Demethylbelamcandaquinone B (Dmcq B) Is the Active Compound of Marantodes pumilum var. alata (Blume) Kuntze with Osteoanabolic Activities

 ,
,
Abstract
:1. Introduction
2. Results
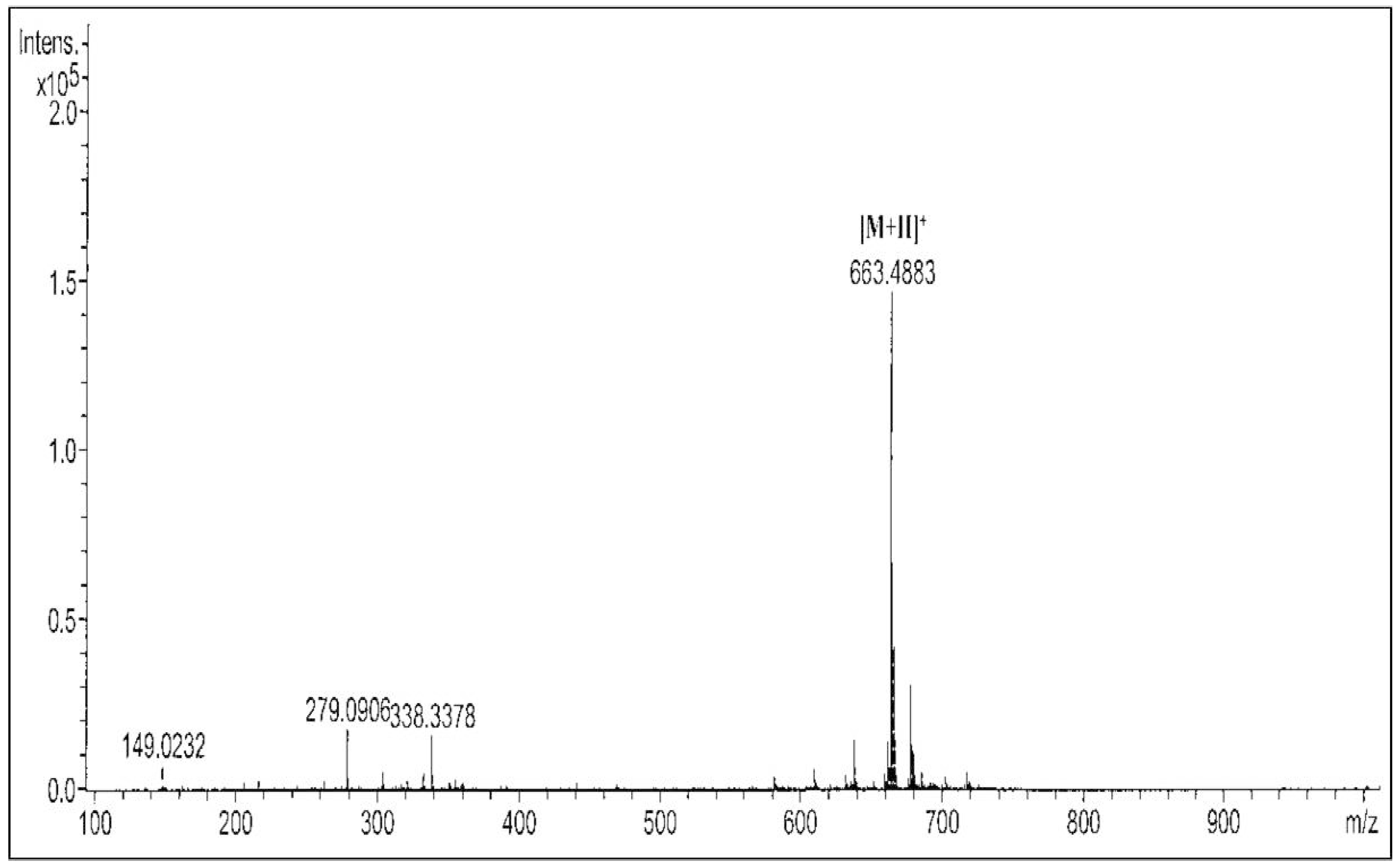
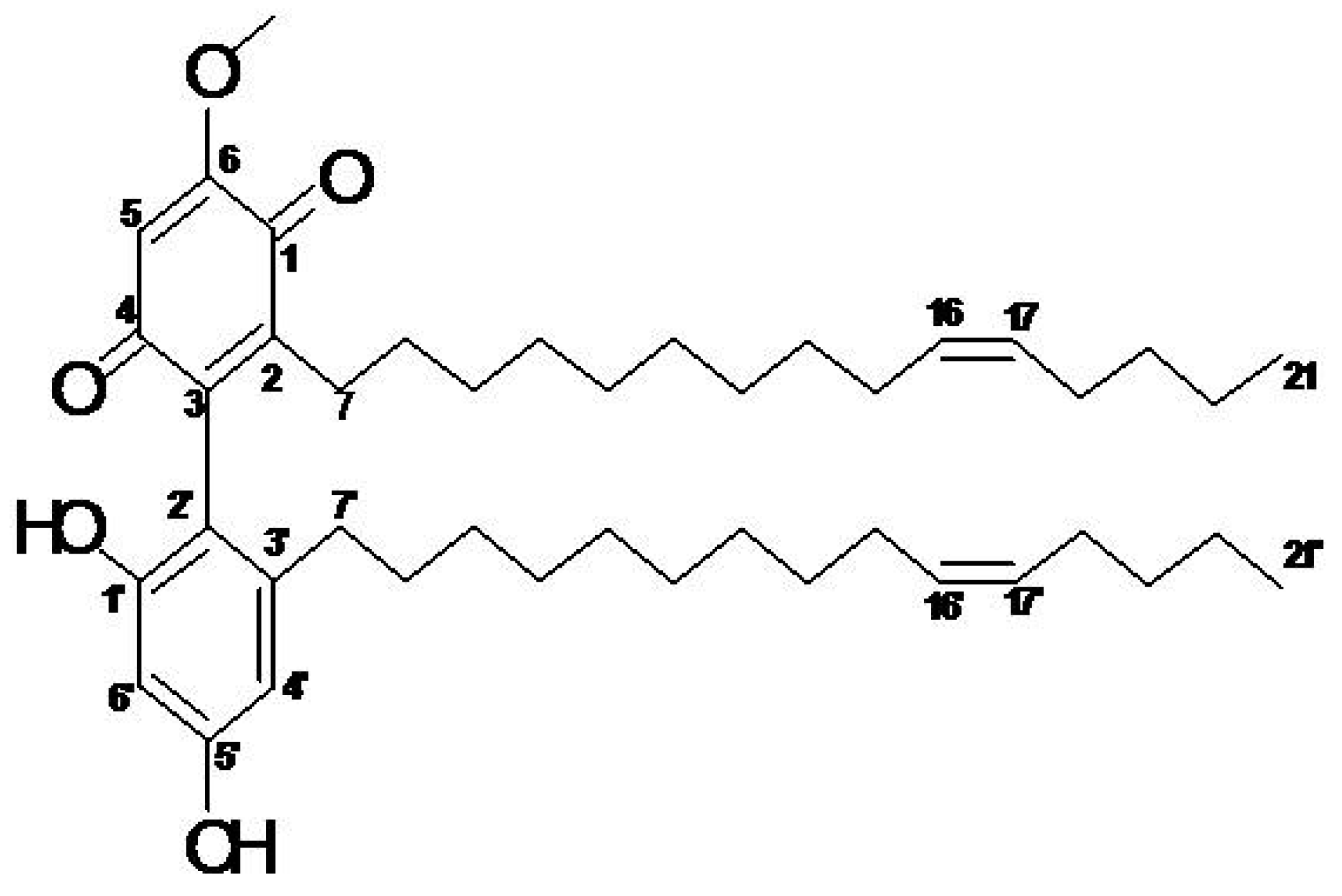
2.1. Active Compound of M. pumilum var. alata
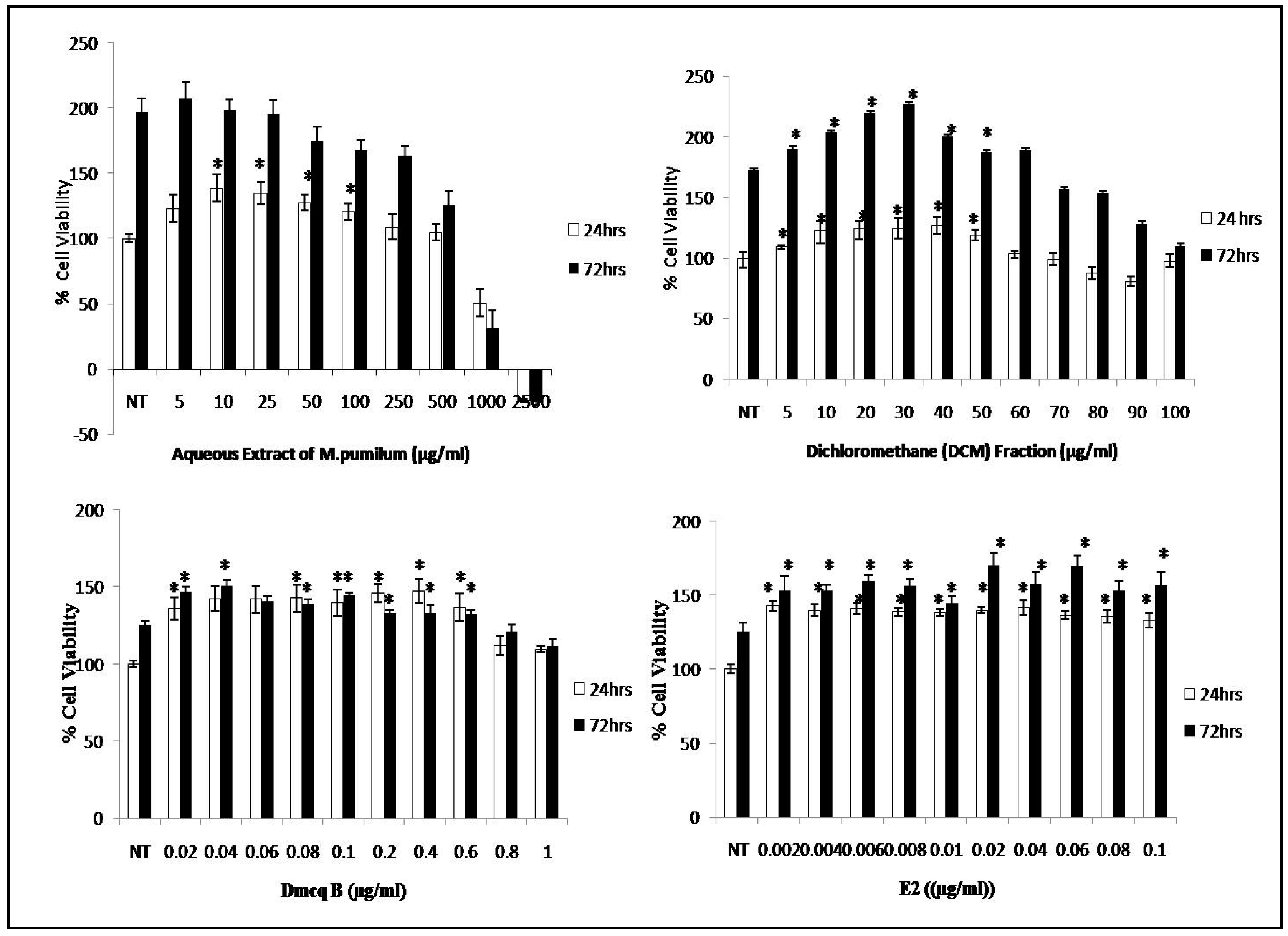
2.2. Promotion of Viable Cells with M. pumilum var. alata Treatments
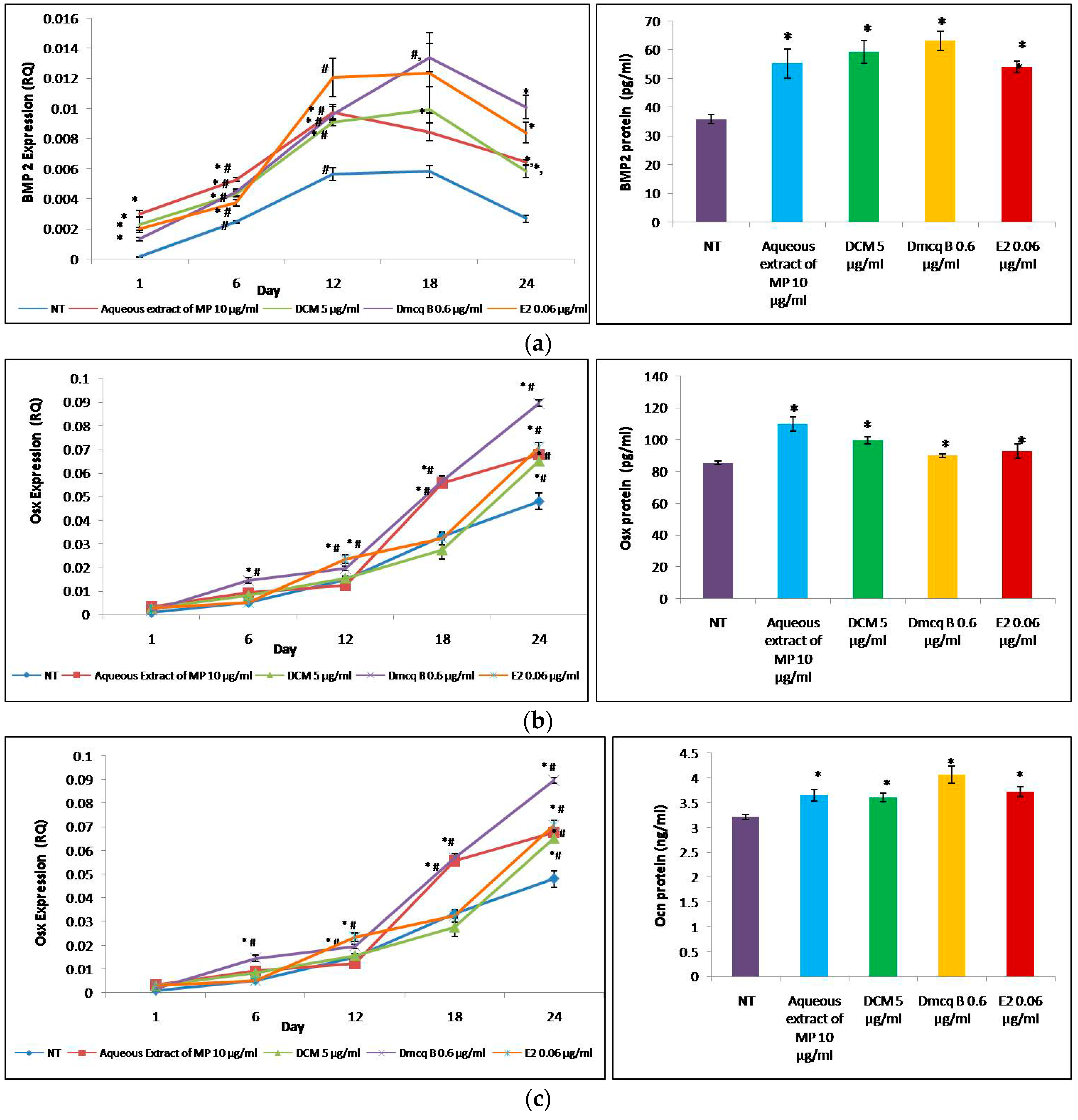
2.3. Modulation of Crude Extract, DCM Fraction and Dmcq B on Early Markers of Osteoblast Differentiation
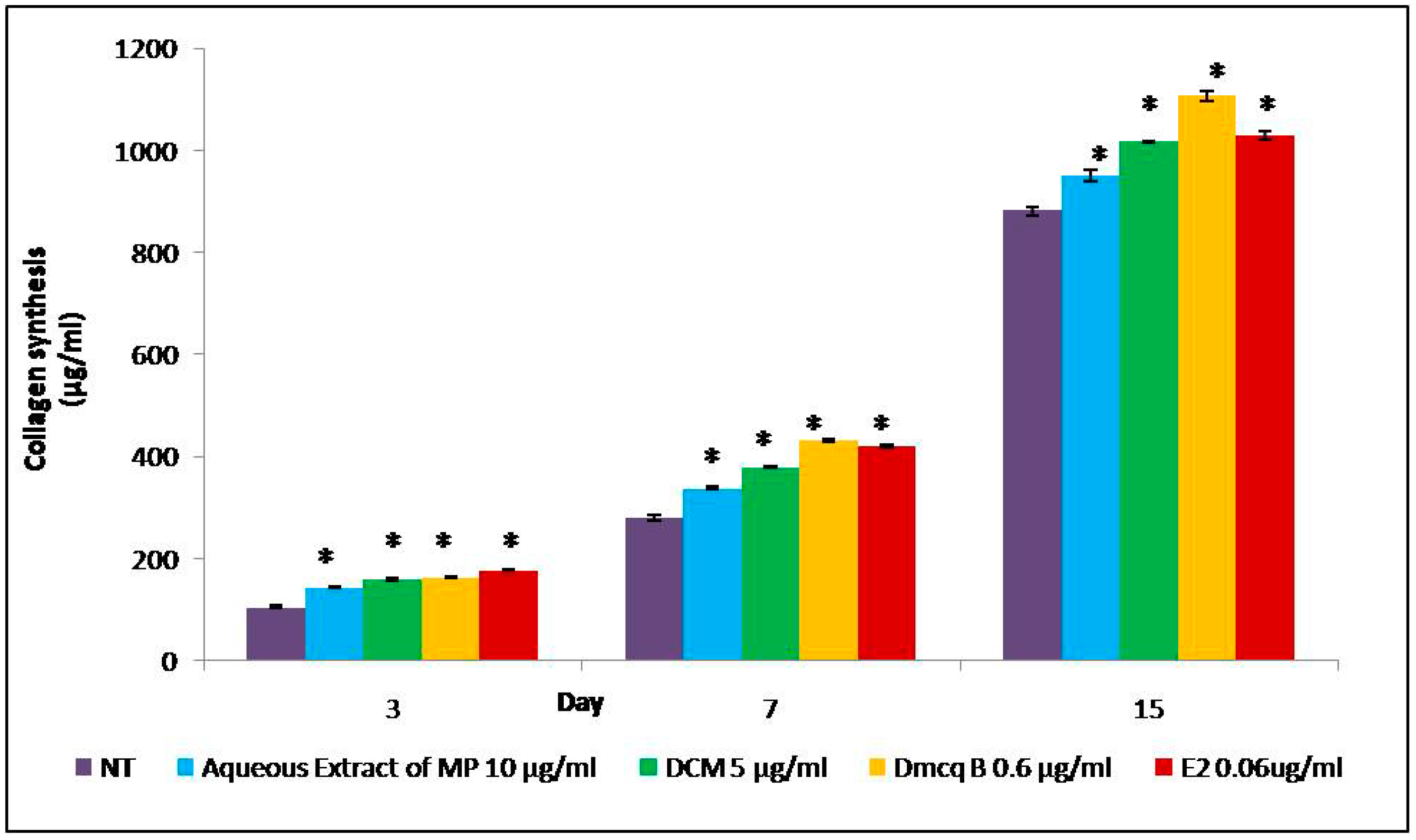
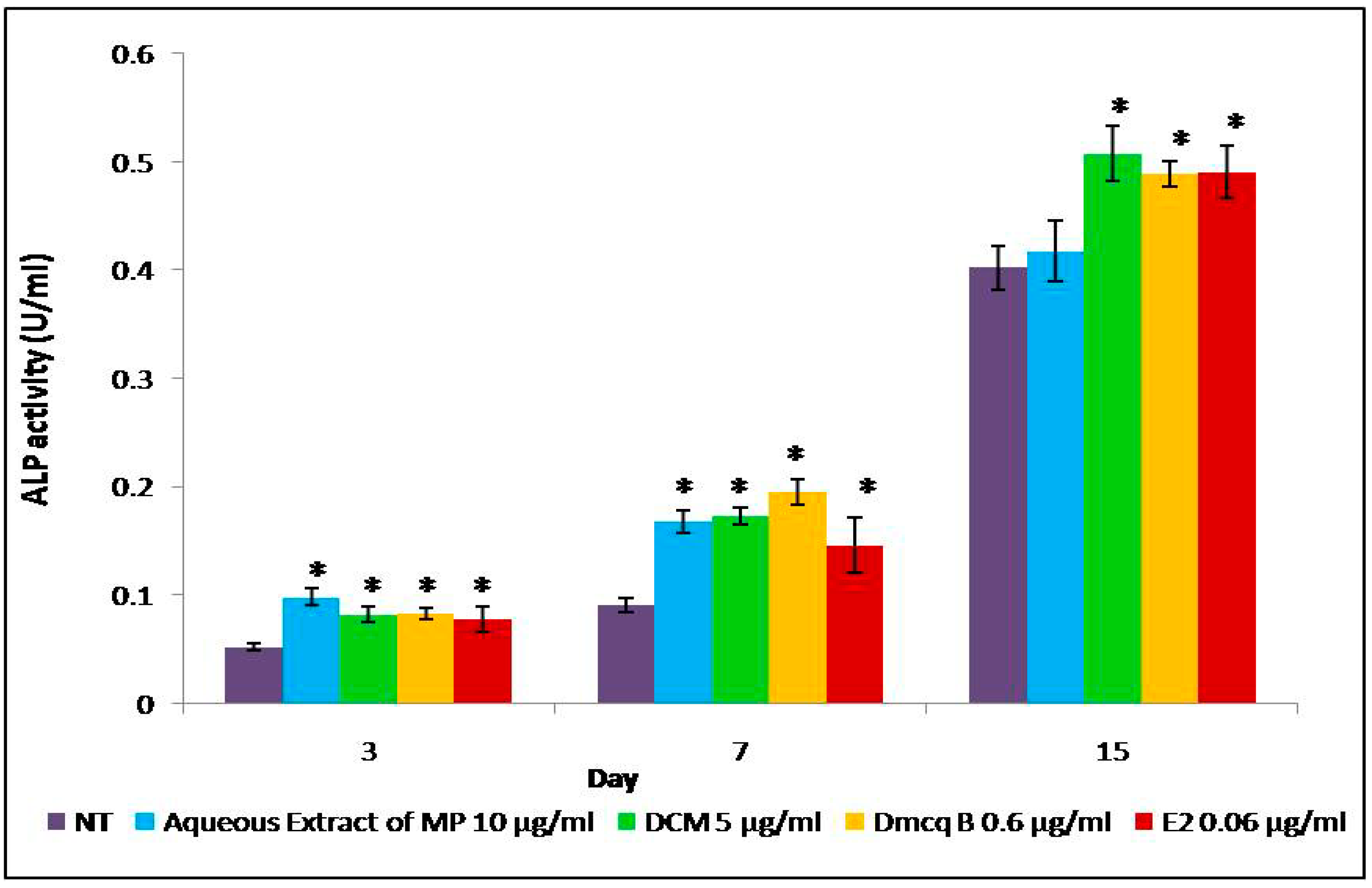
2.4. Modulation of Crude Extract, DCM Fraction and Dmcq B on Osteoblast Differentiation
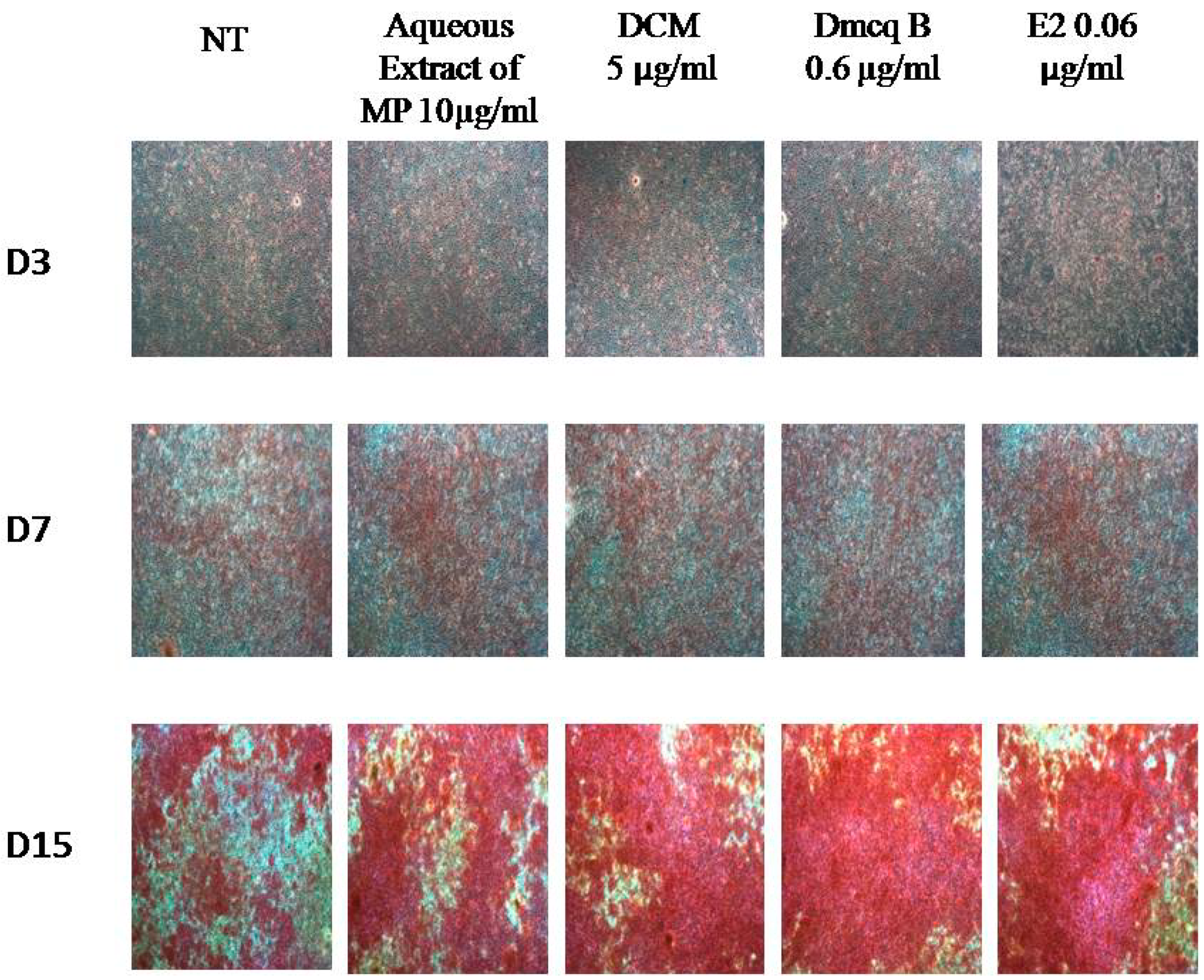
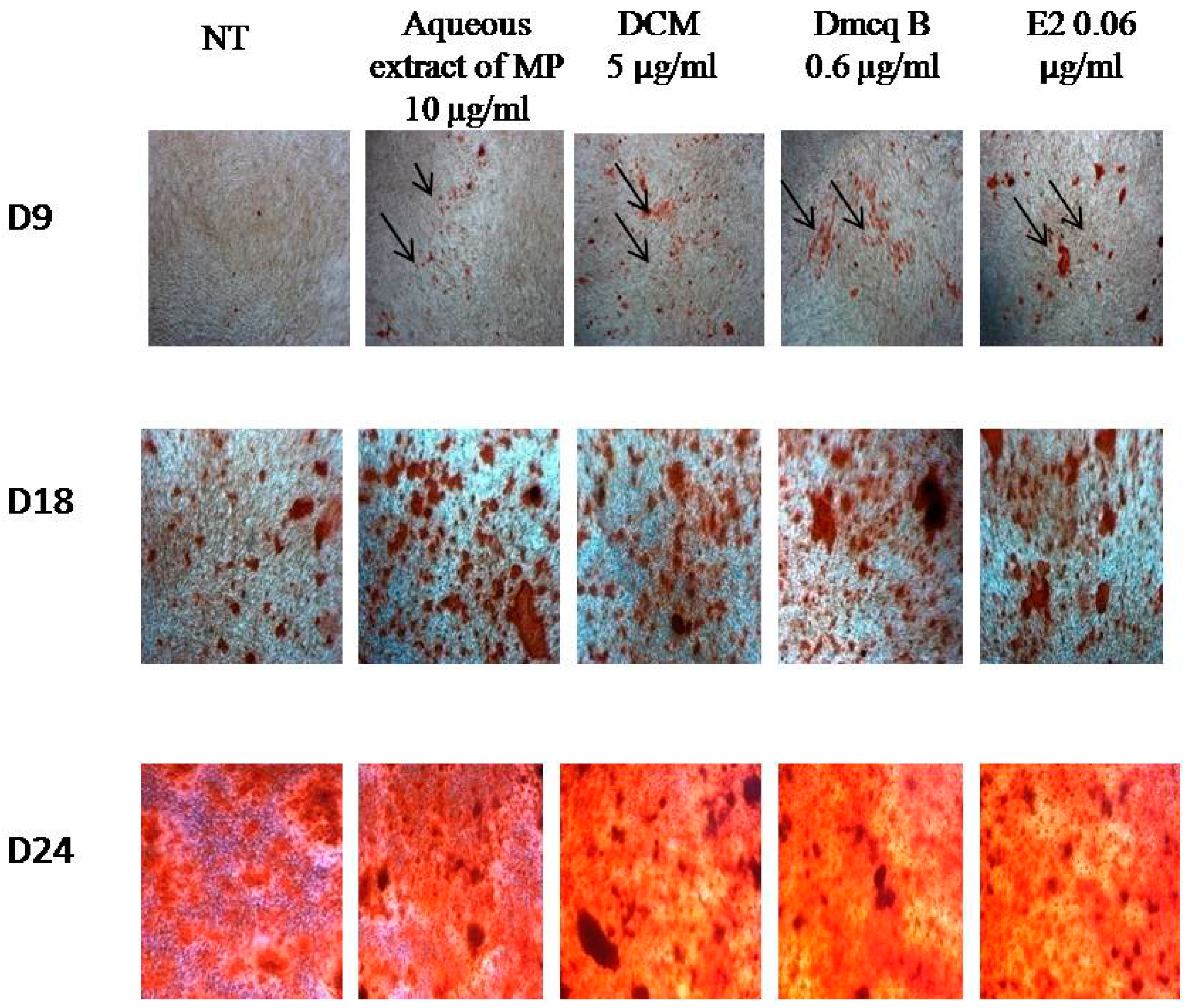
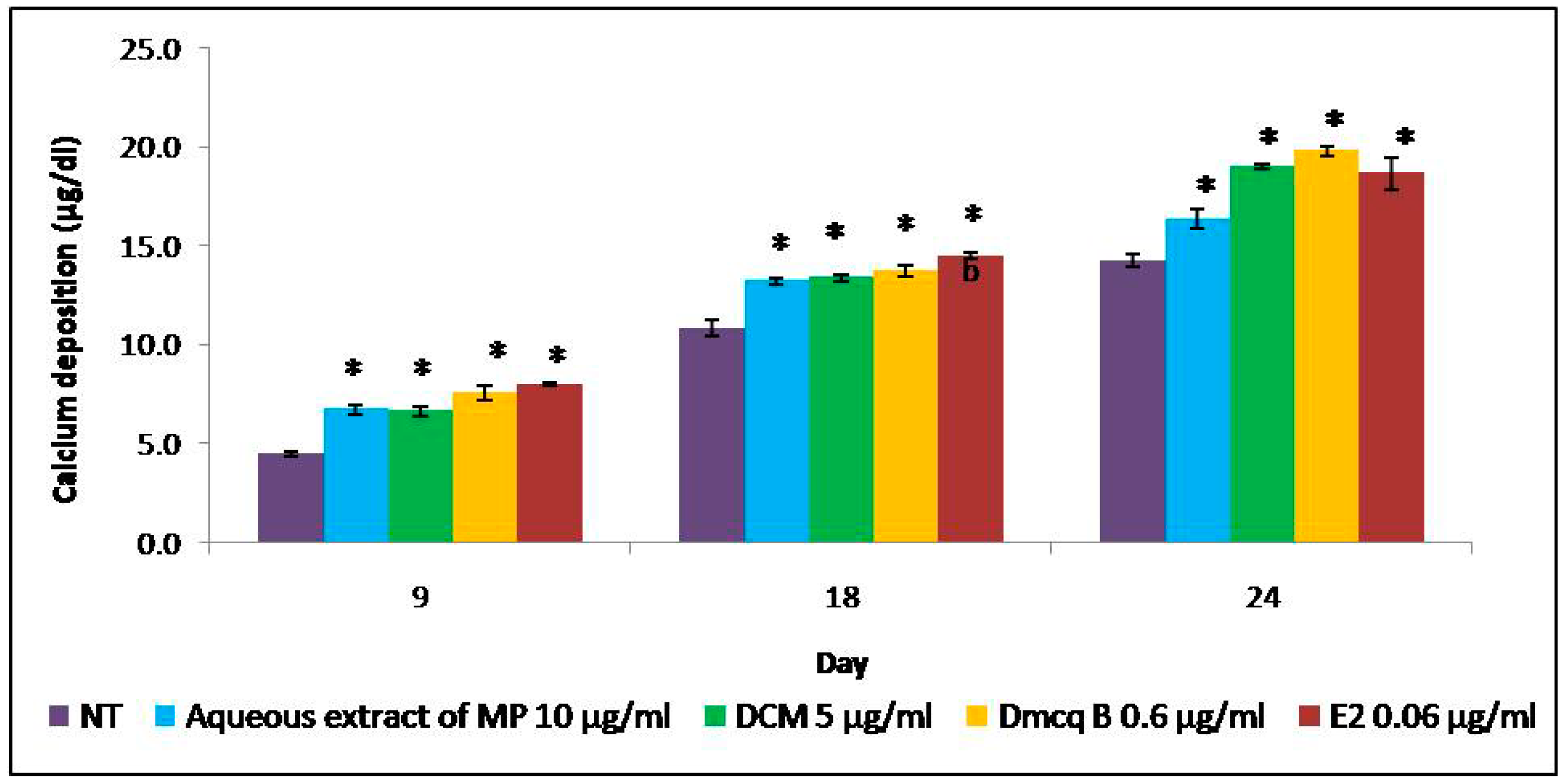
2.5. Stimulation Effects of Crude Extract, DCM Fraction and Dmcq B in Mineralization
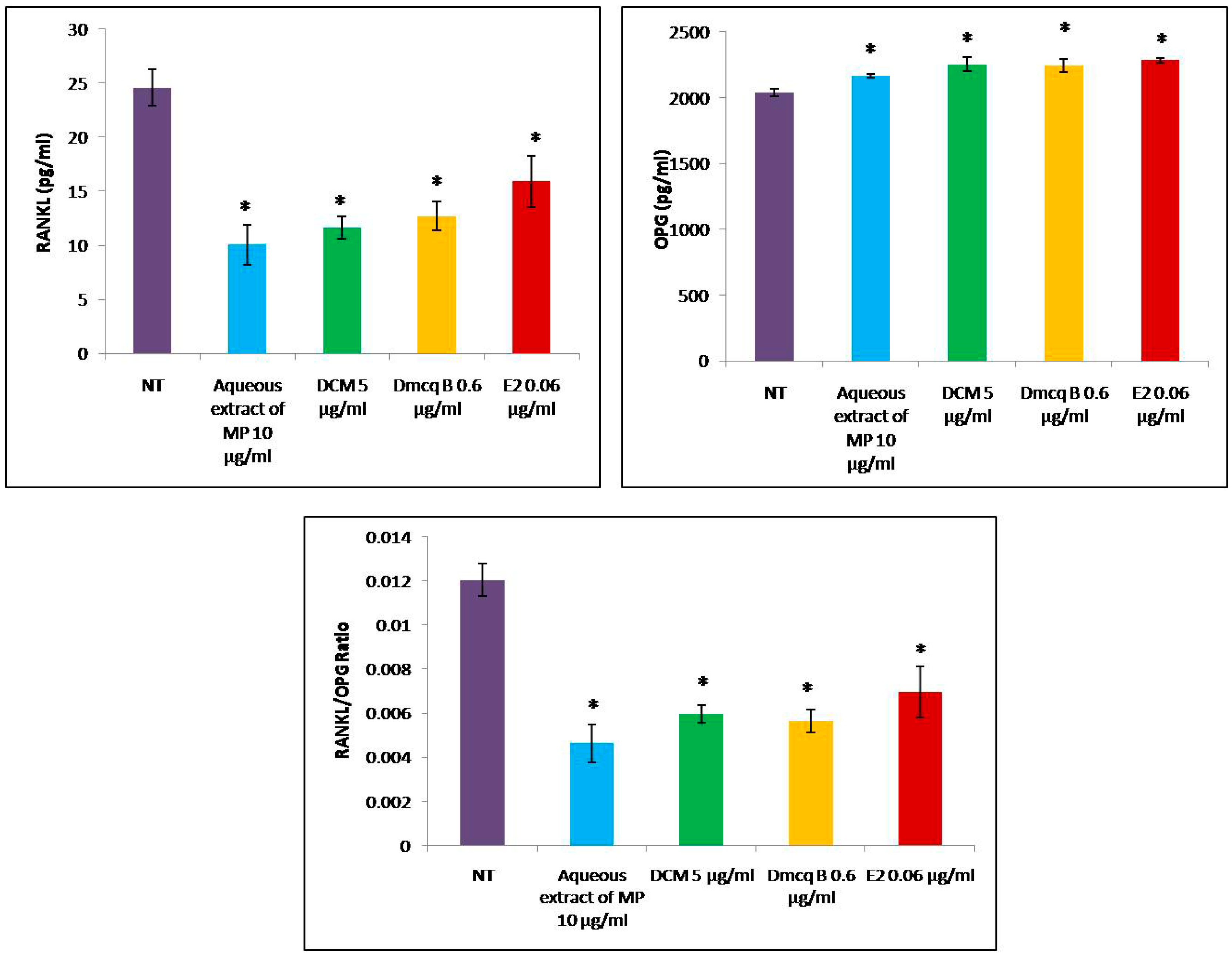
2.6. Reduction of RANKL/OPG Ratio by Crude Extract, DCM Fraction and Dmcq B
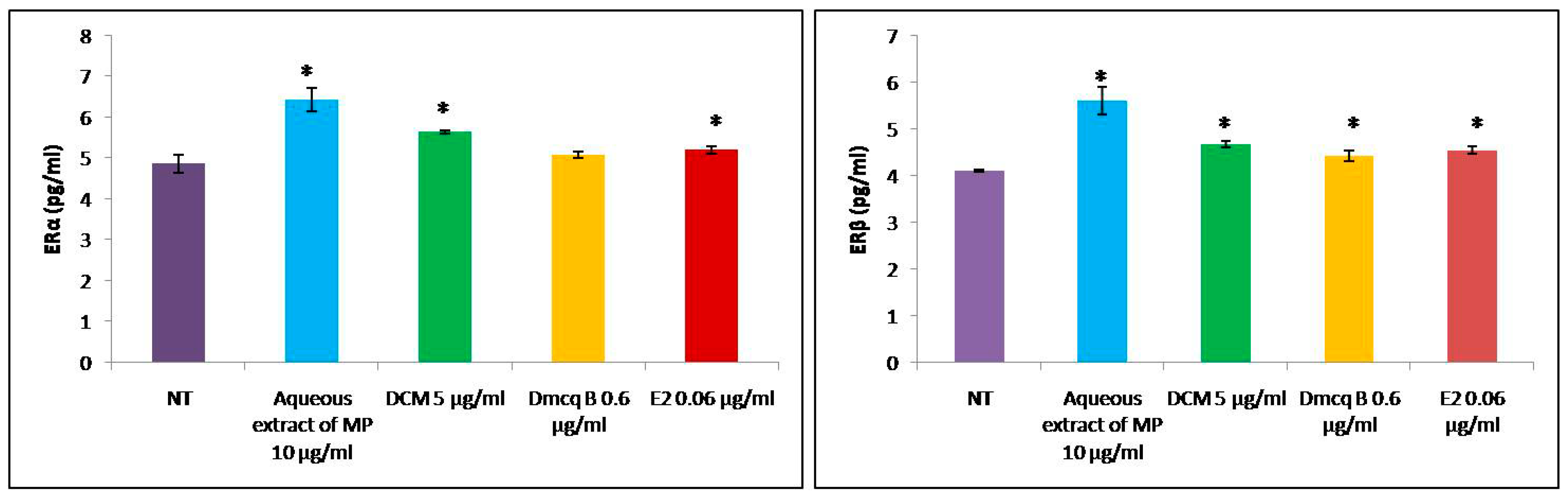
2.7. Effects of Crude Extract, DCM Fraction and Dmcq B on ER Protein Expression
3. Discussion
4. Materials and Methods
4.1. Isolation and Identification of M. pumilum var. alata Active Compound
4.2. Cell Culture
4.3. Determination of Cell Viability
4.4. Collagen Content
4.5. ALP Activity
4.6. Quantitative Analysis of Gene Expression by Real-Time RT-PCR
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Mineralization
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allen, M.R.; Burr, D.V. Bone and bone remodelling. In Basic and Applied Bone Biology; Elsevier: New York, NY, USA, 2014; pp. 75–90. [Google Scholar]
- Drake, T.; Clarke, B.; Lewiecki, E.M. The pathophysiology and treatment of osteoporosis. Clin. Ther. 2015, 37, 1837–1850. [Google Scholar] [CrossRef] [PubMed]
- Lobo, R.A. Hormone-replacement therapy: Current thinking. Nat. Rev. Endocrinol. 2017, 13, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Fisher, B.; Costantino, J.P.; Wickerham, D.L.; Redmond, C.K.; Kavanah, M.; Cronin, M.W.; Vogel, V.; Robidoux, A.; Dimitrov, N.; Atkins, J.; et al. Tamoxifen for prevention of breast cancer: Report of the National Surgical Adjuvant Breast and Bowel Project P-1 Study. J. Natl. Cancer Inst. 1998, 90, 1371–1388. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, B.; Black, D.M.; Mitlak, B.H.; Knickerbocker, R.K.; Nickelsen, T.; Genant, H.K.; Christiansen, C.; Delmas, P.D.; Zanchetta, J.R.; Stakkestad, J.; et al. Reduction of vertebral fracture risk in postmenopausal women with osteoporosis treated with raloxifene: Results from a 3-year randomized clinical trial. Multiple Outcomes of Raloxifene Evaluation (MORE) Investigators. JAMA 1999, 282, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Silverman, S.L.; Christiansen, C.; Genant, H.K. Efficacy of bazedoxifene in reducing new vertebral fracture risk in postmenopausal women with osteoporosis: Results from a 3-year, randomized, placebo-, and active-controlled clinical trial. J. Bone Miner. Res. 2008, 23, 1923–1934. [Google Scholar] [CrossRef] [PubMed]
- Cummings, S.R.; Ensrud, K.; Delmas, P.D.; LaCroix, A.Z.; Vukicevic, S.; Reid, D.M.; Goldstein, S.; Sriram, U.; Lee, U.; Thompson, J.; et al. Lasofoxifene in postmenopausal women with osteoporosis. N. Engl. J. Med. 2010, 362, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Bedell, S.; Nachtigall, M.; Naftolin, F. The pro and cons of plant estrogens for menopause. J. Steroid Biochem. Mol. Biol. 2014, 139, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Atteritano, M.; Mazzafero, S.; Frisina, A.; Cannata, M.L.; Bitto, A.; D’ Anna, R.; Squadrito, F.; Frisina, N. Genistein effects on quantitative ultrasound parameters and bone mineral density in postmenopausal women. Osteoporos. Int. 2009, 20, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Kumar, D.; Dubey, P.; Chandanan, A.; Singh, P. The effects of isoflavones on bone density in postmenopausal women at tertiary centre. Int. J. Reprod. Contracept. Obstet. Gynecol. 2015, 4, 1763–1765. [Google Scholar] [CrossRef]
- Guo, A.J.; Choi, R.C.; Zheng, K.Y.; Chen, V.P.; Dong, T.T.; Wang, Z.; Vollmer, G.; Lau, D.T.; Tsim, K.W. Kaempferol as a flavonoid induces osteoblastic differentiation via estrogen receptor signaling. Chin. Med. 2012, 7, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, M.; Yamamoto, H.; Sato, T.; Mizuha, Y.; Kawai, Y.; Taketani, Y.; Kato, S.; Terao, J.; Inakuma, T.; Takeda, E. Dietary quercetin inhibits bone loss without effect on the uterus in ovariectomized mice. J. Bone Miner. Metab. 2009, 27, 673–681. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Kawaguchi, K.; Kiyama, R. Differential and directional estrogenic signaling pathways induced by enterolignans and their precursors. PLoS ONE 2017, 12, e0171390. [Google Scholar] [CrossRef] [PubMed]
- Chiou, W.; Lee, C.; Liao, J.; Chen, C. 8-Prenylkaempferol accelerates osteoblast maturation through bone morphogenetic protein-2/p38 pathway to activate Runx2 transcription. Life Sci. 2011, 88, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Quarles, L.D.; Song, T.; Pan, W.; Zhou, H.; Xiao, Z. Resveratrol enhances proliferation and osteoblastic differentiation in human mesenchymal stem cells via ER-dependent ERK1/2 activation. Phytomedicine 2007, 14, 806–814. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, M.; Mohd, M.A. Traditional Malay Medicinal Plants; Fajar Bakti: Kuala Lumpur, Malaysia, 1994. [Google Scholar]
- Avula, B.; Wang, Y.H.; Ali, Z.; Smillie, T.J.; Khan, I.A. Quantitative determination of triperpene saponins and alkenated-phenolics from Labisia pumila using an LCUV/ELSD method and confirmation by LC-ESI-TOF. Planta Med. 2011, 77, 1742–1748. [Google Scholar] [CrossRef] [PubMed]
- Hisham, D.M.N.; Mohd Lip, J.; Mohd Noh, J.; Normah, A.; Nurul Nabilah, M.F. Identification and isolation of methyl gallate as a polar chemical marker for Labisia pumila Benth. J. Trop. Agric. Food Sci. 2011, 39, 279–284. [Google Scholar]
- Karimi, E.; Jaafar, H.Z.E. HPLC and GC–MS determination of bioactive compounds in microwave obtained extracts ofthree varieties of Labisia pumila Benth. Molecules 2011, 16, 6791–6805. [Google Scholar] [CrossRef] [PubMed]
- Ali, Z.; Khan, I.A. Alkyl phenols and saponins from the roots of Labisia pumila (Kacip Fatimah). Phytochemistry 2011, 72, 2075–2080. [Google Scholar] [CrossRef] [PubMed]
- Al-Mekhlafi, N.A.; Shaari, K.; Abas, F.; Kneer, R.; Jeyaraj, E.J.; Stanslas, J.; Yamamoto, N.; Honda, T.; Lajis, N.H. Alkenylresorcinols and cytotoxic activity of the constituents isolated from Labisia pumila. Phytochemistry 2012, 80, 42–44. [Google Scholar] [CrossRef] [PubMed]
- Karimi, E.; Hawa, Z.E.J.; Ahmad, S. Phytochemical analysis and antimicrobial activities of methanolic extracts of leaf, stem and root from different varieties of Labisia pumila Benth. Molecules 2011, 16, 4438–4450. [Google Scholar] [CrossRef] [PubMed]
- Karimi, E.; Jaafar, H.Z.E.; Ahmad, S. Antifungal anti-inflammatory and cytotoxicity activities of three varieties of Labisia pumila Benth: From microwave obtained extracts. BMC Complement. Altern. Med. 2013, 13, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuagam, L.; Okechukwu, P.N. The study of antinociceptive effects of MeOH and DCM crude extracts from the leaves of Labisia pumila. Glob. J. Pharm. 2011, 5, 181–185. [Google Scholar]
- Husniza, H. Estrogenic and Androgenic Activities of Kacip Fatimah (Labisia pumila); Institute of Medical Research, Ministry of Health Malaysia: Kuala Lumpur, Malaysia, 2002; p. 8. [Google Scholar]
- Wahab, N.; Yusoff, W.; Shuib, A.; Wan, N.; Khatiza, H. Labisia pumila has similar effects to estrogen on the reproductive hormones of ovariectomized rats. Int. J. Herbal Plant Med. 2011, 2, 1–6. [Google Scholar]
- Ayida, A.; Wan Nazaimoon, W.M.; Norsyam, W.N.; Farihah, H.S.; Azian, A.L. Effect of water extract of Labisia pumila var. alata on aorta of ovariectomized Sprague Dawley rats. Pak. J. Nutr. 2008, 7, 208–213. [Google Scholar]
- Fazliana, M.; Gu, H.F.; Östenson, C.-G.; Yusoff, M.M.; Wan Nazaimoon, W.M. Labisia pumila extract down-regulates hydroxysteroid (11-beta) dehydrogenase 1 expression and corticosterone levels in ovariectomized rats. J. Nat. Med. 2012, 66, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Effendy, N.M.; Khamis, M.F.; Soelaiman, I.N.; Shuid, A.N. The effects of Labisia pumila on postmenopausal osteoporotic rat model: Dose and time-dependent micro-CT analysis. J. X-ray Sci. Technol. 2014, 22, 503–518. [Google Scholar]
- Shuid, A.N.; Ping, L.L.; Muhammad, N.; Mohamed, N.; Soelaiman, I.M. The effects of Labisia pumila var. alata on bone markers and bone calcium in a rat model of post-menopausal osteoporosis. J. Ethnopharmacol. 2011, 133, 538–542. [Google Scholar]
- Fathilah, S.N.; Nazrun Shuid, A.; Mohamed, N.; Muhammad, N.; Soelaiman, I.N. Labisia pumila protects the bone of estrogen-deficient rat model: A histomorphometric study. J. Ethnopharmacol. 2012, 142, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Fathilah, S.N.; Mohamed, N.; Muhammad, N.; Mohamed, I.N.; Soelaiman, I.N.; Shuid, A.N. Labisia pumila regulates bone-related genes expressions in postmenopausal osteoporosis model. BMC Complement. Altern. Med. 2013, 13, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Effendy, N.M.; Shuid, A.N. Time and dose-dependent effects of Labisia pumila on bone oxidative status of postmenopausal osteoporosis rat model. Nutrients 2014, 6, 3288–3302. [Google Scholar] [CrossRef] [PubMed]
- Sirotkin, A.; Harrath, A.H. Phytoestrogen and their effects. Eur. J. Pharmacol. 2014, 741, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Bao, L.; Qin, L.; Liu, L. Anthraquinone compounds from Morinda officinalis inhibit osteoclastic bone resorption in vitro. Chem.-Biol. Interact. 2011, 194, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.U.; Shin, H.K.; Min, Y.K.; Kim, S.H. Emodin accelerates osteoblast differentiation through phosphatidylinositol 3-kinase activation and bone morphogenetic protein-2 gene expression. Int. Immunopharmacol. 2008, 8, 741–747. [Google Scholar] [CrossRef] [PubMed]
- Wirries, A.; Schubert, A.; Zimmermann, R.; Jabari, S.; Rucholtz, S.; El-Najjar, N. Thymoquinone accelerates osteoblast differentiation and activates bone morphogenetic protein-2 end ERK pathway. Int. Immunopharmacol. 2013, 15, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Dianita, R.; Jantan, I.; Amran, A.Z.; Jalil, J. Protective effects of Labisia pumila var. alata on biochemical and histopathological alterations of cardiac muscle cells in isoproterenol-induced myocardial infarction rats. Molecules 2015, 20, 4746–4763. [Google Scholar] [PubMed]
- Czekanska, E.M.; Stoddart, M.J.; Richards, R.G.; Hayes, J.S. In search of an osteoblast cell model for in vitro research. Eur. Cells Mater. 2012, 24, 1–17. [Google Scholar] [CrossRef]
- Chung, H.; Kim, W.K.; Oh, J.; Kim, M.; Shin, J.; Lee, J.; Ha, I.; Lee, S.K. Anti-osteoporotic activity of harpagoside by up-regulation of the BMP2 and Wnt signaling pathways in osteoblasts and suppression of differentiation in osteoclast. J. Nat. Prod. 2017, 80, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Haase, H.; Young, W.G.; Bartold, M. Development and transplantation of a mineralized matrix formed by osteoblasts in vitro for bone regeneration. Cell Transplant. 2004, 13, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C. Transcriptional regulation of bone formation by the osteoblast-specific transcription factor Osx. J. Orthop. Surg. Res. 2010, 5, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, X.; Zhang, Z.; Feng, J.Q.; Dusevich, V.M.; Sinha, K.; Zhang, H.; Darnay, B.G.; de Crombrugghe, B. Multiple functions of Osterix are required for bone growth and homeostasis in postnatal mice. Proc. Natl. Acad. Sci. USA 2010, 107, 12919–12924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoch, M.L.; Clemens, T.L.; Riddle, R.C. New insights into the biology of osteocalcin. Bone 2015, 82, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.; Stein, G. Concepts of osteoblast growth and differentiation: Basis for modulation of bone cell development and tissue formation. Crit. Rev. Oral Biol. Med. 1992, 3, 269–305. [Google Scholar] [CrossRef] [PubMed]
- Balcerzak, M.; Hamade, E.; Zhang, L.; Pikula, S.; Azzar, G.; Raddison, J.; Pikula, J.B.; Buchet, R. The Roles of annexins and alkaline phosphatase in mineralization process. Acta Biochim. Pol. 2003, 50, 1019–1038. [Google Scholar] [PubMed]
- Tsao, Y.T.; Huang, Y.J.; Wu, H.H.; Liu, Y.A.; Liu, Y.S.; Lee, O.K. Osteocalcin mediates biomineralization during osteogenic maturation in human mesenchymal stromal cells. Int. J. Mol. Sci. 2017, 18, 159–172. [Google Scholar] [CrossRef] [PubMed]
- Weitzmann, M.N. The role of inflammatory cytokines, the RANKL/OPG axis, and the immunoskeletal interface in physiological bone turnover and osteoporosis. Scientifica 2013, 2013, 125705. [Google Scholar] [CrossRef] [PubMed]
- Canciani, E.; Dellavia, C.; GioiaMarazzi, M.; Augusti, D.; Carmagnola, D.; Vianello, E.; Canullo, L.; Galliera, E. RNA isolation from alveolar bone and gene expression analysis of RANK, RANKL and OPG: A new tool to monitor bone remodeling and healing in different bone substitutes used for prosthetic rehabilitation. Arch. Oral Biol. 2017, 80, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Bord, S.; Horner, A.; Beavan, S.; Compson, J. Estrogen receptors alpha and beta are differentially expressed in developing human bone. J. Clin. Endocrinol. Metab. 2001, 86, 2309–2314. [Google Scholar] [PubMed]
- Khalid, A.B.; Krum, S.A. Estrogen receptors alpha and beta in bone. Bone 2016, 87, 103–135. [Google Scholar] [CrossRef] [PubMed]
- Jordan, V.C.; MacGregor Schafer, J.I.; Levenson, A.S.; Liu, H.; Pease, K.M.; Simons, L.A.; Zapf, J.W. Molecular classification of estrogens. Cancer Res. 2001, 61, 6619–6623. [Google Scholar] [PubMed]
- Bourgoin-Voillard, S.; Gallo, D.; Laïos, I.; Cleeren, A.; Bali, L.E.; Jacquot, Y.; Nonclercq, D.; Laurent, G.; Tabet, J.-C.; Leclercq, G. Capacity of type I and II ligands to confer to estrogen receptor alpha an appropriate conformation for the recruitment of coactivators containing a LxxLL motif-Relationship with the regulation of receptor level and ERE-dependent transcription in MCF-7 cells. Biochem. Pharmacol. 2010, 79, 746–757. [Google Scholar] [PubMed]
- Han, D.H.; Denison, M.S.; Tachibana, H.; Yamada, K. Relationship between estrogen receptor-binding and estrogenic activities of environmental estrogens and suppression by flavonoids. Biosci. Biotechnol. Biochem. 2002, 66, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.Y.; Yang, H.J.; Song, Y.M.; Kim, I.S.; Hwang, S.J. Estrogen modulates bone morphogenetic protein-induced sclerostin expression through the Wnt signaling pathway. Tissue Eng. Part A 2015, 21, 2076–2088. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Li, L.; Kim, H.J. Fibrillar assembly and stability of collagen coating on titanium for improved osteoblast responses. J. Biomed. Mater. Res. 2005, 75, 629–638. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No | Primer Sequence (5′-3′) | Product Size (bp) |
|---|---|---|---|
| β-actin | NM_007393.5 | F: gaagagctatgagctgcctga R: gcactgtgttggcatagaggt | 185 |
| BMP2 | NM_007561.4 | F: gtgccctggctgctatgg R: tgccgcctccatcatgtt | 546 |
| Osx | NM_130458.3 | F: gcaagaggttcactcgctct R: gtggtcgcttctggtaaagc | 110 |
| Ocn | NM_031368 | F: gcgctctgtctctctgacct R: aagcagggtcaagctcacat | 181 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad Hairi, H.; Jamal, J.A.; Aladdin, N.A.; Husain, K.; Mohd Sofi, N.S.; Mohamed, N.; Mohamed, I.N.; Shuid, A.N. Demethylbelamcandaquinone B (Dmcq B) Is the Active Compound of Marantodes pumilum var. alata (Blume) Kuntze with Osteoanabolic Activities. Molecules 2018, 23, 1686. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23071686
Ahmad Hairi H, Jamal JA, Aladdin NA, Husain K, Mohd Sofi NS, Mohamed N, Mohamed IN, Shuid AN. Demethylbelamcandaquinone B (Dmcq B) Is the Active Compound of Marantodes pumilum var. alata (Blume) Kuntze with Osteoanabolic Activities. Molecules. 2018; 23(7):1686. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23071686
Chicago/Turabian StyleAhmad Hairi, Haryati, Jamia Azdina Jamal, Nor Ashila Aladdin, Khairana Husain, Noor Suhaili Mohd Sofi, Norazlina Mohamed, Isa Naina Mohamed, and Ahmad Nazrun Shuid. 2018. "Demethylbelamcandaquinone B (Dmcq B) Is the Active Compound of Marantodes pumilum var. alata (Blume) Kuntze with Osteoanabolic Activities" Molecules 23, no. 7: 1686. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules23071686