
Synthesis and Immunological Evaluation of a Single Molecular Construct MUC1 Vaccine Containing l-Rhamnose Repeating Units


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
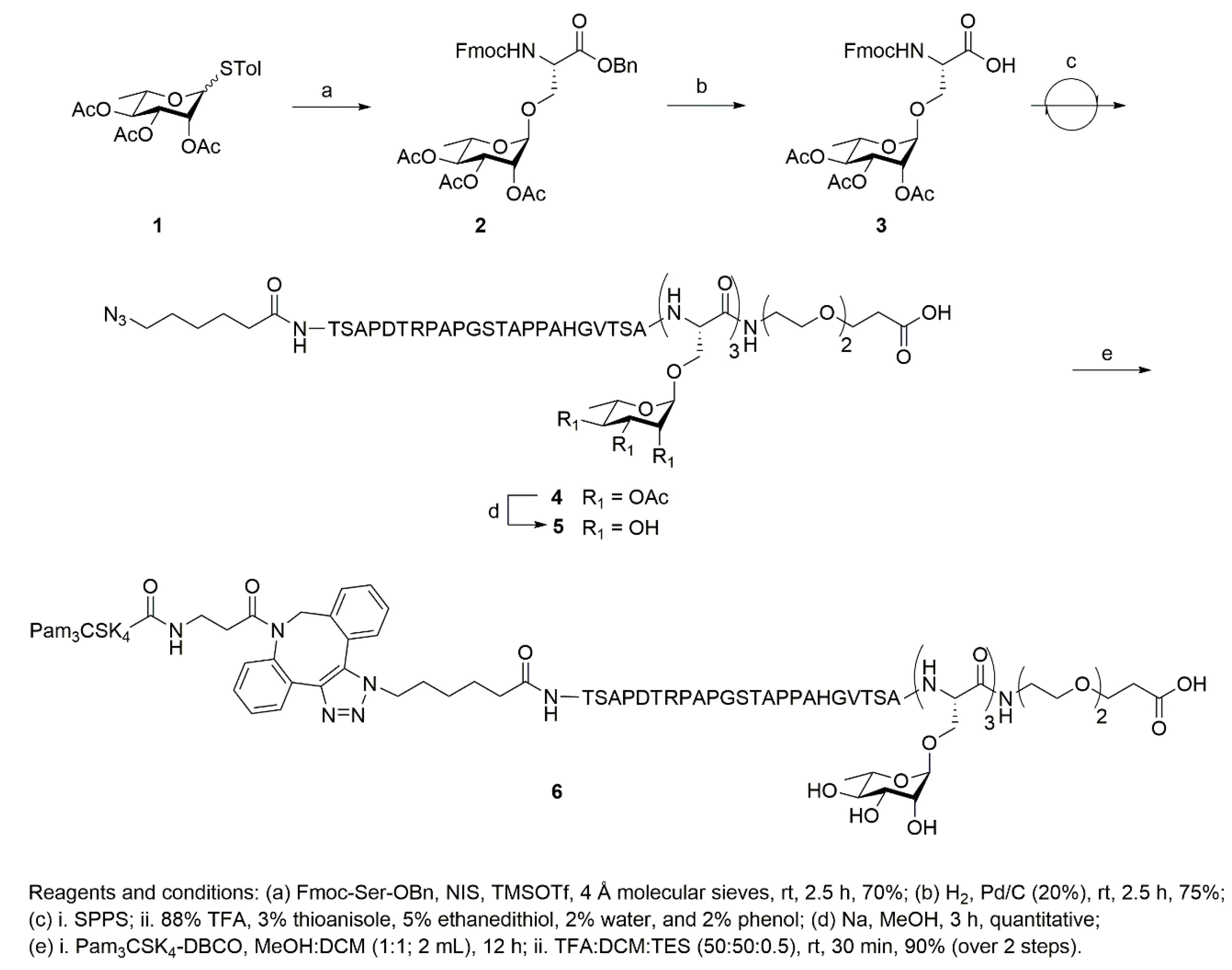
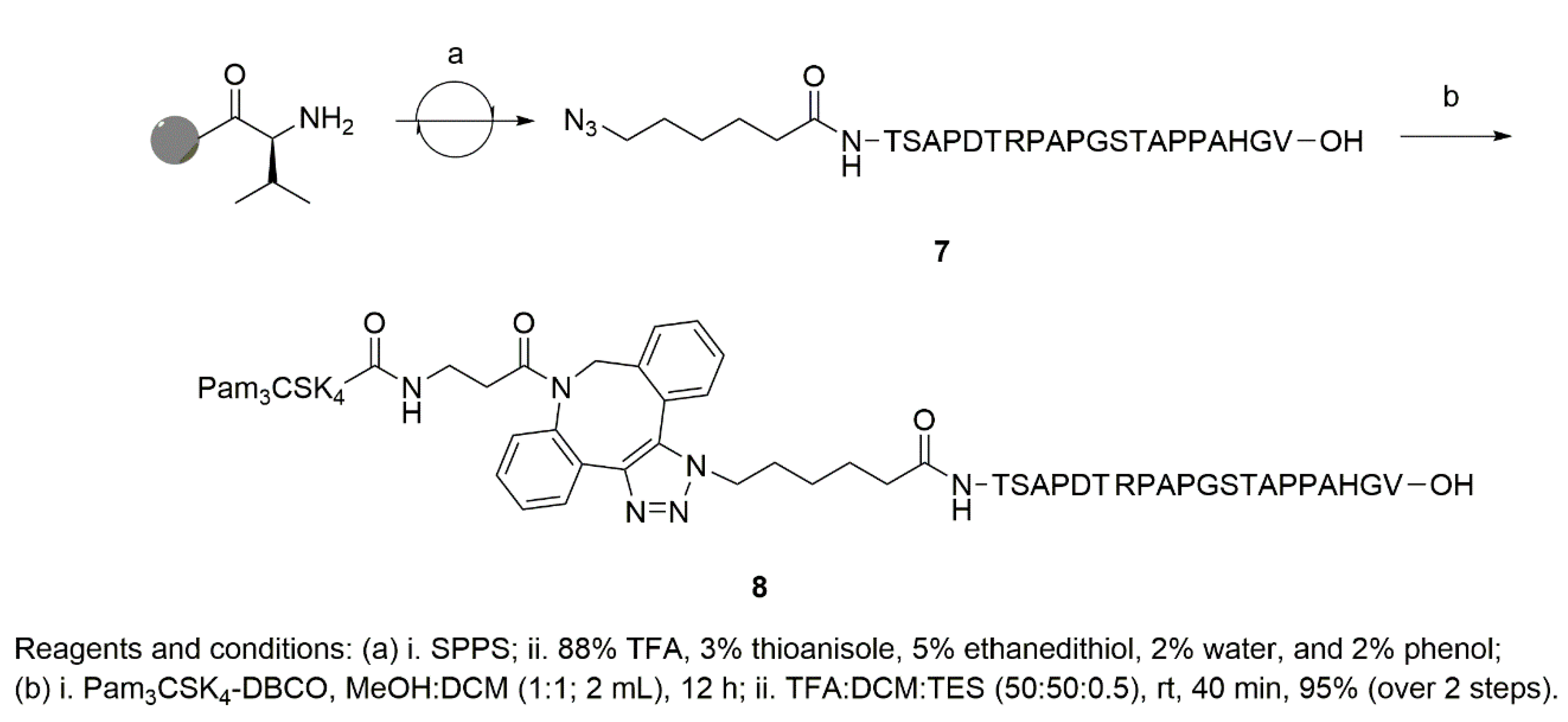
2. Results and Discussion
3. Materials and Methods
3.1. Mouse Strain and Immunization
3.2. ELISA Assay
3.3. Proliferation Assay
3.4. Synthesis of Pam3CSK4-DBCO-MUC1
3.4.1. Synthesis of N-[(9H-fluoren-9-yl)methoxycarbonyl]-O-((2,3,4-tri-O-acetyl)-α-l-rhamnopyranosyl)-l-serine Phenylmethyl Ester (Fmoc-l-Ser(Ac3-α-Rhamose)-OBn) (2)
3.4.2. Synthesis of N-[(9H-fluoren-9-yl)methoxycarbonyl]-O-((2,3,4-tri-O-acetyl)-α-l-rhamnopyranosyl)-l-serine (Fmoc-l-Ser(Ac3-α-Rhamose)-OH) (3)
3.4.3. Synthesis of Azide-Terminated Peptide Sequences, (4) and (7)
3.4.4. Synthesis of the Single Molecule Construct of Pam3CSK4-DBCO-MUC1-(Rha)3 (6)
3.4.5. Synthesis of Pam3CSK4-DBCO-MUC1 (8)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nath, S.; Mukherjee, P. MUC1: A multifaceted oncoprotein with a key role in cancer progression. Trends. Mol. Med. 2014, 20, 332–342. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.K.; Wall, K.A. Immunological Evaluation of Recent MUC1 Glycopeptide Cancer Vaccines. Vaccines 2016, 4, 25. [Google Scholar] [CrossRef]
- Apostolopoulos, V.; Stojanovska, L.; Gargosky, S.E. MUC1 (CD227): A multi-tasked molecule. Cell Mol. Life Sci. 2015, 72, 4475–4500. [Google Scholar] [CrossRef] [PubMed]
- Hanisch, F.-G.; Müller, S. MUC1: The polymorphic appearance of a human mucin. Glycobiology 2000, 10, 439–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor-Papadimitriou, J.; Gendler, S. Molecular aspects of mucins. Cancer Rev. 1988, 11, 11–24. [Google Scholar]
- Lakshminarayanan, V.; Supekar, N.T.; Wei, J.; McCurry, D.B.; Dueck, A.C.; Kosiorek, H.E.; Trivedi, P.P.; Bradley, J.M.; Madsen, C.S.; Pathangey, L.B.; et al. MUC1 Vaccines, Comprised of Glycosylated or Non-Glycosylated Peptides or Tumor-Derived MUC1, Can Circumvent Immunoediting to Control Tumor Growth in MUC1 Transgenic Mice. PLoS ONE 2016, 11, e0145920. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Degliangeli, F.; Palitzsch, B.; Gerlitzki, B.; Kunz, H.; Schmitt, E.; Fiammengo, R.; Westerlind, U. Glycopeptide-functionalized gold nanoparticles for antibody induction against the tumor associated mucin-1 glycoprotein. Bioorg. Med. Chem. 2016, 24, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Qiu, L.; Yan, Z.; Lin, Z.; Cao, M.; Hu, C.; Wang, Z.; Wang, J.; Yu, Y.; Cheng, X.; et al. Cytotoxic T cell responses are enhanced by antigen design involving the presentation of MUC1 peptide on cholera toxin B subunit. Oncotarget 2015, 6, 34537–34548. [Google Scholar] [CrossRef] [Green Version]
- Glaffig, M.; Palitzsch, B.; Stergiou, N.; Schull, C.; Strassburger, D.; Schmitt, E.; Frey, H.; Kunz, H. Enhanced immunogenicity of multivalent MUC1 glycopeptide antitumour vaccines based on hyperbranched polymers. Org. Biomol. Chem. 2015, 13, 10150–10154. [Google Scholar] [CrossRef] [Green Version]
- Thompson, P.; Lakshminarayanan, V.; Supekar, N.T.; Bradley, J.M.; Cohen, P.A.; Wolfert, M.A.; Gendler, S.J.; Boons, G.J. Linear synthesis and immunological properties of a fully synthetic vaccine candidate containing a sialylated MUC1 glycopeptide. Chem. Commun. 2015, 51, 10214–10217. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, C.; Xia, Q.; Wang, P.; Li, B.; Lu, Z.; Sun, J.; Wu, H.; Yu, B.; Wu, J.; et al. Antitumor effect of adenoviral vector prime protein boost immunity targeting the MUC1 VNTRs. Oncol. Rep. 2014, 31, 1437–1444. [Google Scholar] [CrossRef]
- Sarkar, S.; Salyer, A.C.; Wall, K.A.; Sucheck, S.J. Synthesis and immunological evaluation of a MUC1 glycopeptide incorporated into l-rhamnose displaying liposomes. Bioconjug. Chem. 2013, 24, 363–375. [Google Scholar] [CrossRef] [Green Version]
- Lakshminarayanan, V.; Thompson, P.; Wolfert, M.A.; Buskas, T.; Bradley, J.M.; Pathangey, L.B.; Madsen, C.S.; Cohen, P.A.; Gendler, S.J.; Boons, G.J. Immune recognition of tumor-associated mucin MUC1 is achieved by a fully synthetic aberrantly glycosylated MUC1 tripartite vaccine. Proc. Natl. Acad. Sci. USA 2012, 109, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Karmakar, P.; Lee, K.; Sarkar, S.; Wall, K.A.; Sucheck, S.J. Synthesis of a Liposomal MUC1 Glycopeptide-Based Immunotherapeutic and Evaluation of the Effect of l-Rhamnose Targeting on Cellular Immune Responses. Bioconjug. Chem. 2016, 27, 110–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.K.; Vartak, A.; Karmakar, P.; Sucheck, S.J.; Wall, K.A. Augmenting Vaccine Immunogenicity through the Use of Natural Human Anti-rhamnose Antibodies. ACS Chem. Biol. 2018, 13, 2130–2142. [Google Scholar] [CrossRef] [PubMed]
- Villadangos, J.A.; Cardoso, M.; Steptoe, R.J.; van Berkel, D.; Pooley, J.; Carbone, F.R.; Shortman, K. MHC class II expression is regulated in dendritic cells independently of invariant chain degradation. Immunity 2001, 14, 739–749. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.K.; Wall, K.A. Use of Dendritic Cell Receptors as Targets for Enhancing Anti-Cancer Immune Responses. Cancers 2019, 11, 418. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez-Martinez, E.; Planes, R.; Anselmi, G.; Reynolds, M.; Menezes, S.; Adiko, A.C.; Saveanu, L.; Guermonprez, P. Cross-Presentation of Cell-Associated Antigens by MHC Class I in Dendritic Cell Subsets. Front. Immunol. 2015, 6, 363. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, S.; Lombardo, S.A.; Herner, D.N.; Talan, R.S.; Wall, K.A.; Sucheck, S.J. Synthesis of a single-molecule L-rhamnose-containing three-component vaccine and evaluation of antigenicity in the presence of anti-L-rhamnose antibodies. J. Am. Chem. Soc. 2010, 132, 17236–17246. [Google Scholar] [CrossRef]
- Sheridan, R.T.; Hudon, J.; Hank, J.A.; Sondel, P.M.; Kiessling, L.L. Rhamnose glycoconjugates for the recruitment of endogenous anti-carbohydrate antibodies to tumor cells. Chembiochem 2014, 15, 1393–1398. [Google Scholar] [CrossRef] [Green Version]
- Oyelaran, O.; McShane, L.M.; Dodd, L.; Gildersleeve, J.C. Profiling human serum antibodies with a carbohydrate antigen microarray. J. Proteome. Res. 2009, 8, 4301–4310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.K.; Vartak, A.; Sucheck, S.J.; Wall, K.A. Liposomal Fc Domain Conjugated to a Cancer Vaccine Enhances Both Humoral and Cellular Immunity. ACS Omega 2019, 4, 5204–5208. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.A.; Felix, A.M. Pegylated peptides II: Solid-phase synthesis of amino-, carboxy-and side-chain pegylated peptides. Int. J. Pept. Protein Res. 1994, 43, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Veeneman, G.; Van Leeuwen, S.; Van Boom, J. Iodonium ion promoted reactions at the anomeric centre. II An efficient thioglycoside mediated approach toward the formation of 1, 2-trans linked glycosides and glycosidic esters. Tetrahedron Lett. 1990, 31, 1331–1334. [Google Scholar] [CrossRef]
- Fügedi, P.; Garegg, P.J.; Lönn, H.; Norberg, T. Thioglycosides as glycosylating agents in oligosaccharide synthesis. Glycoconj. J. 1987, 4, 97–108. [Google Scholar] [CrossRef]
- Greene, T.W.; Wuts, P.G. Protective Groups in Organic Synthesis. Wiley: New York, NY, USA, 1999. [Google Scholar]
- Nandedkar-Kulkarni, N.; Vartak, A.R.; Sucheck, S.J.; Wall, K.A.; Quinn, A.; Morran, M.P.; McInerney, M.F. Development of a bioconjugate platform for modifying the immune response of autoreactive cytotoxic T lymphocytes involved in type 1 diabetes. Bioconjugate Chem. 2019, 30, 2049–2059. [Google Scholar] [CrossRef]
- Kaiser, A.; Gaidzik, N.; Becker, T.; Menge, C.; Groh, K.; Cai, H.; Li, Y.M.; Gerlitzki, B.; Schmitt, E.; Kunz, H. Fully synthetic vaccines consisting of tumor-associated MUC1 glycopeptides and a lipopeptide ligand of the toll-like receptor 2. Angew. Chem. Int. Ed. 2010, 49, 3688–3692. [Google Scholar] [CrossRef]
- Matheu, M.P.; Sen, D.; Cahalan, M.D.; Parker, I. Generation of bone marrow derived murine dendritic cells for use in 2-photon imaging. JoVE (J. Vis. Exp.) 2008, e773. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are in limited quantity and are not generally available. |




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hossain, M.K.; Vartak, A.; Sucheck, S.J.; Wall, K.A. Synthesis and Immunological Evaluation of a Single Molecular Construct MUC1 Vaccine Containing l-Rhamnose Repeating Units. Molecules 2020, 25, 3137. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25143137
Hossain MK, Vartak A, Sucheck SJ, Wall KA. Synthesis and Immunological Evaluation of a Single Molecular Construct MUC1 Vaccine Containing l-Rhamnose Repeating Units. Molecules. 2020; 25(14):3137. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25143137
Chicago/Turabian StyleHossain, Md Kamal, Abhishek Vartak, Steven J. Sucheck, and Katherine A. Wall. 2020. "Synthesis and Immunological Evaluation of a Single Molecular Construct MUC1 Vaccine Containing l-Rhamnose Repeating Units" Molecules 25, no. 14: 3137. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25143137