
Evaluating the Effect of Azole Antifungal Agents on the Stress Response and Nanomechanical Surface Properties of Ochrobactrum anthropi Aspcl2.2


Abstract
:1. Introduction
2. Results
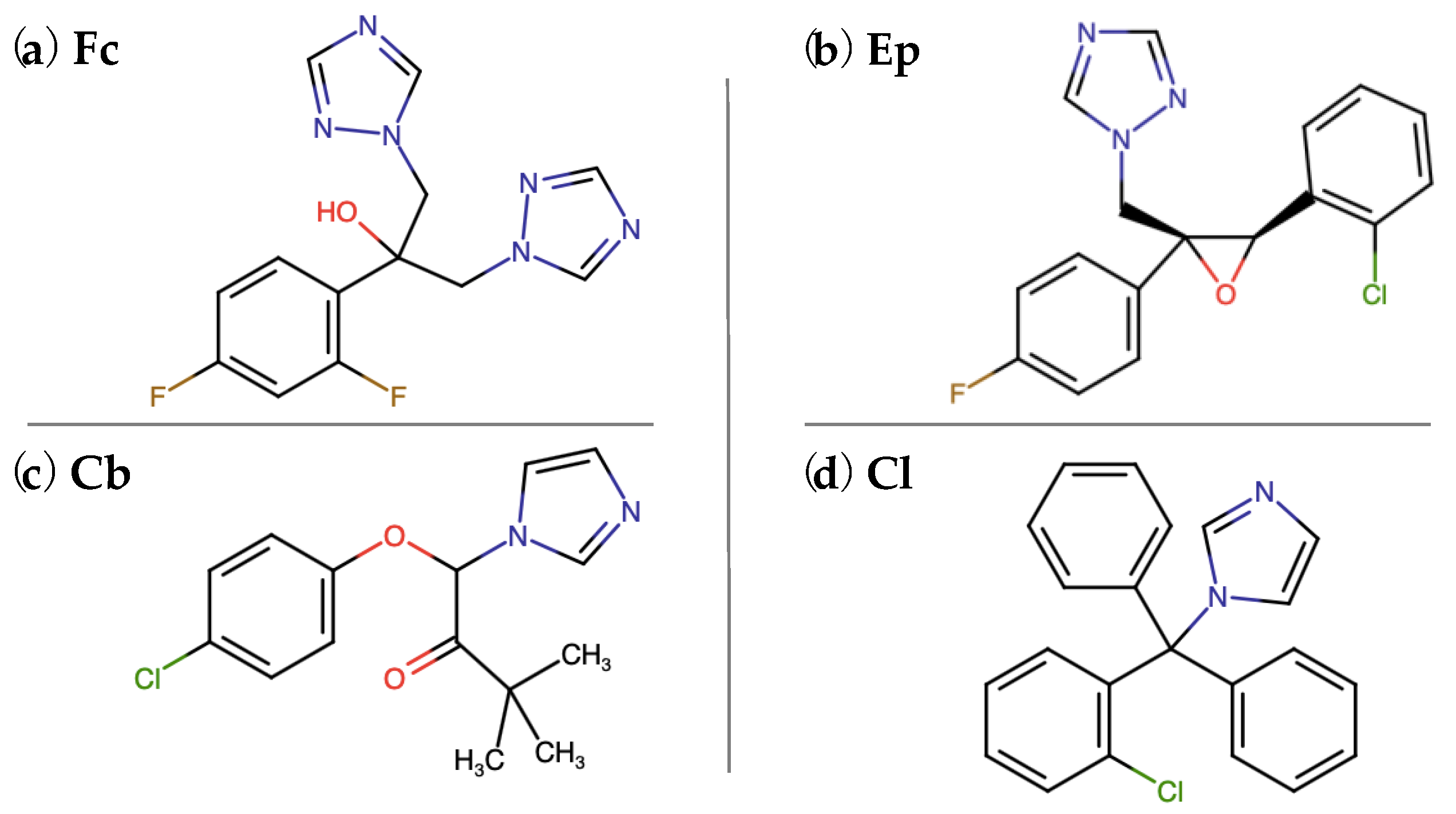
2.1. Microbial Stress Response to Azole Antifungal Agents
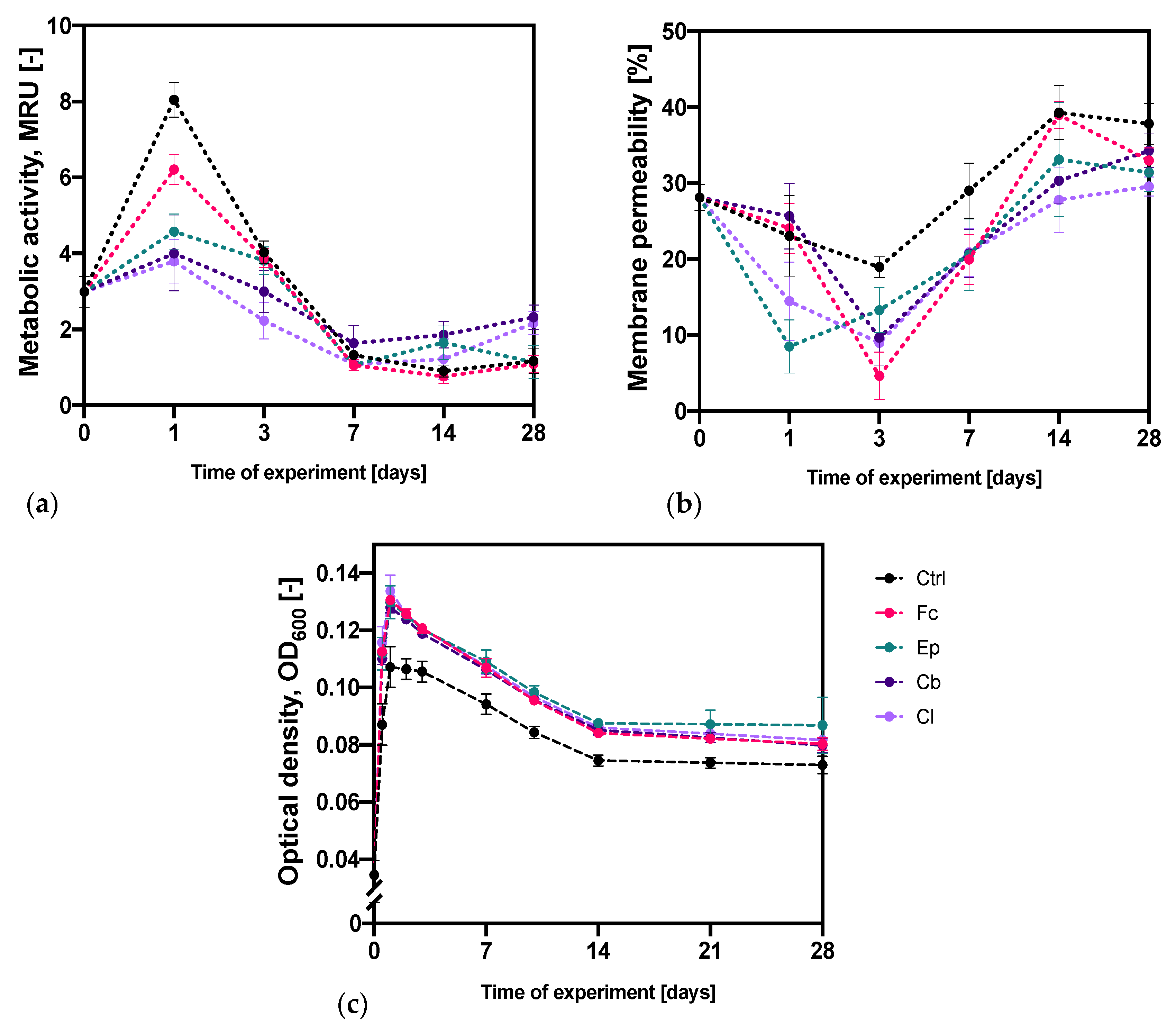
2.1.1. Analysis of the Metabolic Activity and Membrane Permeability
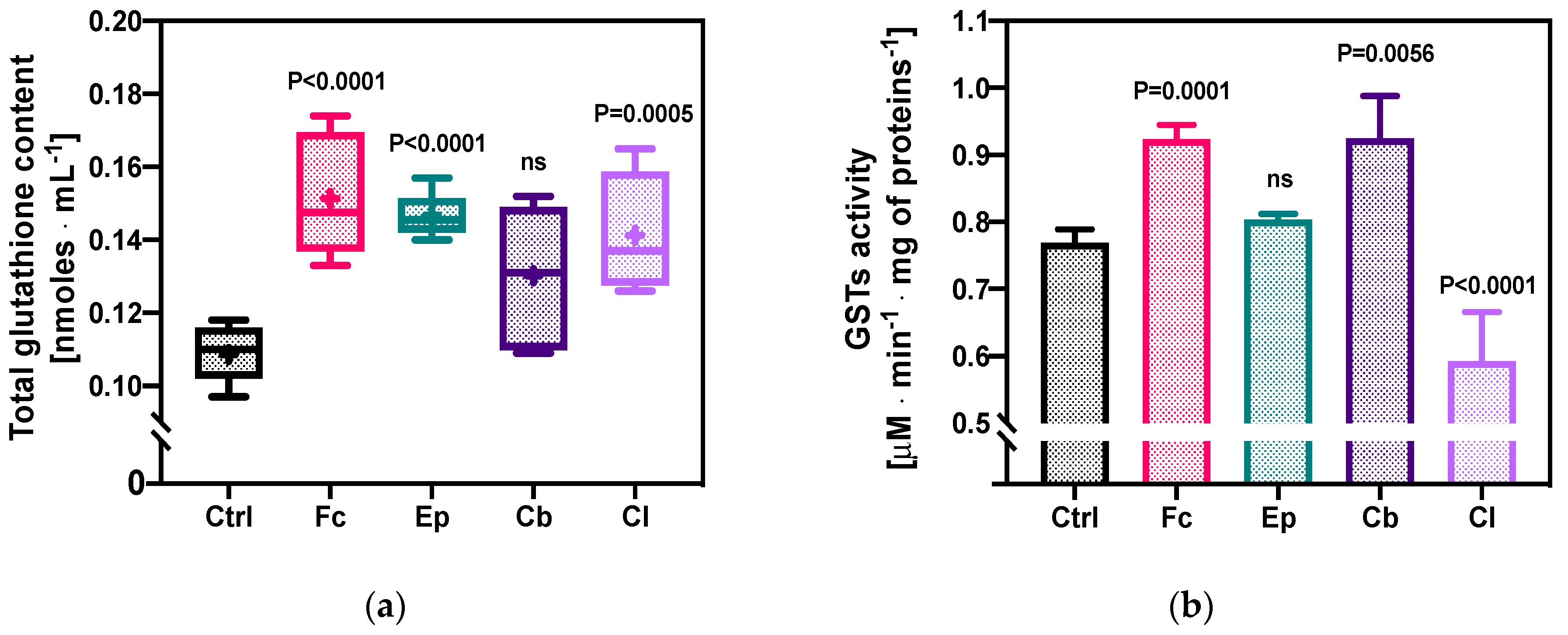
2.1.2. Total Glutathione Content and GSTs’ Enzymatic Activity
2.2. Bacterial Cell Topography and Surface Roughness Determination
2.3. Microbial Cell Surface Properties
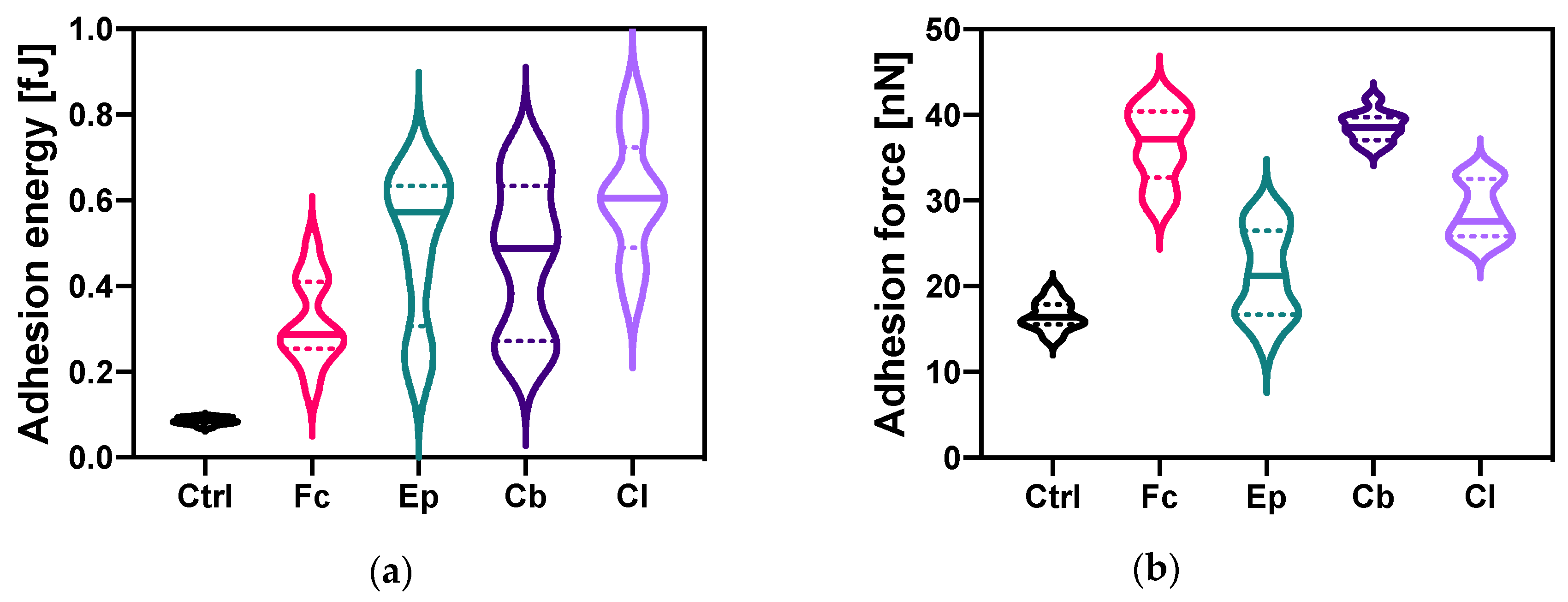
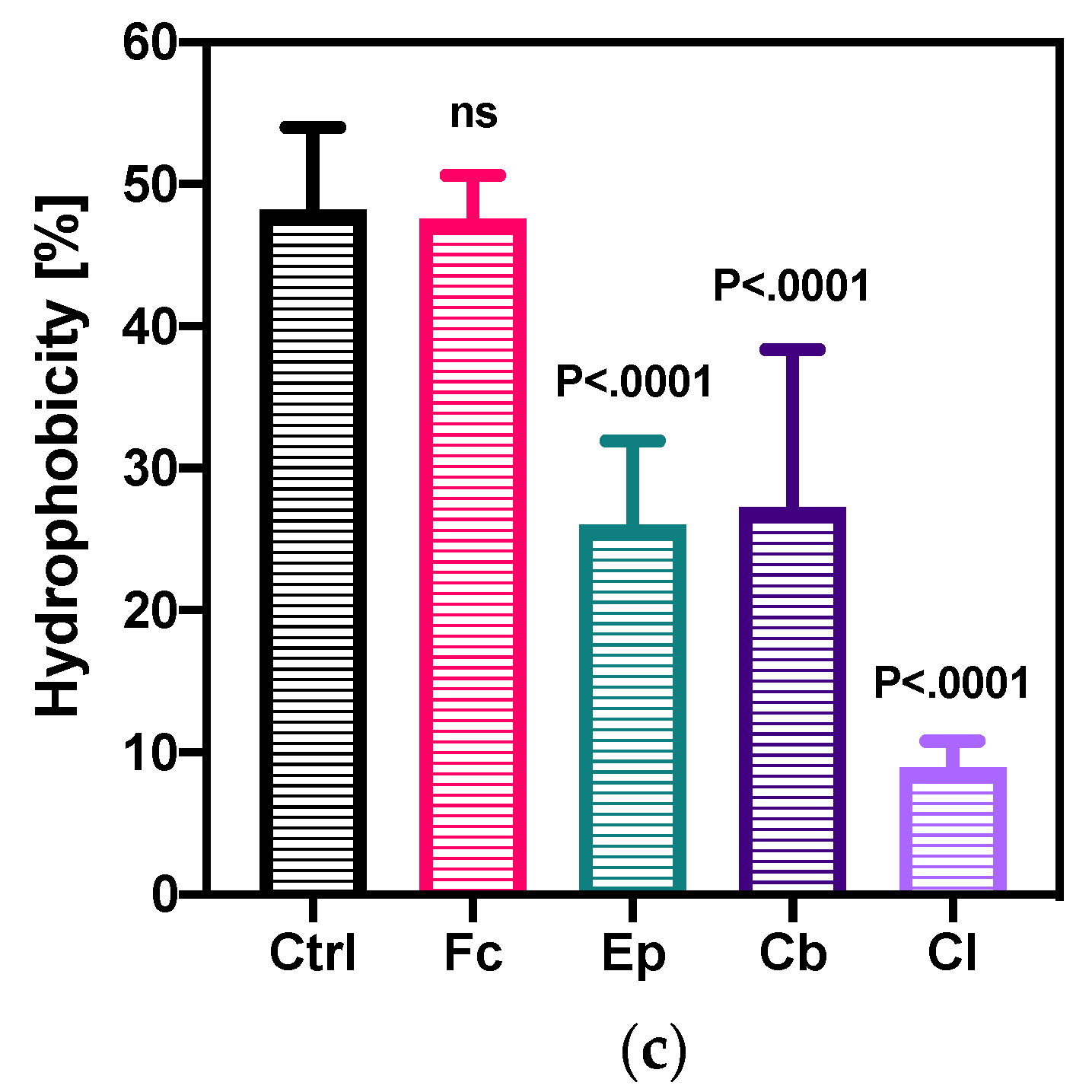
2.3.1. Changes in The Bacterial Cell Adhesion
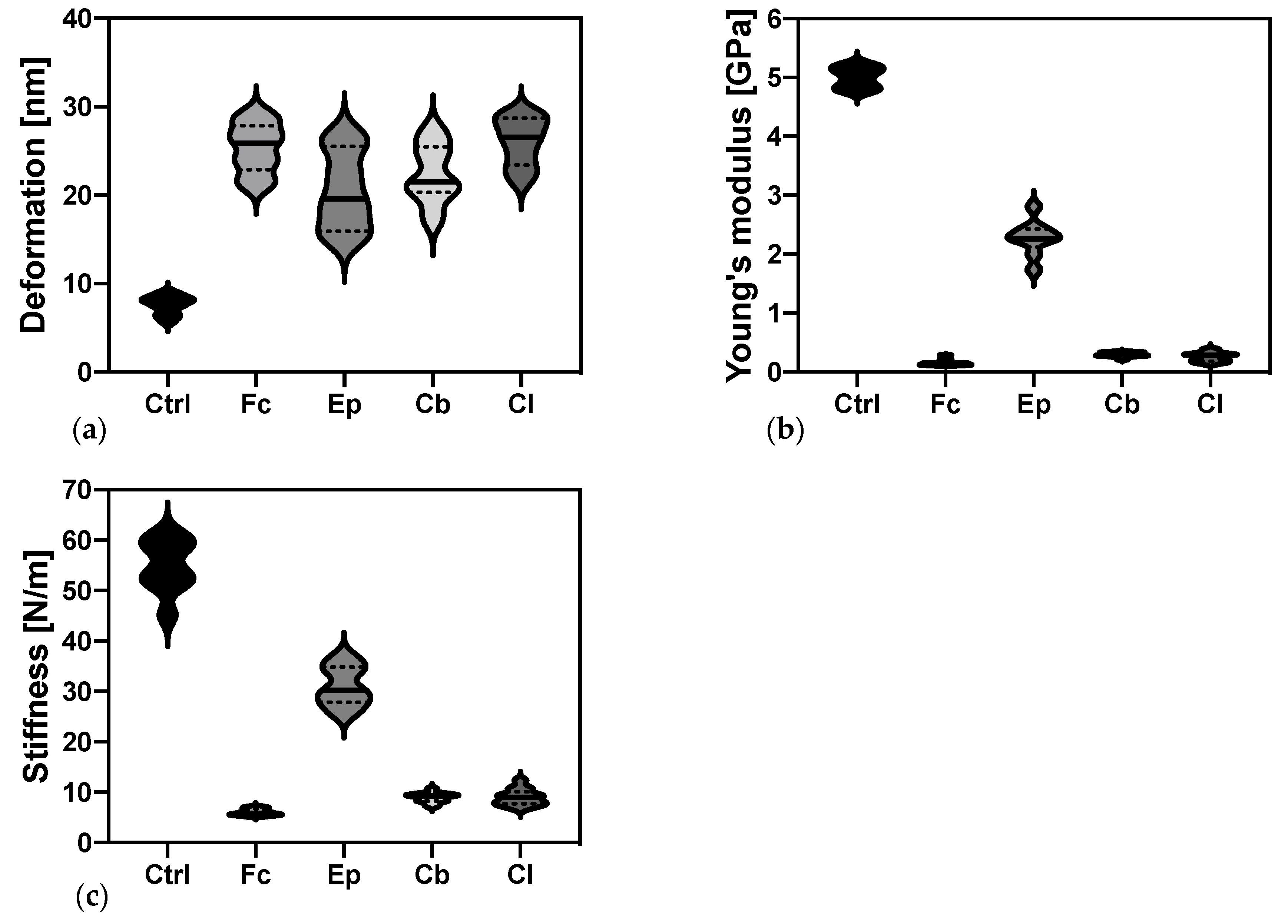
2.3.2. Elastic Properties of O. anthropi AspCl2.2 Determined by Atomic Force Microscopy
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Bacterial Strain and Culture Conditions
4.3. Analysis of the Metabolic Activity, Total Glutathione and GSTs
4.4. Preparation of Bacterial Samples for AFM Analysis and Bacterial Cells’ Surface Characterization by AFM
4.5. Nanomachanics of Bacterial Cells
4.6. Analysis of Cell Surface Hydrophobicity and Cell Surface Permeability
4.7. Statisticl Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Sheehan, D.J.; Hitchcock, C.A.; Sibley, C.M. Current and Emerging Azole Antifungal Agents. Clin. Microbiol. Rev. 1999, 12, 40–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.-F.; Ying, G.-G. Occurrence, fate and ecological risk of five typical azole fungicides as therapeutic and personal care products in the environment: A review. Environ. Int. 2015, 84, 142–153. [Google Scholar] [CrossRef]
- Tran, N.H.; Reinhard, M.; Gin, K.Y.-H. Occurrence and fate of emerging contaminants in municipal wastewater treatment plants from different geographical regions-a review. Water Res. 2018, 133, 182–207. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Huang, Q.; Zhang, K.; Yu, Y.; Wang, Z.; Wang, C. Distribution, behavior and fate of azole antifungals during mechanical, biological, and chemical treatments in sewage treatment plants in China. Sci. Total Environ. 2012, 426, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Pacholak, A.; Simlat, J.; Zgoła-Grześkowiak, A.; Kaczorek, E. Biodegradation of clotrimazole and modification of cell properties after metabolic stress and upon addition of saponins. Ecotoxicol. Environ. Saf. 2018, 161, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Crowley, P.D.; Gallagher, H.C. Clotrimazole as a pharmaceutical: Past, present and future. J. Appl. Microbiol. 2014, 117, 611–617. [Google Scholar] [CrossRef]
- Chen, Z.-F.; Ying, G.-G.; Jiang, Y.-X.; Yang, B.; Lai, H.-J.; Liu, Y.-S.; Pan, C.-G.; Peng, F.-Q. Photodegradation of the azole fungicide fluconazole in aqueous solution under UV-254: Kinetics, mechanistic investigations and toxicity evaluation. Water Res. 2014, 52, 83–91. [Google Scholar] [CrossRef]
- Santos, S.R.C.J.; Campos, E.V.; Sanches, C.; Gomez, D.S.; Ferreira, M.C. Fluconazole plasma concentration measurement by liquid chromatography for drug monitoring of burn patients. Clinics 2010, 65, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Paz-Alvarez, M.; Pudney, P.D.A.; Hadgraft, J.; Lane, M.E. Topical delivery of climbazole to mammalian skin. Int. J. Pharm. 2018, 549, 317–324. [Google Scholar] [CrossRef]
- Parker, J.E.; Warrilow, A.G.S.; Price, C.L.; Mullins, J.G.L.; Kelly, D.E.; Kelly, S.L. Resistance to antifungals that target CYP51. J. Chem. Biol. 2014, 7, 143–161. [Google Scholar] [CrossRef] [Green Version]
- Comber, S.; Gardner, M.; Sörme, P.; Leverett, D.; Ellor, B. Active pharmaceutical ingredients entering the aquatic environment from wastewater treatment works: A cause for concern? Sci. Total Environ. 2018, 613–614, 538–547. [Google Scholar] [CrossRef] [PubMed]
- Jones, V.; Gardner, M.; Ellor, B. Concentrations of trace substances in sewage sludge from 28 wastewater treatment works in the UK. Chemosphere 2014, 111, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Lees, K.; Fitzsimons, M.; Snape, J.; Tappin, A.; Comber, S. Pharmaceuticals in soils of lower income countries: Physico-chemical fate and risks from wastewater irrigation. Environ. Int. 2016, 94, 712–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Launay, M.A.; Dittmer, U.; Steinmetz, H. Organic micropollutants discharged by combined sewer overflows – Characterisation of pollutant sources and stormwater-related processes. Water Res. 2016, 104, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Tsui, M.M.P.; Leung, H.W.; Lam, P.K.S.; Murphy, M.B. Seasonal occurrence, removal efficiencies and preliminary risk assessment of multiple classes of organic UV filters in wastewater treatment plants. Water Res. 2014, 53, 58–67. [Google Scholar] [CrossRef]
- Tran, N.H.; Urase, T.; Ngo, H.H.; Hu, J.; Ong, S.L. Insight into metabolic and cometabolic activities of autotrophic and heterotrophic microorganisms in the biodegradation of emerging trace organic contaminants. Bioresour. Technol. 2013, 146, 721–731. [Google Scholar] [CrossRef]
- Chen, Z.-F.; Ying, G.-G.; Liu, Y.-S.; Zhang, Q.-Q.; Zhao, J.-L.; Liu, S.-S.; Chen, J.; Peng, F.-J.; Lai, H.-J.; Pan, C.-G. Triclosan as a surrogate for household biocides: An investigation into biocides in aquatic environments of a highly urbanized region. Water Res. 2014, 58, 269–279. [Google Scholar] [CrossRef]
- Lacey, C.; Basha, S.; Morrissey, A.; Tobin, J.M. Occurrence of pharmaceutical compounds in wastewater process streams in Dublin, Ireland. Environ. Monit. Assess. 2012, 184, 1049–1062. [Google Scholar] [CrossRef]
- Huang, Q.; Wang, Z.; Wang, C.; Peng, X. Chiral profiling of azole antifungals in municipal wastewater and recipient rivers of the Pearl River Delta, China. Environ. Sci. Pollut. Res. 2013, 20, 8890–8899. [Google Scholar] [CrossRef]
- Kahle, M.; Buerge, I.J.; Hauser, A.; Müller, M.D.; Poiger, T. Azole Fungicides: Occurrence and Fate in Wastewater and Surface Waters. Environ. Sci. Technol. 2008, 42, 7193–7200. [Google Scholar] [CrossRef]
- Zgoła-Grześkowiak, A.; Grześkowiak, T. Application of dispersive liquid-liquid microextraction followed by HPLC-MS/MS for the trace determination of clotrimazole in environmental water samples: Sample Preparation. J. Sep. Sci. 2013, 36, 2514–2521. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.-Q.; Ying, G.-G.; Chen, Z.-F.; Liu, Y.-S.; Liu, W.-R.; Zhao, J.-L. Multimedia fate modeling and risk assessment of a commonly used azole fungicide climbazole at the river basin scale in China. Sci. Total Environ. 2015, 520, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Jobling, S.; Williams, R.; Johnson, A.; Taylor, A.; Gross-Sorokin, M.; Nolan, M.; Tyler, C.R.; van Aerle, R.; Santos, E.; Brighty, G. Predicted Exposures to Steroid Estrogens in U.K. Rivers Correlate with Widespread Sexual Disruption in Wild Fish Populations. Environ. Health Perspect. 2006, 114, 32–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, A.M.S.; Goldstone, J.V.; Lamb, D.C.; Kubota, A.; Lemaire, B.; Stegeman, J.J. Identification, modeling and ligand affinity of early deuterostome CYP51s, and functional characterization of recombinant zebrafish sterol 14α-demethylase. Biochim. et Biophys. Acta (BBA) Gen. Subj. 2014, 1840, 1825–1836. [Google Scholar] [CrossRef] [Green Version]
- Baudiffier, D.; Hinfray, N.; Ravaud, C.; Creusot, N.; Chadili, E.; Porcher, J.-M.; Schulz, R.W.; Brion, F. Effect of in vivo chronic exposure to clotrimazole on zebrafish testis function. Environ. Sci. Pollut. Res. 2013, 20, 2747–2760. [Google Scholar] [CrossRef] [Green Version]
- Geiger, E.; Hornek-Gausterer, R.; Saçan, M.T. Single and mixture toxicity of pharmaceuticals and chlorophenols to freshwater algae Chlorella vulgaris. Ecotoxicol. Environ. Saf. 2016, 129, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Chonova, T.; Labanowski, J.; Cournoyer, B.; Chardon, C.; Keck, F.; Laurent, É.; Mondamert, L.; Vasselon, V.; Wiest, L.; Bouchez, A. River biofilm community changes related to pharmaceutical loads emitted by a wastewater treatment plant. Environ. Sci. Pollut. Res. 2018, 25, 9254–9264. [Google Scholar] [CrossRef]
- Ager, D.; Evans, S.; Li, H.; Lilley, A.K.; van der Gast, C.J. Anthropogenic disturbance affects the structure of bacterial communities. Environ. Microbiol. 2010, 12, 670–678. [Google Scholar] [CrossRef]
- Ebele, A.J.; Abou-Elwafa Abdallah, M.; Harrad, S. Pharmaceuticals and personal care products (PPCPs) in the freshwater aquatic environment. Emerg. Contam. 2017, 3, 1–16. [Google Scholar] [CrossRef]
- Guan, N.; Li, J.; Shin, H.; Du, G.; Chen, J.; Liu, L. Microbial response to environmental stresses: From fundamental mechanisms to practical applications. Appl. Microbiol. Biotechnol. 2017, 101, 3991–4008. [Google Scholar] [CrossRef]
- Villagrasa, E.; Ballesteros, B.; Obiol, A.; Millach, L.; Esteve, I.; Solé, A. Multi-approach analysis to assess the chromium(III) immobilization by Ochrobactrum anthropi DE2010. Chemosphere 2020, 238, 124663. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Hou, R.; Gao, S.; Sun, Z.; Li, X.; Zhai, L.; Jin, Y.; Zhu, Q.; Liao, Y.; Tian, K. First Isolation and Characterization of Ochrobactrum anthropi from Pig. Engineering 2020, 6, 49–55. [Google Scholar] [CrossRef]
- Seleem, M.N.; Ali, M.; Boyle, S.M.; Mukhopadhyay, B.; Witonsky, S.G.; Schurig, G.G.; Sriranganathan, N. Establishment of a Gene Expression System in Ochrobactrum anthropi. AEM 2006, 72, 6833–6836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.-W.; Xu, M.; Ma, X.-W.; Tong, Z.-H.; Liu, D.-F. Isolation and characterization of a chlorate-reducing bacterium Ochrobactrum anthropi XM-1. J. Hazard. Mater. 2019, 380, 120873. [Google Scholar] [CrossRef] [PubMed]
- Horváthová, H.; Lászlová, K.; Dercová, K. Bioremediation vs. Nanoremediation: Degradation of Polychlorinated Biphenyls (PCBS) Using Integrated Remediation Approaches. Water Air Soil Pollut. 2019, 230, 204. [Google Scholar] [CrossRef]
- Hou, X.; Huang, L.; Zhou, P.; Tian, F.; Tao, Y.; Li Puma, G. Electrosynthesis of acetate from inorganic carbon (HCO3−) with simultaneous hydrogen production and Cd(II) removal in multifunctional microbial electrosynthesis systems (MES). J. Hazard. Mater. 2019, 371, 463–473. [Google Scholar] [CrossRef]
- Lei, X.; Jia, Y.; Chen, Y.; Hu, Y. Simultaneous nitrification and denitrification without nitrite accumulation by a novel isolated Ochrobactrum anthropic LJ81. Biores. Technol. 2019, 272, 442–450. [Google Scholar] [CrossRef]
- Gwenzi, W.; Chaukura, N. Organic contaminants in African aquatic systems: Current knowledge, health risks, and future research directions. Sci. Total Environ. 2018, 619–620, 1493–1514. [Google Scholar] [CrossRef]
- Löffler, D.; Römbke, J.; Meller, M.; Ternes, T.A. Environmental fate of pharmaceuticals in water/sediment systems. Environ. Sci. Technol. 2005, 39, 5209–5218. [Google Scholar] [CrossRef]
- Yan, B.; Ye, F.; Gao, D. Residues of the fungicide epoxiconazole in rice and paddy in the Chinese field ecosystem: Fungicide epoxiconazole residues in Chinese rice and paddy fields. Pest. Manag. Sci. 2015, 71, 65–71. [Google Scholar] [CrossRef]
- Itri, R.; Junqueira, H.C.; Mertins, O.; Baptista, M.S. Membrane changes under oxidative stress: The impact of oxidized lipids. Biophys. Rev. 2014, 6, 47–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergheim, M.; Gminski, R.; Spangenberg, B.; Debiak, M.; Bürkle, A.; Mersch-Sundermann, V.; Kümmerer, K.; Gieré, R. Antibiotics and sweeteners in the aquatic environment: Biodegradability, formation of phototransformation products, and in vitro toxicity. Environ. Sci. Pollut. Res. 2015, 22, 18017–18030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Niu, Z.; Yin, K.; Liu, F.; Chen, L. Degradation of furazolidone by bacteria Acinetobacter calcoaceticus T32, Pseudomonas putida SP1 and Proteus mirabilis V7. Int. Biodeterior. Biodegrad. 2013, 77, 45–50. [Google Scholar] [CrossRef]
- Oberoi, A.S.; Philip, L. Variation in toxicity during the biodegradation of various heterocyclic and homocyclic aromatic hydrocarbons in single and multi-substrate systems. Ecotoxicol. Environ. Saf. 2017, 135, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Smułek, W.; Zdarta, A.; Pacholak, A.; Runka, T.; Kaczorek, E. Increased biological removal of 1-chloronaphthalene as a result of exposure: A study of bacterial adaptation strategies. Ecotoxicol. Environ. Saf. 2019, 185, 109707. [Google Scholar] [CrossRef]
- Cao, P.; Wall, D. The Fluidity of the Bacterial Outer Membrane Is Species Specific: Bacterial Lifestyles and the Emergence of a Fluid Outer Membrane. BioEssays 2020, 1900246. [Google Scholar] [CrossRef]
- Monostori, P.; Wittmann, G.; Karg, E.; Túri, S. Determination of glutathione and glutathione disulfide in biological samples: An in-depth review. J. Chromatogr. B 2009, 877, 3331–3346. [Google Scholar] [CrossRef]
- Smirnova, G.V.; Oktyabrsky, O.N. Glutathione in Bacteria. Biochemistry (Moscow) 2005, 70, 1199–1211. [Google Scholar] [CrossRef]
- Allocati, N.; Federici, L.; Masulli, M.; Di Ilio, C. Glutathione transferases in bacteria: Bacterial GSTs. FEBS J. 2009, 276, 58–75. [Google Scholar] [CrossRef]
- Storia, A.L. Atomic force microscopy analysis shows surface structure changes in carvacrol-treated bacterial cells. Res. Microbiol. 2011, 9. [Google Scholar] [CrossRef]
- Uzoechi, S.C.; I Abu-Lail, N. Variations in the Morphology, Mechanics and Adhesion of Persister and Resister E. coli Cells in Response to Ampicillin: AFM Study. Antibiotics 2020, 9, 235. [Google Scholar] [CrossRef] [PubMed]
- Potthoff, E.; Ossola, D.; Zambelli, T.; Vorholt, J.A. Bacterial adhesion force quantification by fluidic force microscopy. Nanoscale 2015, 7, 4070–4079. [Google Scholar] [CrossRef] [PubMed]
- El-Kirat-Chatel, S.; Beaussart, A.; Mathelié-Guinlet, M.; Dufrêne, Y.F. The importance of force in microbial cell adhesion. Curr. Opin. Colloid Interface Sci. 2020, 47, 111–117. [Google Scholar] [CrossRef]
- Francius, G.; Domenech, O.; Mingeot-Leclercq, M.P.; Dufrêne, Y.F. Direct Observation of Staphylococcus aureus Cell Wall Digestion by Lysostaphin. JB 2008, 190, 7904–7909. [Google Scholar] [CrossRef] [Green Version]
- Pacholak, A.; Smułek, W.; Zgoła-Grześkowiak, A.; Kaczorek, E. Nitrofurantoin—Microbial Degradation and Interactions with Environmental Bacterial Strains. IJERPH 2019, 16, 1526. [Google Scholar] [CrossRef] [Green Version]
- Kaczorek, E.; Sałek, K.; Guzik, U.; Dudzińska-Bajorek, B. Cell surface properties and fatty acids composition of Stenotrophomonas maltophilia under the influence of hydrophobic compounds and surfactants. New Biotechnol. 2013, 30, 173–182. [Google Scholar] [CrossRef]
- Wang, H.; Cheng, H.; Wang, F.; Wei, D.; Wang, X. An improved 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) reduction assay for evaluating the viability of Escherichia coli cells. J. Microbiol. Methods 2010, 82, 330–333. [Google Scholar] [CrossRef]
- Ambalam, P.; Kondepudi, K.K.; Nilsson, I.; Wadström, T.; Ljungh, Å. Bile stimulates cell surface hydrophobicity, Congo red binding and biofilm formation of Lactobacillus strains. FEMS Microbiol. Lett. 2012, 333, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Halder, S.; Yadav, K.K.; Sarkar, R.; Mukherjee, S.; Saha, P.; Haldar, S.; Karmakar, S.; Sen, T. Alteration of Zeta potential and membrane permeability in bacteria: A study with cationic agents. SpringerPlus 2015, 4. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ctrl | Fc | Ep | Cb | Cl | |
|---|---|---|---|---|---|
| Cell length [μm] | 0.62 ± 0.08 | 0.75 ± 0.08 * | 0.71 ± 0.16 | 0.76 ± 0.13 * | 0.78 ± 0.10 * |
| Cell width [μm] | 0.34 ± 0.06 | 0.43 ± 0.06 | 0.49 ± 0.12 * | 0.47 ± 0.04 * | 0.37 ± 0.06 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pacholak, A.; Burlaga, N.; Kaczorek, E. Evaluating the Effect of Azole Antifungal Agents on the Stress Response and Nanomechanical Surface Properties of Ochrobactrum anthropi Aspcl2.2. Molecules 2020, 25, 3348. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25153348
Pacholak A, Burlaga N, Kaczorek E. Evaluating the Effect of Azole Antifungal Agents on the Stress Response and Nanomechanical Surface Properties of Ochrobactrum anthropi Aspcl2.2. Molecules. 2020; 25(15):3348. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25153348
Chicago/Turabian StylePacholak, Amanda, Natalia Burlaga, and Ewa Kaczorek. 2020. "Evaluating the Effect of Azole Antifungal Agents on the Stress Response and Nanomechanical Surface Properties of Ochrobactrum anthropi Aspcl2.2" Molecules 25, no. 15: 3348. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25153348