Effects of Diethyl Phosphate, a Non-Specific Metabolite of Organophosphorus Pesticides, on Serum Lipid, Hormones, Inflammation, and Gut Microbiota


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
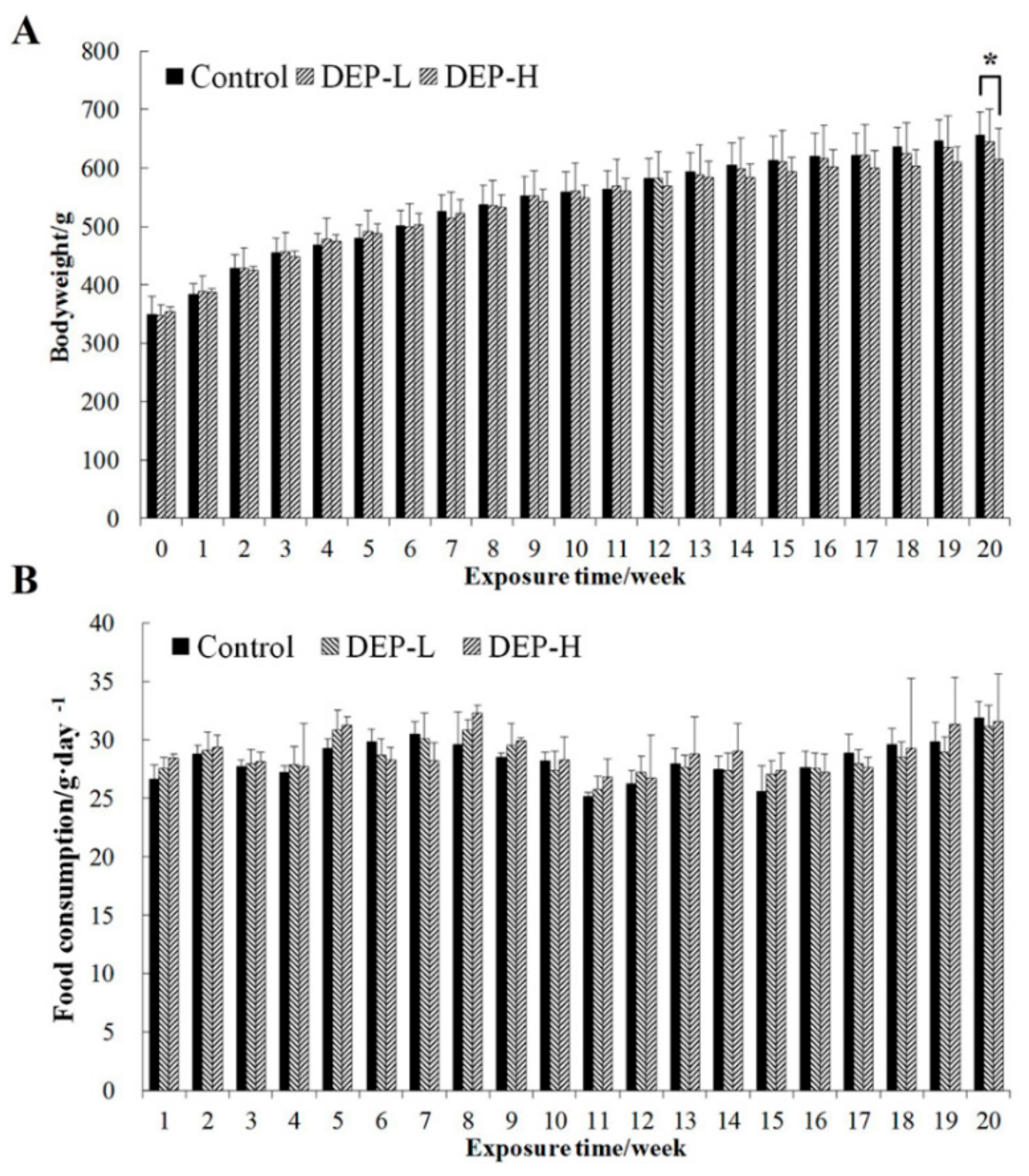
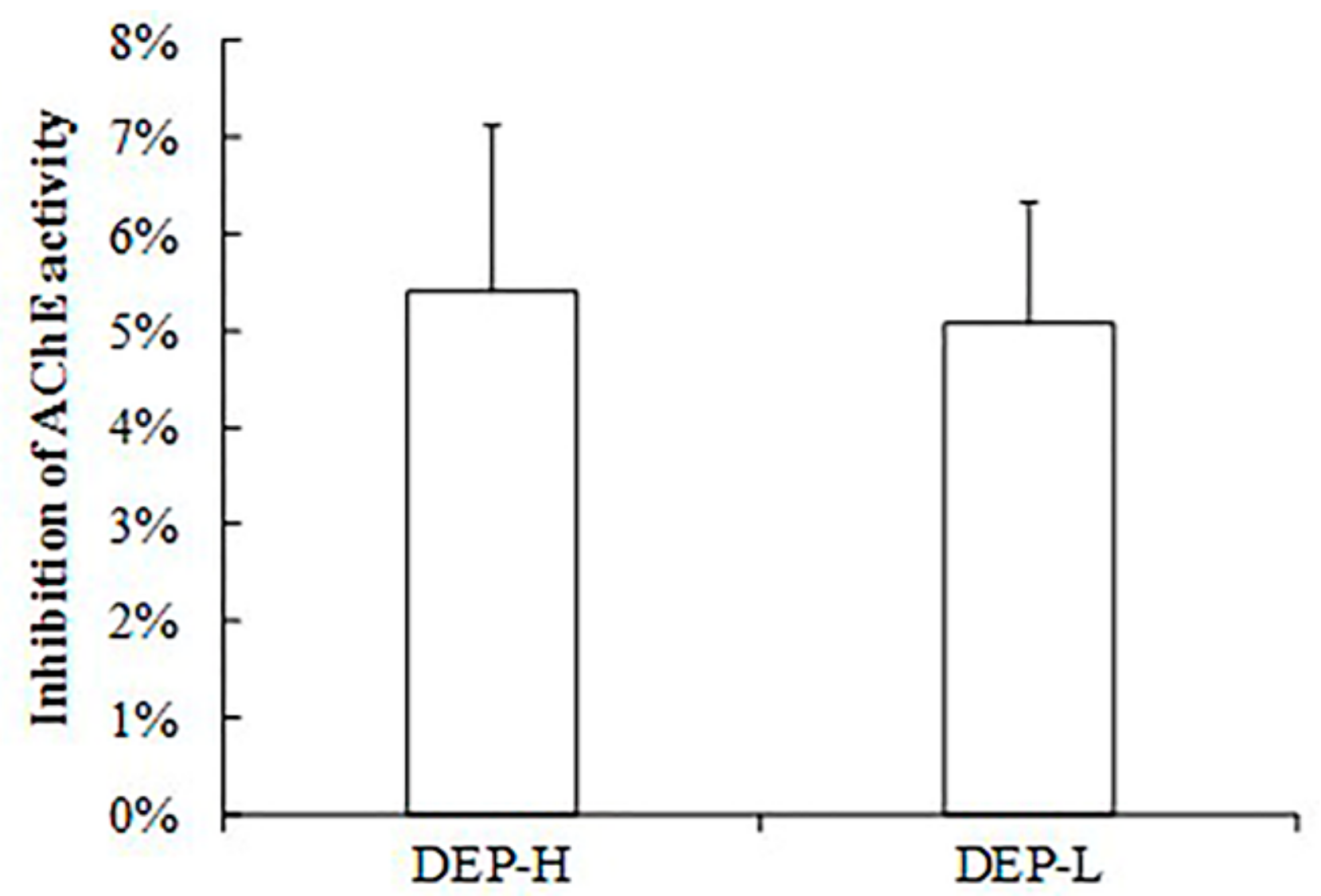
2.1. Effects of DEP Exposure on Body Weight and Food Composition
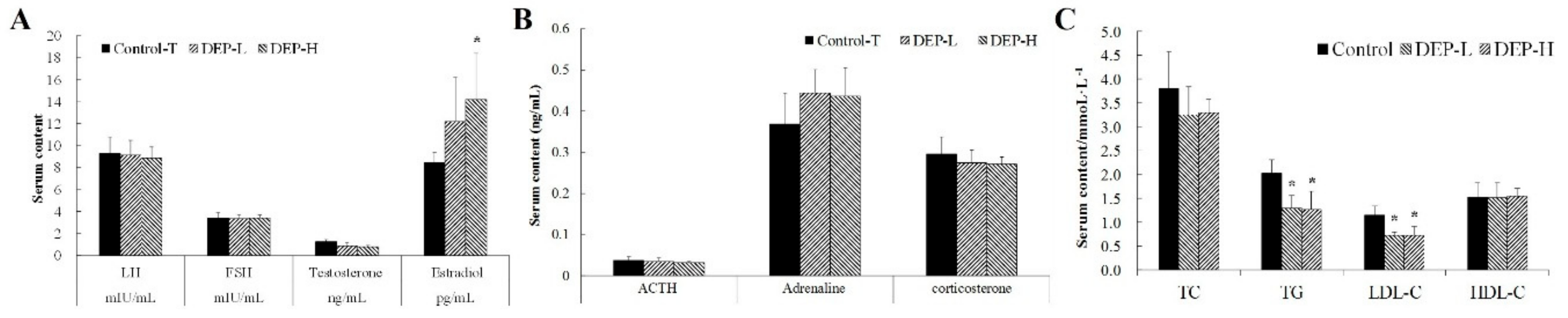
2.2. Effects of DEP Exposure on Serum Levels of Hormones and Lipids
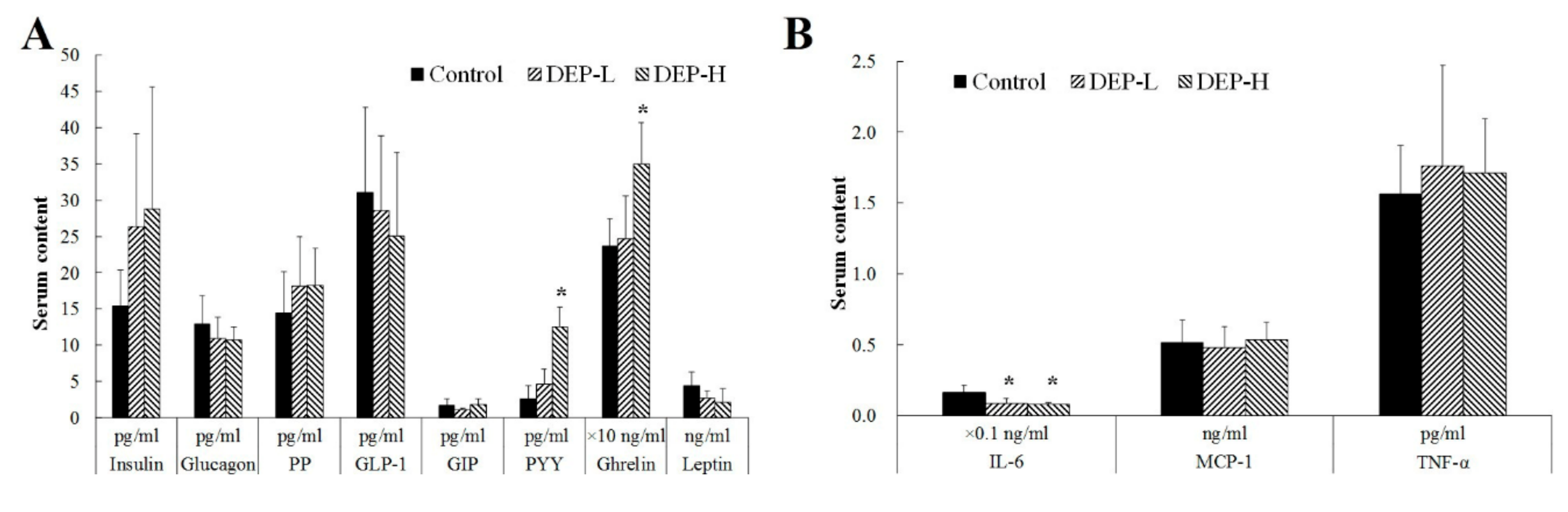
2.3. Effects of DEP on Gut Endocrine Cells and the Immune System
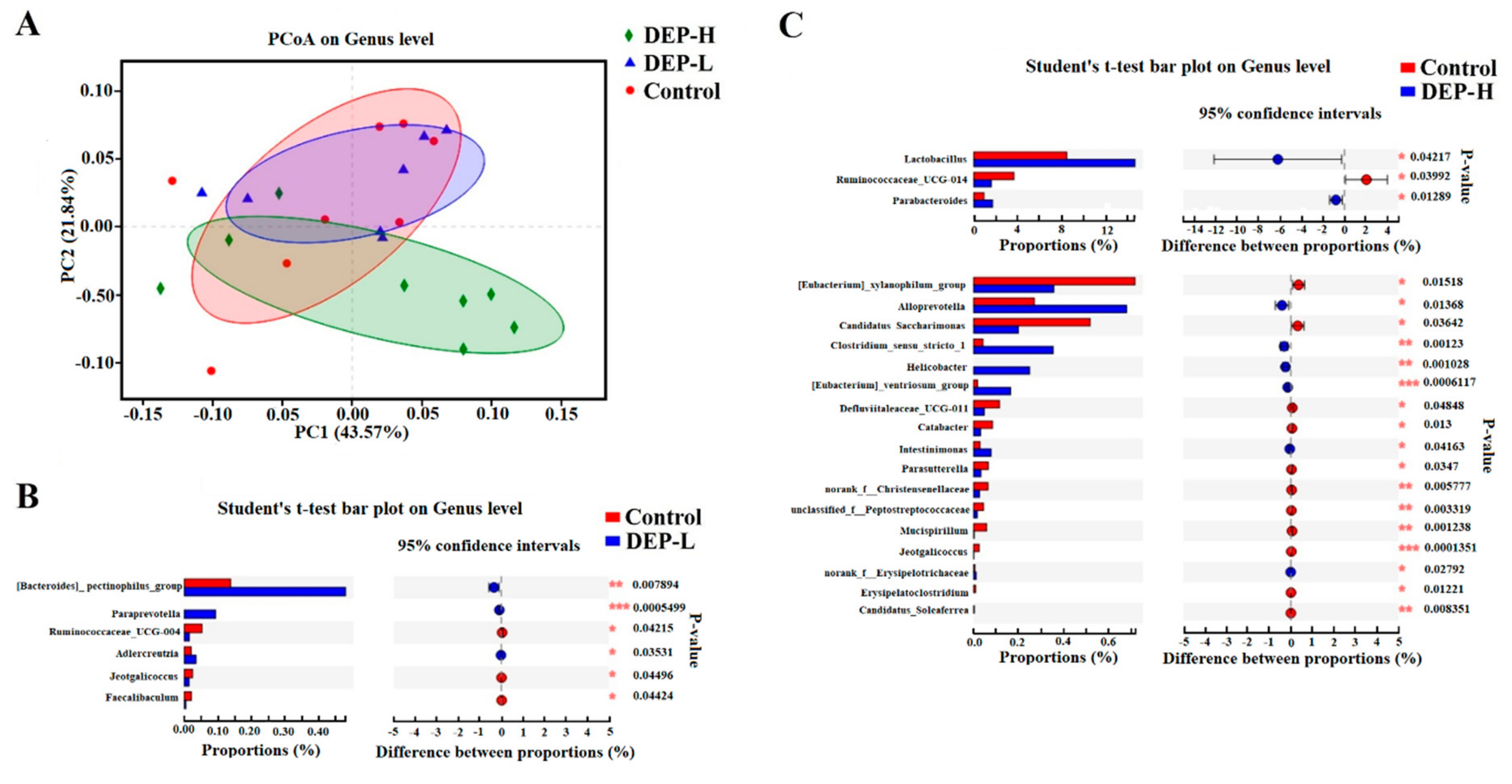
2.4. Effects of DEP on Gut Microbiota
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Fecal Sample Collection and Bacterial DNA Extraction
4.3. AChE Activity Measurements
4.4. Serum Lipid Profiles
4.5. Hormones and Cytokine Measurements
4.6. S rRNA Gene Sequencing
4.7. Gut Microbiota Analysis
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- USDA (United States Department of Agriculture). Pesticide Data Program: Annual summary, calendar year 2011. Available online: http://www.ams.usda.gov/AMSv1.0/getfile?dDocName=stelprdc5102692 (accessed on 3 October 2018).
- PRiF (Expert Committee on Pesticide Residues in Food). Pesticide residues monitoring programme for quarter 1 2013. Available online: http://www.pesticides.gov.uk/Resources/CRD/PRiF/Documents/Results%20and%20Reports/2013/2013%20Q1%20Final.pdf. (accessed on 3 December 2013).
- FSANZ (Food Standards Australia New Zealand). The 23rd Australian Total Diet Survey. Available online: http://www.foodstandards.gov.au/publications/pages/23rdaustraliantotald5367.aspx (accessed on 18 December 2013).
- Jain, R.B.; Consultant, P.; Georgia, D. Levels of dialkylphosphate metabolites in urine among general U.S. population. Environ. Toxicol. Pharmacol. 2016, 43, 74–82. [Google Scholar] [CrossRef]
- Sudakin, D.L.; Stone, D.L. Dialkyl phosphates as biomarkers of organophosphates: The current divide between epidemiology and clinical toxicology. Clin. Toxicol. 2011, 49, 771–781. [Google Scholar] [CrossRef] [PubMed]
- Berman, T.; Goldsmith, R.; Göen, T.; Spungen, J.; Novack, L.; Levine, H.; Amitai, Y.; Shohat, T.; Grotto, I. Urinary concentrations of organophosphate pesticide metabolites in adults in Israel: Demographic and dietary predictors. Environ. Int. 2013, 60, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Berman, T.; Göen, T.; Novack, L.; Beacher, L.; Grinshpan, L.; Segev, D.; Tordjman, K. Urinary concentrations of organophosphate and carbamate pesticides in residents of a vegetarian community. Environ. Int. 2016, 96, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Hodgson, E.; Rose, R.L. Organophosphorus Chemicals: Potent Inhibitors of the Human Metabolism of Steroid Hormones and Xenobiotics. Drug Metab. Rev. 2006, 38, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Koutros, S. 389 Risk of total and aggressive prostate cancer and pesticide use in the Agricultural Health Study. Occup. Environ. Med. 2013, 70, 59–74. [Google Scholar] [CrossRef]
- Evangelou, E.; Ntritsos, G.; Chondrogiorgi, M.; Kavvoura, F.K.; Hernández, A.F.; Ntzani, E.E.; Tzoulaki, I. Exposure to pesticides and diabetes: A systematic review and meta-analysis. Environ. Int. 2016, 91, 60–68. [Google Scholar] [CrossRef]
- Miranda-Contreras, L.; Gómez-Pérez, R.; Rojas, G.; Cruz, I.; Berrueta, L.; Salmen, S.; Colmenares, M.; Barreto, S.; Balza, A.; Zavala, L.; et al. Occupational Exposure to Organophosphate and Carbamate Pesticides Affects Sperm Chromatin Integrity and Reproductive Hormone Levels among Venezuelan Farm Workers. J. Occup. Heal. 2013, 55, 195–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar-Garduno, C.; Lacasana, M.; Blanco-Munoz, J.; Rodriguez-Barranco, M.; Hernandez, A.F.; Bassol, S.; Gonzalez-Alzaga, B.; Cebrian, M.E. Changes in male hormone profile after occupational organophosphate exposure. A longitudinal study. Toxicology 2013, 307, 55–65. [Google Scholar] [PubMed]
- Ghisari, M.; Long, M.; Tabbo, A.; Bonefeld-Jørgensen, E.C. Effects of currently used pesticides and their mixtures on the function of thyroid hormone and aryl hydrocarbon receptor in cell culture. Toxicol. Appl. Pharmacol. 2015, 284, 292–303. [Google Scholar]
- Chebab, S.; Mekircha, F.; Leghouchi, E. Potential protective effect of Pistacia lentiscus oil against chlorpyrifos-induced hormonal changes and oxidative damage in ovaries and thyroid of female rats. Biomed. Pharmacother. 2017, 96, 1310–1316. [Google Scholar] [CrossRef]
- Adedara, I.A.; Owoeye, O.; Ajayi, B.O.; Awogbindin, I.O.; Rocha, J.B.; Farombi, E.O. Diphenyl diselenide abrogates chlorpyrifos-induced hypothalamic-pituitary-testicular axis impairment in rats. Biochem. Biophys. Commun. 2018, 503, 171–176. [Google Scholar] [CrossRef]
- Peiris, D.C.; Dhanushka, T. Low doses of chlorpyrifos interfere with spermatogenesis of rats through reduction of sex hormones. Environ. Sci. Pollut. 2017, 24, 20859–20867. [Google Scholar] [CrossRef] [PubMed]
- Mosbah, R.; Yousef, M.I.; Maranghi, F.; Mantovani, A. Protective role of Nigella sativa oil against reproductive toxicity, hormonal alterations, and oxidative damage induced by chlorpyrifos in male rats. Toxicol Ind. Health 2016, 32, 1266–1277. [Google Scholar] [CrossRef] [PubMed]
- Akande, M.G.; Shittu, M.; Uchendu, C.; Yaqub, L.S. Taurine ameliorated thyroid function in rats co-administered with chlorpyrifos and lead. Vet. Res. Commun. 2016, 40, 123–129. [Google Scholar] [CrossRef]
- Ahmad, S.N.; Nasir, M.; Naureen, A.; Andleeb, S.; Raees, K.; Abbas, T.; Younis, A.; Ahmad, K.R. Histopathological and micrometric studies of diazinon exposure on thyroid and parathyroid tissues in mice. Biologia (Pakistan) 2018, 64, 10. [Google Scholar]
- Raees, K.; Ishfaq, R.; Ullah, A.; Tahir, M.Z.; Abbas, T.; Tahir, H.M.; Mukhtar, M.K.; Arshad, M.; Khan, S.Y.; Ahmad, K.R. Histological and micrometric effects of diazinon exposure on adrenal medulla and cortex in mice. J. Appl. 2012, 40, 267–272. [Google Scholar] [CrossRef]
- Geng, X.; Shao, H.; Zhang, Z.; Ng, J.C.; Peng, C. Malathion-induced testicular toxicity is associated with spermatogenic apoptosis and alterations in testicular enzymes and hormone levels in male Wistar rats. Environ. Toxicol. Pharmacol. 2015, 39, 659–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, D.; Sangha, G.K.; Khera, K.S. Triazophos-induced oxidative stress and histomorphological changes in ovary of female Wistar rats. Pestic. Biochem. Physiol. 2015, 117, 9–18. [Google Scholar] [CrossRef]
- Jallouli, M.; Dhouib, I.E.B.; Lasram, M.; Gharbi, N.; El Fazaa, S. Disruption of steroidogenesis after dimethoate exposure and efficacy of N-acetylcysteine in rats: An old drug with new approaches. Environ. Sci. Pollut. 2016, 23, 7975–7984. [Google Scholar] [CrossRef]
- Leoni, C.; Balduzzi, M.; Buratti, F.M.; Testai, E. The contribution of human small intestine to chlorpyrifos biotransformation. Toxicol. Lett. 2012, 215, 42–48. [Google Scholar] [CrossRef]
- Harishankar, M.K.; Sasikala, C.; Ramya, M. Efficiency of the intestinal bacteria in the degradation of the toxic pesticide, chlorpyrifos. 3 Biotech 2013, 3, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-J.; Li, S.; Gan, R.-Y.; Zhou, T.; Xu, D.-P.; Li, H.-B.; Sugumaran, M. Impacts of Gut Bacteria on Human Health and Diseases. Int. J. Mol. Sci. 2015, 16, 7493–7519. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.A.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Fang, B.; Li, J.W.; Zhang, M.; Ren, F.Z.; Pang, G.F. Chronic chlorpyrifos exposure elicits diet-specific effects on metabolism and the gut microbiome in rats. Food Chem. Toxicol. 2018, 111, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Condette, C.J.; Khorsi-Cauet, H.; Morlière, P.; Zabijak, L.; Reygner, J.; Bach, V.; Gay-Quéheillard, J. Increased Gut Permeability and Bacterial Translocation after Chronic Chlorpyrifos Exposure in Rats. PLoS ONE 2014, 9, e102217. [Google Scholar] [CrossRef]
- Li, W.; Ehrich, M. Transient alterations of the blood-brain barrier tight junction and receptor potential channel gene expression by chlorpyrifos. J. Appl. Toxicol. 2013, 33, 1187–1191. [Google Scholar] [CrossRef]
- Vismaya; Rajini, P.S. Oral exposure to the organophosphorus insecticide, Monocrotophos induces intestinal dysfunction in rats. Food. Chem. Toxicol. 2014, 71, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Zeng, Z.; Fu, Z.; Jin, Y. Oral imazalil exposure induces gut microbiota dysbiosis and colonic inflammation in mice. Chemosphere 2016, 160, 349–358. [Google Scholar] [CrossRef]
- Jin, C.; Zeng, Z.; Fu, Z.; Jin, Y. Chronic exposure to endosulfan induces inflammation in murine colon via β-catenin expression and IL-6 production. Chemosphere 2016, 13, 842–849. [Google Scholar]
- Jin, Y.; Wu, S.; Zeng, Z.Y.; Fu, Z. Effects of environmental pollutants on gut microbiota. Environ. Pollut. 2017, 222, 1–9. [Google Scholar] [CrossRef]
- Jin, Y.; Zeng, Z.; Wu, Y.; Zhang, S.; Fu, Z. Oral Exposure of Mice to Carbendazim Induces Hepatic Lipid Metabolism Disorder and Gut Microbiota Dysbiosis. Toxicol. Sci. 2015, 147, 116–126. [Google Scholar] [CrossRef] [Green Version]
- Reygner, J.; Joly Condette, C.; Bruneau, A.; Delanaud, S.; Rhazi, L.; Depeint, F.; Abdennebi-Najar, L.; Bach, V.; Mayeur, C.; Khorsi-Cauet, H. Changes in Composition and Function of Human Intestinal Microbiota Exposed to Chlorpyrifos in Oil as Assessed by the SHIME® Model. Int. J. Environ. Res. Public Health 2016, 13, 1088. [Google Scholar] [CrossRef]
- Velmurugan, G.; Ramprasath, T.; Swaminathan, K.; Mithieux, G.; Rajendhran, J.; Dhivakar, M.; Parthasarathy, A.; Babu, D.D.V.; Thumburaj, L.J.; Freddy, A.J.; et al. Gut microbial degradation of organophosphate insecticides-induces glucose intolerance via gluconeogenesis. Genome Biol. 2017, 18, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Driver, J.H.; Li, Y.; Ross, J.H.; Krieger, R.I. Dialkylphosphates (DAPs) in Fruits and Vegetables May Confound Biomonitoring in Organophosphorus Insecticide Exposure and Risk Assessment. J. Agric. Food Chem. 2008, 56, 10638–10645. [Google Scholar] [CrossRef] [PubMed]
- Pesticide residues in food 2002 Joint FAO/WHO Meeting on Pesticide Residues TRIAZOPHOS. Available online: http://www.inchem.org/documents/jmpr/jmpmono/2002pr14.htm (accessed on 10 May 2019).
- Wang, H.-P.; Liang, Y.-J.; Long, D.-X.; Chen, J.-X.; Hou, W.-Y.; Wu, Y.-J. Metabolic Profiles of Serum from Rats after Subchronic Exposure to Chlorpyrifos and Carbaryl. Chem. Toxicol. 2009, 22, 1026–1033. [Google Scholar] [CrossRef]
- Fu, Y.; Xie, W. Analysis of exposure levels of several common environmental pollutants in medical examination population. J. Qiqihar Uni. Med. 2013, 34, 2897–2898. (In Chinese) [Google Scholar]
- National Health Commission of the People’s Republic of China. The Report on Nutrition and Chronic Diseases of Chinese Residents. Available online: http://www.nhc.gov.cn/jkj/s5879/201506/4505528e65f3460fb88685081ff158a2.shtml (accessed on 15 April 2019).
- Urine 24-Hour Volume. Available online: https://medlineplus.gov/ency/article/003425.htm (accessed on 29 March 2019).
- Omoike, O.E.; Lewis, R.C.; Meeker, J.D. Association between urinary biomarkers of exposure to organophosphate insecticides and serum reproductive hormones in men from NHANES 1999-2002. Reprod Toxicol 2015, 53, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Meeker, J.D.; Ravi, S.R.; Barr, D.B.; Hauser, R. Circulating estradiol in men is inversely related to urinary metabolites of nonpersistent insecticides. Reprod. Toxicol. 2008, 25, 184–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korljan, B.; Bagatin, J.; Kokić, S.; Matulić, N.B.; Ostojić, S.B.; Deković, A. The impact of hormone replacement therapy on metabolic syndrome components in perimenopausal women. Med. Hypotheses 2010, 74, 162–163. [Google Scholar] [CrossRef] [PubMed]
- Whitcroft, S.I.; Crook, D.; Marsh, M.S.; Ellerington, M.C.; I Whitehead, M.; Stevenson, J.C. Long-term effects of oral and transdermal hormone replacement therapies on serum lipid and lipoprotein concentrations. Obstet. Gynecol. 1994, 84, 222–226. [Google Scholar]
- Luo, F.; Huang, W.-Y.; Guo, Y.; Ruan, G.-Y.; Peng, R.; Li, X.-P. 17β-estradiol lowers triglycerides in adipocytes via estrogen receptor α and it may be attenuated by inflammation. Lipids Heal. 2017, 16, 182. [Google Scholar] [CrossRef]
- Filippatos, T.; Liberopoulos, E.; Pavlidis, N.; Elisaf, M.; Mikhailidis, D.; Filippatos, T.; Liberopoulos, E.; Elisaf, M. Effects of hormonal treatment on lipids in patients with cancer. N.a. Rev. 2009, 35, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Lecomte, M.J.; Bertolus, C.; Ramanantsoa, N.; Saurini, F.; Callebert, J.; Sénamaud-Beaufort, C.; Ringot, M.; Bourgeois, T.; Matrot, B.; Collet, C.; et al. Acetylcholine modulates the hormones of the growth hormone/insulin like growth factor-1 axis during development in mice. Endocrinology 2018, 159, 1844–1859. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, K.; Sumi, G.; Kanamori, C.; Nakajima, T.; Tsuzuki, T.; Cho, H.; Nishigaki, A.; Okada, H.; Kanzaki, H. Effects of ovarian hormone treatment on the gene expression of muscarinic acetylcholine receptors in the ovariectomized rat myometrium. J. Steroid Biochem. Mol. Boil. 2014, 143, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Wang, Q.; Liu, Y.-P.; Xu, L.-F.; Li, Y.-F.; Hu, J.-X.; Jiang, M.; Zhang, J.-P.; Zhang, M.-R.; Yu, H.-M.; et al. Anti-androgen effects of the pyrethroid pesticide cypermethrin on interactions of androgen receptor with corepressors. Toxicology 2013, 311, 178–183. [Google Scholar] [CrossRef]
- Xu, Z.; Wang, C.; Dong, X.; Hu, T.; Wang, L.; Zhao, W.; Zhu, S.; Li, G.; Hu, Y.; Gao, Q.; et al. Chronic alcohol exposure induced gut microbiota dysbiosis and its correlations with neuropsychic behaviors and brain BDNF/Gabra1 changes in mice. BioFactors 2018, 45, 187–199. [Google Scholar] [CrossRef]
- Shaw, K.A.; Bertha, M.; Hofmekler, T.; Chopra, P.; Vatanen, T.; Srivatsa, A.; Prince, J.; Kumar, A.; Sauer, C.; Zwick, M.E.; et al. Dysbiosis, inflammation, and response to treatment: A longitudinal study of pediatric subjects with newly diagnosed inflammatory bowel disease. Genome Med. 2016, 8, 75. [Google Scholar] [CrossRef]
- Bajer, L.; Kverka, M.; Kostovcik, M.; Macinga, P.; Dvorak, J.; Stehlikova, Z.; Brezina, J.; Wohl, P.; Spicak, J.; Drastich, P. Distinct gut microbiota profiles in patients with primary sclerosing cholangitis and ulcerative colitis. World J. Gastroenterol. 2017, 23, 4548–4558. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chia, N.; Kalari, K.R.; Yao, J.Z.; Novotna, M.; Soldan, M.M.P.; Luckey, D.H.; Marietta, E.V.; Jeraldo, P.R.; Chen, X.; et al. Multiple sclerosis patients have a distinct gut microbiota compared to healthy controls. Sci. Rep. 2016, 6, 28484. [Google Scholar] [CrossRef] [Green Version]
- Peterson, L.W.; Artis, D. Intestinal epithelial cells: Regulators of barrier function and immune homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef]
- Alonso, C.; Vicario, M.; Pigrau, M.; Lobo, B.; Santos, J.; Pastor, M.P. Intestinal Barrier Function and the Brain-Gut Axis. Purine and Pyrimidine Metabolism in Man V 2014, 817, 73–113. [Google Scholar]
- Heni, M.; Kullmann, S.; Gallwitz, B.; Haring, H.-U.; Preissl, H.; Fritsche, A. Dissociation of GLP-1 and insulin association with food processing in the brain: GLP-1 sensitivity despite insulin resistance in obese humans. Mol. Metab. 2015, 4, 971–976. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Jun, H.-S. Anti-diabetic actions of glucagon-like peptide-1 on pancreatic beta-cells. Metab. Clin. Exp. 2014, 63, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.M.; Du, X.X.; Jiang, H.; Xie, J.X. Ghrelin and Neurodegenerative Disorders-a Review. Mol. Neurobiol. 2017, 54, 1144–1155. [Google Scholar] [CrossRef]
- Stadlbauer, U.; Woods, S.C.; Langhans, W.; Meyer, U. PYY3-36: Beyond food intake. Front. Neuroendocrinol. 2015, 38, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Al-Asmakh, M.; Stukenborg, J.-B.; Reda, A.; Anuar, F.; Strand, M.-L.; Hedin, L.; Pettersson, S.; Söder, O. The Gut Microbiota and Developmental Programming of the Testis in Mice. PLoS ONE 2014, 9, 103809. [Google Scholar] [CrossRef]
- Gérard, P. Gastrointestinal Tract: Microbial Metabolism of Steroids. In Handbook of Hydrocarbon and Lipid Microbiology; Springer Nature: Basingstoke, UK, 2010; pp. 3133–3140. [Google Scholar]
- Lu, N.; Li, M.; Lei, H.; Jiang, X.; Tu, W.; Lu, Y.; Xia, D. Butyric acid regulates progesterone and estradiol secretion via cAMP signaling pathway in porcine granulosa cells. J. Steroid Biochem. Mol. Boil. 2017, 172, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Yue, S.; Yang, Z.; Feng, W.; Meng, X.; Wang, A.; Peng, C.; Wang, C.; Yan, D.; Yan, Z. Oral hydroxysafflor yellow A reduces obesity in mice by modulating the gut microbiota and serum metabolism. Pharmacol. Res. 2018, 134, 40–50. [Google Scholar] [CrossRef]
- Shetty, S.A.; Lagkouvardos, I.; Ritari, J.; Chamlagain, B.; Douillard, F.P.; Paulin, L.; Piironen, V.; Clavel, T.; Plugge, C.M.; De Vos, W.M.; et al. Comparative genomics and physiology of the butyrate-producing bacterium Intestinimonas butyriciproducens. Environ. Microbiol. Rep. 2016, 8, 1024–1037. [Google Scholar]
- Palm, N.W.; De Zoete, M.R.; Flavell, R.A. Immune–microbiota interactions in health and disease. Clin. Immunol. 2015, 159, 122–127. [Google Scholar] [CrossRef]
- Lyte, M. Microbial Endocrinology in the Microbiome-Gut-Brain Axis: How Bacterial Production and Utilization of Neurochemicals Influence Behavior. PLOS Pathog. 2013, 9, e1003726. [Google Scholar] [CrossRef] [PubMed]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. 2015, 39, 424–429. [Google Scholar] [CrossRef]
- Chambers, E.S.; Morrison, D.J.; Frost, G. Control of appetite and energy intake by SCFA: What are the potential underlying mechanisms? Proc. Nutr. Soc. 2015, 74, 328–336. [Google Scholar] [CrossRef]
- Marathe, N.P.; Ranade, D.R.; Shouche, Y.S.; Lanjekar, V.B. Clostridium punense sp. nov. an obligate anaerobic bacteria isolated from healthy human faeces. Int. J. Syst. Evol. Microbiol. 2015, 65, 4749–4756. [Google Scholar]
- Lesniewska, V.; Rowland, I.; Cani, P.D.; Neyrinck, A.M.; Delzenne, N.M.; Naughton, P.J. Effect on Components of the Intestinal Microflora and Plasma Neuropeptide Levels of Feeding Lactobacillus delbrueckii, Bifidobacterium lactis, and Inulin to Adult and Elderly Rats. Appl. Environ. Microbiol. 2006, 72, 6533–6538. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Masuda, K.; Kishimoto, T. Regulation of IL-6 in Immunity and Diseases. Purine Pyrimidine Metab. Man V 2016, 941, 79–88. [Google Scholar]
- Ye, Z.; Zhang, N.; Wu, C.; Zhang, X.; Wang, Q.; Huang, X.; Du, L.; Cao, Q.; Tang, J.; Zhou, C.; et al. A metagenomic study of the gut microbiome in Behcet’s disease. Microbiome 2018, 6, 135. [Google Scholar] [CrossRef] [PubMed]
- Lv, L.-X.; Fang, D.-Q.; Shi, D.; Chen, D.-Y.; Yan, R.; Zhu, Y.-X.; Chen, Y.-F.; Shao, L.; Guo, F.-F.; Wu, W.-R.; et al. Alterations and correlations of the gut microbiome, metabolism and immunity in patients with primary biliary cirrhosis. Environ. Microbiol. 2016, 18, 2272–2286. [Google Scholar] [CrossRef]
- Kang, Y.; Li, Y.; Du, Y.; Guo, L.; Chen, M.; Huang, X.; Yang, F.; Hong, J.; Kong, X. Konjaku flour reduces obesity in mice by modulating the composition of the gut microbiota. Int. J. Obes. 2018, 1. [Google Scholar] [CrossRef]
- Hu, Y.-L.; Pang, W.; Huang, Y.; Zhang, Y.; Zhang, C.-J. The Gastric Microbiome Is Perturbed in Advanced Gastric Adenocarcinoma Identified Through Shotgun Metagenomics. Front. Microbiol. 2018, 8, 433. [Google Scholar] [CrossRef] [Green Version]
- Blosse, A.; Lehours, P.; Wilson, K.T.; Gobert, A.P. Helicobacter: Inflammation, immunology, and vaccines. Helicobacter 2018, 23, e12517. [Google Scholar] [CrossRef]
- Wang, F.; Meng, W.; Wang, B.; Qiao, L. Helicobacter pylori-induced gastric inflammation and gastric cancer. Cancer Lett. 2014, 345, 196–202. [Google Scholar] [CrossRef]
- Barcellini, W.; Rizzardi, G.; Borghi, M.; Nicoletti, F.; Fain, C.; Del Papa, N.; Meroni, P. In vitro type-1 and type-2 cytokine production in systemic lupus erythematosus: Lack of relationship with clinical disease activity. Lupus 1996, 5, 139–145. [Google Scholar] [CrossRef]
- Hirano, D.; Nagashima, M.; Ogawa, R.; Yoshino, S. Serum levels of interleukin 6 and stress related substances indicate mental stress condition in patients with rheumatoid arthritis. J. Rheumatol. 2001, 28, 490–495. [Google Scholar]
- Oh, Y.S.; Bae, G.D.; Park, E.-Y.; Jun, H.-S. MicroRNA-181c Inhibits Interleukin-6-mediated Beta Cell Apoptosis by Targeting TNF-α Expression. Molecules 2019, 24, 1410. [Google Scholar] [CrossRef] [PubMed]
- Knüpfer, H.; Preiss, R. Serum interleukin-6 levels in colorectal cancer patients—A summary of published results. Int. J. Colorectal Dis. 2010, 25, 135–140. [Google Scholar] [CrossRef]
- Köchl, S.; Niederstätter, H.; Parson, W. DNA extraction and quantitation of forensic samples using the phenol-chloroform method and real-time PCR. Methods Mol. Boil. (Clifton, N.J.) 2005, 297, 18. [Google Scholar]
- Calderón, K.; Spor, A.; Breuil, M.C.; Bru, D.; Bizouard, F.; Violle, C.; Barnard, R.L.; Philippot, L. Effectiveness of ecological rescue for altered soil microbial communities and functions. ISME J. 2017, 11, 272. [Google Scholar] [CrossRef] [PubMed]
- White, J.R.; Nagarajan, N.; Pop, M. Statistical Methods for Detecting Differentially Abundant Features in Clinical Metagenomic Samples. PLOS Comput. Boil. 2009, 5, e1000352. [Google Scholar] [CrossRef]
- Zheng, J.; Xiao, X.-H.; Zhang, Q.; Mao, L.-L.; Yu, M.; Xu, J.-P.; Wang, T. Correlation of placental microbiota with fetal macrosomia and clinical characteristics in mothers and newborns. Oncotarget 2017, 8, 82314–82325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Sample Availability: Samples of the compounds are available from the authors. |





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, F.; Li, J.; Pang, G.; Ren, F.; Fang, B. Effects of Diethyl Phosphate, a Non-Specific Metabolite of Organophosphorus Pesticides, on Serum Lipid, Hormones, Inflammation, and Gut Microbiota. Molecules 2019, 24, 2003. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24102003
Yang F, Li J, Pang G, Ren F, Fang B. Effects of Diethyl Phosphate, a Non-Specific Metabolite of Organophosphorus Pesticides, on Serum Lipid, Hormones, Inflammation, and Gut Microbiota. Molecules. 2019; 24(10):2003. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24102003
Chicago/Turabian StyleYang, Fangwei, Jinwang Li, Guofang Pang, Fazheng Ren, and Bing Fang. 2019. "Effects of Diethyl Phosphate, a Non-Specific Metabolite of Organophosphorus Pesticides, on Serum Lipid, Hormones, Inflammation, and Gut Microbiota" Molecules 24, no. 10: 2003. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24102003