Isolation of a Novel Flavanonol and an Alkylresorcinol with Highly Potent Anti-Trypanosomal Activity from Libyan Propolis

 ,
,  , and
, and
Abstract
:
1. Introduction
2. Results
2.1. Anti-Helminthic Activity of the Propolis Samples
2.2. Isolation and Characterization of Compounds from Samples P1, P2, P7 and P9
2.3. Characterization of Compound 4 as 3-Acetoxy-5,7,3′-trihydroxy-4′-methoxyflavanone or Taxifolin-3-acetyl-4′-methyl ether
2.4. Testing of the Antiparasite Activity of the Isolated Compounds
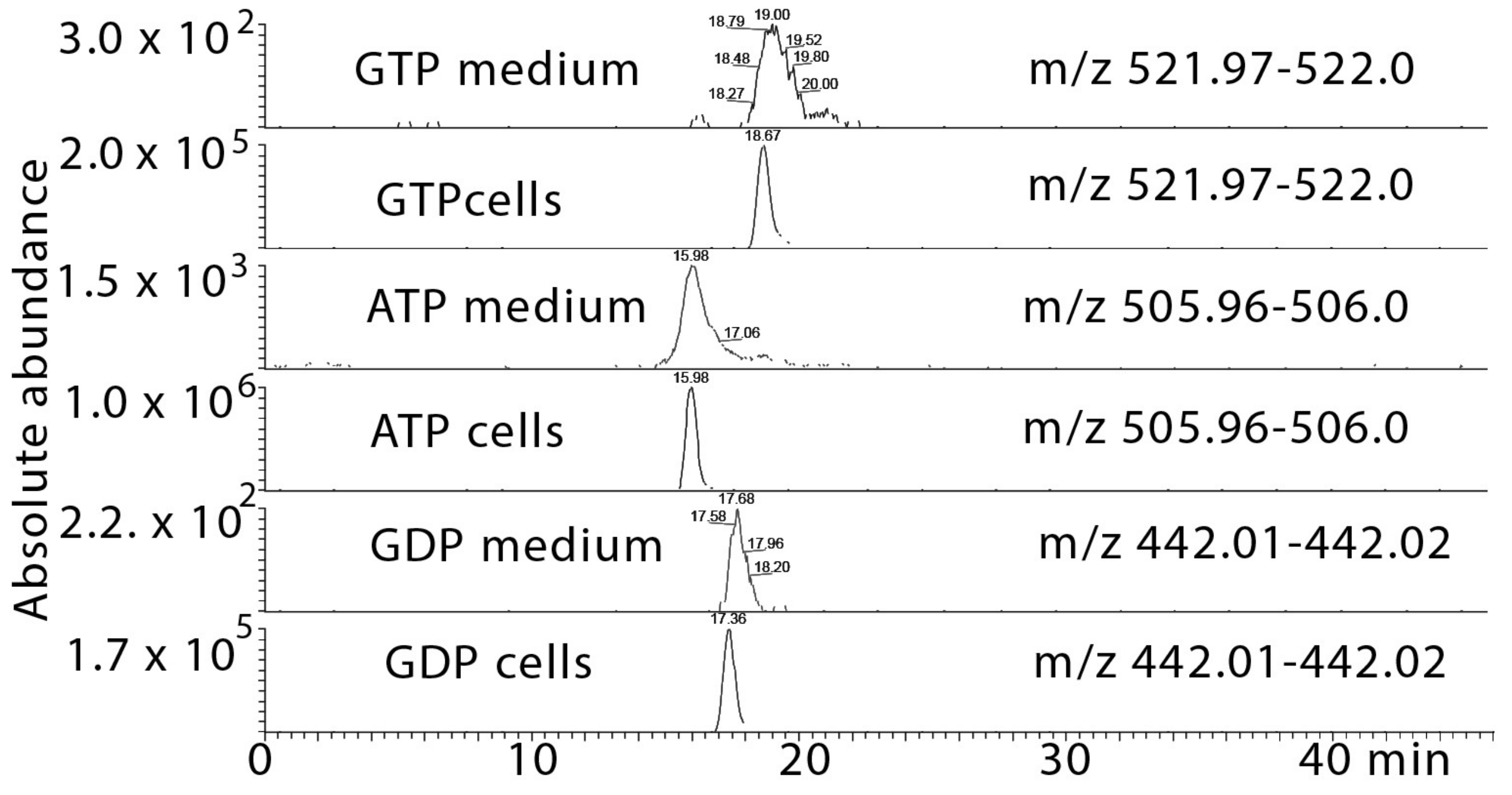
2.5. Metabolomic Profiling of the Effect of the Cardol-Rich Fraction 10 against T. brucei
3. Discussion
4. Materials and Methods
4.1. Reagents and Materials
4.2. Extraction of Propolis Samples
4.3. Column Chromatography
4.4. Purification of Column Fractions Using MPLC
4.5. Liquid Chromatography-High Resolution Mass Spectrometry (LC-HRMS) and HPLC with UV/Evaporative Light Scattering Detection (ELSD)
4.6. Nuclear Magnetic Resonance (NMR) Spectroscopy
4.7. Software and Data Processing for LC-HR-MS
4.8. In Vitro Testing against T. brucei and P. falciparum and Cytotoxicity
4.9. In Vitro Anti-Helminthic Activity of P1–P12 Against Trichinella spiralis
4.10. Metabolomics Experiments
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Almutairi, S.; Eapen, B.; Chundi, S.M.; Akhalil, A.; Siheri, W.; Clements, C.; Fearnley, J.; Watson, D.G.; Edrada-Ebel, R. New anti-trypanosomal active prenylated compounds from African propolis. Phytochem. Lett. 2014, 10, 35–39. [Google Scholar] [CrossRef]
- Falcão, S.I.; Vale, N.; Cos, P.; Gomes, P.; Freire, C.; Maes, L.; Vilas-Boas, M. In vitro evaluation of Portuguese propolis and floral sources for antiprotozoal, antibacterial and antifungal activity. Phytother. Res. 2014, 28, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Omar, R.M.; Igoli, J.; Gray, A.I.; Ebiloma, G.U.; Clements, C.; Fearnley, J.; Ebel, R.A.; Zhang, T.; De Koning, H.P.; Watson, D.G. Chemical characterisation of Nigerian red propolis and its biological activity against Trypanosoma brucei. Phytochem. Anal. 2016, 27, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Siheri, W.; Igoli, J.O.; Gray, A.I.; Nasciemento, T.G.; Zhang, T.; Fearnley, J.; Clements, C.J.; Carter, K.C.; Carruthers, J.; Edrada-Ebel, R.; et al. The isolation of antiprotozoal compounds from Libyan propolis. Phytother. Res. 2014, 28, 1756–1760. [Google Scholar] [CrossRef] [PubMed]
- Olayemi, K.I. Therapeutic potentials of Nigerian insect-propolis against the malarial parasite, Plasmodium berghei (Haemosporida: Plasmodidae). Am. J. Drug Disc. Dev. 2014, 4, 241–247. [Google Scholar] [CrossRef]
- Siheri, W.; Zhang, T.; Ebiloma, G.U.; Biddau, M.; Woods, N.; Hussain, M.Y.; Clements, C.J.; Fearnley, J.; Ebel, R.E.; Paget, T.; et al. Chemical and antimicrobial profiling of propolis from different regions within Libya. PLoS ONE 2016, 11, e0155355. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Fattah, N.S.; Nada, O.H. Effect of propolis versus metronidazole and their combined use in treatment of acute experimental giardiasis. J. Egypt Soc. Parasitol. 2007, 37, 691–710. [Google Scholar] [PubMed]
- Alday-Provencio, S.; Diaz, G.; Rascon, L.; Quintero, J.; Alday, E.; Robles-Zepeda, R.; Garibay-Escobar, A.; Astiazaran, H.; Hernandez, J.; Velazquez, C. Sonoran propolis and some of its chemical constituents inhibit in vitro growth of Giardia lamblia trophozoites. Planta Med. 2015, 81, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Hassan, S.E.; Abou-El-Dobal, S.K.; Hegazi, A.G. Bioassay of Egyptian propolis on Toxocara vitulorum adult worms. World Appl. Sci. J. 2016, 34, 283–289. [Google Scholar]
- Siheri, W.; Alenezi, S.; Tusiimire, J.; Watson, D.G. The chemical and biological properties of propolis. In Bee Products-Chemical and Biological Properties; Springer: Cham, Switzerland, 2017; pp. 137–178. [Google Scholar]
- Abd El-Rahman, S.S. West-Libyan propolis and rosemary have synergistic anti-tumor effect against 12-O-tetradecanoylphorbol 13-acetate-induced skin tumor in BULB/C mice previously initiated with 7,12-dimethylbenz [a] anthracene. Basic Appl. Pathol. 2010, 3, 46–51. [Google Scholar] [CrossRef]
- Sarkez, N.H. Antimicrobial properties of Libyan propolis against Staphylococcus aureus. Libyan J. Med. Res. 2014, 8, 50–55. [Google Scholar]
- Azab, E.A.; Algridi, M.A.; Lashkham, N.M. Hypolipidemic and antiatherogenic effects of aqueous extract of Libyan propolis in lead acetate intoxicated male albino mice. IJSR 2015, 4, 1060–1068. [Google Scholar]
- Azab, A.E.; Lashkham, N.M.; Albasha, M.O. Haematoprotective and hypolipidemic effects of aqueous extract of Libyan propolis against sodium nitrite induced haematotoxicity and hyperlipidemia in Guinea pigs. Am. J. Biosci. Bioeng. 2015, 3, 22–32. [Google Scholar]
- Stevens, J.F.; Wollenweber, E.; Ivancic, M.; Hsu, V.L.; Sundberg, S.; Deinzer, M.L. Leaf surface flavonoids of Chrysothamnus. Phytochemistry 1999, 51, 771–780. [Google Scholar] [CrossRef]
- Bridges, D.; Gould, M.K.; Nerima, B.; Mäser, P.; Burchmore, R.J.S.; De Koning, H.P. Loss of the high affinity pentamidine transporter is responsible for high levels of cross-resistance between arsenical and diamidine drugs in African trypanosomes. Mol. Pharmacol. 2007, 71, 1098–1108. [Google Scholar] [CrossRef] [PubMed]
- Popova, M.P.; Graikou, K.; Chinou, I.; Bankova, V.S. GC-MS profiling of diterpene compounds in Mediterranean propolis from Greece. J. Agric. Food Chem. 2010, 58, 3167–3176. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Fang, J.; Cheng, Y. Labdanes from Cryptomeria japonica. Phytochemistry 1994, 37, 1109–1114. [Google Scholar]
- Kardar, M.N.; Zhang, T.; Coxon, G.D.; Watson, D.G.; Fearnley, J.; Seidel, V. Characterisation of triterpenes and new phenolic lipids in Cameroonian propolis. Phytochemistry 2014, 106, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Petrova, A.; Popova, M.; Kuzmanova, C.; Tsvetkova, I.; Naydenski, H.; Muli, E.; Bankova, V. New biologically active compounds from Kenyan propolis. Fitoterapia 2010, 81, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Omar, R.; Siheri, W.; Al Mutairi, S.; Clements, C.; Fearnley, J.; Edrada-Ebel, R.; Watson, D. Chromatographic analysis with different detectors in the chemical characterisation and dereplication of African propolis. Talanta 2014, 120, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Trusheva, B.; Popova, M.; Koendhori, E.B.; Tsvetkova, I.; Naydenski, C.; Bankova, V. Indonesian propolis: Chemical composition, biological activity and botanical origin. Nat. Prod. Res. 2011, 25, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Maia, F.J.N.; Ribeiro, V.G.P.; Almeida, M.O.; Lomonaco, D.; Mafezoli, J.; Mazzetto, S.E. Study of antioxidant activity of a phenyl phosphorylated compound derived from hydrogenated cardol by thermogravimetric analysis. Br. J. Appl. Sci. Technol. 2013, 3, 546. [Google Scholar] [CrossRef]
- Silva, M.S.S.; De Lima, S.G.; Oliveira, E.H.; Lopes, J.A.D.; Chaves, M.H.; Reis, F.A.M.; Citó, A.M.G.L. Anacardic acid derivatives from Brazilian propolis and their antibacterial activity. Ecl. Quím. São Paulo 2008, 33, 53–58. [Google Scholar] [CrossRef]
- Rakotomanga, M.; Blanc, S.; Gaudin, K.; Chaminade, P.; Loiseau, P.M. Miltefosine affects lipid metabolism in Leishmania donovani promastigotes. Antimicrob. Agents Chemother. 2007, 51, 1425–1430. [Google Scholar] [CrossRef] [PubMed]
- Kozubek, A.; Tyman, J.H. Resorcinolic lipids, the natural non-isoprenoid phenolic amphiphiles and their biological activity. Chem. Rev. 1999, 99, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Cho, Y.S.; Park, E.J.; Kim, J.; Oh, W.K.; Lee, H.S.; Ahn, J.S. Phospholipase Cγ1 inhibitory principles from the sarcotestas of Ginkgo biloba. J. Nat. Prod. 1998, 61, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Araim, Y.; Kimm, S.; Hamm, J.; Usukim, T. Synthesis and biological evaluation of bilobol and adipostatin A. J. Asian Nat. Prod. Res. 2011, 13, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Ebiloma, G.U.; Ayuga, T.D.; Balogun, E.O.; Gil, L.A.; Donachie, A.; Kaiser, M.; Herraiz, T.; Inaoka, D.K.; Shiba, T.; Harada, S.; et al. Inhibition of trypanosome alternative oxidase without its N-terminal mitochondrial targeting signal (ΔMTS-TAO) by cationic and non-cationic 4-hydroxybenzoate and 4-alkoxybenzaldehyde derivatives active against T. brucei and T. congolense. Eur. J. Med. Chem. 2018, 150, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Alonezi, S.; Tusiimire, J.; Wallace, J.; Dufton, M.J.; Parkinson, J.A.; Young, L.C.; Clements, C.J.; Park, J.K.; Jeon, J.W.; Ferro, V.A.; et al. Metabolomic profiling of the effects of melittin on cisplatin resistant and cisplatin sensitive ovarian cancer cells using mass spectrometry and biolog microarray technology. Metabolites 2016, 6, 35. [Google Scholar] [CrossRef] [PubMed]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Not available. |



{kind=link}
{kind=link}
{kind=link}
| Propolis Origin (Number of Samples) | Extract Investigated | Analysis Conducted | Biological Activity Observed |
|---|---|---|---|
| Surman City, West Libya, (One sample) | Ethanol extract/CAPE | TLC investigation | Cytotoxicity and Antioxidant activity [11] |
| Zawia City, West-Libya, (One sample) | Ethanol extract | Partial purification | Inhibitory effect against S. aureus [12] |
| Alaquria and Tokra, North East Libya (Two samples) | Ethanol extract and purified compounds | LC-MS-GC-MS, HPLC-UV/ELSD, NMR, diterpenes, lignin compounds | High activity against T. brucei, L. donovani [4] |
| Surman City, West Libya, (One sample) | Aqueous extract | None | In vivo hypolipidaemic and antiatherogenic effects in mice [13] |
| Surman City, West Libya (One sample) | Aqueous extract | None | Hepatoprotective and hypolipidemic effects in guinea pigs [14] |
| Different Geographic areas West, East, South East and South West of Libya (12 samples) | Ethanol extract | LC-MS, PCA, analysis combined with HCA | Activity against T. brucei, L. donovani, P. falciparum, C. fasciculata and M. marinum [6] |
| Propolis Samples | Inhibition of T. spiralis 1 µg/mL | 10 µg/mL |
|---|---|---|
| P1 | 19.5 ± 3.5 | 63.1 ± 0.5 |
| P2 | 38.9 ± 0.1 | 57.2 ± 6.4 |
| P3 | 51.9 ± 0.1 | 63.6 ± 0.3 |
| P4 | 59.3 ± 0.1 | 61.3 ± 0.7 |
| P5 | 4.77 ± 0.21 | 56.2 ± 5.2 |
| Nitazoxanide a | 28.7 ± 11.8 | |
| Levamisole b | 56.7 ± 3.9 |
| No. | Yield (mg) | Name | Propolis Sample | Molecular Formula | m/z -Ve Ion | Class of Compound |
|---|---|---|---|---|---|---|
| (1) | 17.7 | 13-Epitorulosolol | P1 | C20H34O2 | 305.4812 | Diterpene |
| (2) | 22.7 | Demethylpiperitol | P2 | C19H18O6 | 341.1211 | Lignan |
| (3) | 20.3 | 5′-Methoxypiperitol | P2 | C21H22O7 | 385.1136 | Lignan |
| (4) | 17.7 | Taxifolin-3-acetyl-4′methyl ether | P1 | C18H16O8 | 359.0766 | Flavanone |
| (5) | 25.7 | Cycloartanol | P7 | C30H50O | 425.3821 | Cycloartane triterpene |
| (6) | 29.2 | Mangiferolic acid | P7 | C30H48O3 | 455.7123 | Cycloartane triterpene |
| (7) | 21.7 | Mangiferonic acid | P7 | C30H46O3 | 453.6934 | Cycloartane triterpene |
| (8) | 41.8 | Ambolic acid C31H51O3 | P7 | C31H50O3 | 469.729 | Cycloartane triterpene |
| (9) | 33.8 | 27-Hydroxymangiferonic acid | P7 | C30H46O4 | 469.6982 | Cycloartane triterpene |
| (10) | 37.8 | Cardol plus mangiferolic acid (6) | P7 | C21H34O2 | 317.2489, 455.7123 | Resorcinol, Cycloartane |
| (11) | 27.1 | Acetylisocupressic acid | P9 | C20H32O3 | 319.1711 | Diterpene |
| (12) | 25.4 | Agathadiol | P9 | C20H34O2 | 305.4838 | Diterpene |
| (13) | 22.3 | Isocupressic acid | P9 | C20H32O3 | 319.4791 | Diterpene |
| (14) | 22.2 | Isoagatholal | P9 | C20H32O2 | 303.2412 | Diterpene |
| Compound | T. brucei (s427) a | T. brucei (B48) a | P. falciparuma |
|---|---|---|---|
| (2) | 2.7 ± 0.2 | 2.68 ± 0.04 | 17.5 ± 0.1 |
| (3) | 13.1 ± 0.1 | 12.4 ± 1.6 | - |
| (5) | 3.7 ± 0.1 | 3.42 ± 0.08 | * |
| (6) | - | - | * |
| (7) | 14.6 ± 0.2 | 14.7 ± 0.4 | 49.2 ± 9.5 |
| (8) | - | - | * |
| (9) | 35.2 ± 0.6 | 34.9 ± 0.3 | * |
| (10) | 0.70 ± 0.03 | 0.70 ± 0.06 | 12.4 ± 2.1 |
| (11) | 25.0 ± 0.2 | 25.6 ± 0.9 | - |
| (12) | 7.0 ± 0.6 | 6.90 ± 0.45 | - |
| (13) | 3.0 ± 0.1 | 2.73 ± 0.11 | * |
| (14) | 10.4 ± 0.1 | 10.2 ± 0.8 | |
| Pentamidine 1 | 0.00012 ± 0.00003 | 0.255 ± 0.009 | - |
| Chloroquine 1 | - | - | 0.0034 ± 0.00003 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siheri, W.; Ebiloma, G.U.; Igoli, J.O.; Gray, A.I.; Biddau, M.; Akrachalanont, P.; Alenezi, S.; Alwashih, M.A.; Edrada-Ebel, R.; Muller, S.; et al. Isolation of a Novel Flavanonol and an Alkylresorcinol with Highly Potent Anti-Trypanosomal Activity from Libyan Propolis. Molecules 2019, 24, 1041. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24061041
Siheri W, Ebiloma GU, Igoli JO, Gray AI, Biddau M, Akrachalanont P, Alenezi S, Alwashih MA, Edrada-Ebel R, Muller S, et al. Isolation of a Novel Flavanonol and an Alkylresorcinol with Highly Potent Anti-Trypanosomal Activity from Libyan Propolis. Molecules. 2019; 24(6):1041. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24061041
Chicago/Turabian StyleSiheri, Weam, Godwin U. Ebiloma, John O. Igoli, Alexander I. Gray, Marco Biddau, Pilaslak Akrachalanont, Samya Alenezi, Mohammad A. Alwashih, RuAngelie Edrada-Ebel, Sylke Muller, and et al. 2019. "Isolation of a Novel Flavanonol and an Alkylresorcinol with Highly Potent Anti-Trypanosomal Activity from Libyan Propolis" Molecules 24, no. 6: 1041. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules24061041