
Insights into Penicillium brasilianum Secondary Metabolism and Its Biotechnological Potential

 ,
,
Abstract
:1. Introduction
2. Penicillium brasilianum’s Environmental Sources and Isolation Methodologies
3. Secondary Metabolites Production in Penicillium brasilianum
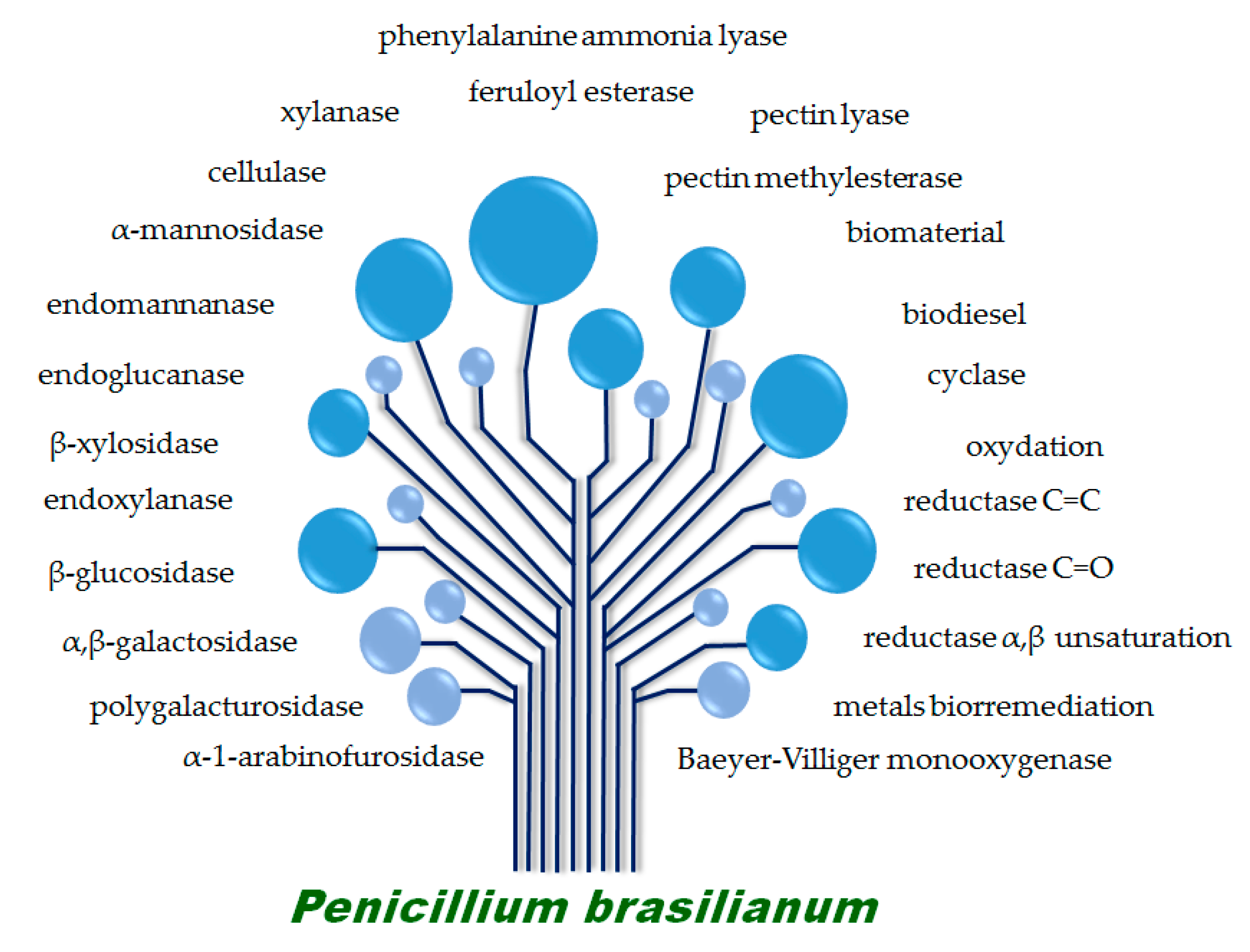
4. Biotechnological Potential of P. brasilianum
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Visagie, C.; Houbraken, J.; Frisvad, J.; Hong, S.B.; Klaassen, C.; Perrone, G.; Seifert, K.; Varga, J.; Yaguchi, T.; Samson, R. Identification and nomenclature of the genus penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.S.; Hong, S.B.; Go, S.J. First report of Penicillium brasilianum and P. daleae isolated from soil in Korea. Mycobiology 2005, 33, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Kozlovskii, A.G.; Zhelifonova, V.P.; Antipova, T.V. Fungi of the genus penicillium as producers of physiologically active compounds (review). Prikl. Biokhim. Mikrobiol. 2013, 49, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Ropars, J.; De la Vega, R.R.; Villavicencio, M.L.; Branca, A. Diversity and mechanisms of genomic adaptation in penicillium. In Aspergillus and Penicillium in the Post-Genomic Era; DeVries, R.G., Andersen, M.R., Eds.; Caister Academic Press: Lyngby, Denmark, 2016; pp. 27–42. [Google Scholar]
- Rancic, A.; Sokovic, M.; Karioti, A.; Vukojevic, J.; Skaltsa, H. Isolation and structural elucidation of two secondary metabolites from the filamentous fungus Penicillium ochrochloron with antimicrobial activity. Environ. Toxicol. Pharmacol. 2006, 22, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Lucas, E.M.F.; Castro, M.C.M.; Takahashi, J.A. Antimicrobial properties of sclerotiorin, isochromophilone vi and pencolide, metabolites from a brazilian cerrado isolate of Penicillium sclerotiorum van beyma. Braz. J. Microbiol. 2007, 38, 785–789. [Google Scholar] [CrossRef]
- Nicoletti, R.; Lopez-Gresa, M.P.; Manzo, E.; Carella, A.; Ciavatta, M.L. Production and fungitoxic activity of sch 642305, a secondary metabolite of Penicillium canescens. Mycopathologia 2007, 163, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Kwon, O.E.; Rho, M.C.; Song, H.Y.; Lee, S.W.; Chung, M.Y.; Lee, J.H.; Kim, Y.H.; Lee, H.S.; Kim, Y.K. Phenylpyropene a and b, new inhibitors of acyl-coa: Cholesterol acyltransferase produced by penicillium griseofulvum f1959. J. Antibiot. (Tokyo) 2002, 55, 1004–1008. [Google Scholar] [CrossRef] [PubMed]
- Geris dos Santos, R.M.; Rodrigues-Fo, E. Meroterpenes from Penicillium sp. found in association with melia azedarach. Phytochemistry 2002, 61, 907–912. [Google Scholar] [CrossRef]
- Petrini, O.; Sieber, T.N.; Toti, L.; Viret, O. Ecology, metabolite production, and substrate utilization in endophytic fungi. Nat. Toxins 1992, 1, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Fill, T.P.; Geris dos Santos, R.M.; Barisson, A.; Rodrigues-Fo, E.; Souza, A.Q. Co-production of bisphenylpropanoid amides and meroterpenes by an endophytic Penicillium brasilianum found in the root bark of melia azedarach. Z. Naturforsch. C 2009, 64, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Makishima, D.; Akiyama, K.; Hayashi, H. New convulsive compounds, brasiliamides a and b, from Penicillium brasilianum batista jv-379. Biosci. Biotechnol. Biochem. 2002, 66, 1697–1705. [Google Scholar] [CrossRef] [PubMed]
- Schürmann, B.T.M.; Sallum, W.S.T.; Takahashi, J.A. Austin, dehydroaustin and other metabolites from Penicillium brasilianum. Quím. Nova 2010, 33, 1044–1046. [Google Scholar] [CrossRef]
- Bander, K.I.; Al-Sanafi, A.E.; Abbas, A.R.K.H. Isolation and identification of antibiotics produced by Penicillium brasilianum batista isolated from salahaddin province soils. Thi-Qar Med. J. 2009, 3, 71–87. [Google Scholar]
- Thygesen, A.; Thomsena, A.B.; Schmidta, A.S.; Jørgensen, H.; Ahringc, B.K.; Olsson, L. Production of cellulose and hemicellulose-degrading enzymes by filamentous fungi cultivated on wet-oxidised wheat straw. Enzym. Microb. Technol. 2003, 32, 606–615. [Google Scholar] [CrossRef]
- Zeni, J.; Cence, K.; Grando, C.E.; Valduga, E. Screening of pectinase-producing microorganisms with polygalacturonase activity. Appl. Biochem. Biotechnol. 2011, 163, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Zeni, J.; Gomes, J.; Ambroszini, É.; Basso, A.P.; Toniazzo, G.; Valduga, E. Experimental design applied to the optimization and partial characterization of pectin liase from a newly isolated Penicillium brasilianum. Braz. Arch. Biol. Technol. 2014, 57, 908–915. [Google Scholar] [CrossRef]
- Zhelifonova, V.P.; Antipova, T.V.; Kozlovsky, A.G. Secondary metabolites in taxonomy of the Penicillium fungi. Microbiology 2010, 79, 277–286. [Google Scholar] [CrossRef]
- Lazarus, C.M.; Williams, K.; Bailey, A.M. Reconstructing fungal natural product biosynthetic pathways. Nat. Prod. Rep. 2014, 31, 1339–1347. [Google Scholar] [CrossRef] [PubMed]
- Fujita, T.; Hayashi, H. New brasiliamide congeners, brasiliamides c, d and e, from Penicillium brasilianum batista jv-379. Biosci. Biotechnol. Biochem. 2004, 68, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Fill, T.P.; Pereira, G.K.; Santos, R.M.G.; Rodrigues-Fo, E. Four additional meroterpenes produced by Penicillium sp. found in association with melia azedarach. Possible biosynthetic intermediates to austin. Z. Naturforsch. B 2007, 62, 1035–1044. [Google Scholar] [CrossRef]
- Fill, T.P.; Asenha, H.B.; Marques, A.S.; Ferreira, A.G.; Rodrigues-Fo, E. Time course production of indole alkaloids by an endophytic strain of Penicillium brasilianum cultivated in rice. Nat. Prod. Res. 2013, 27, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Inokoshi, J.; Nakamura, Y.; Hongbin, Z.; Uchida, R.; Nonaka, K.; Masuma, R.; Tomoda, H. Spirohexalines, new inhibitors of bacterial undecaprenyl pyrophosphate synthase, produced by Penicillium brasilianum fki-3368. J. Antibiot. (Tokyo) 2013, 66, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, H.; Mukaihara, M.; Murao, S.; Arai, M.; Lee, A.Y.; Clardy, J. Acetoxydehydroaustin, a new bioactive compound, and related compound neoaustin from Penicillium sp. Mg–11. Biosci. Biotechnol. Biochem. 1994, 58, 334–338. [Google Scholar] [CrossRef]
- Solomon, P.S.; Waters, O.D.; Oliver, R.P. Decoding the mannitol enigma in filamentous fungi. Trends Microbiol. 2007, 15, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, R.M.; Rodrigues-Fo, E. Further meroterpenes produced by Penicillium sp., an endophyte obtained from melia azedarach. Z. Naturforsch. C 2003, 58, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.M.G.; Rodrigues-Fo, E. Structures of meroterpenes produced by Penicillium sp., an endophytic fungus found associated with melia azedarach. J. Braz. Chem. Soc. 2003, 14, 722–727. [Google Scholar] [CrossRef]
- Geris, R.; Rodrigues-Fo, E.; Garcia da Silva, H.H.; Garcia da Silva, I. Larvicidal effects of fungal meroterpenoids in the control of Aedes aegypti L., the main vector of dengue and yellow fever. Chem. Biodivers. 2008, 5, 341–345. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, S.; Furutani, S.; Hirata, K.; Hayashi, H.; Matsuda, K. Three austin family compounds from Penicillium brasilianum exhibit selective blocking action on cockroach nicotinic acetylcholine receptors. Neurotoxicology 2011, 32, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Maganhi, S.H.; Fill, T.P.; Rodrigues-Fo, E.; Caracelli, I.; Zukerman-Schpector, J. Preaustinoid a: A meroterpene produced by Penicillium sp. Acta Crystallogr. Sect. E Struct Rep. Online 2009, 65, 221. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.W.; Kim, S.W. New antifungal activity of penicillic acid against phytophthora species. Biotechnol. Lett. 2004, 26, 695–698. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, S.Y.; Takahashi, M.; Shen, Y.; Kaihou, S.; Tomiyama, T.; Yazawa, M.; Tamai, Y.; Sin, Y.; Kazusaka, A.; Terazawa, M. A phytochemical in the edible tamogi-take mushroom (Pleurotus cornucopiae), d-mannitol, inhibits ace activity and lowers the blood pressure of spontaneously hypertensive rats. Biosci. Biotechnol. Biochem. 2005, 69, 1603–1605. [Google Scholar] [CrossRef] [PubMed]
- Okuyama, E.; Yamazaki, M.; Kobayashi, K.; Sakurai, T. Paraherquonin, a new meroterpenoid from Penicillium paraherquei. Tetrahedron Lett. 1983, 24, 3113–3114. [Google Scholar] [CrossRef]
- Stierle, D.B.; Stierle, A.A.; Patacini, B. The berkeleyacetals, three meroterpenes from a deep water acid mine waste penicillium. J. Nat. Prod. 2007, 70, 1820–1823. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.Y.; Zhang, Q.; Li, H.; Gao, J.M. Antimicrobial and allelopathic metabolites produced by Penicillium brasilianum. Nat. Prod. Res. 2015, 29, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Koyama, N.; Inokoshi, J.; Tomoda, H. Anti-infectious agents against mrsa. Molecules 2012, 18, 204–224. [Google Scholar] [CrossRef] [PubMed]
- Inokoshi, J.; Nakamura, Y.; Komada, S.; Komatsu, K.; Umeyama, H.; Tomoda, H. Inhibition of bacterial undecaprenyl pyrophosphate synthase by small fungal molecules. J. Antibiot. 2016, 69, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, Y.; Iwabuchi, T.; Fujimoto, T.; Awakawa, T.; Nakashima, Y.; Mori, T.; Zhang, H.; Hayashi, F.; Abe, I. Discovery of key dioxygenases that diverged the paraherquonin and acetoxydehydroaustin pathways in Penicillium brasilianum. J. Am. Chem. Soc. 2016, 138, 12671–12677. [Google Scholar] [CrossRef] [PubMed]
- Cueto, M.; Macmillan, J.B.; Jensen, P.R.; Fenical, W. Tropolactones A–D, four meroterpenoids from a marine-derived fungus of the genus Aspergillus. Phytochemistry 2006, 67, 1826–1831. [Google Scholar] [CrossRef] [PubMed]
- Chexal, K.K.; Springer, J.P.; Clardy, J.; Cole, R.J.; Kirksey, J.W.; Dorner, J.W.; Cutler, H.G.; Strawter, B.J. Austin, a novel polyisoprenoid mycotoxin from aspergillus ustus. J. Am. Chem. Soc. 1976, 98, 6748–6750. [Google Scholar] [CrossRef] [PubMed]
- Stierle, D.B.; Stierle, A.A.; Patacini, B.; McIntyre, K.; Girtsman, T.; Bolstad, E. Berkeleyones and related meroterpenes from a deep water acid mine waste fungus that inhibit the production of interleukin 1-β from induced inflammasomes. J. Nat. Prod. 2011, 74, 2273–2277. [Google Scholar] [CrossRef] [PubMed]
- Keromnes, J.; Thouvenot, D. Role of penicillic acid in the phytotoxicity of Penicillium cyclopium and Penicillium canescens to the germination of corn seeds. Appl. Environ. Microbiol. 1985, 49, 660–663. [Google Scholar] [PubMed]
- Aftab, A.; Monawwar Eqbal, M. Effect of penicillic acid on some biochemical changes in germinating maize seeds. Geobios (Jodhpur) 2002, 29, 161–163. [Google Scholar]
- Stierle, D.B.; Stierle, A.A.; Hobbs, J.D.; Stokken, J.; Clardy, J. Berkeleydione and berkeleytrione, new bioactive metabolites from an acid mine organism. Org. Lett. 2004, 6, 1049–1052. [Google Scholar] [CrossRef] [PubMed]
- Cole, R.J.; Cox, R.H. Handbook of Toxic Fungal Metabolites; Academic Press: London, UK, 1981. [Google Scholar]
- Knaus, H.G.; McManus, O.B.; Lee, S.H.; Schmalhofer, W.A.; Garcia-Calvo, M.; Helms, L.M.; Sanchez, M.; Giangiacomo, K.; Reuben, J.P.; Smith, A.B.; et al. Tremorgenic indole alkaloids potently inhibit smooth muscle high-conductance calcium-activated potassium channels. Biochemistry 1994, 33, 5819–5828. [Google Scholar] [CrossRef] [PubMed]
- Fill, T.P.; Pallini, H.F.; Amaral, L.S.; Silva, J.V.; Bidóia, D.L.; Peron, F.; Garcia, F.P.; Nakamura, C.V.; Rodrigues-Fo, E. Copper and manganese cations alter secondary metabolism in the fungus Penicillium brasilianum. J. Braz. Chem. Soc. 2016, 27, 1444–1451. [Google Scholar]
- Kawahara, T.; Takagi, M.; Shin-ya, K. Three new depsipeptides, jbir-113, jbir-114 and jbir-115, isolated from a marine sponge-derived Penicillium sp. Fs36. J. Antibiot. 2011, 65, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Hutchison, R.D.; Steyn, P.S.; Van Rensburg, S.J. Viridicatumtoxin, a new mycotoxin from Penicillium viridicatum westling. Toxicol. Appl. Pharmacol. 1973, 24, 507–509. [Google Scholar] [CrossRef]
- Bendele, A.M.; Carlton, W.W.; Nelson, G.E.; Peterson, R.E.; Grove, M.D. Viridicatumtoxin mycotoxicosis in mice and rats. Toxicol. Lett. 1984, 22, 287–291. [Google Scholar] [CrossRef]
- Bladt, T.; Dürr, C.; Knudsen, P.; Kildgaard, S.; Frisvad, J.; Gotfredsen, C.; Seiffert, M.; Larsen, T. Bio-activity and dereplication-based discovery of ophiobolins and other fungal secondary metabolites targeting leukemia cells. Molecules 2013, 18, 14629–14650. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.C.; Nielsen, J. Development of fungal cell factories for the production of secondary metabolites: Linking genomics and metabolism. Synth. Syst. Biotechnol. 2017, 2, 5–12. [Google Scholar] [CrossRef]
- Nielsen, J.C.; Grijseels, S.; Prigent, S.; Ji, B.; Dainat, J.; Nielsen, K.F.; Frisvad, J.C.; Workman, M.; Nielsen, J. Global analysis of biosynthetic gene clusters reveals vast potential of secondary metabolite production in Penicillium species. Nat. Microbiol. 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Wood, T.M. Properties of cellulolytic enzyme systems. Biochem. Soc. Trans. 1985, 13, 407–410. [Google Scholar] [CrossRef] [PubMed]
- Hoondal, G.S.; Tiwari, R.P.; Tewari, R.; Dahiya, N.; Beg, Q.K. Microbial alkaline pectinases and their industrial applications: A review. Appl. Microbiol. Biotechnol. 2002, 59, 409–418. [Google Scholar] [PubMed]
- Henning, J.; Torny, E.; Johan, B.; Folke, T.; Lisbeth, O. Purification and characterization of five cellulases and one xylanase from Penicillium brasilianum ibt 20888. Enzy. Microb. Technol. 2003, 32, 851–861. [Google Scholar]
- Jorgensen, H.; Morkeberg, A.; Krogh, K.B.; Olsson, L. Growth and enzyme production by three Penicillium species on monosaccharides. J. Biotechnol 2004, 109, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, H.; Olsson, L. Production of cellulases by Penicillium brasilianum ibt 20888—Effect of substrate on hydrolytic performance. Enzym. Microb. Technol. 2006, 38, 381–390. [Google Scholar] [CrossRef]
- Panagiotou, G.; Granouillet, P.; Olsson, L. Production and partial characterization of arabinoxylan-degrading enzymes by Penicillium brasilianum under solid-state fermentation. Appl. Microbiol. Biotechnol. 2006, 72, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Dilokpimol, A.; Mäkelä, M.R.; Aguilar-Pontes, M.V.; Benoit-Gelber, I.; Hildén, K.S.; Vries, R.P. Diversity of fungal feruloyl esterases: Updated phylogenetic classification, properties, and industrial applications. Biotechnol. Biofuels 2016, 9, 231. [Google Scholar] [CrossRef] [PubMed]
- Saha, B.C. Alpha-l-arabinofuranosidases: Biochemistry, molecular biology and application in biotechnology. Biotechnol. Adv. 2000, 18, 403–423. [Google Scholar] [CrossRef]
- Panagiotou, G.; Olavarria, R.; Olsson, L. Penicillium brasilianum as an enzyme factory; the essential role of feruloyl esterases for the hydrolysis of the plant cell wall. J. Biotechnol. 2007, 130, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Zeni, J.; Pili, J.; Cence, K.; Toniazzo, G.; Treichel, H.; Valduga, E. Characterization of novel thermostable polygalacturonases from Penicillium brasilianum and aspergillus niger. Bioprocess. Biosyst. Eng. 2015, 38, 2497–2502. [Google Scholar] [CrossRef] [PubMed]
- Zeni, J.; Ambrozini, É.; Pili, J.; Cence, K.; Backes, G.T.; Valduga, E. Production and characterization of Penicillium brasilianum pectinases with regard to industrial application. Biocatal. Biotransform. 2016, 33, 270–278. [Google Scholar] [CrossRef]
- Krogh, K.B.; Harris, P.V.; Olsen, C.L.; Johansen, K.S.; Hojer-Pedersen, J.; Borjesson, J.; Olsson, L. Characterization and kinetic analysis of a thermostable gh3 beta-glucosidase from Penicillium brasilianum. Appl. Microbiol. Biotechnol. 2010, 86, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Bojarová, P.; Petrásková, L.; Ferrandi, E.E.; Monti, D.; Pelantová, H.; Kuzma, M.; Simerská, P.; Křen, V. Glycosyl azides—An alternative way to disaccharides. Adv. Synth. Catal. 2007, 349, 1514–1520. [Google Scholar] [CrossRef]
- Fill, T.P.; da Silva, B.F.; Rodrigues-Fo, E. Biosynthesis of phenylpropanoid amides by an endophytic Penicillium brasilianum found in root bark of melia azedarach. J. Microbiol. Biotechnol. 2010, 20, 622–629. [Google Scholar] [PubMed]
- MacDonald, M.J.; D’Cunha, G.B. A modern view of phenylalanine ammonia lyase. Biochem. Cell. Biol. 2007, 85, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Nabe, K.; Izuo, N.; Nakamichi, K.; Chibata, I. Production of l-phenylalanine from trans-cinnamic acid with Rhodotorula glutinis containing l-phenylalanine ammonia-lyase activity. Appl. Environ. Microbiol. 1981, 42, 773–778. [Google Scholar] [PubMed]
- Fritz, R.R.; Hodgins, D.S.; Abell, C.W. Phenylalanine ammonia-lyase. Induction and purification from yeast and clearance in mammals. J. Biol. Chem. 1976, 251, 4646–4650. [Google Scholar] [PubMed]
- Kawasaki Watanabe, S.; Hernandez-Velazco, G.; Iturbe-Chiñas, F. Phenylalanine ammonia lyase from Sporidiobolus pararoseus and Rhodosporidium toruloides: Application for phenylalanine and tyrosine deamination. World J. Microbiol. Biotechnol. 1992, 8, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Santos-Fo, F.; Fill, T.P.; Nakamura, J.; Monteiro, M.R.; Rodrigues-Fo, E. Endophytic fungi as a source of biofuel precursors. J. Microbiol. Biotechnol. 2011, 21, 728–733. [Google Scholar] [CrossRef] [PubMed]
- Helwani, Z.; Othman, M.R.; Azizb, N.; Fernandob, W.J.N.; Kimc, J. Technologies for production of biodiesel focusing on green catalytic techniques: A review. Fuel Process. Technol. 2009, 90, 1502–1514. [Google Scholar] [CrossRef]
- Marques, M.V.; Naciuk, F.F.; Mello, A.M.S.; Seibel, N.M.; Fontoura, L.A.M. Fatty ester content determination in soybean methyl biodiesel by gas chromatography using ethyl oleate as internal standard. Quím. Nova 2010, 33, 978–980. [Google Scholar] [CrossRef]
- Kubo, A.M.; Gorup, L.F.; Amaral, L.S.; Filho, E.R.; Camargo, E.R. Kinetic control of microtubule morphology obtained by assembling gold nanoparticles on living fungal biotemplates. Bioconjug. Chem. 2016, 27, 2337–2345. [Google Scholar] [CrossRef] [PubMed]
- Borges, K.B.; Borges Wde, S.; Pupo, M.T.; Bonato, P.S. Endophytic fungi as models for the stereoselective biotransformation of thioridazine. Appl. Microbiol. Biotechnol. 2007, 77, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Borges, K.B.; Borges, W.S.; Durán-Patrón, R.; Pupo, M.T.; Bonato, P.S.; Collado, I.G. Stereoselective biotransformations using fungi as biocatalysts. Tetrahedron Asymmetry 2009, 20, 385–397. [Google Scholar] [CrossRef]
- Fill, T.P.; da Silva, J.V.; de Oliveira, K.T.; da Silva, B.F.; Rodrigues-Fo, E. Oxidative potential of some endophytic fungi using 1-indanone as a substrate. J. Microbiol. Biotechnol. 2012, 22, 832–837. [Google Scholar] [CrossRef] [PubMed]
- Din, Z.U.; Fill, T.P.; Donatoni, M.C.; Dos Santos, C.A.; Brocksom, T.J.; Rodrigues-Fo, E. Microbial diversification of diels-alder cycloadducts by whole cells of Penicillium brasilianum. Mol. Divers. 2016, 20, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Horn, F.; Linde, J.; Mattern, D.J.; Walther, G.; Guthke, R.; Brakhage, A.A.; Valiante, V. Draft genome sequence of the fungus Penicillium brasilianum mg11. Genome Announc. 2015, 3, 1–2. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Compound | Chemical Structure | Molecular Formula | Bioactivity | Reference |
|---|---|---|---|---|
| Austin 1 |  | C27H32O9 | Antibacterial, Antagonists on neuronal nicotinic acetylcholine receptors | [13,21,24,25] |
| Austinol 2 |  | C25H30O7 | [24] | |
| Dehydroaustinol 3 |  | C25H28O8 | [24] | |
| Austinolide 4 |  | C22H26O9 | Antibacterial | [9,11,13,21,26,27] |
| Austinoneol 5 |  | C24H30O6 | Antibacterial | [27] |
| Dehydroaustin 6 |  | C27H30O9 | Antagonists on neuronal nicotinic acetylcholine receptors, Insecticide | [13,24,25,28] |
| Acetoxydehydroaustin 7 |  | C29H32O11 | Antibacterial, Antagonists on neuronal nicotinic acetylcholine receptors, Insecticide | [13,24,25,27,29] |
| Neoaustin 8 |  | C25H30O6 | Antibacterial | [24,27] |
| Isoaustinone 9 |  | C25H30O6 | Antibacterial | [9,21,26,27] |
| Preaustinoid A 10 |  | C26H36O6 | Antibacterial, Inhibition of Caspase-1 | [9,30] |
| Preaustinoid A1 11 |  | C26H36O7 | Inhibition of Caspase-1 | [9,13,21,26,27,30] |
| Preaustinoid A2 12 |  | C26H34O7 | [26] | |
| Preaustinoid A3 13 |  | C26H32O7 | [21] | |
| Preaustinoid B 14 |  | C26H36O6 | Antibacterial | [9] |
| Preaustinoid B1 15 |  | C26H36O6 | [26] | |
| Preaustinoid B2 16 |  | C24H34O5 | [9,11,13,21,26,27] | |
| Brasiliamide A 17 |  | C24H26N2O6 | Bacteriostatic, Convulsive activity | [11,12,20] |
| Brasiliamide B 18 |  | C24H26N2O5 | Antibacterial, Convulsive activity | [11,12,20] |
| Brasiliamide C 19 |  | C24H26N2O5 | Convulsive activity | [11,12,20] |
| Brasiliamide D 20 |  | C24H28N2O5 | Convulsive activity | [11,12,20] |
| Brasiliamide E 21 |  | C22H26N2O4 | [11,12,20] | |
| Brasiliamide F 22 |  | C22H22N2O5 | Antibacterial | [11,20] |
| Penicillic acid 23 |  | C8H10O4 | Antibacterial, herbicide, Inhibit germination of fungal spores | [13,31] |
| d-mannitol 24 |  | C6H14O6 | Antibacterial Anti-hypertensive | [13,32] |
| Verruculogen 25 |  | C27H33N3O7 | Tremorgenic, Antibacterial, Week antiparasitary | [9,23,24,33,34] |
| Verruculogen TR-2 26 |  | C22H27N3O6 | [22] | |
| Verruculogen TR-2 epimer 27 |  | C22H27N3O6 | [22] | |
| Isoroquefortine C 28 |  | C22H23N5O2 | Antifungal | [35] |
| Griseofulvin 29 |  | C17H17ClO6 | Antibacterial, Antifungal | [35] |
| Ergosterol peroxide 30 |  | C28H44O3 | [35] | |
| 3β-Hydroxy-(22E,24R)-ergosta-5,8,22-trien-7-one 31 |  | C28H43O2 | [35] | |
| Cerevisterol 32 |  | C28H46O3 | [35] | |
| (22E,24R)-6β-Methoxyergosta-7,22-diene-3β,5α-diol 33 |  | C29H48O3 | [35] | |
| JBIR 113 34 |  | C31H42N5O7 | [36] | |
| JBIR 114 35 |  | C30H39N5O7 | [36] | |
| JBIR 115 36 |  | C30H39N5O7 | [36] | |
| Spirohexaline 37 |  | C31H32O10 | Antibacterial, | [23,24] |
| Viridicatumtoxin 38 |  | C30H31NO10 | Antibacterial, Antifungal, Cytotoxic against lymphocytic leukemia | [23,24,37] |
| Paraherquonin 39 |  | C24H28O7 | - | [38] |
| Berkeleydione 40 |  | C26H32O7 | Inhibition of Metalloproteinase- 3 and Caspase-1 | [38] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazioli, J.M.; Amaral, L.D.S.; Fill, T.P.; Rodrigues-Filho, E. Insights into Penicillium brasilianum Secondary Metabolism and Its Biotechnological Potential. Molecules 2017, 22, 858. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22060858
Bazioli JM, Amaral LDS, Fill TP, Rodrigues-Filho E. Insights into Penicillium brasilianum Secondary Metabolism and Its Biotechnological Potential. Molecules. 2017; 22(6):858. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22060858
Chicago/Turabian StyleBazioli, Jaqueline Moraes, Luciana Da Silva Amaral, Taícia Pacheco Fill, and Edson Rodrigues-Filho. 2017. "Insights into Penicillium brasilianum Secondary Metabolism and Its Biotechnological Potential" Molecules 22, no. 6: 858. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules22060858