
Activity Dependence of a Novel Lectin Family on Structure and Carbohydrate-Binding Properties

 ,
,
Abstract
:1. Introduction
2. Novel Lectin Family Structure
3. Glycan Specificity
4. Effect on Bacteria and Fungi
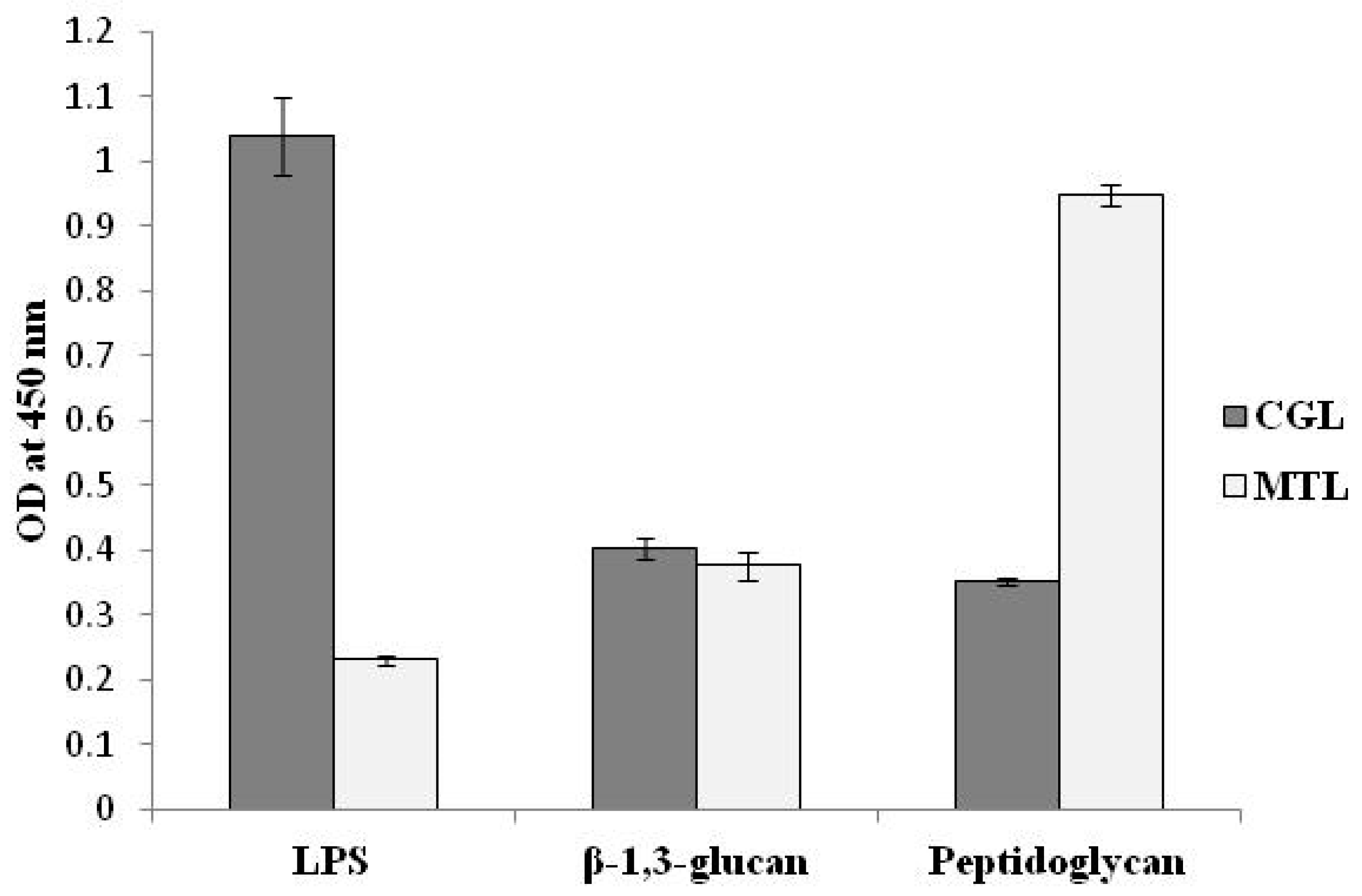
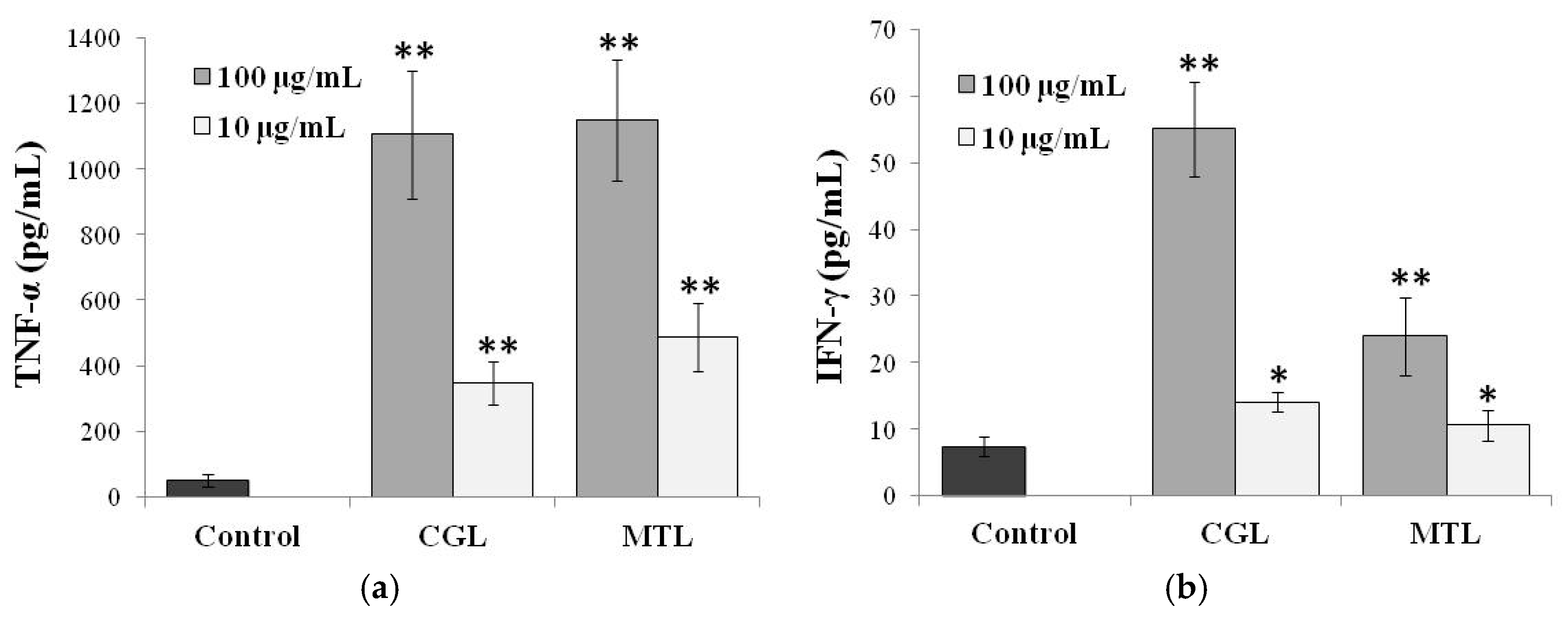
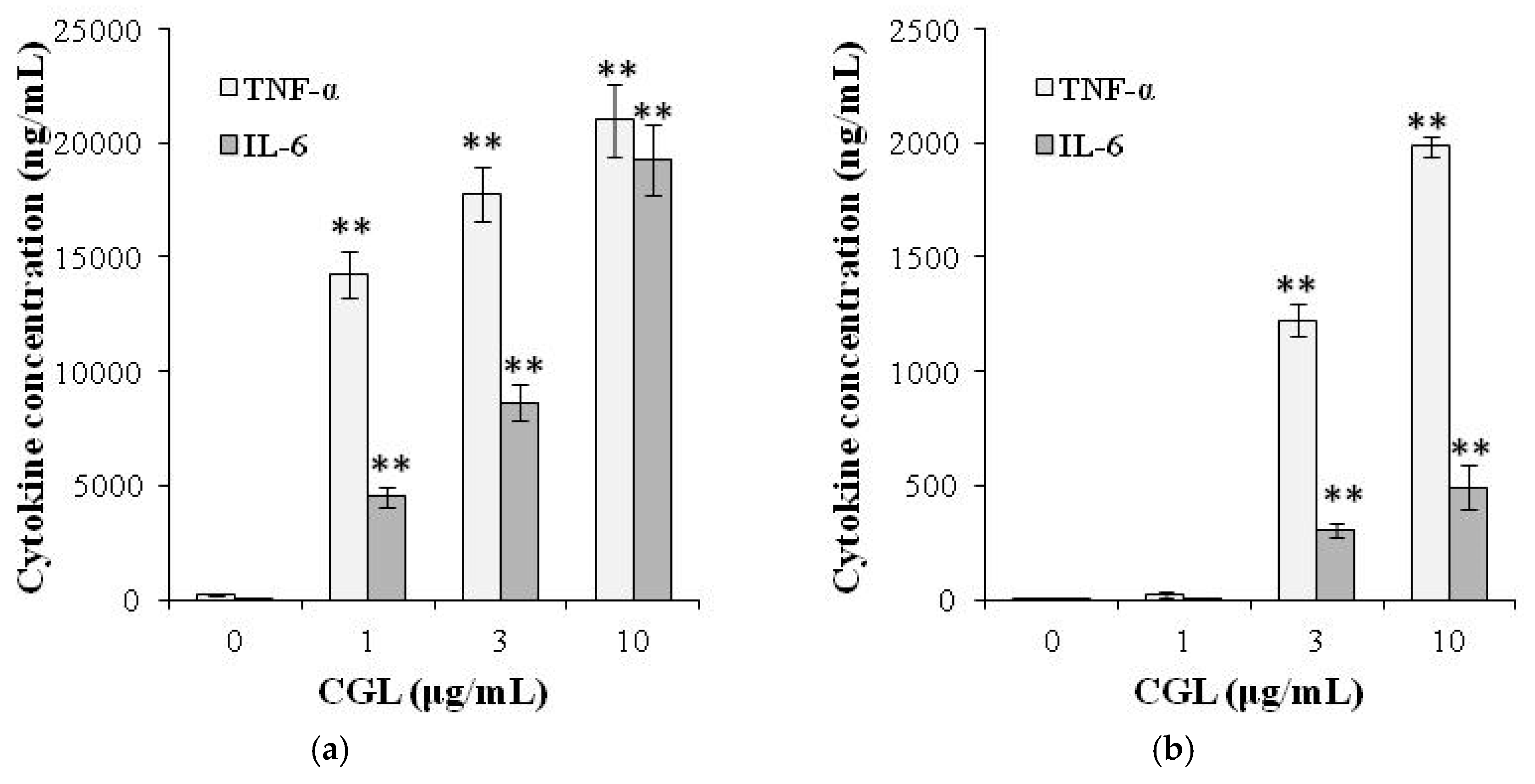
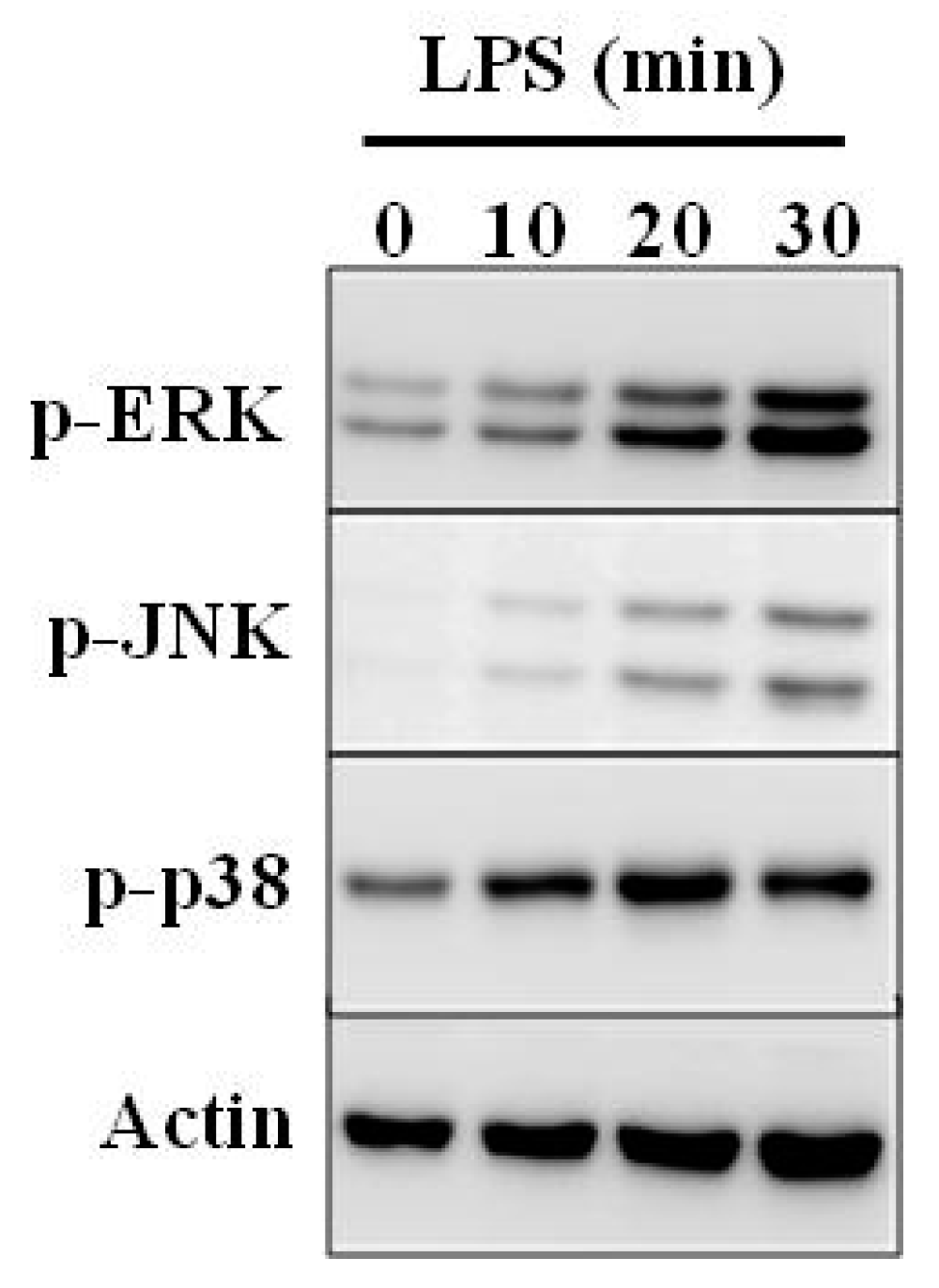
5. Immunomodulatory Activity
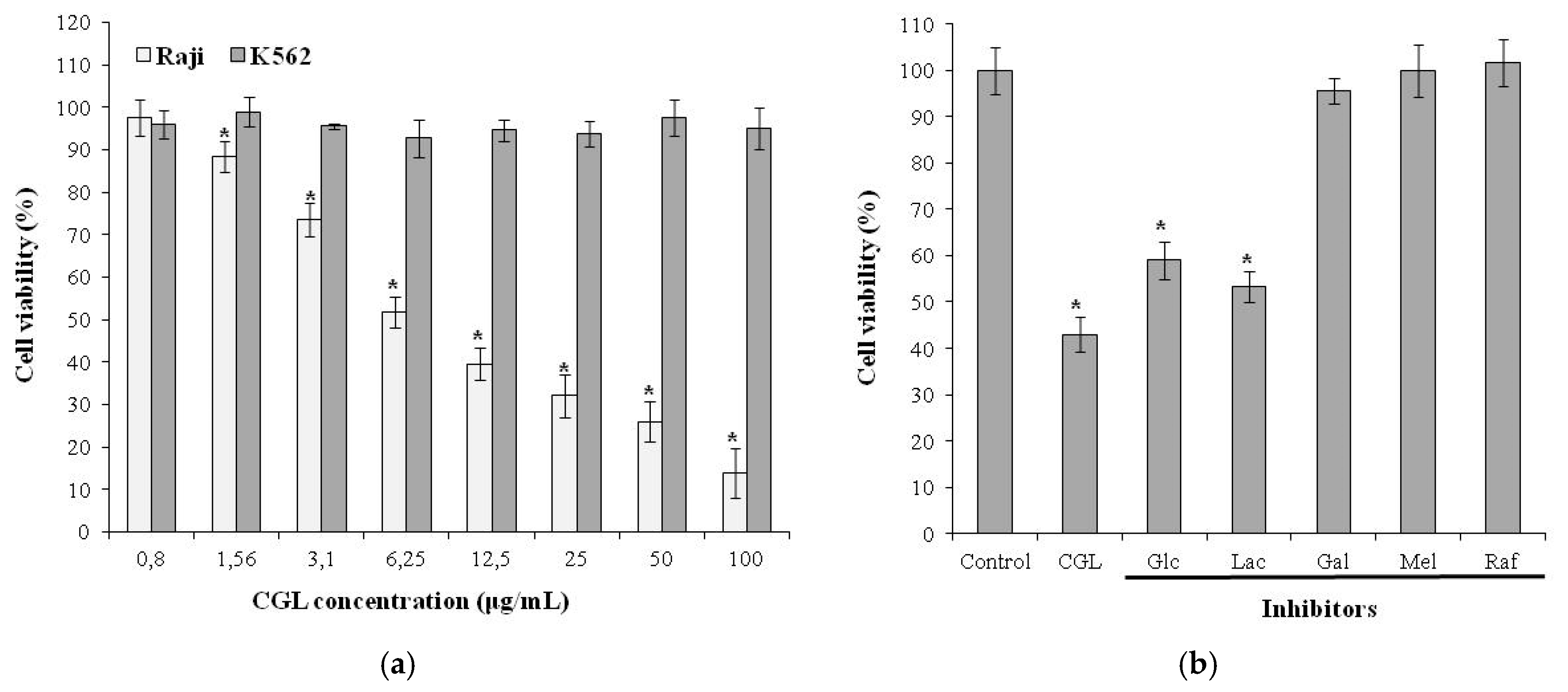
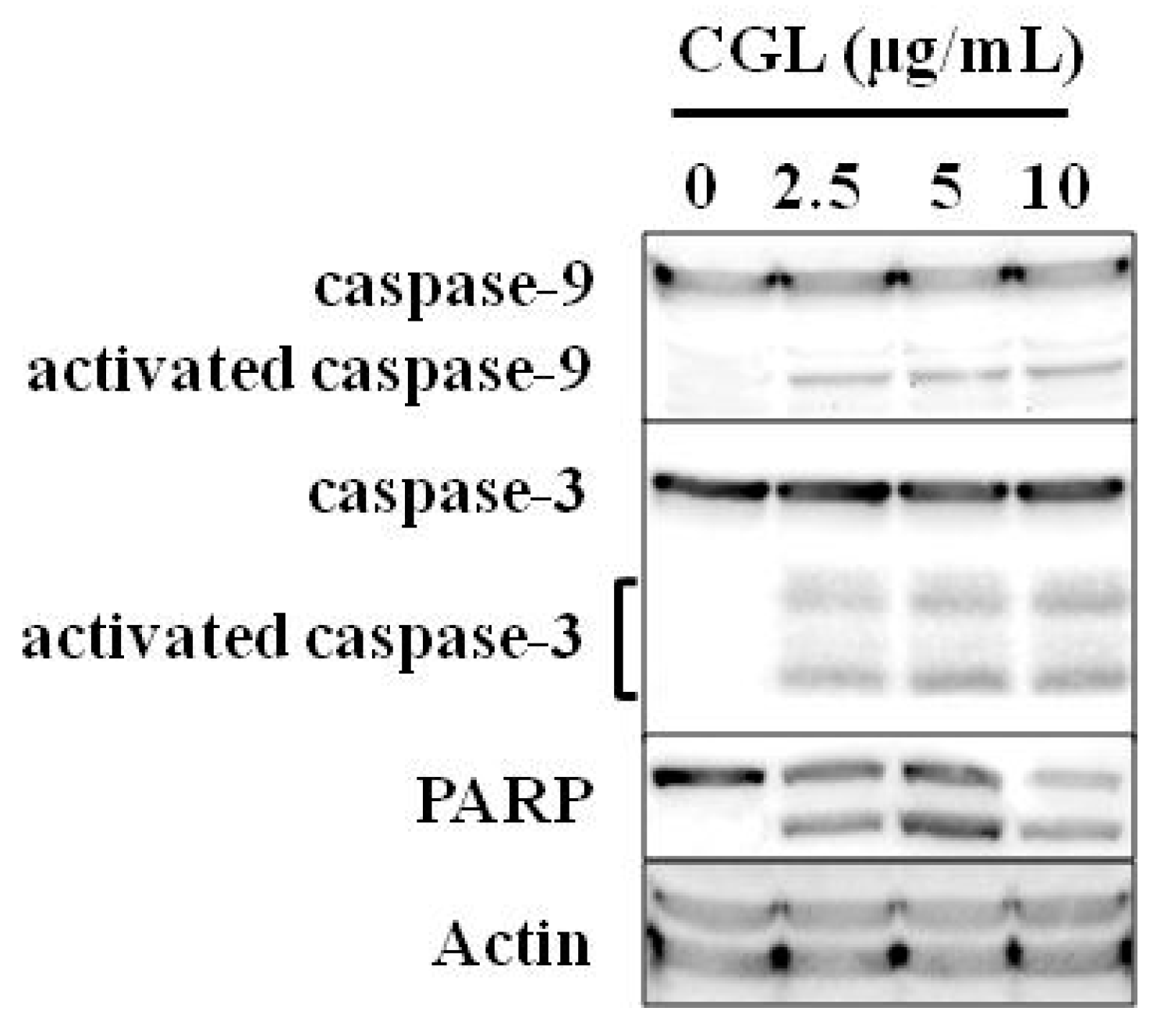
6. Effect on Tumor Cells
7. Conclusions
Funding
Conflicts of Interest
References
- Sharon, N.; Lis, H. Lectins; Springer: Dordrecht, The Netherlands, 2007; ISBN 978-1-4020-6605-4. [Google Scholar]
- McGreal, E.; Martinez-Pomares, L.; Gordon, S. Divergent roles for C-type lectins expressed by cells of the innate immune system. Mol. Immunol. 2004, 41, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N. Lectins: Carbohydrate-specific Reagents and Biological Recognition Molecules. J. Biol. Chem. 2007, 282, 2753–2764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Luo, Y.; Zhou, T.-T.; Zhang, W.-Z. Could plant lectins become promising anti-tumour drugs for causing autophagic cell death? Cell Prolif. 2013, 46, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.R.D.; Mota, C.M.; Ribeiro, D.P.; Noleto, P.G.; Andrade, W.B.F.; Souza, M.A.; Silva, N.M.; Mineo, T.W.P.; Mineo, J.R.; Silva, D.A.O. Adjuvant and immunostimulatory effects of a D-galactose-binding lectin from Synadenium carinatum latex (ScLL) in the mouse model of vaccination against neosporosis. Vet. Res. 2012, 43, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.-H.; Kong, J.; Li, W.; Molchanova, V.; Chikalovets, I.; Belogortseva, N.; Luk’yanov, P.; Zheng, Y.-T. A beta-galactose-specific lectin isolated from the marine worm Chaetopterus variopedatus possesses anti-HIV-1 activity. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 142, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Allen, H.J.; Kisailus, E.C. Glycoconjugates: Composition, Structure, and Function; Dekker: New York, NY, USA, 1992; ISBN 9780824784317. [Google Scholar]
- Chatterjee, B.P.; Adhya, M. Lectins with Varying Specificity and Biological Activity from Marine Bivalves. In Marine Proteins and Peptides; John Wiley & Sons, Ltd: Chichester, UK, 2013; pp. 41–68. ISBN 9781118375082. [Google Scholar]
- Adhya, M.; Singha, B. Gal/GalNAc specific multiple lectins in marine bivalve Anadara granosa. Fish Shellfish Immunol. 2016, 50, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Belogortseva, N.I.; Molchanova, V.I.; Kurika, A.V.; Skobun, A.S.; Glazkova, V.E. Isolation and characterization of new GalNAc/Gal-specific lectin from the sea mussel Crenomytilus grayanus. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1998, 119, 45–50. [Google Scholar] [CrossRef]
- Chikalovets, I.V.; Kovalchuk, S.N.; Litovchenko, A.P.; Molchanova, V.I.; Pivkin, M.V.; Chernikov, O.V. A new Gal/GalNAc-specific lectin from the mussel Mytilus trossulus: Structure, tissue specificity, antimicrobial and antifungal activity. Fish Shellfish Immunol. 2016, 50, 27–33. [Google Scholar] [CrossRef]
- Fujii, Y.; Dohmae, N.; Takio, K.; Kawsar, S.M.; Matsumoto, R.; Hasan, I.; Koide, Y.; Kanaly, R.A.; Yasumitsu, H.; Ogawa, Y.; et al. A lectin from the mussel Mytilus galloprovincialis has a highly novel primary structure and induces glycan-mediated cytotoxicity of globotriaosylceramide-expressing lymphoma cells. J. Biol. Chem. 2012, 287, 44772–44783. [Google Scholar] [CrossRef] [Green Version]
- Chikalovets, I.V.; Mizgina, T.O.; Molchanova, V.I.; Ovcharenko, Y.S.; Chernikov, O.V. Isolation and Characterization of Lectin from the Scallop Patinopecten yessoensis. Chem. Nat. Compd. 2017, 53, 717–721. [Google Scholar] [CrossRef]
- Hasan, I.; Gerdol, M.; Fujii, Y.; Rajia, S.; Koide, Y.; Yamamoto, D.; Kawsar, S.M.A.; Ozeki, Y. CDNA and Gene Structure of MytiLec-1, A Bacteriostatic R-Type Lectin from the Mediterranean Mussel (Mytilus galloprovincialis). Mar. Drugs 2016, 14, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovalchuk, S.N.; Chikalovets, I.V.; Chernikov, O.V.; Molchanova, V.I.; Li, W.; Rasskazov, V.A.; Lukyanov, P.A. CDNA cloning and structural characterization of a lectin from the mussel Crenomytilus grayanus with a unique amino acid sequence and antibacterial activity. Fish Shellfish Immunol. 2013, 35, 1320–1324. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Gerdol, M.; Hasan, I.; Koide, Y.; Matsuzaki, R.; Ikeda, M.; Rajia, S.; Ogawa, Y.; Kawsar, S.M.A.; Ozeki, Y. Phylogeny and Properties of a Novel Lectin Family with β-Trefoil Folding in Mussels. Trends Glycosci. Glycotechnol. 2018, 30, J155–J168. [Google Scholar] [CrossRef] [Green Version]
- Terada, D.; Kawai, F.; Noguchi, H.; Unzai, S.; Hasan, I.; Fujii, Y.; Park, S.-Y.; Ozeki, Y.; Tame, J.R.H. Crystal structure of MytiLec, a galactose-binding lectin from the mussel Mytilus galloprovincialis with cytotoxicity against certain cancer cell types. Sci. Rep. 2016, 6, 28344. [Google Scholar] [CrossRef]
- Liao, J.H.; Chien, C.T.; Wu, H.Y.; Huang, K.F.; Wang, I.; Ho, M.R.; Tu, I.F.; Lee, I.M.; Li, W.; Shih, Y.L.; et al. A Multivalent Marine Lectin from Crenomytilus grayanus Possesses Anti-cancer Activity through Recognizing Globotriose Gb3. J. Am. Chem. Soc. 2016, 138, 4787–4795. [Google Scholar] [CrossRef]
- Gerdol, M.; Venier, P. An updated molecular basis for mussel immunity. Fish Shellfish Immunol. 2015, 46, 17–38. [Google Scholar] [CrossRef] [Green Version]
- Chikalovets, I.V.; Kondrashina, A.S.; Chernikov, O.V.; Molchanova, V.I.; Luk’yanov, P.A. Isolation and general characteristics of lectin from the mussel Mytilus trossulus. Chem. Nat. Compd. 2013, 48, 1058–1061. [Google Scholar] [CrossRef]
- Chichvarkhin, A.I.; Kartavtsev, I.F.; Kafanov, A.I. Genetic connections between some species of Mytilidae (Mollusca: Bivalvia) from the northern part of the Pacific Ocean. Genetika 2000, 36, 1206–1220. [Google Scholar]
- Houser, J.; Komárek, J.; Kostlánová, N.; Cioci, G.; Imberty, A.; Wimmerová, M. Protein oligomerization in Aleuria aurantia lectin family—Importance and difficulties. Mater. Struct. Chem. Biol. Phys. Technol. 2012, 19, 20–21. [Google Scholar]
- Chikalovets, I.V.; Molchanova, V.I.; Chernikov, O.V.; Luk’Yanov, P.A. Domain organization of lectin from the mussel Crenomytilus grayanus. Chem. Nat. Compd. 2014, 50, 706–709. [Google Scholar] [CrossRef]
- Cho, M.; Cummings, R.D. Galectin-1, a beta-galactoside-binding lectin in Chinese hamster ovary cells. I. Physical and chemical characterization. J. Biol. Chem. 1995, 270, 5198–5206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terada, D.; Voet, A.R.D.; Noguchi, H.; Kamata, K.; Ohki, M.; Addy, C.; Fujii, Y.; Yamamoto, D.; Ozeki, Y.; Tame, J.R.H.; et al. Computational design of a symmetrical β-trefoil lectin with cancer cell binding activity. Sci. Rep. 2017, 7, 5943. [Google Scholar] [CrossRef] [PubMed]
- Chernikov, O.; Kuzmich, A.; Chikalovets, I.; Molchanova, V.; Hua, K.-F. Lectin CGL from the sea mussel Crenomytilus grayanus induces Burkitt’s lymphoma cells death via interaction with surface glycan. Int. J. Biol. Macromol. 2017, 104, 508–514. [Google Scholar] [CrossRef]
- Oyelaran, O.; Gildersleeve, J.C. Glycan arrays: Recent advances and future challenges. Curr. Opin. Chem. Biol. 2009, 13, 406–413. [Google Scholar] [CrossRef] [Green Version]
- Cheung, R.C.; Wong, J.H.; Pan, W.; Chan, Y.S.; Yin, C.; Dan, X.; Ng, T.B. Marine lectins and their medicinal applications. Appl. Microbiol. Biotechnol. 2015, 99, 3755–3773. [Google Scholar] [CrossRef]
- Toubiana, M.; Gerdol, M.; Rosani, U.; Pallavicini, A.; Venier, P.; Roch, P. Toll-like receptors and MyD88 adaptors in Mytilus: Complete cds and gene expression levels. Dev. Comp. Immunol. 2013, 40, 158–166. [Google Scholar] [CrossRef]
- Kubler-Kielb, J.; Lai, W.-T.; Schneerson, R.; Vinogradov, E. The structure of the Escherichia coli O148 lipopolysaccharide core region and its linkage to the O-specific polysaccharide. Carbohydr. Res. 2011, 346, 150–152. [Google Scholar] [CrossRef] [Green Version]
- Poxton, I.R. Teichoic Acids, Lipoteichoic Acids and Other Secondary Cell Wall and Membrane Polysaccharides of Gram-Positive Bacteria. In Molecular Medical Microbiology; Academic Press: Cambridge, MA, USA, 2015; pp. 91–103. ISBN 9780123971692. [Google Scholar]
- Vollmer, W. Peptidoglycan. In Molecular Medical Microbiology; Academic Press: Cambridge, MA, USA, 2015; pp. 105–124. ISBN 9780123971692. [Google Scholar]
- Gaudet, R.G.; Sintsova, A.; Buckwalter, C.M.; Leung, N.; Cochrane, A.; Li, J.; Cox, A.D.; Moffat, J.; Gray-Owen, S.D. Cytosolic detection of the bacterial metabolite HBP activates TIFA-dependent innate immunity. Science 2015, 348, 1251–1255. [Google Scholar] [CrossRef]
- Wollin, R.; Creeger, E.S.; Rothfield, L.I.; Stocker, B.A.; Lindberg, A.A. Salmonella typhimurium mutants defective in UDP-D-galactose:lipopolysaccharide alpha 1,6-D-galactosyltransferase. Structural, immunochemical, and enzymologic studies of rfaB mutants. J. Biol. Chem. 1983, 258, 3769–3774. [Google Scholar]
- Klein, R.A.; Hartmann, R.; Egge, H.; Behr, T.; Fischer, W. The aqueous solution structure of a lipoteichoic acid from Streptococcus pneumoniae strain R6 containing 2,4-diamino-2,4,6-trideoxy-galactose: Evidence for conformational mobility of the galactopyranose ring. Carbohydr. Res. 1996, 281, 79–98. [Google Scholar] [CrossRef]
- Valiente, E.; Jiménez, N.; Merino, S.; Tomás, J.M.; Amaro, C. Vibrio vulnificus biotype 2 serovar E gne but not galE is essential for lipopolysaccharide biosynthesis and virulence. Infect. Immun. 2008, 76, 1628–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado-Aguayo, W.; Teneb, J.; Gallardo-Escárate, C. A galectin with quadruple-domain from red abalone Haliotis rufescens involved in the immune innate response against to Vibrio anguillarum. Fish Shellfish Immunol. 2014, 40, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Shen, H.; Zhou, H.; Dong, Y.; Lin, Z. A tandem-repeat galectin from blood clam Tegillarca granosa and its induced mRNA expression response against bacterial challenge. Genes Genom. 2013, 35, 733–740. [Google Scholar] [CrossRef]
- Moreira, R.; Balseiro, P.; Romero, A.; Dios, S.; Posada, D.; Novoa, B.; Figueras, A. Gene expression analysis of clams Ruditapes philippinarum and Ruditapes decussatus following bacterial infection yields molecular insights into pathogen resistance and immunity. Dev. Comp. Immunol. 2012, 36, 140–149. [Google Scholar] [CrossRef] [Green Version]
- Chikalovets, I.V.; Chernikov, O.V.; Pivkin, M.V.; Molchanova, V.I.; Litovchenko, A.P.; Li, W.; Lukyanov, P.A. A lectin with antifungal activity from the mussel Crenomytilus grayanus. Fish Shellfish Immunol. 2015, 42, 503–507. [Google Scholar] [CrossRef]
- Van Parijs, J.; Joosen, H.M.; Peumans, W.J.; Geuns, J.M.; Van Laere, A.J. Effect of the Urtica dioica agglutinin on germination and cell wall formation of Phycomyces blakesleeanus Burgeff. Arch. Microbiol. 1992, 158, 19–25. [Google Scholar] [CrossRef]
- Zvereva, L.V.; Vysotskaya, M.A. Filamentous Fungi Associated with Bivalve Mollusks from Polluted Biotopes of Ussuriiskii Bay, Sea of Japan. Russ. J. Mar. Biol. 2005, 31, 382–385. [Google Scholar] [CrossRef]
- Kang, H.K.; Lee, H.H.; Seo, C.H.; Park, Y. Antimicrobial and immunomodulatory properties and applications of marine-derived proteins and peptides. Mar. Drugs 2019, 17, 350. [Google Scholar] [CrossRef] [Green Version]
- Malagoli, D.; Sacchi, S.; Ottaviani, E. Lectins and cytokines in celomatic invertebrates: Two tales with the same end. Invertebr. Surviv. J. 2010, 7, 1–10. [Google Scholar]
- Hosono, M.; Sugawara, S.; Matsuda, A.; Tatsuta, T.; Koide, Y.; Hasan, I.; Ozeki, Y.; Nitta, K. Binding profiles and cytokine-inducing effects of fish rhamnose-binding lectins on Burkitt’s lymphoma Raji cells. Fish Physiol. Biochem. 2014, 40, 1559–1572. [Google Scholar] [CrossRef] [PubMed]
- Yamanishi, T.; Yamamoto, Y.; Hatakeyama, T.; Yamaguchi, K.; Oda, T. CEL-I, an invertebrate N-acetylgalactosamine-specific C-type lectin, induces TNF-alpha and G-CSF production by mouse macrophage cell line RAW264.7 cells. J. Biochem. 2007, 142, 587–595. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.H.; Mun, S.H.; Ko, N.Y.; Lee, J.H.; Jun, M.H.; Seo, J.Y.; Kim, Y.M.; Choi, W.S.; Her, E. The synergistic effect of phytohemagglutinin and interferon-gamma on the expression of tumor necrosis factor-alpha from RAW 264.7 cells. Immunol. Lett. 2005, 98, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Chernikov, O.V.; Wong, W.-T.; Li, L.-H.; Chikalovets, I.V.; Molchanova, V.I.; Wu, S.-H.; Liao, J.-H.; Hua, K.-F. A GalNAc/Gal-specific lectin from the sea mussel Crenomytilus grayanus modulates immune response in macrophages and in mice. Sci. Rep. 2017, 7, 6315. [Google Scholar] [CrossRef] [PubMed]
- Lambeth, J.D. NOX enzymes and the biology of reactive oxygen. Nat. Rev. Immunol. 2004, 4, 181–189. [Google Scholar] [CrossRef]
- West, A.P.; Brodsky, I.E.; Rahner, C.; Woo, D.K.; Erdjument-Bromage, H.; Tempst, P.; Walsh, M.C.; Choi, Y.; Shadel, G.S.; Ghosh, S. TLR signalling augments macrophage bactericidal activity through mitochondrial ROS. Nature 2011, 472, 476–480. [Google Scholar] [CrossRef] [Green Version]
- Fan, H.; Cook, J.A. Molecular mechanisms of endotoxin tolerance. J. Endotoxin Res. 2004, 10, 71–84. [Google Scholar] [CrossRef]
- Wheeler, D.S.; Lahni, P.M.; Denenberg, A.G.; Poynter, S.E.; Wong, H.R.; Cook, J.A.; Zingarelli, B. Induction of endotoxin tolerance enhances bacterial clearance and survival in murine polymicrobial sepsis. Shock 2008, 30, 267–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furtak, V.A.; Kurika, A.V.; Belogortseva, N.I.; Chikalovets, I.V.; Kleshch, Y. Cell localization of mucin-type receptors assayed with novel GalNac/Gal-specific lectin from sea mussel Crenomytilus grayanus in human colon tumors. Bull. Exp. Biol. Med. 1999, 128, 1039–1041. [Google Scholar] [CrossRef]
- Nudelman, E.; Kannagi, R.; Hakomori, S.; Parsons, M.; Lipinski, M.; Wiels, J.; Fellous, M.; Tursz, T. A glycolipid antigen associated with Burkitt lymphoma defined by a monoclonal antibody. Science 1983, 220, 509–511. [Google Scholar] [CrossRef]
- Cao, Y.; Merling, A.; Karsten, U.; Goletz, S.; Punzel, M.; Kraft, R.; Butschak, G.; Schwartz-Albiez, R. Expression of CD175 (Tn), CD175s (sialosyl-Tn) and CD176 (Thomsen-Friedenreich antigen) on malignant human hematopoietic cells. Int. J. Cancer 2008, 123, 89–99. [Google Scholar] [CrossRef] [PubMed]
- Geiger, P.; Mayer, B.; Wiest, I.; Schulze, S.; Jeschke, U.; Weissenbacher, T. Binding of galectin-1 to breast cancer cells MCF7 induces apoptosis and inhibition of proliferation in vitro in a 2D- and 3D- cell culture model. BMC Cancer 2016, 16, 870. [Google Scholar] [CrossRef] [Green Version]
- Freire, T.; Bay, S.; von Mensdorff-Pouilly, S.; Osinaga, E. Molecular Basis of Incomplete O-Glycan Synthesis in MCF-7 Breast Cancer Cells: Putative Role of MUC6 in Tn Antigen Expression. Cancer Res. 2005, 65, 7880–7887. [Google Scholar] [CrossRef] [Green Version]
- Hasan, I.; Sugawara, S.; Fujii, Y.; Koide, Y.; Terada, D.; Iimura, N.; Fujiwara, T.; Takahashi, K.G.; Kojima, N.; Rajia, S.; et al. MytiLec, a Mussel R-Type Lectin, Interacts with Surface Glycan Gb3 on Burkitt’s Lymphoma Cells to Trigger Apoptosis through Multiple Pathways. Mar. Drugs 2015, 13, 7377–7389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, H.Y.; Yang, X. Proteases for cell suicide: Functions and regulation of caspases. Microbiol. Mol. Biol. Rev. 2000, 64, 821–846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stennicke, H.R.; Salvesen, G.S. Properties of the caspases. Biochim. Biophys. Acta 1998, 1387, 17–31. [Google Scholar] [CrossRef]
- Dias, N.; Bailly, C. Drugs targeting mitochondrial functions to control tumor cell growth. Biochem. Pharmacol. 2005, 70, 1–12. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lectin | MW (Da) | Thermal Stability | pH Dependence | Metal Ion Dependence | Localization in Mussel Tissue | Reference |
|---|---|---|---|---|---|---|
| CGL | 16,953 | Stable until 50 °C | 8–10 | Not dependent | Mantle | [10,15] |
| MTL | 16,492 | Stable until 50 °C | 9–10 | Not dependent | Mantle | [11,21] |
| MytiLec-1 | 16,812 | N.D. 1 | N.D. | Not dependent | Mantle | [12] |
| Carbohydrate | Minimum Inhibitory Concentration, mM | ||
|---|---|---|---|
| CGL [10] | MTL [21] | MytiLec-1 [12] | |
| N-Acetyl-d-galactosamine | 1.4 | 0.7 | 1.6 |
| N-Acetyl-d-glucosamine | No inhibition at 80 | >50 | No inhibition at 100 |
| d-Galactose | 5.4 | 1.7 | 3.1 |
| d-Glucose | No inhibition at 80 | >50 | >50 |
| d-Fucose | No inhibition at 80 | >50 | >50 |
| d-Talose | 5.4 | N.D. 1 | 25 |
| Lactose | No inhibition at 80 | 29.2 | 50 |
| Melibiose | 2.0 | N.D. | 1.6 |
| Raffinose | 1.8 | N.D. | N.D. |
| Glycoproteins | Minimum Inhibitory Concentration, mg/mL | ||
| BSM | 0.007 | 0.0156 | No inhibition at 2 |
| Asialo-BSM | 0.0017 | N.D. | 0.2 |
| Fetuin | 2 | 0.0156 | No inhibition at 2 |
| Asialofetuin | 0.03 | 0.0156 | 0.2 |
| Glycan # | Compound | Fluorescence (%) 1 | |
|---|---|---|---|
| CGL [27] | MTL 2 | ||
| 559 | Galα1-3Galβ1-4GlcNAcβ1-2Mana1-6(Galα1-3Galβ1-4GlcNAcβ1-2Manα1-3)Manβ1-4GlcNAcβ1-4GlcNAc-Sp24 | 100 | 87.7 |
| 587 | Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-6(Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-2)Manα1-6(Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-2Manα1-3)Manβ1-4GlcNAcβ1-4(Fucα1-6)GlcNAcβ-Sp24 | 98.7 | 100 |
| 362 | Galα1-3Galβ1-4GlcNAcβ1-2Manα1-6(Galα1-3Galβ1-4GlcNAcβ1-2Manα1-3)Manβ1-4GlcNAcβ1-4GlcNAcβ-Sp20 | 96.4 | 91.5 |
| 402 | Galα1-4Galβ1-3GlcNAcβ1-2Manα1-6(Galα1-4Galβ1-3GlcNAcβ1-2Manα1-3)Manβ1-4GlcNAcβ1-4GlcNAcβ-Sp19 | 78.2 | 72.5 |
| 583 | Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-6(Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-2)Manα1-6(Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-3Galβ1-4GlcNAcβ1-2Manα1-3)Manβ1-4GlcNAcβ1-4(Fucα1-6)GlcNAcβ-Sp24 | 66.5 | 72.4 |
| 122 | Galα1-4Galβ1-4Glcβ-Sp0 | 24.5 | 23.3 |
| 72 | Fucα1-2Galβ1-4(Fucα1-3)GlcNAcβ-Sp8 | 0.15 | 0.03 |
| Bacteria | Binding Activity 1 | Agglutination 2 | Growth Suppressive Activity (%) | |||||
|---|---|---|---|---|---|---|---|---|
| CGL | MTL | CGL | MTL | MytiLec-1 | CGL | MTL | MytiLec-1 | |
| Candida albicans | 0.65 ± 0.01 | 0.44 ± 0.05 | ++ | +++ | N.D. 3 | - 4 | - | N.D. |
| Vibrio proteolyticus | 1.42 ± 0.04 | 0.37 ± 0.14 | +++ | ++ | N.D. | 39.9 ± 5 | - | N.D. |
| Escherichia coli | 1.63 ± 0.09 | 0.48 ± 0.06 | ++ | + | ++ | 46 ± 5 | 10.6 ± 7 | 58 ± 5 |
| Bacillus subtilis | 0.74 ± 0.07 | 0.26 ± 0.07 | +++ | ++ | ++ | 85 ± 8 | 62 ± 6 | 74 ± 8 |
| Staphylococcus aureus | 0.39 ± 0.06 | 0.49 ± 0.01 | ++ | ++ | + | 68 ± 6 | - | 61 ± 36 |
| CGL (μg/mL) | CFU (×106) | Killed Bacteria (×106) | |
|---|---|---|---|
| 1 h after Infection | 24 h after Infection | ||
| 0 | 1680 ± 370 | 724 ± 196 | 956 |
| 10 | 1984 ± 792 | 448 ± 166 | 1536 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chikalovets, I.; Filshtein, A.; Molchanova, V.; Mizgina, T.; Lukyanov, P.; Nedashkovskaya, O.; Hua, K.-F.; Chernikov, O. Activity Dependence of a Novel Lectin Family on Structure and Carbohydrate-Binding Properties. Molecules 2020, 25, 150. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010150
Chikalovets I, Filshtein A, Molchanova V, Mizgina T, Lukyanov P, Nedashkovskaya O, Hua K-F, Chernikov O. Activity Dependence of a Novel Lectin Family on Structure and Carbohydrate-Binding Properties. Molecules. 2020; 25(1):150. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010150
Chicago/Turabian StyleChikalovets, Irina, Alina Filshtein, Valentina Molchanova, Tatyana Mizgina, Pavel Lukyanov, Olga Nedashkovskaya, Kuo-Feng Hua, and Oleg Chernikov. 2020. "Activity Dependence of a Novel Lectin Family on Structure and Carbohydrate-Binding Properties" Molecules 25, no. 1: 150. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25010150