
Sampling for DUS Test of Flower Colors of Ranunculus asiaticus L. in View of Spatial and Temporal Changes of Flower Colorations, Anthocyanin Contents, and Gene Expression Levels

Abstract
:1. Introduction
2. Results
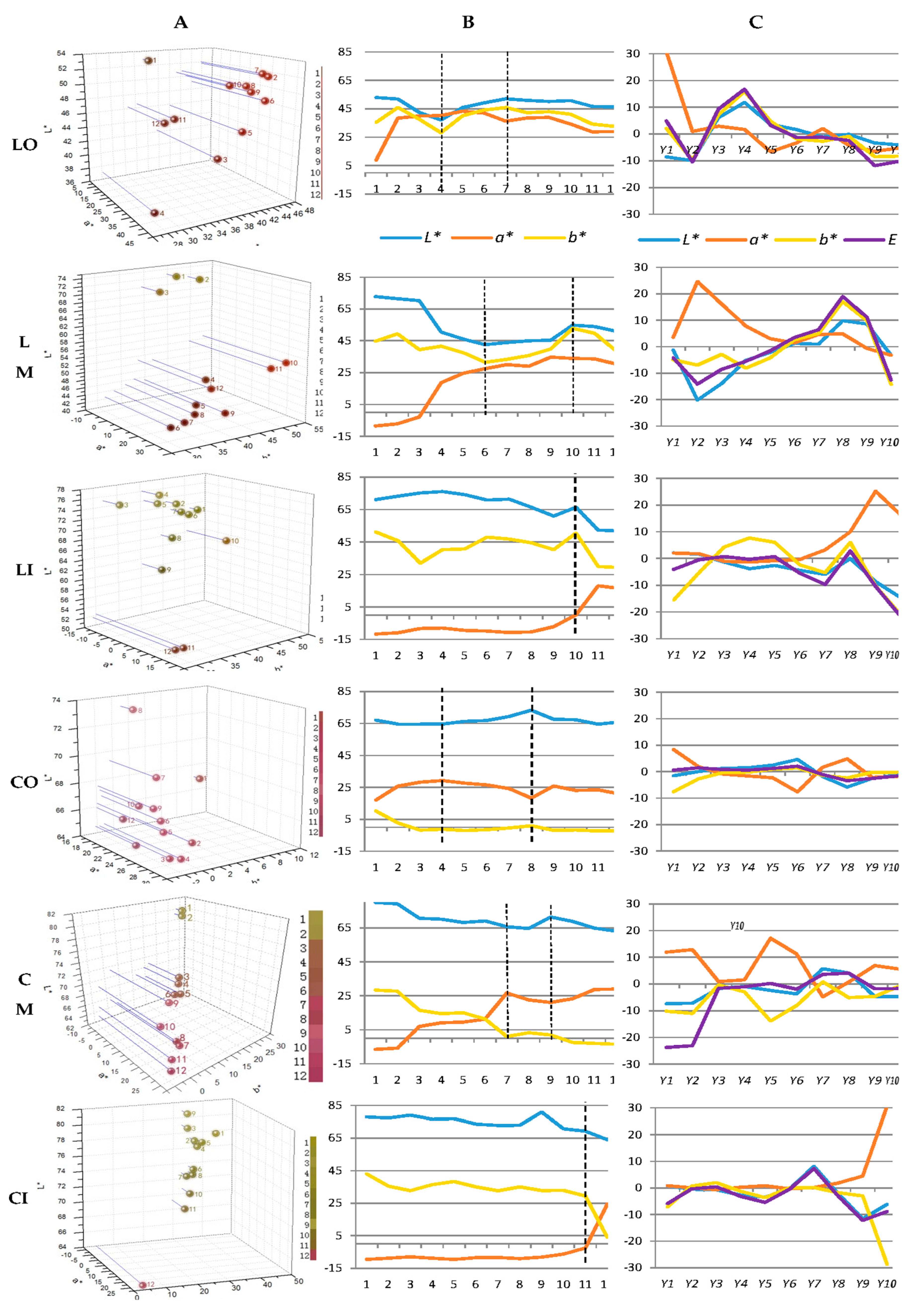
2.1. Spatial and Temporal Change Traits of Flower Coloration
2.1.1. Expression States of Flower Coloration
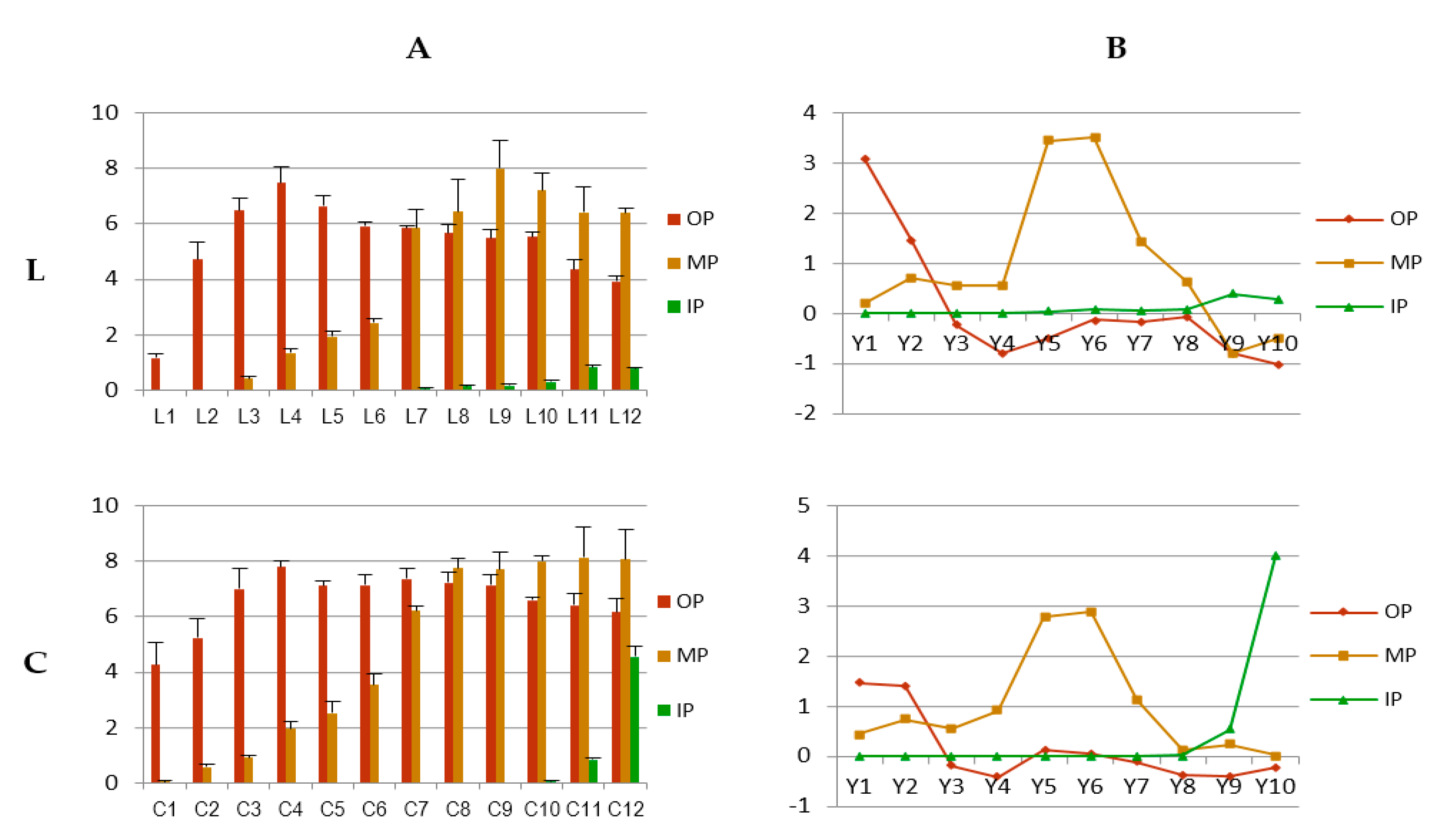
2.1.2. Uniformity of Flower Coloration
2.1.3. Stability of Flower Coloration
2.2. Metabolic Evidence of Anthocyanin Content for the Change Traits of Flower Colorations
2.3. Molecular Evidence of Gene Expression Patterns for the Change Traits of Flower Colorations
2.3.1. Selecting of Reference Genes for Quantitative Real-Time PCR
2.3.2. Gene Screening for Expression Pattern Analysis
2.3.3. Change Traits of Expression Levels of c72570, c130622, and c83020
2.4. Correlation among Color Phenotypes, Anthocyanin Contents, and Gene Expression Patterns
3. Discussion
4. Materials and Methods
4.1. Spatial and Temporal Change Traits of Flower Coloration
4.2. Measurement of Color
4.3. Measurement of Anthocyanin Content and Tentative Identification of Anthocyanin Components
4.4. Expression Patterns of Related Gene
4.5. Change Traits and Model of Change Dynamic of Flower Coloration
Author Contributions
Funding
Conflicts of Interest
References
- Azadi, P.; Bagheri, H.; Nalousi, A.M.; Nazari, F.; Chandler, S.F. Current status and biotechnological advances in genetic engineering of ornamental plants. Biotechnol. Adv. 2016, 34, 1073–1090. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aal, E.M.; Hucl, P.; Rabalski, I. Compositional and antioxidant propterties of anthocyanin-rich products prepared from purple wheat. Food Chem. 2018, 254, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Zha, J.; Koffas, M.A.G. Production of anthocyanins in metabolically engineered microorganisms: Current status and perspectives. Synth. Syst. Biotechnol. 2017, 2, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Ali, H.M.; Almagribi, W.; Al-Rashidi, M.N. Antiradical and reductant activities of anthocyanidins and anthocyanins structure-activity relationship and synthesis. Food Chem. 2016, 194, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Zhang, J.H.; Yang, X.H.; Wang, J.M.; Li, Y.G.; Zhang, P.; Mao, J.; Huang, Q.M.; Tang, H. Diversity in flower colorations of Ranunculus asiaticus L. revealed by anthocyanin biosynthesis pathway in view of gene composition, gene expression patterns and color phenotype. Environ. Sci. Pollut. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Dhooghe, E.; Grunewald, W.; Reheul, D.; Goetghebeur, P.; Van Labeke, M.C. Floral characteristics and gametophyte development of Anemone coronaria L. and Ranunculus asiaticus L. (Ranunculaceae). Sci. Hortic. 2012, 138, 73–80. [Google Scholar] [CrossRef]
- Zhang, J.H.; Liu, Y.F.; Yang, X.H.; Huang, Q.M.; Wang, J.M.; Zhang, P.; Zhang, H.; Hou, X.W.; Guan, J.J. Guidelines for the Conduct of Tests for Distinctness, Uniformity and Stability–Persian Buttercup (Ranunculus asiaticus L.); Chinese Agricultural Press: Beijing, China, 2014; pp. 4–16. [Google Scholar]
- Liu, Y.F.; Wang, J.M.; Zhang, J.H.; Li, Y.G.; Yang, X.H.; Zhang, P.; Huang, Q.M.; Tang, H. Characterization of SSRs and SNPs based on transcriptome of Ranunculus asiaticus L. and development of anthocyanin-related SSRs and SNPs. Nanosci. Nanotechnol. Lett. 2018, 10, 267–273. [Google Scholar] [CrossRef]
- Liu, Y.F.; Zhang, J.H.; Lü, B.; Yang, X.H.; Li, Y.G.; Wang, Y.; Wang, J.M.; Zhang, H.; Guan, J.J. Statistic analysis on quantitative characteristics for developing the DUS test guideline of Ranunculus asiaticus L. J. Integr. Agric. 2013, 12, 971–978. [Google Scholar] [CrossRef]
- Warren, D. Image analysis in chrysanthemum DUS testing. Comput. Electron. Agric. 2016, 18, 240–245. [Google Scholar] [CrossRef]
- Lee, H.H.; Hong, K.S. Automatic recognition of flower species in the natural environment. Image Vis. Comput. 2017, 61, 98–114. [Google Scholar] [CrossRef]
- Krishnaveni, S.; Pethalakshmi, A. Toward automatic quality detection of Jasmenum flower. Inf. Commun. Tech. Express 2017, 3, 148–153. [Google Scholar] [CrossRef]
- Li, X.L.; Qian, X.K.; Lü, X.; Wang, X.H.; Ji, N.; Zhang, M.S.; Ren, M.J. Upregulated structural and regulatory genes involved in anthocyanin biosynthesis for coloration of purple grains during the middle and late grain-filling stages. Plant Physiol. Biochem. 2018, 130, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Gu, C.; Liao, L.; Zhou, H.; Wang, L.; Deng, X.B.; Han, Y.P. Constitutive activation of an anthocyanin regulatory gene PcMYB10.6 is related to red coloration in purple-foliage plum. PLoS ONE 2015, 10, e0135159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, W.N.; Wang, Y.J.; Gao, S.L.; Du, D.N.; Fu, J.X.; Dong, L. Anthocyanin biosynthesis and accumulation in developing flowers of tree peony (Paeonia suffruticosa) ‘Luoyang Hong’. Postharvest Biol. Technol. 2014, 97, 11–22. [Google Scholar] [CrossRef]
- Zhang, W.X.; Jiang, Z.H.; Qiu, J.; Wei, H.L.; Cao, F.L. Studies on sequence dynamic distribution pattern of flower color parameters of Ornamental Crabapple. Acta Horic. Sin. 2013, 40, 505–514. (In Chinese) [Google Scholar] [CrossRef]
- Li, Q.; Wang, J.; Sun, H.Y.; Shang, X. Flower color patterning in pansy (Viola × wittrockiana Gams.) is caused by the differential expression of three genes from the anthocyanin pathway in acyanic and cyanic flower areas. Plant Physiol. Biochem. 2014, 84, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Albert, N.W.; Zhang, H.B.; Arathoon, S.; Boase, M.R.; Ngo, H.; Schwinn, K.K.; Davies, K.M.; Lewis, D.H. Temporal and spatial regulation of anthocyanin biosynthesis provide diverse flower colour intensities and patterning in Cymbidium orchid. Planta 2014, 240, 983–1002. [Google Scholar] [CrossRef] [PubMed]
- Zhong, P.X.; Wang, L.S.; Li, S.S.; Xu, Y.J.; Zhu, M.L. The changes of floral color and pigments composition during the flowering period in Paeonia lactiflora Pallas. Acta Horic. Sin. 2012, 39, 2271–2282. (In Chinese) [Google Scholar] [CrossRef]
- Gordillo, B.; Sigurdson, G.T.; Lao, F.; González-Miret, M.L.; Heredia, F.J.; Giusti, M.M. Assessment of the color modulation and stability of naturally copigmented anthocyanin-grape colorants with different level of purification. Food Res. Int. 2018, 106, 791–799. [Google Scholar] [CrossRef]
- Yager, R.R. Toward a language for specifying summarizing statistics. IEEE Trans. Syst. Man Cybern. B Cybern. 2003, 33, 177–187. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Chen, C.; Hu, W.Y.; Chen, L.J.; Zhan, J.T. Influence of source credibility on consumer acceptance of genetically modified foods in China. Sustainability 2016, 8, 899. [Google Scholar] [CrossRef]
- Georne, W.R.G. Nonlinear decision weights in choice under uncertainty. Manag. Sci. 1999, 45, 74–85. [Google Scholar]
- Oertel, A.; Matros, A.; Hartmann, A.; Arapitsas, P.; Dehmer, K.J.; Marten, S.; Mock, H.P. Metabolite profiling of red and blue potatoes revealed cultivar and tissue specific patterns for anthocyanins and other polyphenols. Planta 2017, 246, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Koyama, K.; Kamiqakiuchi, H.; Iwashita, K.; Mochioka, R.; Goto-Yamamoto, N. Polyphenolic diversity and characterization in the redpurple berries of East Asian wild Vitis species. Phytochemistry 2017, 134, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Xu, Y.X.; Zhang, D.L.; Zhuang, H.F.; Wu, J.Q.; Sun, G.L. An acyltransferase gene that putatively functions in anthocyanin modification was horizontally transferred from Fabaceae into the genus Cuscuta. Plant Divers. 2016, 38, 149–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Sun, Y.; Huang, J.R. Anthocyanin modification in Arabidopsis. Plant Physiol. J. 2013, 49, 101–110. (In Chinese) [Google Scholar] [CrossRef]
- Sasaki, N.; Matsuba, Y.; Abe, Y.; Okamura, M.; Momose, M.; Umemoto, N.; Nakayama, M.; Itoh, Y.; Ozeke, Y. Recent advances in understanding the anthocyanin modification steps in carnation flowers. Sci. Hortic. 2013, 163, 37–45. [Google Scholar] [CrossRef]
- Zhang, B.; Yang, X.S.; Li, N.N.; Zhu, X.; Sheng, W.J.; He, F.; Duan, C.Q.; Han, S.Y. Colorimetric study of malvidin-3-O-glucoside copigmented by phenolic compounds: The effect of molar ratio, temperature, pH, and ethanol content on color expression of red wine model solutions. Food Res. Int. 2017, 102, 468–477. [Google Scholar] [CrossRef]
- Lorenzis, G.D.; Rustioni, L.; Parisi, S.G.; Zoli, F.; Brancadoro, L. Anthocyanin biosynthesis during berry development in corvina grape. Sci. Hortic. 2016, 212, 74–80. [Google Scholar] [CrossRef]
- Gu, Z.Y.; Chen, H.; Yang, R.N.; Ran, M.H. Identification of DFR as a promoter of anthocyanin accumulation in poinsettia (Euphorbia pulcherrima, willd. Ex Klotzsch) bracts under short-day conditions. Sci. Hortic. 2018, 236, 158–165. [Google Scholar] [CrossRef]
- Valiñas, M.A.; Lanteri, M.L.; Have, A.T.; Andreu, A.B. Chlorogenic acid, anthocyanin and flavan-3-ol biosynthesis in flesh and skin of Andean potato tubers (Solanum tuberosum subsp. andigena). Food Chem. 2017, 229, 837–846. [Google Scholar] [CrossRef]
- Carmona, L.; Alquézar, B.; Marques, V.V.; Peña, L. Anthocyanin biosynthesis and accumulation in blood oranges during postharvest storage at different low temperatures. Food Chem. 2017, 237, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Olivares, D.; Contreras, C.; Muñoz, V.; Rivera, S.; González-Agüero, M.; Retamales, J.; Defilippia, B.G. Relationship among color development, anthocyanin and pigment-related gene expression in ‘Crimson Seedless’ grapes treated with abscisic acid and sucrose. Plant Physiol. Biochem. 2017, 115, 286–297. [Google Scholar] [CrossRef] [PubMed]
- Rosas, I.D.; Ponce, M.T.; Malovini, E.; Deis, L.; Cavagnaro, B.; Cavagnaro, P. Loss of anthocyanins and modification of the anthocyanin profiles in grape berries of Malbec and Bonarda grown under high temperature conditions. Plant Sci. 2017, 258, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, Y.H.; Hsieh, J.F. The conversion and deglycosylation of isoflavones and anthocyanins in black soymilk process. Food Chem. 2018, 261, 8–14. [Google Scholar] [CrossRef]
- Cao, K.; Ding, T.Y.; Mao, D.M.; Zhu, G.R.; Fang, W.C.; Chen, C.W.; Wang, X.W.; Wang, L.R. Transcriptome analysis reveals novel genes involved in anthocyanin biosynthesis in the flesh of peach. Plant Physiol. Biochnol. 2018, 123, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Lou, Q.; Liu, Y.L.; Qi, Y.Y.; Jiao, S.Z.; Tian, F.F.; Jiang, L.; Wang, Y.J. Transcriptome sequencing and metabolite analysis reveals the role of delphinidin metabolism in flower colour in grape hyacinth. J. Exp. Bot. 2014, 65, 3157–3164. [Google Scholar] [CrossRef] [PubMed]
- Margherita, B.; Giampiero, C.; Pierre, D. Field performance of tissue-cultured plants of Ranunculus asiaticus L. Sci. Hortic. 1996, 66, 229–239. [Google Scholar] [CrossRef]
- Hondo, K.; Sukhumpinij, P.; Kakihara, F. Flower color and pigments in yellow-flowered hybrid progeny raised from the interspecific cross Pelargonium quinquelobatum white-flowered geraniums. Sci. Hortic. 2015, 195, 145–153. [Google Scholar] [CrossRef]
- Giusti, M.M.; Wrolstad, R.E. Characterization and measurement of anthocyanins by UV-visible spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, F1.2.1–F1.2.13. [Google Scholar] [CrossRef]
- Du, L.J.; Wang, Q.P.; Shao, J.L.; Wang, L.; Liu, X.Y.; He, L.Z. Optimization on testing conditions of anthocyanins in Lycium ruthenicum by response surface methodology. Food Nutr. Chin. 2016, 22, 50–54. (In Chinese) [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef]
- Jin, X.H.; Fu, J.X.; Dai, S.L.; Sun, Y.; Hong, Y. Reference gene selection for qPCR analysis in cineraria developing flowers. Sci. Hortic. 2013, 153, 64–70. [Google Scholar] [CrossRef]
- Hellemans, J.; Mortier, G.; Paepe, A.D.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhong, H.Y.; Kuang, J.F.; Li, J.G.; Lu, W.J.; Chen, J.Y. Validation of reference genes for RT-qPCR studies of gene expression in banana fruit under different experimental conditions. Planta 2011, 234, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Andersen, C.L.; Jensen, J.L.; Ørntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: ‘A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets’. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| OP | L* | a* | b* | C* | MP | L* | a* | b* | C* | IP | L* | a* | b* | C* |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L1 | 52.9 ± 1.0 | 8.9 ± 0.8 | 35.4 ± 1.6 | 36.5 | L1 | 72.8 ± 1.9 | −8.5 ± 0.7 | 44.8 ± 1.6 | 45.6 | L1 | 71.2 ± 2.5 | −11.7 ± 2.0 | 51.1 ± 2.1 | 52.5 |
| L2 | 51.8 ± 0.8 | 38.4 ± 1.3 | 45.9 ± 1.2 | 59.8 | L2 | 71.5 ± 2.4 | −7.2 ± 1.0 | 49.4 ± 1.5 | 49.9 | L2 | 73.2 ± 2.1 | −10.9 ± 1.2 | 45.7 ± 1.8 | 47.0 |
| L3 | 42.3 ± 0.9 | 39.9 ± 1.2 | 37.5 ± 1.3 | 54.8 | L3 | 70.2 ± 2.0 | −2.9 ± 0.7 | 39.4 ± 1.6 | 39.5 | L3 | 75.3 ± 2.2 | −8.2 ± 1.2 | 32.0 ± 1.7 | 33.0 |
| L4 | 37.1 ± 1.5 | 40.1 ± 1.3 | 28.1 ± 0.7 | 49.0 | L4 | 50.5 ± 1.7 | 18.7 ± 1.4 | 41.6 ± 1.8 | 45.6 | L4 | 76.1 ± 1.9 | −8.0 ± 1.1 | 40.2 ± 1.8 | 41.0 |
| L5 | 45.8 ± 2.1 | 43.3 ± 0.9 | 40.0 ± 1.0 | 59.0 | L5 | 46.1 ± 1.3 | 24.5 ± 1.3 | 37.6 ± 1.8 | 44.9 | L5 | 74.1 ± 2.0 | −9.5 ± 1.4 | 40.7 ± 1.9 | 41.8 |
| L6 | 49.2 ± 1.2 | 41.9 ± 1.3 | 44.1 ± 1.2 | 60.8 | L6 | 42.6 ± 1.5 | 27.4 ± 1.2 | 31.5 ± 1.3 | 41.8 | L6 | 70.8 ± 2.1 | −9.8 ± 1.1 | 48.0 ± 1.8 | 49.0 |
| L7 | 52.0 ± 1.1 | 36.3 ± 0.8 | 45.8 ± 0.8 | 58.4 | L7 | 43.8 ± 1.7 | 30.0 ± 1.8 | 33.3 ± 1.8 | 44.9 | L7 | 71.4 ± 2.0 | −10.8 ± 1.0 | 46.8 ± 1.8 | 48.0 |
| L8 | 50.9 ± 0.7 | 38.5 ± 0.6 | 42.2 ± 0.7 | 57.2 | L8 | 45.0 ± 1.7 | 29.0 ± 1.2 | 35.8 ± 2.0 | 46.0 | L8 | 66.4 ± 2.0 | −10.4 ± 1.0 | 44.5 ± 1.6 | 45.7 |
| L9 | 50.1 ± 0.9 | 38.9 ± 0.4 | 42.9 ± 0.7 | 57.9 | L9 | 45.4 ± 1.6 | 34.7 ± 1.1 | 39.8 ± 1.2 | 52.8 | L9 | 61.0 ± 2.4 | −7.1 ± 1.2 | 40.2 ± 2.0 | 40.8 |
| L10 | 50.7 ± 1.1 | 34.2 ± 0.9 | 41.2 ± 1.5 | 53.5 | L10 | 54.9 ± 2.0 | 33.9 ± 1.7 | 52.8 ± 1.0 | 62.8 | L10 | 66.4 ± 1.9 | 0.2 ± 0.5 | 50.5 ± 1.6 | 50.5 |
| L11 | 46.6 ± 0.8 | 28.7 ± 1.0 | 34.2 ± 1.3 | 44.7 | L11 | 54.0 ± 1.4 | 33.6 ± 1.5 | 49.7 ± 1.4 | 60.0 | L11 | 52.3 ± 1.8 | 18.1 ± 1.4 | 30.0 ± 1.5 | 35.0 |
| L12 | 46.3 ± 1.5 | 28.9 ± 0.9 | 32.7 ± 1.2 | 43.6 | L12 | 50.9 ± 1.4 | 30.3 ± 1.2 | 38.5 ± 1.4 | 49.0 | L12 | 51.7 ± 1.3 | 16.4 ± 1.2 | 29.3 ± 1.3 | 33.6 |
| C1 | 67.0 ± 1.1 | 17.1 ± 1.1 | 10.3 ± 0.7 | 19.9 | C1 | 80.0 ± 2.1 | −6.5 ± 1.3 | 28.4 ± 1.6 | 29.1 | C1 | 78.0 ± 2.0 | −9.6 ± 1.1 | 43.0 ± 1.7 | 44.1 |
| C2 | 64.6 ± 0.9 | 25.8 ± 1.3 | 2.7 ± 1.1 | 26.0 | C2 | 79.2 ± 1.6 | −5.8 ± 1.1 | 27.8 ± 2.0 | 28.4 | C2 | 77.3 ± 2.2 | −8.8 ± 1.3 | 35.5 ± 1.7 | 36.6 |
| C3 | 64.5 ± 1.7 | 28.3 ± 1.5 | −1.7 ± 0.8 | 28.3 | C3 | 70.7 ± 2.0 | 6.9 ± 1.1 | 16.6 ± 1.2 | 17.9 | C3 | 79.1 ± 2.0 | −8.0 ± 1.0 | 32.7 ± 1.6 | 33.7 |
| C4 | 64.6 ± 1.2 | 29.3 ± 2.0 | −1.2 ±0.2 | 29.3 | C4 | 70.2 ± 1.9 | 9.1 ± 1.0 | 14.5 ± 1.2 | 17.1 | C4 | 76.5 ± 1.9 | −8.8 ± 1.2 | 36.4 ± 1.7 | 37.5 |
| C5 | 66.3 ± 0.8 | 27.6 ± 1.0 | −2.0 ± 0.3 | 27.7 | C5 | 68.2 ± 1.7 | 9.5 ± 1.4 | 15.1 ± 1.2 | 17.8 | C5 | 76.9 ± 1.7 | −9.6 ± 1.2 | 38.3 ± 1.4 | 39.5 |
| C6 | 66.8 ± 1.0 | 26.5 ± 0.6 | −1.5 ± 0.3 | 26.5 | C6 | 69.1 ± 2.2 | 11.3 ± 0.6 | 11.4 ± 0.8 | 16.0 | C6 | 73.6 ± 2.1 | −8.5 ± 1.1 | 35.1 ± 1.3 | 36.1 |
| C7 | 69.3 ± 0.8 | 24.3 ± 0.6 | −0.4 ± 0.3 | 24.3 | C7 | 65.7 ± 2.3 | 26.7 ± 1.0 | 1.0 ± 0.7 | 26.7 | C7 | 72.8 ± 1.9 | −8.3 ± 1.0 | 32.6 ± 1.4 | 33.7 |
| C8 | 73.2 ± 0.7 | 18.3 ± 0.7 | 1.1 ± 0.2 | 18.3 | C8 | 64.7 ± 1.6 | 22.5 ± 1.6 | 3.3 ± 0.9 | 22.8 | C8 | 72.9 ± 2.0 | −9.1 ± 1.0 | 35.0 ± 1.7 | 36.2 |
| C9 | 67.6 ± 0.8 | 25.8 ± 0.6 | −1.8 ± 0.2 | 25.9 | C9 | 71.4 ± 2.3 | 21.1 ± 1.0 | 1.9 ± 0.8 | 21.2 | C9 | 80.9 ± 1.8 | −8.2 ± 1.1 | 32.7 ± 1.7 | 33.7 |
| C10 | 67.2 ± 1.1 | 23.1 ± 0.5 | −1.6 ± 0.2 | 23.2 | C10 | 68.8 ± 1.8 | 23.3 ± 1.2 | −2.4 ± 0.9 | 23.4 | C10 | 70.7 ± 1.9 | −6.3 ± 1.2 | 32.9 ± 1.8 | 33.4 |
| C11 | 64.5 ± 1.5 | 23.4 ± 0.9 | −2.2 ± 0.5 | 23.5 | C11 | 64.9 ± 2.1 | 28.7 ± 1.1 | −3.0 ± 1.2 | 28.9 | C11 | 69.3 ± 1.4 | −2.5 ± 1.2 | 29.4 ± 1.4 | 29.5 |
| C12 | 66.0 ± 1.7 | 20.9 ± 1.5 | −2.0 ± 0.4 | 21.0 | C12 | 63.1 ± 1.4 | 29.0 ± 0.9 | −3.5 ± 1.1 | 29.2 | C12 | 64.1 ± 1.3 | 24.4 ± 1.1 | 4.1 ± 1.1 | 24.7 |
| Protein Name | Gene Name | Unigene Number 1 | |
|---|---|---|---|
| Backbone forming | chalcone synthase | CHS | 13 (9) |
| chalcone isomerase | CHI | 3 (3) | |
| flavanone 3-hydroxylase | F3H | 32 (21) | |
| flavanoid 3′-hydroxylase | F3′H | 5 (1) | |
| flavanoid 3′5′-hydroxylase | F3′5′H | 9 (6) | |
| dihydroflavonol 4-reductase | DFR | 39 (27) | |
| anthocyanidin synthase | ANS | 1 (1) | |
| Anthocyanidin modification | anthocyanidin 3-O-glucoside-2′′-O-glucosyltransferase | 3GGT (UGAT) | 20 (2) |
| anthocyanidin 3-O-glucosyltransferase | 3GT (GT, UFGT) | 18 (1) | |
| anthocyanidin 5,3-O-glucosyltransferase | 3,5GT (GT1) | 6 (0) | |
| anthocyanidin 3-O-glucoside-2′’-O-xylosyltransferase | 3G2′′GT (UGT79B1, A3G2XYLT) | 6 (3) | |
| anthocyanidin-3-O-glucoside-2′′-O-rhamnosyltransferase | 3GRT (RT) | 6 (2) | |
| anthocyanidin-5-O-glucosyltransferase | 5GT (HGT8, UGT75C1) | 4 (0) | |
| anthocyanidin-3-O-glucoside-3′-O-beta-glucosyltransferase | 3′GT | 1 (0) | |
| anthocyanin 3-O-glucoside-6′′-O-coumaroyltransferase | 3AT | 5 (0) | |
| anthocyanin 3-O-glucoside-6′′-O-malonyltransferase | 3MAT | 3 (0) | |
| anthocyanin 5-O-glucoside-6′′′′-aromatic acyltransferase | 5AT | 3 (0) | |
| flavonoid O-methyltransferase | FMT | 2 (0) |
| L* | a* | b* | TA | |
|---|---|---|---|---|
| a* | −0.905 ** | |||
| b* | 0.487 ** | −0.257 | ||
| TA | −0.783 ** | 0.914 ** | −0.110 | |
| c72570 | −0.905 ** | 0.944 ** | −0.302 | 0.871 ** |
| L* | a* | b* | TA | c130622 | |
|---|---|---|---|---|---|
| a* | −0.867 ** | ||||
| b* | 0.825 ** | −0.984 ** | |||
| TA | −0.777 ** | 0.960 ** | −0.960 ** | ||
| c130622 | −0.782 ** | 0.893 ** | −0.876 ** | 0.845 ** | |
| c83020 | −0.697 ** | 0.871 ** | −0.862 ** | 0.906 ** | 0.797 ** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Lin, T.; Du, L.; Wang, J.; Yang, X.; Zhang, J.; Zhang, P.; Li, Y.; Shi, J.; Yang, X. Sampling for DUS Test of Flower Colors of Ranunculus asiaticus L. in View of Spatial and Temporal Changes of Flower Colorations, Anthocyanin Contents, and Gene Expression Levels. Molecules 2019, 24, 615. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24030615
Liu Y, Lin T, Du L, Wang J, Yang X, Zhang J, Zhang P, Li Y, Shi J, Yang X. Sampling for DUS Test of Flower Colors of Ranunculus asiaticus L. in View of Spatial and Temporal Changes of Flower Colorations, Anthocyanin Contents, and Gene Expression Levels. Molecules. 2019; 24(3):615. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24030615
Chicago/Turabian StyleLiu, Yanfang, Tao Lin, Lijuan Du, Jiangmin Wang, Xiaohong Yang, Jianhua Zhang, Peng Zhang, Yangang Li, Junfeng Shi, and Xuhong Yang. 2019. "Sampling for DUS Test of Flower Colors of Ranunculus asiaticus L. in View of Spatial and Temporal Changes of Flower Colorations, Anthocyanin Contents, and Gene Expression Levels" Molecules 24, no. 3: 615. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24030615