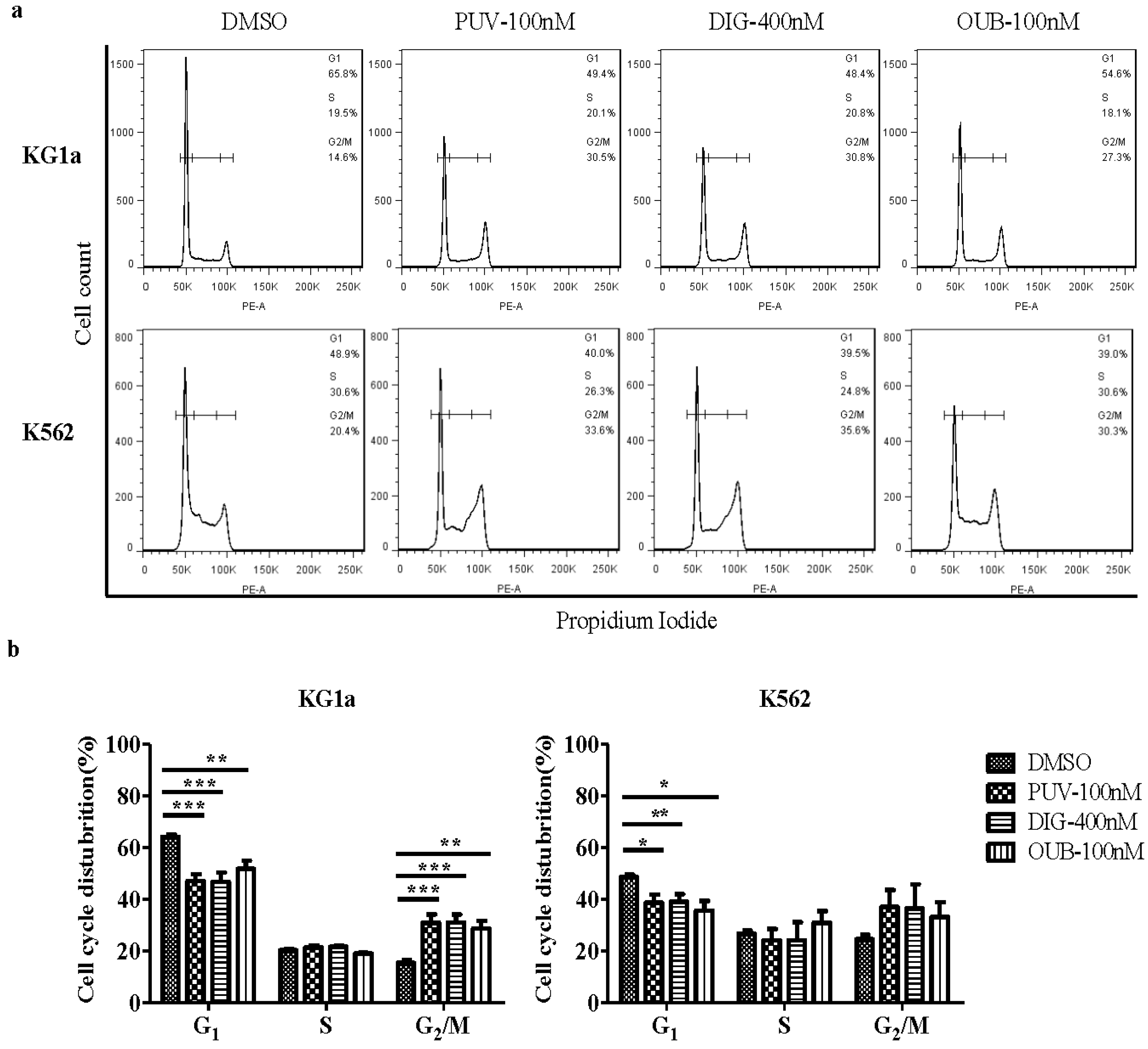
2.4. Peruvoside Induces G2/M Phase Arrest in KG1a and K562 Cells
2.5. Peruvoside Does Not Alter the Intracellular Reactive Oxygen Species (ROS) Level in Leukemia Cells
2.6. Peruvoside Does Not Affect the Mitochondrial Membrane Potential (MMP) in Leukemia Cells
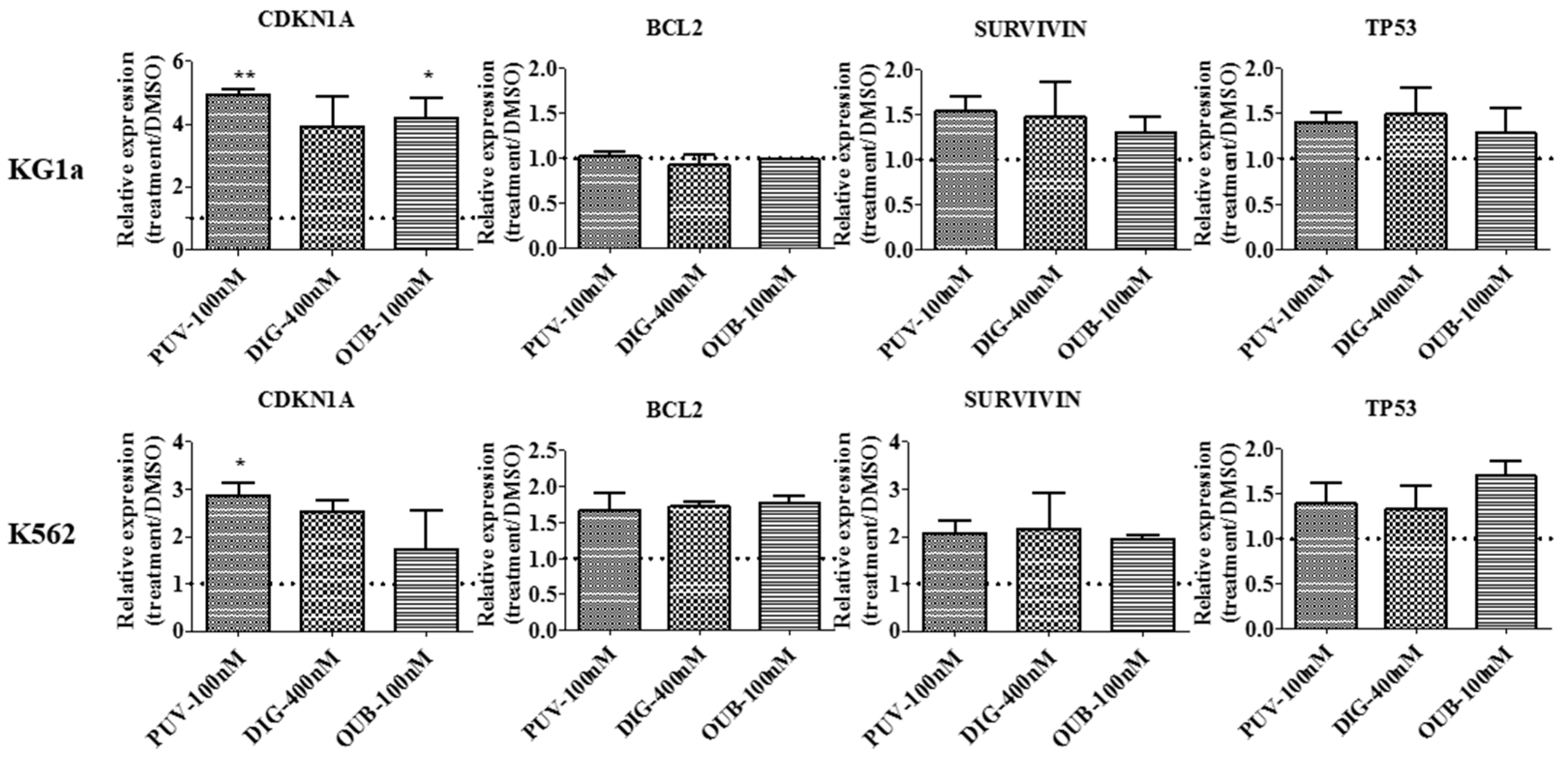
2.7. Peruvoside Induces Up-Regulation of Cyclin-Dependent Kinase Inhibitor 1A
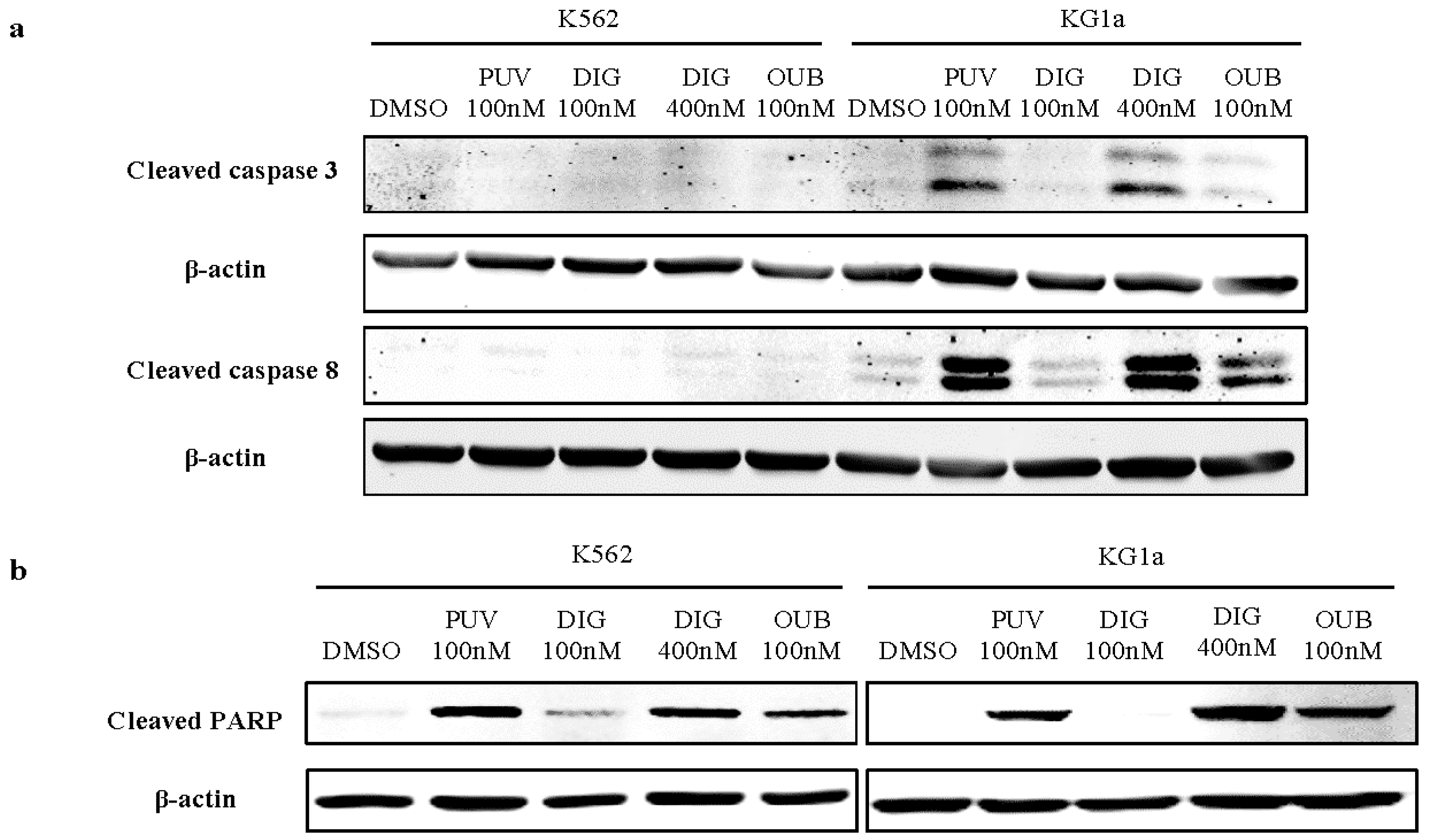
2.8. Peruvoside Activates the Cleavage of Caspase Proteins in Leukemia Cells
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. MTT Cell Viability Assay
4.4. Apoptosis and Cell Cycle Analyses
4.5. Reactive Oxygen Species Analysis
4.6. Mitochondrial Membrane Potential Analysis
4.7. Western Blot Analysis
4.8. Real-Time Polymerase Chain Reaction Analysis
4.9. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| LSC | Leukemic stem cell |
| CGs | Cardiac glycosides |
| AML | Acute myeloid leukemia |
| CML | Chronic myelogenous leukemia |
| PBMCs | Peripheral blood mononuclear cells |
| ROS | Reactive oxygen species |
| MMP | Mitochondrial membrane potential |
| PUV | Peruvoside |
| DIG | Digitoxin |
| OUB | Ouabain |
References
- Huntly, B.J.; Shigematsu, H.; Deguchi, K.; Lee, B.H.; Mizuno, S.; Duclos, N.; Rowan, R.; Amaral, S.; Curley, D.; Williams, I.R.; et al. MOZ-TIF2, but not BCR-ABL, confers properties of leukemic stem cells to committed murine hematopoietic progenitors. Cancer Cell 2004, 6, 587–596. [Google Scholar] [CrossRef] [PubMed]
- Krivtsov, A.V.; Twomey, D.; Feng, Z.; Stubbs, M.C.; Wang, Y.; Faber, J.; Levine, J.E.; Wang, J.; Hahn, W.C.; Gilliland, D.G.; et al. Transformation from committed progenitor to leukaemia stem cell initiated by MLL-AF9. Nature 2006, 442, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Renneville, A.; Roumier, C.; Biggio, V.; Nibourel, O.; Boissel, N.; Fenaux, P.; Preudhomme, C. Cooperating gene mutations in acute myeloid leukemia: A review of the literature. Leukemia 2008, 22, 915–931. [Google Scholar] [CrossRef] [PubMed]
- Guzman, M.L.; Rossi, R.M.; Neelakantan, S.; Li, X.J.; Corbett, C.A.; Hassane, D.C.; Becker, M.W.; Bennett, J.M.; Sullivan, E.; Lachowicz, J.L.; et al. An orally bioavailable parthenolide analog selectively eradicates acute myelogenous leukemia stem and progenitor cells. Blood 2007, 110, 4427–4435. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.R.; Eom, J.I.; Kim, S.J.; Jeung, H.K.; Cheong, J.W.; Kim, J.S.; Min, Y.H. Myeloperoxidase expression as a potential determinant of parthenolide-induced apoptosis in leukemia bulk and leukemia stem cells. J. Pharmacol. Exp. Ther. 2010, 335, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Rahimtoola, S.H.; Tak, T. The use of digitalis in heart failure. Curr. Probl. Cardiol. 1996, 21, 781–853. [Google Scholar] [CrossRef]
- Demers, C.; McKelvie, R.S.; Yusuf, S. The role of digitalis in the treatment of heart failure. Role Digit. Treat. Heart Fail. 1999, 10, 353–360. [Google Scholar] [CrossRef]
- Prassas, I.; Diamandis, E.P. Novel therapeutic applications of cardiac glycosides. Nat. Rev. Drug Discov. 2008, 7, 926–935. [Google Scholar] [CrossRef] [PubMed]
- Elbaz, H.A.; Stueckle, T.A.; Tse, W.; Rojanasakul, Y.; Dinu, C.Z. Digitoxin and its analogs as novel cancer therapeutics. Exp. Hematol. Oncol. 2012, 1, 4. [Google Scholar] [CrossRef] [PubMed]
- Kometiani, P.; Liu, L.; Askari, A. Digitalis-induced signaling by Na+/K+-atpase in human breast cancer cells. Mol. Pharmacol. 2005, 67, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Frese, S.; Frese-Schaper, M.; Andres, A.C.; Miescher, D.; Zumkehr, B.; Schmid1, RA. Cardiac glycosides initiate Apo2L/TRAIL-induced apoptosis in non-small cell lung cancer cells by up-regulation of death receptors 4 and 5. Cancer Res. 2006, 66, 5867–5874. [Google Scholar] [CrossRef] [PubMed]
- Felth, J.; Rickardson, L.; Rosén, J.; Wickström, M.; Fryknäs, M.; Lindskog, M.; Bohlin, L.; Gullbo, J. Cytotoxic effects of cardiac glycosides in colon cancer cells, alone and in combination with standard chemotherapeutic drugs. J. Nat. Prod. 2009, 72, 1969–1974. [Google Scholar] [CrossRef] [PubMed]
- Hallbook, H.; Felth, J.; Eriksson, A.; Fryknas, M.; Bohlin, L.; Larsson, R.; Gullbo, J. Ex vivo activity of cardiac glycosides in acute leukaemia. PLoS ONE 2011, 6, e15718. [Google Scholar] [CrossRef] [PubMed]
- Tailler, M.; Senovilla, L.; Lainey, E.; Thepot, S.; Metivier, D.; Sebert, M.; Baud, V.; Billot, K.; Fenaux, P.; Galluzzi, L.; et al. Antineoplastic activity of ouabain and pyrithione zinc in acute myeloid leukemia. Oncogene 2012, 31, 3536–3546. [Google Scholar] [CrossRef] [PubMed]
- Barry, W.H.; Hasin, Y.; Smith, T.W. Sodium pump inhibition, enhanced calcium influx via sodium-calcium exchange, and positive inotropic response in cultured heart cells. Circ. Res. 1985, 56, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Cerella, C.; Dicato, M.; Diederich, M. Assembling the puzzle of anti-cancer mechanisms triggered by cardiac glycosides. Mitochondrion 2013, 13, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Prassas, I.; Karagiannis, G.S.; Batruch, I.; Dimitromanolakis, A.; Datti, A.; Diamandis, E.P. Digitoxin-induced cytotoxicity in cancer cells is mediated through distinct kinase and interferon signaling networks. Mol. Cancer Ther. 2011, 10, 2083–2093. [Google Scholar] [CrossRef] [PubMed]
- Calderon-Montano, J.M.; Burgos-Moron, E.; Orta, M.L.; Maldonado-Navas, D.; Garcia-Dominguez, I.; Lopez-Lazaro, M. Evaluating the cancer therapeutic potential of cardiac glycosides. Biomed. Res. Int. 2014. [Google Scholar] [CrossRef] [PubMed]
- Abe, F.; Yamauchi, T.; Wan, A.S. Cardiac glycosides from the leaves of thevetia neriifolia. Phytochemistry 1992, 31, 3189–3193. [Google Scholar] [CrossRef]
- Ye, Y.X.; Yang, X.R. Inhibitory action of peruvoside and neriifolin on Na+, K(+)-ATPase. Zhongguo Yao Li Xue Bao 1990, 11, 491–494. [Google Scholar] [PubMed]
- Li, H.; Zhou, H.; Wang, D.; Qiu, J.; Zhou, Y.; Li, X.; Rosenfeld, M.G.; Ding, S.; Fu, X.D. Versatile pathway-centric approach based on high-throughput sequencing to anticancer drug discovery. Proc. Natl. Acad. Sci. USA 2012, 109, 4609–4614. [Google Scholar] [CrossRef] [PubMed]
- She, M.R.; Niu, X.Q.; Chen, X.L.; Li, J.G.; Zhou, M.H.; He, Y.J.; Le, Y.; Guo, K.Y. Resistance of leukemic stem-like cells in AML cell line KG1a to natural killer cell-mediated cytotoxicity. Cancer Lett. 2012, 318, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Cuozzo, F.; Raciti, M.; Bertelli, L.; Parente, R.; di Renzo, L. Pro-death and pro-survival properties of ouabain in U937 lymphoma derived cells. J. Exp. Clin. Cancer Res. 2012, 31, 95. [Google Scholar] [CrossRef] [PubMed]
- Kulikov, A.; Eva, A.; Kirch, U.; Boldyrev, A. Scheiner-Bobis, G. Ouabain activates signaling pathways associated with cell death in human neuroblastoma. Biochim. Biophys. Acta 2007, 1768, 1691–1702. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.T.; Chueh, S.C.; Teng, C.M.; Guh, J.-H. Investigation of ouabain-induced anticancer effect in human androgen-independent prostate cancer PC-3 cells. Biochem. Pharmacol. 2004, 67, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Dowling, R.J.O.; Goodwin, P.J.; Stambolic, V. Understanding the benefit of metformin use in cancer treatment. BMC Med. 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Ladzaro, M.; Pastor, N.; Azrak, S.S.; Ayuso, M.J.; Austin, C.A.; Cortes, F. Digitoxin inhibits the growth of cancer cell lines at concentrations commonly found in cardiac patients. J. Nat. Prod. 2005, 68, 1642–1645. [Google Scholar] [CrossRef] [PubMed]
- Xie, Z.; Cai, T. Na+-K+-atpase-mediated signal transduction: From protein interaction to cellular function. Mol. Interv. 2003, 3, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Haux, J.; Klepp, O.; Spigset, O.; Tretli, S. Digitoxin medication and cancer; case control and internal dose-response studies. BMC Cancer 2001, 1, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Lázaro, M. Digitoxin as an anticancer agent with selectivity for cancer cells: Possible mechanisms involved. Expert Opin. Ther. Targets 2007, 11, 1043–1053. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of compounds are available commercially.








© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feng, Q.; Leong, W.S.; Liu, L.; Chan, W.-I. Peruvoside, a Cardiac Glycoside, Induces Primitive Myeloid Leukemia Cell Death. Molecules 2016, 21, 534. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21040534
Feng Q, Leong WS, Liu L, Chan W-I. Peruvoside, a Cardiac Glycoside, Induces Primitive Myeloid Leukemia Cell Death. Molecules. 2016; 21(4):534. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21040534
Chicago/Turabian StyleFeng, Qian, Wa Seng Leong, Liang Liu, and Wai-In Chan. 2016. "Peruvoside, a Cardiac Glycoside, Induces Primitive Myeloid Leukemia Cell Death" Molecules 21, no. 4: 534. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules21040534