Dihydroartemisinin Regulates the Th/Treg Balance by Inducing Activated CD4+ T cell Apoptosis via Heme Oxygenase-1 Induction in Mouse Models of Inflammatory Bowel Disease

Abstract
:1. Introduction
2. Results
2.1. DHA Ameliorates OXA-Induced Colitis in a Dose-Dependent Manner in Vivo
2.2. DHA Ameliorates TNBS-Induced Colitis in a Dose-Dependent Manner in Vivo
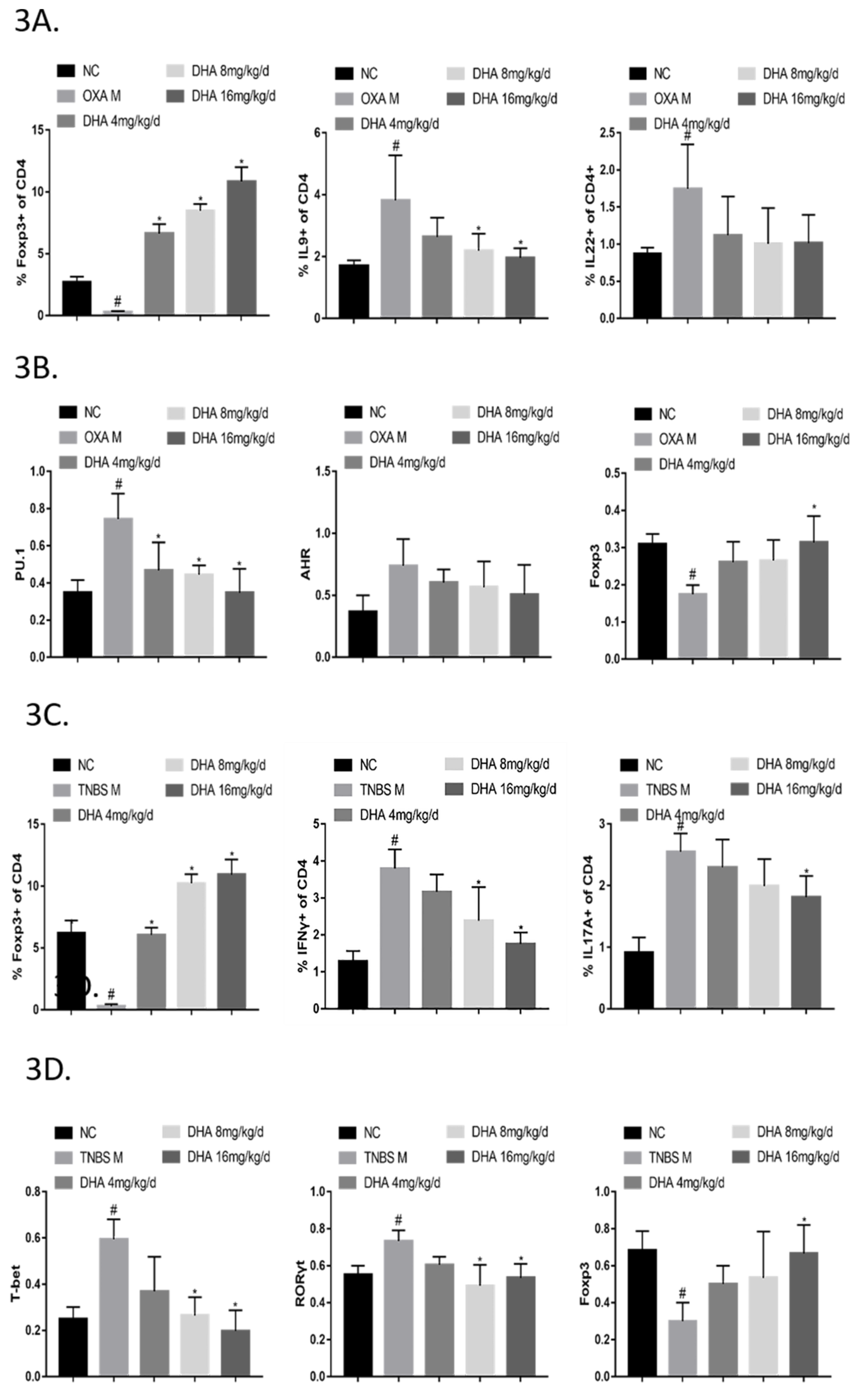
2.3. DHA Regulates the Th/Treg Balance in OXA- and TNBS-Induced Colitis
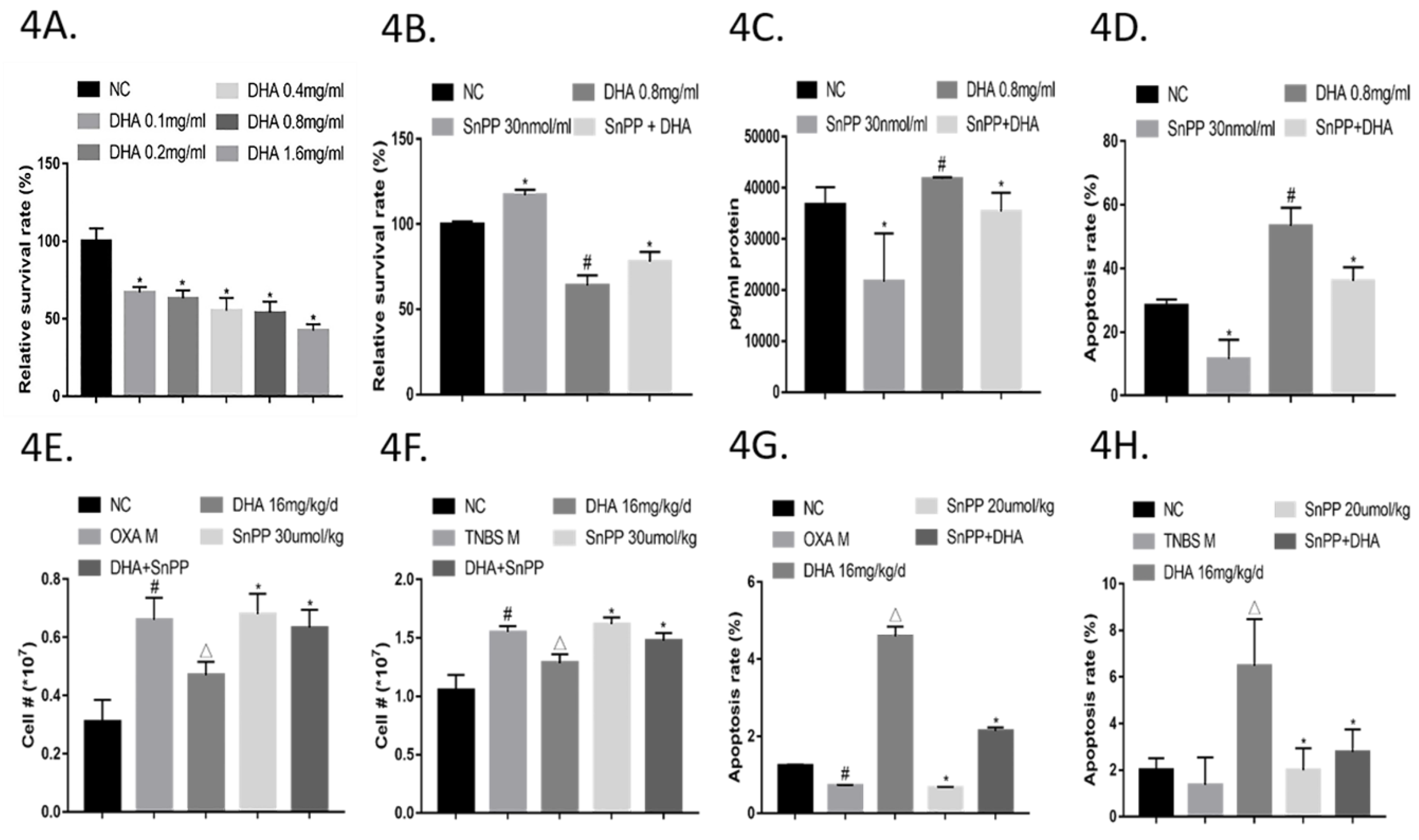
2.4. DHA Suppresses Activated CD4+ T Cell Subsets by Inducing Apoptosis via HO-1
2.5. DHA Ameliorates OXA- and TNBS-Induced Colitis via HO-1
2.6. DHA Regulates the Th/Treg Balance in OXA- and TNBS-Induced Colitis via HO-1
3. Discussion
4. Materials and Methods
4.1. Colitis Induction and Disease Assessment
4.2. In Vivo Administration of DHA or SnPP
4.3. Histopathology
4.4. Preparation of Spleen Lymphocytes and LPMCs
4.5. Flow Cytometry
4.6. Quantitative RT-PCR
4.7. T Cell Apoptosis Assays
4.8. ELISA and Immunoblotting
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rieder, F.; Fiocchi, C. Mechanisms, Management, and Treatment of Fibrosis in Patients with Inflammatory Bowel Diseases. Gastroenterology 2017, 152, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.G. The global burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Ananthakrishnan, A.N. Epidemiology and risk factors for IBD. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Kirchgesner, J.; Beaugerie, L. Increased risk of acute arterial events in young patients and severely active IBD: A nationwide French cohort study. Gut 2018, 67, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Choi, C.R.; Bakir, I.A. Clonal evolution of colorectal cancer in IBD. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Mowat, C.; Cole, A. Guidelines for the management of inflammatory bowel disease in adults. Gut 2011, 60, 571–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, B.; Dailey, F. Patient Understanding of the Risks and Benefits of Biologic Therapies in Inflammatory Bowel Disease: Insights from a Large-scale Analysis of Social Media Platforms. Inflamm. Bowel Dis. 2017, 23, 1057–1064. [Google Scholar] [CrossRef]
- Tu, Y. The discovery of artemisinin (qinghaosu) and gifts from Chinese medicine. Nat. Med. 2011, 17, 1217–1220. [Google Scholar] [CrossRef]
- Tu, Y. Artemisinin-A Gift from Traditional Chinese Medicine to the World (Nobel Lecture). Angew. Chem. Int. Ed. Engl. 2016, 55, 10210–10226. [Google Scholar] [CrossRef]
- Zhao, Y.G.; Wang, Y. Dihydroartemisinin ameliorates inflammatory disease by its reciprocal effects on Th and regulatory T cell function via modulating the mammalian target of rapamycin pathway. J. Immunol. 2012, 189, 4417–4425. [Google Scholar] [CrossRef]
- Fan, M.; Li, Y. DC32, a Dihydroartemisinin Derivative, Ameliorates Collagen-Induced Arthritis through an Nrf2-p62-Keap1 Feedback Loop. Front. Immunol. 2018, 9, 2762. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ueno, A. Crossover Subsets of CD4+ T Lymphocytes in the Intestinal Lamina Propria of Patients with Crohn’s Disease and Ulcerative Colitis. Dig. Dis. Sci. 2017, 62, 2357–2368. [Google Scholar] [CrossRef] [PubMed]
- Omenetti, S.; Pizarro, T.T. The Treg/Th17 Axis: A Dynamic Balance Regulated by the Gut Microbiome. Front. Immunol. 2015, 6, 639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegazy, A.N.; West, N.R. Circulating and Tissue-Resident CD4+ T Cells with Reactivity to Intestinal Microbiota Are Abundant in Healthy Individuals and Function Is Altered During Inflammation. Gastroenterology 2017, 153, 1320–1337. [Google Scholar] [CrossRef] [PubMed]
- Gui, X.; Li, J. Histopathological Features of Inflammatory Bowel Disease are Associated with Different CD4+ T Cell Subsets in Colonic Mucosal Lamina Propria. J. Crohns Colitis 2018, 12, 1448–1458. [Google Scholar] [CrossRef]
- Zhou, R.; Chang, Y. JNK Pathway-Associated Phosphatase/DUSP22 Suppresses CD4+ T-Cell Activation and Th1/Th17-Cell Differentiation and Negatively Correlates with Clinical Activity in Inflammatory Bowel Disease. Front. Immunol. 2017, 8, 781. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, K.; Hwang, Y. TH9 cells that express the transcription factor PU.1 drive T cell-mediated colitis via IL-9 receptor signaling in intestinal epithelial cells. Nat. Immunol. 2014, 15, 676–686. [Google Scholar] [CrossRef]
- Tang, M.S.; Bowcutt, R. Integrated Analysis of Biopsies from Inflammatory Bowel Disease Patients Identifies SAA1 as a Link Between Mucosal Microbes with TH17 and TH22 Cells. Inflamm. Bowel Dis. 2017, 23, 1544–1554. [Google Scholar] [CrossRef] [Green Version]
- Poulos, T.L. Heme enzyme structure and function. Chem. Rev. 2014, 114, 3919–3962. [Google Scholar] [CrossRef]
- Schipper, H.M.; Song, W. The sinister face of heme oxygenase-1 in brain aging and disease. Prog. Neurobiol. 2019, 172, 40–70. [Google Scholar] [CrossRef]
- Meunier, B.; Robert, A. Heme as trigger and target for trioxane-containing antimalarial drugs. Acc. Chem. Res. 2010, 43, 1444–1451. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, C.J. Haem-activated promiscuous targeting of artemisinin in Plasmodium falciparum. Nat. Commun. 2015, 6, 10111. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lai, W. BLT1 in dendritic cells promotes Th1/Th17 differentiation and its deficiency ameliorates TNBS-induced colitis. Cell. Mol. Immunol. 2018, 15, 1047–1056. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, S.; Popp, V. Chemically induced mouse models of acute and chronic intestinal inflammation. Nat. Protoc. 2017, 12, 1295–1309. [Google Scholar] [CrossRef]
- Yan, Y.X.; Shao, M.J. Artemisinin analogue SM934 ameliorates DSS-induced mouse ulcerative colitis via suppressing neutrophils and macrophages. Acta Pharmacol. Sin. 2018, 39, 1633–1644. [Google Scholar] [CrossRef]
- Ge, X.; Chen, Z. The effects of dihydroartemisinin on inflammatory bowel disease-related bone loss in a rat model. Exp. Biol. Med. (Maywood) 2018, 243, 715–724. [Google Scholar] [CrossRef]
- Yang, Z.; Ding, J. Immunomodulatory and anti-inflammatory properties of artesunate in experimental colitis. Curr. Med. Chem. 2012, 19, 4541–4551. [Google Scholar] [CrossRef]
- Rohr, J.C.; Gerlach, C. Single cell behavior in T cell differentiation. Trends Immunol. 2014, 35, 170–177. [Google Scholar] [CrossRef]
- Cravens, P.D.; Hussain, R.Z. Lymph node-derived donor encephalitogenic CD4+ T cells in C57BL/6 mice adoptive transfer experimental autoimmune encephalomyelitis highly express GM-CSF and T-bet. J. Neuroinflamm. 2011, 8, 73. [Google Scholar] [CrossRef]
- Chyuan, I.T.; Tsai, H.F. TRAIL-Mediated Suppression of T Cell Receptor Signaling Inhibits T Cell Activation and Inflammation in Experimental Autoimmune Encephalomyelitis. Front. Immunol. 2018, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.F.; He, S.J. SM934, a water-soluble derivative of arteminisin, exerts immunosuppressive functions in vitro and in vivo. Int. Immunopharmacol. 2009, 9, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Simon, T.; Pogu, J. Inhibition of effector antigen-specific T cells by intradermal administration of heme oxygenase-1 inducers. J. Autoimmun. 2017, 81, 44–55. [Google Scholar] [CrossRef] [PubMed]
- McDaid, J.; Yamashita, K. Heme oxygenase-1 modulates the allo-immune response by promoting activation-induced cell death of T cells. FASEB J. 2005, 19, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.; Zhang, Q. 5-Aminolevulinic acid with ferrous iron induces permanent cardiac allograft acceptance in mice via induction of regulatory cells. J. Heart Lung Transplant. 2015, 34, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Chora, A.A.; Fontoura, P. Heme oxygenase-1 and carbon monoxide suppress autoimmune neuroinflammation. J. Clin. Investig. 2007, 117, 438–447. [Google Scholar] [CrossRef]
- Mangano, K.; Cavalli, E. Involvement of the Nrf2/HO-1/CO axis and therapeutic intervention with the CO-releasing molecule CORM-A1, in a murine model of autoimmune hepatitis. J. Cell. Physiol. 2018, 233, 4156–4165. [Google Scholar] [CrossRef]
- Fagone, P.; Mangano, K. Carbon monoxide-releasing molecule-A1 (CORM-A1) improves clinical signs of experimental autoimmune uveoretinitis (EAU) in rats. Clin. Immunol. 2015, 157, 198–204. [Google Scholar] [CrossRef]
- Fagone, P.; Mangano, K. Prevention of clinical and histological signs of proteolipid protein (PLP)-induced experimental allergic encephalomyelitis (EAE) in mice by the water-soluble carbon monoxide-releasing molecule (CORM)-A1. Clin. Exp. Immunol. 2011, 163, 368–374. [Google Scholar] [CrossRef]
- Nikolic, I.; Saksida, T. Pharmacological application of carbon monoxide ameliorates islet-directed autoimmunity in mice via anti-inflammatory and anti-apoptotic effects. Diabetologia 2014, 57, 980–990. [Google Scholar] [CrossRef]
- Sebastián, V.P.; Salazar, G.A. Heme Oxygenase-1 as a Modulator of Intestinal Inflammation Development and Progression. Front. Immunol. 2018, 9, 1956. [Google Scholar] [CrossRef]
- Günther, S.; Fagone, P. Role of MIF and D-DT in immune-inflammatory, autoimmune, and chronic respiratory diseases: From pathogenic factors to therapeutic targets. Drug Discov. Today 2019, 24, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Mangano, K.; Mazzon, E. Pathogenic role for macrophage migration inhibitory factor in glioblastoma and its targeting with specific inhibitors as novel tailored therapeutic approach. Oncotarget 2018, 9, 17951–17970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, V.; Kindt, N. Macrophage migration inhibitory factor involvement in breast cancer (Review). Int. J. Oncol. 2015, 47, 1627–1633. [Google Scholar] [CrossRef] [PubMed]
- Lechien, J.R.; Nassri, A. Role of macrophage migration inhibitory factor in head and neck cancer and novel therapeutic targets: A systematic review. Head Neck 2017, 39, 2573–2584. [Google Scholar] [CrossRef] [PubMed]
- Morris, K.T.; Nofchissey, R.A. Chronic macrophage migration inhibitory factor exposure induces mesenchymal epithelial transition and promotes gastric and colon cancers. PLoS ONE 2014, 9, e98656. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Chen, W. Artesunate inhibits type I interferon-induced production of macrophage migration inhibitory factor in patients with systemic lupus erythematosus. Lupus 2017, 26, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Santucci, L.; Fiorucci, S. Galectin-1 suppresses experimental colitis in mice. Gastroenterology 2003, 124, 1381–1394. [Google Scholar] [CrossRef]
- Okazawa, A.; Kanai, T. Th1-mediated intestinal inflammation in Crohn’s disease may be induced by activation of lamina propria lymphocytes through synergistic stimulation of interleukin-12 and interleukin-18 without T cell receptor engagement. Am. J. Gastroenterol. 2002, 97, 3108–3117. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds Dihydroartemisinin are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Weight Loss | Stool Consistency | Blood |
|---|---|---|---|
| 0 | None | Normal | Negative hemoculture |
| 1 | 1–5% | Soft but still formed | Negative hemoculture |
| 2 | 6–10% | Soft | Positive hemoculture |
| 3 | 11–18% | Very soft; wet | Blood traces in stool visible |
| 4 | >18% | Watery diarrhea | Gross rectal bleeding |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, S.C.; Wang, Y.J.; Li, Y.J.; Cai, W.Y.; Weng, X.G.; Li, Q.; Chen, Y.; Yang, Q.; Zhu, X.X. Dihydroartemisinin Regulates the Th/Treg Balance by Inducing Activated CD4+ T cell Apoptosis via Heme Oxygenase-1 Induction in Mouse Models of Inflammatory Bowel Disease. Molecules 2019, 24, 2475. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24132475
Yan SC, Wang YJ, Li YJ, Cai WY, Weng XG, Li Q, Chen Y, Yang Q, Zhu XX. Dihydroartemisinin Regulates the Th/Treg Balance by Inducing Activated CD4+ T cell Apoptosis via Heme Oxygenase-1 Induction in Mouse Models of Inflammatory Bowel Disease. Molecules. 2019; 24(13):2475. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24132475
Chicago/Turabian StyleYan, Si Chao, Ya Jie Wang, Yu Jie Li, Wei Yan Cai, Xiao Gang Weng, Qi Li, Ying Chen, Qing Yang, and Xiao Xin Zhu. 2019. "Dihydroartemisinin Regulates the Th/Treg Balance by Inducing Activated CD4+ T cell Apoptosis via Heme Oxygenase-1 Induction in Mouse Models of Inflammatory Bowel Disease" Molecules 24, no. 13: 2475. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24132475