
The New Coronavirus (SARS-CoV-2): A Comprehensive Review on Immunity and the Application of Bioinformatics and Molecular Modeling to the Discovery of Potential Anti-SARS-CoV-2 Agents

 ,
,  , ,
, ,  , , , , , , add
Show full author list
, , , , , , add
Show full author list
Abstract
:1. Introduction
2. Coronavirus
Etiological Agent
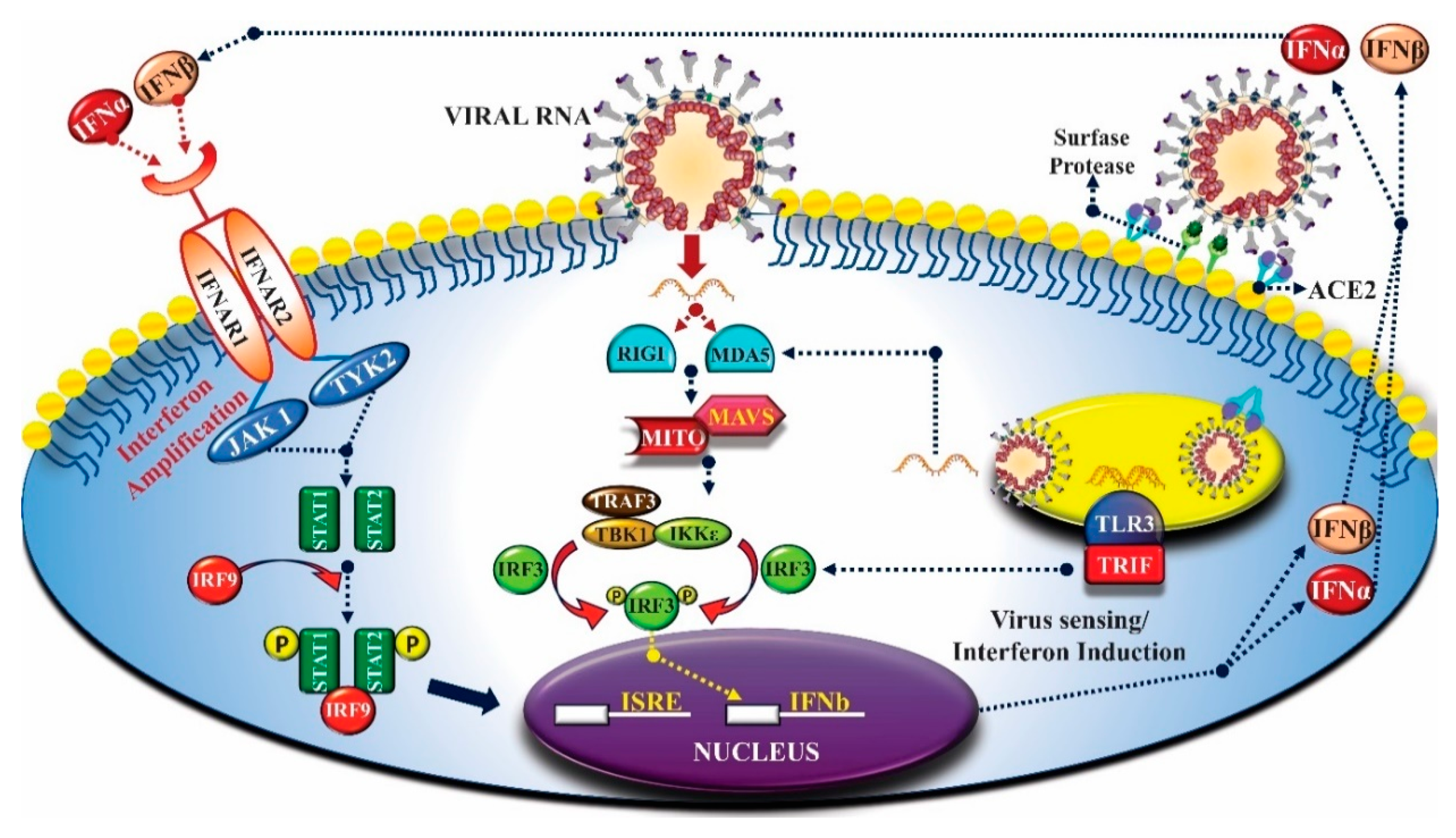
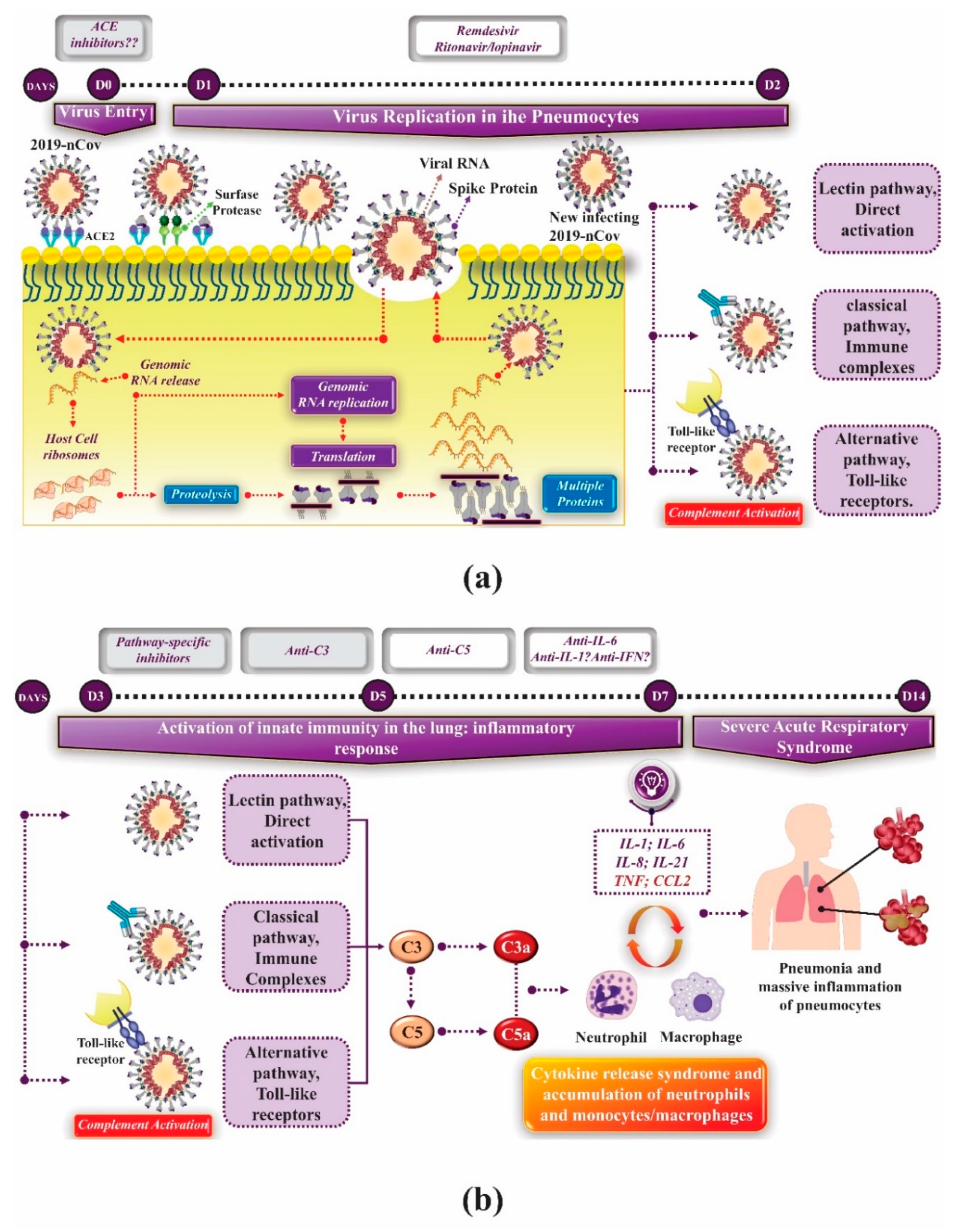
3. Innate Immunity and Coronavirus
4. Adaptative Immunity and Coronavirus
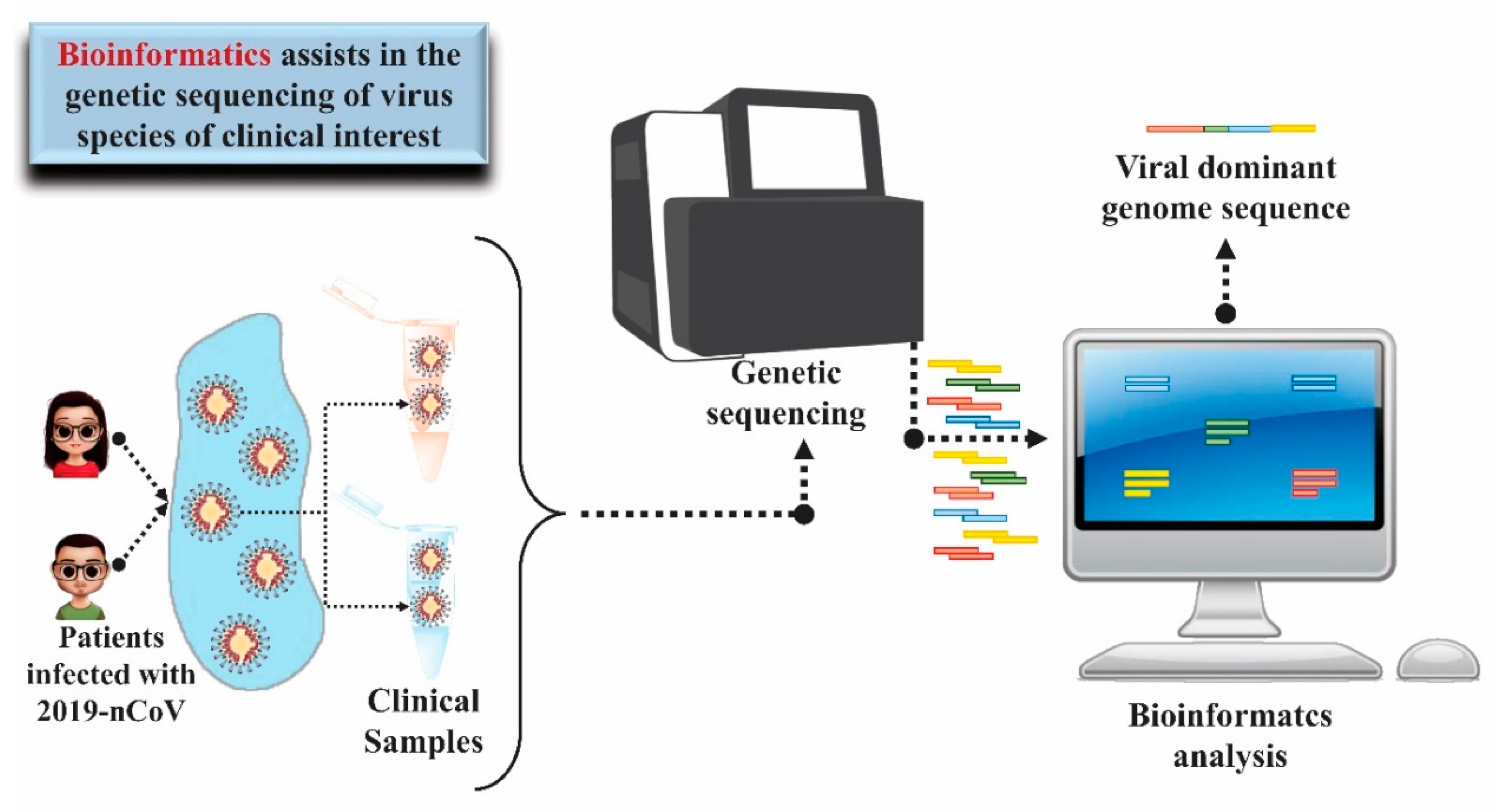
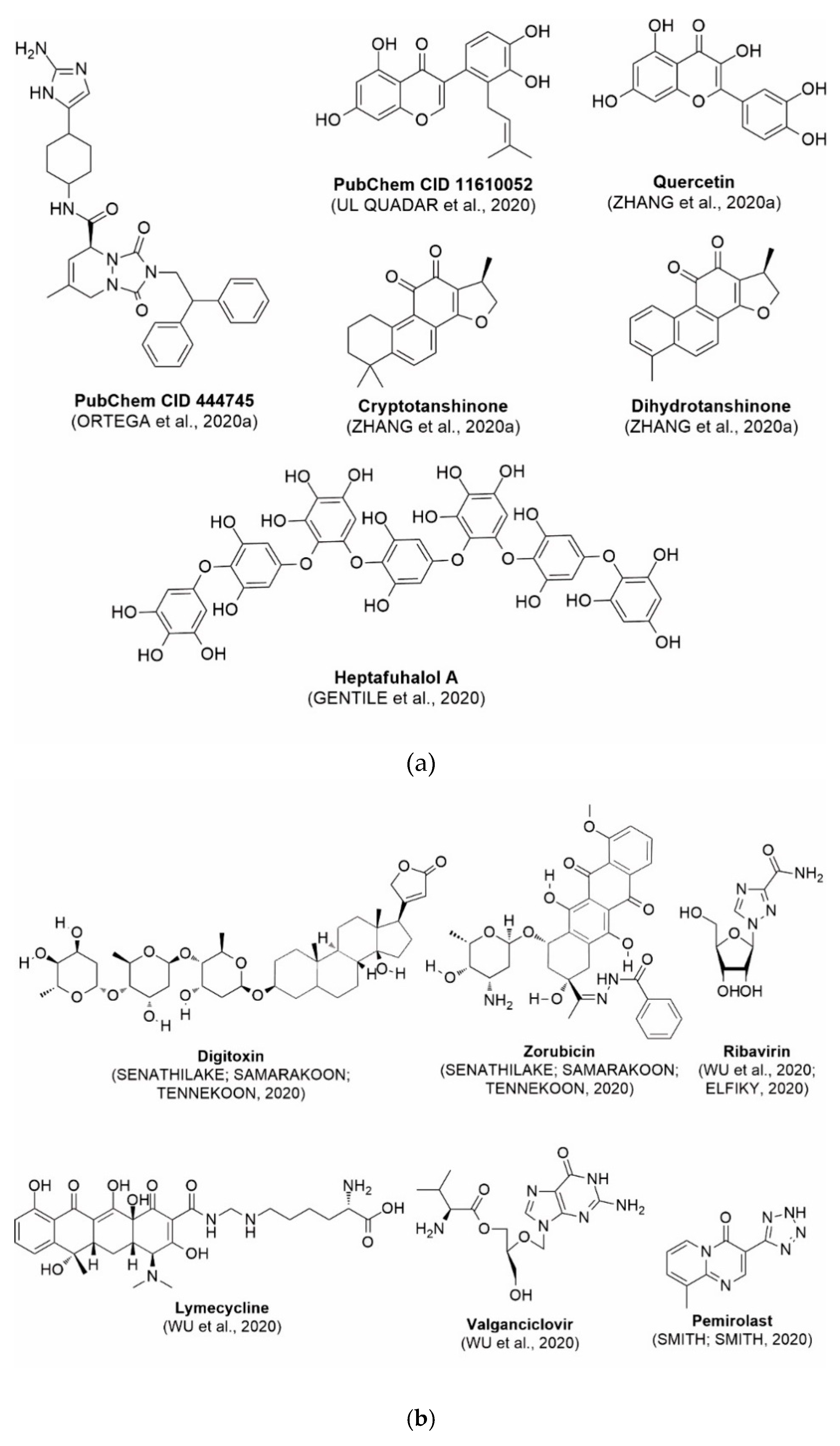
5. Use of Bioinformatics and Molecular Modeling as Technology Applied to Health to Diagnose and Treat COVID-19
6. Future Perspectives on Bioinformatics and Molecular Modeling as Prospecting Tools Against SARS-CoV-2
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Li, Q.; Guan, X.; Wu, P.; Wang, X.; Zhou, L.; Tong, Y.; Ren, R.; Leung, K.S.M.; Lau, E.H.Y.; Wong, J.Y.; et al. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus–Infected Pneumonia. N. Engl. J. Med. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. Severe acute respiratory syndrome related coronavirus: The species and its viruses—A statement of the Coronavirus Study Group. bioRxiv 2020, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Zhou, Y.; Liu, X.; Tan, J. The impact of the COVID-19 epidemic on the utilization of emergency dental services. J. Dent. Sci. 2020. In Press. [Google Scholar] [CrossRef]
- Singh, A.K.; Singh, A.; Shaikh, A.; Singh, R.; Misra, A. Chloroquine and hydroxychloroquine in the treatment of COVID-19 with or without diabetes: A systematic search and a narrative review with a special reference to India and other developing countries. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 1–35. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 6736, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.; Yang, X.; Wang, X.; Hu, B.; Zhang, L.; Zhang, W.; Guo, H.; Jiang, R.; Liu, M.; Chen, Y.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wu, Z.; Ren, X.; Yang, F.; He, G.; Zhang, J.; Dong, J.; Sun, L.; Zhu, Y.; Du, J.; et al. Novel SARS-like betacoronaviruses in bats, China, 2011. Emerg. Infect. Dis. 2013, 19, 989–991. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Zeng, L.P.; Yang, X.L.; Ge, X.Y.; Zhang, W.; Li, B.; Xie, J.Z.; Shen, X.R.; Zhang, Y.Z.; Wang, N.; et al. Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus. PLoS Pathog. 2017, 13, 1–27. [Google Scholar] [CrossRef]
- Wang, N.; Li, S.Y.; Yang, X.L.; Huang, H.M.; Zhang, Y.J.; Guo, H.; Luo, C.M.; Miller, M.; Zhu, G.; Chmura, A.A.; et al. Serological Evidence of Bat SARS-Related Coronavirus Infection in Humans, China. Virol. Sin. 2018, 33, 104–107. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 3405, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19. 2020. Available online: https://www.who.int/dg/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020 (accessed on 4 April 2020).
- WHO Novel Coronavirus (2019-nCoV), Situation Report—12. 2020. Available online: https://www.who.int/timorleste/emergencies/novel-coronavirus-2019/novel-coronavirus-(2019-ncov)-situation-reports (accessed on 4 April 2020).
- Sohrabia, C.; Alsafib, Z.; O’Neilla, N.; Khanb, M.; Kerwanc, A.; Al-Jabirc, A.; Iosifidisa, C.; Aghad, R. World Health Organization declares global emergency: A review of the 2019 novel Coronavirus (COVID-19). Int. J. Surg. 2020, 76, 128–129. [Google Scholar] [CrossRef] [PubMed]
- WHO Novel Coronavirus (COVID-19) Situation. Available online: https://experience.arcgis.com/experience/685d0ace521648f8a5beeeee1b9125cd (accessed on 28 August 2020).
- Li, X.; Zai, J.; Zhao, Q.; Nie, Q.; Li, Y.; Foley, B.T.; Chaillon, A. Evolutionary history, potential intermediate animal host, and cross-species analyses of SARS-CoV-2. J. Med. Virol. 2020, 1–10. [Google Scholar] [CrossRef]
- Robson, B. Computers and viral diseases. Preliminary bioinformatics studies on the design of a synthetic vaccine and a preventative peptidomimetic antagonist against the SARS-CoV-2 (2019-nCoV, COVID-19) coronavirus. Comput. Biol. Med. 2020, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Mullins, I.M.; Siadaty, M.S.; Lyman, J.; Scully, K.; Garrett, C.T.; Miller, W.G.; Muller, R.; Robson, B.; Apte, C.; Weiss, S.; et al. Data mining and clinical data repositories: Insights from a 667,000 patient data set. Comput. Biol. Med. 2007, 36, 1351–1377. [Google Scholar] [CrossRef] [PubMed]
- Robson, B.; Li, J.; Dettinger, R.; Peters, A.; Boyer, S.K. Drug Discovery Using Very Large Numbers of Patents. General Strategy with Extensive Use of Match and Edit Operations. J. Comput. Aided. Mol. Des. 2011, 25, 427–441. [Google Scholar] [CrossRef] [PubMed]
- Robson, B.; Caruso, T.P.; Balis, U.G.J. Suggestions for a Web based universal exchange and inference language for medicine. Comput. Biol. Med. 2013, 43, 2297–2310. [Google Scholar] [CrossRef]
- Frieman, M.; Heise, M.; Baric, R. SARS coronavirus and innate immunity. Virus Res. 2008, 133, 101–112. [Google Scholar] [CrossRef]
- Li, J.; You, Z.; Wang, Q.; Zhou, Z.; Qiu, Y.; Luo, R.; Ge, X. The epidemic of 2019-novel-coronavirus (2019-nCoV) pneumonia and insights for emerging infectious diseases in the future. Microbes Infect. 2020, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Lam, C.W.K.; Wu, A.K.L.; Ip, W.K.; Lee, N.L.S.; Chan, I.H.S.; Lit, L.C.W.; Hui, D.S.C.; Chan, M.H.M.; Chung, S.S.C.; et al. Plasma inflammatory cytokines and chemokines in severe acute respiratory syndrome. Clin. Exp. Immunol. 2004, 136, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahallawi, W.H.; Khabour, O.F.; Zhang, Q.; Makhdoum, H.M.; Suliman, B.A. MERS-CoV infection in humans is associated with a pro-inflammatory Th1 and Th17 cytokine profile. Cytokine 2018, 104, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Vargas, A.R.; Ferreira, A.C.N.N.J.; Diana, P.M.C.; Suassuna, T.D.S. Protocolo de Tratamento do Novo Coronavírus (2019-nCoV), 1st ed.; Ministério da Saúde: Brasília-DF, Brazil, 2020.
- Elfiky, A.A.; Mahdy, S.M.; Elshemey, W.M. Quantitative structure-activity relationship and molecular docking revealed a potency of anti-hepatitis C virus drugs against human corona viruses. J. Med. Virol. 2017, 89, 1040–1047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cascella, M.; Rajnik, M.; Cuomo, A.; Dulebohn, S.C.; Di Napoli, R. Features, Evaluation and Treatment Coronavirus (COVID-19); StatPearls Publishing: Napo, Italy, 2020. [Google Scholar]
- Holmes, K.V. Coronaviruses. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott Willians & Wilkins: Philadelphia, PA, USA, 2001; pp. 1187–1203. [Google Scholar]
- Mahony, J.B.; Richardson, S. Molecular Diagnosis of Severe Acute Respiratory. J. Mol. Diagnostics 2005, 7, 551–559. [Google Scholar] [CrossRef] [Green Version]
- Van Der Hoek, L.; Pyrc, K.; Berkhout, B. Human coronavirus NL63, a new respiratory virus. FEMS Microbiol. Rev. 2006, 30, 760–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, S.R.; Navas-martin, S.; Weiss, S.R.; Navas-martin, S. Coronavirus Pathogenesis and the Emerging Pathogen Severe Acute Respiratory Syndrome Coronavirus. Microbiol. Mol. Biol. Rev. 2005, 69, 635–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woo, P.C.Y.; Lau, S.K.P.; Li, K.S.M.; Poon, R.W.S.; Wong, B.H.L.; Tsoi, H.; Yip, B.C.K.; Huang, Y.; Chan, K.; Yuen, K. Molecular diversity of coronaviruses in bats. Virology 2006, 351, 180–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Cao, R.; Zhang, L.; Yang, X.; Liu, J.; Xu, M.; Shi, Z.; Hu, Z.; Zhong, W.; Xiao, G. Remdesivir and chloroquine effectively inhibit the recently emerged novel coronavirus (2019-nCoV) in vitro. Cell Res. 2020, 30, 269–271. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.F.; To, K.K.; Tse, H.; Jin, D.; Yuen, K. Interspecies transmission and emergence of novel viruses: Lessons from bats and birds. Trends Microbiol. 2013, 21, 1–12. [Google Scholar] [CrossRef]
- Guan, Y.; Cheung, C.L.; Luo, S.W.; Li, P.H.; Zhang, L.J.; Guan, Y.J.; Yuen, K.Y.; Peiris, J.S.M.; Poon, L.L.M. Isolation and Characterization of Viruses Related to the SARS Coronavirus from Animals in Southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [Green Version]
- Poon, L.L.M.; Chu, D.K.W.; Chan, K.H.; Wong, O.K.; Ellis, T.M.; Leung, Y.H.C.; Lau, S.K.P.; Woo, P.C.Y.; Suen, K.Y.; Yuen, K.Y.; et al. Identification of a Novel Coronavirus in Bats. J. Virol. 2009, 79, 2001–2009. [Google Scholar] [CrossRef] [Green Version]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nat. Rev. Microbiol. 2009, 7, 439–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Shi, Z.; Yu, M.; Ren, W.; Smith, C.; Epstein, J.H.; Wang, H.; Crameri, G.; Hu, Z.; Zhang, H.; et al. Bats Are Natural Reservoirs of SARS-Like Coronaviruses. Science 2005, 310, 676–680. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Woo, P.C.Y.; Li, K.S.M.; Huang, Y.; Tsoi, H.; Wong, B.H.L.; Wong, S.S.Y.; Leung, S.; Chan, K.; Yuen, K. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc. Natl. Acad. Sci. USA 2005, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vijgen, L.; Keyaerts, E.; Moe, E.; Thoelen, I.; Wollants, E.; Lemey, P.; Vandamme, A.; Ranst, M. Van Complete Genomic Sequence of Human Coronavirus OC43: Molecular Clock Analysis Suggests a Relatively Recent Zoonotic Coronavirus Transmission Event. J. Virol. 2005, 79, 1595–1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alekseev, K.P.; Vlasova, A.N.; Jung, K.; Hasoksuz, M.; Zhang, X.; Halpin, R.; Wang, S.; Ghedin, E.; Spiro, D.; Saif, L.J. Bovine-Like Coronaviruses Isolated from Four Species of Captive Wild Ruminants Are Homologous to Bovine Coronaviruses, Based on Complete Genomic Sequences. J. Virol. 2008, 82, 12422–12431. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Cebra, C.K.; Baker, R.J.; Mattson, D.E.; Cohen, S.A.; Alvarado, D.E.; Rohrmann, G.F. Analysis of the genome sequence of an alpaca coronavirus. Virology 2007, 365, 198–203. [Google Scholar] [CrossRef] [Green Version]
- Lorusso, A.; Decaro, N.; Schellen, P.; Rottier, P.J.M.; Buonavoglia, C.; Haijema, B.; De Groot, R.J. Gain, Preservation, and Loss of a Group 1a Coronavirus Accessory Glycoprotein. J. Virol. 2008, 82, 10312–10317. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Liu, Q.; Guo, D. Coronaviruses: Genome structure, replication, and pathogenesis. J. Med. Virol. 2020, 92, 418–423. [Google Scholar] [CrossRef]
- Chen, H.; Guo, J.; Wang, C.; Luo, F.; Yu, X.; Zhang, W.; Li, J.; Zhao, D.; Xu, D.; Gong, Q.; et al. Clinical characteristics and intrauterine vertical transmission potential of COVID-19 infection in nine pregnant women: A retrospective review of medical records. Lancet 2020, 395, 809–815. [Google Scholar] [CrossRef] [Green Version]
- Bauch, C.T.; Lloyd-Smith, J.O.; Coffee, M.P.; Galvani, A.P. Dynamically modeling SARS and other newly emerging respiratory illnesses: Past, present, and future. Epidemiology 2005, 16, 791–801. [Google Scholar] [CrossRef] [Green Version]
- Rothan, H.A.; Byrareddy, S.N. The epidemiology and pathogenesis of coronavirus disease (COVID-19) outbreak. J. Autoimmun. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Shang, J.; Graham, R.; Baric, R.S.; Li, F. Receptor recognition by novel coronavirus from Wuhan: An analysis based on decade-long structural studies of SARS. J. Virol. 2020, 84, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaimes, J.A.; Millet, J.K.; Stout, A.E.; André, N.M.; Whittaker, G.R. A tale of two viruses: The distinct spike glycoproteins of feline coronaviruses. Viruses 2020, 12, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belouzard, S.; Chu, V.C.; Whittaker, G.R. Activation of the SARS coronavirus spike protein via sequential proteolytic cleavage at two distinct sites. Proc. Nalt. Acad. Sci. USA 2009, 106, 5871–5876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qing, E.; Gallagher, T. Spotlight SARS Coronavirus Redux. Trends Immunol. 2020, 1–3. [Google Scholar] [CrossRef]
- Katze, M.G.; He, Y.; Gale, M. Viruses and interferon: A fight for supremacy. Nat. Rev. Immunol. 2002, 2, 675–687. [Google Scholar] [CrossRef] [PubMed]
- García-Sastre, A.; Biron, C.A. Type 1 interferons and the virus-host relationship: A lesson in détente. Science 2006, 312, 879–882. [Google Scholar] [CrossRef]
- Sen, G.C. Viruses and Interferons. Annu. Rev. Microbiol. 2001, 55, 255–281. [Google Scholar] [CrossRef]
- De Wit, E.; Van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Li, G.; Fan, Y.; Lai, Y.; Han, T.; Li, Z.; Zhou, P.; Pan, P.; Wang, W.; Hu, D.; Liu, X.; et al. Coronavirus Infections and Immune Responses. J. Med. Virol. 2020, 92, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Gralinski, L.E.; Sheahan, T.P.; Morrison, T.E.; Menachery, V.D.; Jensen, K.; Leist, S.R.; Whitmore, A.; Heise, M.T.; Baric, R.S. Complement Activation Contributes to Severe Acute Respiratory Syndrome Coronavirus Pathogenesis. mBio 2018, 9, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Risitano, A.M.; Mastellos, D.C.; Huber-lang, M.; Yancopoulou, D.; Garlanda, C.; Ciceri, F.; Lambris, J.D. Complement as a target in COVID-19? Nat. Rev. Immunol. 2020, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastellos, D.C.; Ricklin, D.; Lambris, J.D. Clinical promise of next-generation complement therapeutics. Nat. Rev. Drug Discov. 2019, 18, 707–729. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Hu, M.; Zhang, X.; Li, H.; Zhu, L.; Liu, H.; Dong, Q.; Zhang, Z.; Wang, Z.; Hu, Y.; et al. Highly pathogenic coronavirus N protein aggravates lung injury by MASP-2- mediated complement over-activation. medRevix 2020, 1–25. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.; Tortorici, M.A.; Wall, A.; Mcguire, A.T.; Veesler, D.; Walls, A.C.; Park, Y.; Tortorici, M.A.; Wall, A.; et al. Structure, Function, and Antigenicity of the SARS- Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 180, 1–12. [Google Scholar] [CrossRef]
- Channappanavar, R.; Fehr, A.R.; Vijay, R.; Mack, M.; Zhao, J.; Meyerholz, D.K.; Perlman, S. Dysregulated Type I Interferon and Inflammatory Monocyte-Macrophage Responses Cause Lethal Pneumonia in SARS-CoV-Infected Mice Article Dysregulated Type I Interferon and Inflammatory Monocyte-Macrophage Responses Cause Lethal Pneumonia in SARS-CoV-Infecte. Cell Host Microbe 2016, 19, 181–193. [Google Scholar] [CrossRef] [Green Version]
- Lu, G.; Wang, Q.; Gao, G.F. Bat-to-human: Spike features determining ‘host jump’ of coronaviruses SARS-CoV, MERS-CoV, and beyond. Trends Microbiol. 2015, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; He, Y.; Zhou, Y.; Liu, S.; Zheng, B.J. The spike protein of SARS-CoV—A target for vaccine and therapeutic development. Nature 2009, 7, 226–236. [Google Scholar] [CrossRef]
- Li, C.K.; Wu, H.; Yan, H.; Ma, S.; Wang, L.; Zhang, M.; Tang, X.; Temperton, N.J.; Weiss, R.A.; Brenchley, J.M.; et al. T Cell Responses to Whole SARS Coronavirus in Humans. J. Immunol. 2008, 181, 5490–5500. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.J.; Zhao, M.; Liu, K.; Xu, K.; Wong, G.; Tan, W.; Gao, G.F. T-cell immunity of SARS-CoV: Implications for vaccine development against MERS-CoV. Antiviral Res. 2017, 137, 82–92. [Google Scholar] [CrossRef]
- Buonavoglia, C.; Decaro, N.; Martella, V.; Elia, G.; Campolo, M.; Desario, C.; Castagnaro, M.; Tempesta, M. Canine Coronavirus Highly Pathogenic for Dogs. Emerg. Infect. Dis. 2006, 12, 49–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decaro, N.; Campolo, M.; Lorusso, A.; Desario, C.; Mari, V.; Loredana, M.; Elia, G.; Martella, V.; Buonavoglia, C. Experimental infection of dogs with a novel strain of canine coronavirus causing systemic disease and lymphopenia. Vet. Microbiol. 2008, 128, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Marinaro, M.; Mari, V.; Lucia, A.; Tarsitano, E.; Elia, G.; Losurdo, M.; Rezza, G.; Buonavoglia, C.; Decaro, N. Prolonged depletion of circulating CD4+ T lymphocytes and acute monocytosis after pantropic canine coronavirus infection in dogs. Virus Res. 2010, 152, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Decaro, N.; Buonavoglia, C. Canine Coronavirus: Not Only an Enteric Pathogen. Vet. Clin. Small Anim. 2011, 41, 1121–1132. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, D.; Tian, J.; Kang, H.; Guo, D.; Jiang, Q.; Liu, J.; Li, Z. Molecular characterization of HLJ-073, a recombinant canine coronavirus strain from China with an ORF3abc deletion. Arch. Virol. 2019, 164, 2159–2164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Licitra, B.N.; Whittaker, G.R.; Dubovi, E.J.; Duhamel, G.E. Genotypic Characterization of Canine Coronaviruses Associated with Fatal Canine Neonatal Enteritis in the United States. J. Clin. Microbiol. 2014, 52, 4230–4238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DE-Groot-Mijnes, J.D.F.; van Dun, J.M.; van der Most, R.G.; de Groot, R.J. Natural History of a Recurrent Feline Coronavirus Infection and the Role of Cellular Immunity in Survival and Disease. J. Virol. 2005, 79, 1036–1044. [Google Scholar] [CrossRef] [Green Version]
- Rottier, P.J.M.; Nakamura, K.; Schellen, P.; Volders, H.; Haijema, B.J. Acquisition of Macrophage Tropism during the Pathogenesis of Feline Infectious Peritonitis Is Determined by Mutations in the Feline Coronavirus Spike Protein. J. Virol. 2005, 79, 14122–14130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decaro, N.; Buonavoglia, C. An update on canine coronaviruses: Viral evolution and pathobiology. Vet. Microbiol. 2008, 132, 221–234. [Google Scholar] [CrossRef]
- Dunne, P.J.; Faint, J.M.; Gudgeon, N.H.; Fletcher, J.M.; Plunkett, F.J.; Soares, M.V.D.; Hislop, A.D.; Annels, N.E.; Rickinson, A.B.; Salmon, M.; et al. Epstein-Barr virus—Specific CD8+ T cells that re-express CD45RA are apoptosis-resistant memory cells that retain replicative potential. Blood 2002, 100, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Veazey, R.; Ling, B.; Pandrea, I.; Clure, H.M.C.; Lackner, A.; Marx, P. Decreased CCR5 Expression on CD4+ T Cells of SIV-Infected Sooty Mangabeys. AIDS Res. Hum. Retroviruses 2003, 19, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Fan, Y.; Wu, W.; Zhang, F.; Wang, J. Expression of Lymphocytes and Lymphocyte Subsets in Patients with Severe Acute Respiratory Syndrome. Br. Rep. 2003, 37, 857–859. [Google Scholar] [CrossRef] [PubMed]
- Roncati, L.; Gallo, G.; Manenti, A.; Palmieri, B. Renin-angiotensin system: The unexpected flaw inside the human immune system revealed by SARS-CoV-2. Med. Hypotheses 2020, 140. [Google Scholar] [CrossRef]
- Decaro, N.; Elia, G.; Martella, V.; Campolo, M.; Mari, V.; Desario, C.; Stella, M.; Lorusso, E.; Kanellos, T.; Gibbons, R.H.; et al. Immunity after natural exposure to enteric canine coronavirus does not provide complete protection against infection with the new pantropic CB/05 strain. Vaccine 2010, 28, 724–729. [Google Scholar] [CrossRef] [PubMed]
- Savino, W. The Thymus Is a Common Target Organ. PLoS Pathog. 2006, 2, 472–483. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.; Zhao, X.; Li, J.; Niu, P.; Yang, B.; Wu, H.; Wang, W.; Song, H.; Huang, B.; Zhu, N.; et al. Articles Genomic characterisation and epidemiology of 2019 novel coronavirus: Implications for virus origins and receptor binding. Lancet 2020, 395, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Faria, N.R. First Cases of Coronavirus Disease (COVID-19) in Brazil, South America (2 Genomes, 3rd March 2020). Available online: https://virological.org/t/first-cases-of-coronavirus-disease-covid-19-in-brazil-south-america-2-genomes-3rd-march-2020/409 (accessed on 4 September 2020).
- Hagen, J.B. The origins of bioinformatics. Nat. Rev. Genet. 2000, 1, 231–236. [Google Scholar] [CrossRef]
- Chatzou, M.; Magis, C.; Chang, J.M.; Kemena, C.; Bussotti, G.; Erb, I.; Notredame, C. Multiple sequence alignment modeling: Methods and applications. Brief. Bioinform. 2016, 17, 1009–1023. [Google Scholar] [CrossRef] [Green Version]
- Ceraolo, C.; Giorgi, F.M. Genomic variance of the 2019-nCoV coronavirus. J. Med. Virol. 2020, 92, 522–528. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Liu, Y.; Yang, Y.; Zhang, P.; Zhong, W.; Wang, Y.; Wang, Q.; Xu, Y.; Li, M.; Li, X.; et al. Analysis of therapeutic targets for SARS-CoV-2 and discovery of potential drugs by computational methods. Acta Pharm. Sin. B 2020. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990. [Google Scholar] [CrossRef]
- Xiao, K.; Zhai, J.; Feng, Y.; Zhou, N.; Zhang, X.; Zou, J.-J.; Li, N.; Guo, Y.; Li, X.; Shen, X.; et al. Isolation and Characterization of 2019-nCoV-like Coronavirus from Malayan Pangolins. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Andersen, K.G.; Rambaut, A.; Lipkin, W.I.; Holmes, E.C.; Garry, R.F. The proximal origin of SARS-CoV-2. Nat. Med. 2020, 89, 44–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eur. Surveill. 2020, 25, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustin, S.; Huggett, J. qPCR primer design revisited. Biomol. Detect. Quantif. 2017, 14, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Chuang, L.Y.; Cheng, Y.H.; Yang, C.H. Specific primer design for the polymerase chain reaction. Biotechnol. Lett. 2013, 31, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elsalam, K.A. Bioinformatic tools and guideline for PCR primer design. African J. Biotechnol. 2003, 2, 91–100. [Google Scholar]
- GISAID. GISAID Initiative. Available online: https://www.gisaid.org/ (accessed on 4 September 2020).
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T.L. Primer-BLAST: A tool to design target-specific primers for polymerase chain reaction. BMC Bioinformatics 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padhi, B.K.; Pelletier, G.; Shwed, P.S. A bioinformatics workflow for the evaluation of RT-qPCR primer specificity: Application for the assessment of gene expression data reliability in toxicological studies. Regul. Toxicol. Pharmacol. 2020, 111, 104575. [Google Scholar] [CrossRef]
- De Oliveira Diniz, M.; Souza Ferreira, L.C. Biotechnology applied to the development of vaccines. Estud. Avançados 2010, 24, 19–30. [Google Scholar]
- Grifoni, A.; Sidney, J.; Zhang, Y.; Scheuermann, R.H.; Peters, B.; Sette, A.; Grifoni, A.; Sidney, J.; Zhang, Y.; Scheuermann, R.H.; et al. A Sequence Homology and Bioinformatic Approach Can Predict Candidate Targets for Immune Responses to SARS-CoV-2. Cell Host Microbe 2020, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Baruah, V.; Bose, S. Immunoinformatics-aided identification of T cell and B cell epitopes in the surface glycoprotein of 2019-nCoV. J. Med. Virol. 2020, 495–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, S.F.; Quadeer, A.A.; McKay, M.R. Preliminary identification of potential vaccine targets for the COVID-19 Coronavirus (SARS-CoV-2) Based on SARS-CoV Immunological Studies. Viruses 2020, 12, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Everts, M.; Cihlar, T.; Bostwick, J.R.; Whitley, R.J. Accelerating Drug Development: Antiviral Therapies for Emerging Viruses as a Model. Annu. Rev. Pharmacol. Toxicol. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joshi, T.; Xu, D. Quantitative assessment of relationship between sequence similarity and function similarity. BMC Genomics 2007. [Google Scholar] [CrossRef] [Green Version]
- Mirza, M.U.; Froeyen, M. Structural elucidation of SARS-CoV-2 vital proteins: Computational methods reveal potential drug candidates against Main protease, Nsp12 RNA-dependent RNA polymerase and Nsp13 helicase. J. Pharm. Anal. 2020, 1–10. [Google Scholar] [CrossRef]
- Evans, G.; Axford, D.; Owen, R.L. The design of macromolecular crystallography diffraction experiments. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011. [Google Scholar] [CrossRef]
- Saibil, H.R. Macromolecular structure determination by cryo-electron microscopy. Acta Crystallogr. Sect. D Biol. Crystallogr. 2000. [Google Scholar] [CrossRef] [Green Version]
- Bryce, D.L.; Taulelle, F. NMR Crystallography. Acta Crystallogr. Sect. C Struct. Chem. 2017, 73, 126–127. [Google Scholar] [CrossRef] [Green Version]
- Burley, S.K.; Almo, S.C.; Bonanno, J.B.; Capel, M.; Chance, M.R.; Gaasterland, T.; Lin, D.; Šali, A.; Studier, F.W.; Swaminathan, S. Structural genomics: Beyond the Human Genome Project. Nat. Genet. 1999, 23, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Battistuz, T.; Bhat, T.N.; Bluhm, W.F.; Bourne, P.E.; Burkhardt, K.; Feng, Z.; Gilliland, G.L.; Iype, L.; Jain, S.; et al. The protein data bank. Acta Crystallogr. Sect. D Biol. Crystallogr. 2002. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro from COVID-19 virus and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Jedrzejczak, R.; Maltseva, N.I.; Endres, M.; Godzik, A.; Michalska, K.; Joachimiak, A. Crystal structure of Nsp15 endoribonuclease NendoU from SARS-CoV-2. Protein Sci. 2020, 29, 1596–1605. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q. Crystal structure of the 2019-nCoV spike receptor-binding domain bound with the ACE2 receptor. bioRxiv 2020, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [Green Version]
- Baker, M. Making membrane proteins for structures: A trillion tiny tweaks. Nat. Methods 2010, 7, 429–434. [Google Scholar] [CrossRef]
- Muhammed, M.T.; Aki-Yalcin, E. Homology modeling in drug discovery: Overview, current applications, and future perspectives. Chem. Biol. Drug Des. 2019, 93, 12–20. [Google Scholar] [CrossRef] [Green Version]
- Levoin, N.; Calmels, T.; Krief, S.; Danvy, D.; Berrebi-Bertrand, I.; Lecomte, J.M.; Schwartz, J.C.; Capet, M. Homology model versus X-ray structure in receptor-based drug design: A retrospective analysis with the dopamine D3 receptor. ACS Med. Chem. Lett. 2011. [Google Scholar] [CrossRef] [Green Version]
- Miguet, L.; Zhang, Z.; Barbier, M.; Grigorov, M.G. Comparison of a homology model and the crystallographic structure of human 11β-hydroxysteroid dehydrogenase type 1 (11βHSD1) in a structure-based identification of inhibitors. J. Comput. Aided. Mol. Des. 2006. [Google Scholar] [CrossRef]
- Uba, A.I.; Yelekçi, K. Crystallographic structure versus homology model: A case study of molecular dynamics simulation of human and zebrafish histone deacetylase 10. J. Biomol. Struct. Dyn. 2019, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Cui, H.; Gao, Z.; Liu, M.; Lu, S.; Mkandawire, W.; Narykov, O.; Sun, M.; Korkin, D. Structural Genomics of SARS-CoV-2 Indicates Evolutionary Conserved Functional Regions of Viral Proteins. Viruses 2020, 12, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shanker, A.; Bhanu, D.; Alluri, A. Whole Genome Sequence Analysis and Homology Modelling of a 3C Like Peptidase and a Non-Structural Protein 3 of the SARS-CoV-2 Shows Protein Ligand Interaction with an Aza-Peptide and a Noncovalent Lead Inhibitor with Possible Antiviral Properties. New J. Chem. 2020, 44, 9202–9212. [Google Scholar] [CrossRef]
- Qamar, M.T.; Alqahtani, S.M.; Alamri, M.A.; Chen, L.-L. Structural basis of SARS-CoV-2 3CL pro and anti-COVID-19 drug discovery from medicinal plants. J. Pharm. Anal. 2020, 1–26. [Google Scholar] [CrossRef]
- Ortega, J.T.; Serrano, M.L.; Pujol, F.H.; Rangel, H.R. Role of Changes in Sars-Cov-2 Spike Protein in the Interaction With the Human Ace2 Receptor: An in Silico Analysis. EXCLI J. 2020, 19, 410–417. [Google Scholar] [CrossRef]
- Lavecchia, A.; Giovanni, C. Virtual Screening Strategies in Drug Discovery: A Critical Review. Curr. Med. Chem. 2013. [Google Scholar] [CrossRef]
- Fan, J.; Fu, A.; Zhang, L. Progress in molecular docking. Quant. Biol. 2019, 7, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Irwin, J.J.; Shoichet, B.K. ZINC—A free database of commercially available compounds for virtual screening. J. Chem. Inf. Model. 2005. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2019 update: Improved access to chemical data. Nucleic Acids Res. 2019. [Google Scholar] [CrossRef] [Green Version]
- Ortega, J.T.; Serrano, M.L.; Pujol, F.H.; Rangel, H.R. Unrevealing Sequence and Structural Features of Novel Coronavirus Using in Silico Approaches: The Main Protease As Molecular Target. EXCLI J. 2020, 19, 400–409. [Google Scholar] [CrossRef]
- Koehn, F.E.; Carter, G.T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-h.; Wu, K.-l.; Zhang, X.; Deng, S.-q.; Peng, B. In silico screening of Chinese herbal medicines with the potential to directly inhibit 2019 novel coronavirus. J. Integr. Med. 2020, 18, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Gentile, D.; Patamia, V.; Scala, A.; Sciortino, M.T.; Piperno, A. Putative Inhibitors of SARS-CoV-2 Main Protease from A Library of Marine Natural Products: A Virtual Screening and Molecular Modeling Study. Mar. Drugs 2020, 18, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, H.; Li, J.; Xie, H.; Wang, Y. Review of drug repositioning approaches and resources. Int. J. Biol. Sci. 2018, 14, 1232–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senathilake, K.S.; Samarakoon, S.R.; Tennekoon, K.H. Virtual screening of inhibitors against spike glycoprotein of 2019 novel corona virus: A drug repurposing approach. Eur. PMC 2020, 1, 1–10. [Google Scholar] [CrossRef]
- Smith, M.; Smith, J.C. Repurposing Therapeutics for COVID-19: Supercomputer-Based Docking to the SARS-CoV-2 Viral Spike Protein and Viral Spike Protein-Human ACE2 Interface. ChemRxiv 2020. [Google Scholar] [CrossRef]
- Novick, P.A.; Ortiz, O.F.; Poelman, J.; Abdulhay, A.Y.; Pande, V.S. SWEETLEAD: An in silico database of approved drugs, regulated chemicals, and herbal isolates for computer-aided drug discovery. PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed]
- Elfiky, A.A. Anti-HCV, nucleotide inhibitors, repurposing against COVID-19. Life Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ostaszewski, M.; Mazein, A.; Gillespie, M.E.; Kuperstein, I.; Niarakis, A.; Hermjakob, H.; Pico, A.R.; Willighagen, E.L.; Evelo, C.T.; Hasenauer, J.; et al. COVID-19 Disease Map, building a computational repository of SARS-CoV-2 virus-host interaction mechanisms. Sci. Data 2020, 7, 1–4. [Google Scholar] [CrossRef]
- Forster, P.; Forster, L.; Renfrew, C.; Forster, M. Phylogenetic network analysis of SARS-CoV-2 genomes. PNAS 2020, 117, 9241–9243. [Google Scholar] [CrossRef] [Green Version]
- Adebali, O.; Bircan, A.; Çirci, D.; İşlek, B.; Kilinç, Z.; Selçuk, B.; Turhan, B. Phylogenetic analysis of SARS-CoV-2 genomes in Turkey. Turkish J. Biol. 2020, 44, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Fauver, J.; Petrone, M.; Hodcroft, E.; Shioda, K.; Ehrlich, H.; Watts, A.; Vogels, C.; Brito, A.; Alpert, T.; Muyombwe, A.; et al. Coast-to-coast spread of SARS-CoV-2 in the United States revealed by genomic epidemiology. medRxiv 2020, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Nie, Q.; Li, X.; Chen, W.; Liu, D.; Chen, Y.; Li, H.; Li, D.; Tian, M.; Tan, W.; Zai, J. Phylogenetic and phylodynamic analyses of SARS-CoV-2. Virus Res. 2020, 287, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Islam, R.; Parves, M.R.; Paul, A.S.; Uddin, N.; Rahman, M.S.; Mamun, A.A.; Hossain, M.N.; Ali, M.A.; Halim, M.A. A molecular modeling approach to identify effective antiviral phytochemicals against the main protease of SARS-CoV-2. J. Biomol. Struct. Dyn. 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fantini, J.; Di Scala, C.; Chahinian, H.; Yahi, N. Structural and molecular modelling studies reveal a new mechanism of action of chloroquine and hydroxychloroquine against SARS-CoV-2 infection. Int. J. Antimicrob. Agents 2020, 55, 105960. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.P.; Viswanathan, S.; Wang, M.; Sun, L.-Q.; Clark, G.C.; D’Elia, R.V. Current and future developments in the treatment of virus-induced hypercytokinemia. Future Med. Chem. 2017, 9, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Jamilloux, Y.; Henry, T.; Belot, A.; Viel, S.; Fauter, M.; El Jammal, T.; Walzer, T.; François, B.; Sève, P. Should we stimulate or suppress immune responses in COVID-19? Cytokine and anti-cytokine interventions. Autoimmun. Rev. 2020, 19, 102567. [Google Scholar] [CrossRef]
- Plante, J.A.; Fernandez, D.; Mirchandani, D.; Aguilar, P.V.; Sastry, K.J.; Newman, R.A.; Weaver, S.C.; Oncology, N.M. Prophylactic and Therapeutic Inhibition of In Vitro SARS-CoV-2 Replication by Oleandrin. bioRxiv 2020. [Google Scholar] [CrossRef]
- Su, H.; Yao, S.; Zhao, W.; Li, M.; Liu, J.; Shang, W.; Xie, H.; Ke, C.; Hu, H.; Gao, M.; et al. Anti-SARS-CoV-2 activities in vitro of Shuanghuanglian preparations and bioactive ingredients. Acta Pharmacol. Sin. 2020. [Google Scholar] [CrossRef]
- Deftereos, S.G.; Giannopoulos, G.; Vrachatis, D.A.; Siasos, G.D.; Giotaki, S.G.; Gargalianos, P.; Metallidis, S.; Sianos, G.; Baltagiannis, S.; Panagopoulos, P.; et al. Effect of Colchicine vs Standard Care on Cardiac and Inflammatory Biomarkers and Clinical Outcomes in Patients Hospitalized With Coronavirus Disease 2019. JAMA Netw. Open 2020, 3, e2013136. [Google Scholar] [CrossRef]
- Scarsi, M.; Piantoni, S.; Colombo, E.; Airó, P.; Richini, D.; Miclini, M.; Bertasi, V.; Bianchi, M.; Bottone, D.; Civelli, P.; et al. Association between treatment with colchicine and improved survival in a single-centre cohort of adult hospitalised patients with COVID-19 pneumonia and acute respiratory distress syndrome. Ann. Rheum. Dis. 2020, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Pruijssers, A.J.; George, A.S.; Schäfer, A.; Leist, S.R.; Gralinksi, L.E.; Dinnon, K.H.; Yount, B.L.; Agostini, M.L.; Stevens, L.J.; Chappell, J.D.; et al. Remdesivir Inhibits SARS-CoV-2 in Human Lung Cells and Chimeric SARS-CoV Expressing the SARS-CoV-2 RNA Polymerase in Mice. Cell Rep. 2020, 32, 107940. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, D.; Du, G.; Du, R.; Zhao, J.; Jin, Y.; Fu, S.; Gao, L.; Cheng, Z.; Lu, Q.; et al. Remdesivir in adults with severe COVID-19: A randomised, double-blind, placebo-controlled, multicentre trial. Lancet 2020, 395, 1569–1578. [Google Scholar] [CrossRef]
- Antinori, S.; Cossu, M.V.; Ridolfo, A.L.; Rech, R.; Bonazzetti, C.; Pagani, G.; Gubertini, G.; Coen, M.; Magni, C.; Castelli, A.; et al. Compassionate remdesivir treatment of severe Covid-19 pneumonia in intensive care unit (ICU) and Non-ICU patients: Clinical outcome and differences in post-treatment hospitalisation status. Pharmacol. Res. 2020, 158, 104899. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Wang, Y.; Wen, D.; Liu, W.; Wang, J.; Fan, G.; Ruan, L.; Song, B.; Cai, Y.; Wei, M.; et al. A Trial of Lopinavir–Ritonavir in Adults Hospitalized with Severe Covid-19. N. Engl. J. Med. 2020, 382, 1787–1799. [Google Scholar] [CrossRef] [PubMed]
- Hraiech, S.; Bourenne, J.; Kuteifan, K.; Helms, J.; Carvelli, J.; Gainnier, M.; Meziani, F.; Papazian, L. Lack of viral clearance by the combination of hydroxychloroquine and azithromycin or lopinavir and ritonavir in SARS-CoV-2-related acute respiratory distress syndrome. Ann. Intensive Care 2020, 10, 63. [Google Scholar] [CrossRef]
- Hung, I.F.-N.; Lung, K.-C.; Tso, E.Y.-K.; Liu, R.; Chung, T.W.-H.; Chu, M.-Y.; Ng, Y.-Y.; Lo, J.; Chan, J.; Tam, A.R.; et al. Triple combination of interferon beta-1b, lopinavir–ritonavir, and ribavirin in the treatment of patients admitted to hospital with COVID-19: An open-label, randomised, phase 2 trial. Lancet 2020, 395, 1695–1704. [Google Scholar] [CrossRef]
- Dastan, F.; Nadji, S.A.; Saffaei, A.; Marjani, M.; Moniri, A.; Jamaati, H.; Hashemian, S.M.; Baghaei, P.; Abedini, A.; Varahram, M.; et al. Subcutaneous administration of interferon beta-1a for COVID-19: A non-controlled prospective trial. Int. Immunopharmacol. 2020, 85, 106688. [Google Scholar] [CrossRef]
- Tang, W.; Cao, Z.; Han, M.; Wang, Z.; Chen, J.; Sun, W.; Wu, Y.; Xiao, W.; Liu, S.; Chen, E.; et al. Hydroxychloroquine in patients with mainly mild to moderate coronavirus disease 2019: Open label, randomised controlled trial. BMJ 2020, 369, m1849. [Google Scholar] [CrossRef]
- Chen, J.; Liu, D.; Liu, L.; Liu, P.; Xu, Q.; Xia, L.; Ling, Y.; Huang, D.; Song, S.; Zhang, D.; et al. A pilot study of hydroxychloroquine in treatment of patients with common coronavirus disease-19 (COVID-19). J. Zhejiang Univ. 2020, 49, 1–10. [Google Scholar]
- Gautret, P.; Lagier, J.-C.; Parola, P.; Hoang, V.T.; Meddeb, L.; Mailhe, M.; Doudier, B.; Courjon, J.; Giordanengo, V.; Vieira, V.E.; et al. Hydroxychloroquine and azithromycin as a treatment of COVID-19: Results of an open-label non-randomized clinical trial. Int. J. Antimicrob. Agents 2020, 56, 105949. [Google Scholar] [CrossRef] [PubMed]
- Arshad, S.; Kilgore, P.; Chaudhry, Z.S.; Jacobsen, G.; Wang, D.D.; Huitsing, K.; Brar, I.; Alangaden, G.J.; Ramesh, M.S.; McKinnon, J.E.; et al. Treatment with hydroxychloroquine, azithromycin, and combination in patients hospitalized with COVID-19. Int. J. Infect. Dis. 2020, 97, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Ye, F.; Zhang, M.; Cui, C.; Huang, B.; Niu, P.; Liu, X.; Zhao, L.; Dong, E.; Song, C.; et al. In Vitro Antiviral Activity and Projection of Optimized Dosing Design of Hydroxychloroquine for the Treatment of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). Clin. Infect. Dis. 2020, 71, 732–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Cao, R.; Xu, M.; Wang, X.; Zhang, H.; Hu, H.; Li, Y.; Hu, Z.; Zhong, W.; Wang, M. Hydroxychloroquine, a less toxic derivative of chloroquine, is effective in inhibiting SARS-CoV-2 infection in vitro. Cell Discov. 2020, 6, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andreani, J.; Le Bideau, M.; Duflot, I.; Jardot, P.; Rolland, C.; Boxberger, M.; Wurtz, N.; Rolain, J.-M.; Colson, P.; La Scola, B.; et al. In vitro testing of combined hydroxychloroquine and azithromycin on SARS-CoV-2 shows synergistic effect. Microb. Pathog. 2020, 145, 104228. [Google Scholar] [CrossRef] [PubMed]
- Million, M.; Lagier, J.-C.; Gautret, P.; Colson, P.; Fournier, P.-E.; Amrane, S.; Hocquart, M.; Mailhe, M.; Esteves-Vieira, V.; Doudier, B.; et al. Early treatment of COVID-19 patients with hydroxychloroquine and azithromycin: A retrospective analysis of 1061 cases in Marseille, France. Travel Med. Infect. Dis. 2020, 35, 101738. [Google Scholar] [CrossRef]
- Pothen, L.; Yildiz, H.; De Greef, J.; Penaloza, A.; Beauloye, C.; Belkhir, L.; Yombi, J.C. Safe use of hydroxychloroquine and its combination with azithromycin in the context of Sars-CoV-2 outbreak: Clinical experience in a Belgian tertiary center. Travel Med. Infect. Dis. 2020, 36, 101788. [Google Scholar] [CrossRef]
- Lagier, J.-C.; Million, M.; Gautret, P.; Colson, P.; Cortaredona, S.; Giraud-Gatineau, A.; Honoré, S.; Gaubert, J.-Y.; Fournier, P.-E.; Tissot-Dupont, H.; et al. Outcomes of 3,737 COVID-19 patients treated with hydroxychloroquine/azithromycin and other regimens in Marseille, France: A retrospective analysis. Travel Med. Infect. Dis. 2020, 36, 101791. [Google Scholar] [CrossRef]
- Borba, M.G.S.; Val, F.F.A.; Sampaio, V.S.; Alexandre, M.A.A.; Melo, G.C.; Brito, M.; Mourão, M.P.G.; Brito-Sousa, J.D.; Baía-da-Silva, D.; Guerra, M.V.F.; et al. Effect of High vs Low Doses of Chloroquine Diphosphate as Adjunctive Therapy for Patients Hospitalized With Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Infection. JAMA Netw. Open 2020, 3, e208857. [Google Scholar] [CrossRef] [Green Version]
- Caly, L.; Druce, J.D.; Catton, M.G.; Jans, D.A.; Wagstaff, K.M. The FDA-approved drug ivermectin inhibits the replication of SARS-CoV-2 in vitro. Antiviral Res. 2020, 178, 104787. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kiso, M.; Sakai-Tagawa, Y.; Iwatsuki-Horimoto, K.; Imai, M.; Takeda, M.; Kinoshita, N.; Ohmagari, N.; Gohda, J.; Semba, K.; et al. The Anticoagulant Nafamostat Potently Inhibits SARS-CoV-2 S Protein-Mediated Fusion in a Cell Fusion Assay System and Viral Infection In Vitro in a Cell-Type-Dependent Manner. Viruses 2020, 12, 629. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, M.; Toyama, M.; Baba, M. The chemokine receptor antagonist cenicriviroc inhibits the replication of SARS-CoV-2 in vitro. Antiviral Res. 2020, 182, 104902. [Google Scholar] [CrossRef] [PubMed]
- Cingolani, A.; Tummolo, A.M.; Montemurro, G.; Gremese, E.; Larosa, L.; Cipriani, M.C.; Pasciuto, G.; Liperoti, R.; Murri, R.; Pirronti, T.; et al. Baricitinib as rescue therapy in a patient with COVID-19 with no complete response to sarilumab. Infection 2020, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Liu, Y.; Qiu, L.; Liu, X.; Liu, D.; Li, J. Tocilizumab treatment in COVID-19: A single center experience. J. Med. Virol. 2020, 92, 814–818. [Google Scholar] [CrossRef] [PubMed]
- Radbel, J.; Narayanan, N.; Bhatt, P.J. Use of Tocilizumab for COVID-19-Induced Cytokine Release Syndrome. Chest 2020, 158, e15–e19. [Google Scholar] [CrossRef]
- Sinha, P.; Mostaghim, A.; Bielick, C.G.; McLaughlin, A.; Hamer, D.H.; Wetzler, L.; Bhadelia, N.; Fagan, M.; Linas, B.P.; Assoumou, S.A.; et al. Early administration of Interleukin-6 inhibitors for patients with severe Covid-19 disease is associated with decreased intubation, reduced mortality, and increased discharge. Int. J. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Xu, X.; Han, M.; Li, T.; Sun, W.; Wang, D.; Fu, B.; Zhou, Y.; Zheng, X.; Yang, Y.; Li, X.; et al. Effective treatment of severe COVID-19 patients with tocilizumab. Proc. Natl. Acad. Sci. USA 2020, 117, 10970–10975. [Google Scholar] [CrossRef]
- Toniati, P.; Piva, S.; Cattalini, M.; Garrafa, E.; Regola, F.; Castelli, F.; Franceschini, F.; Airò, P.; Bazzani, C.; Beindorf, E.-A.; et al. Tocilizumab for the treatment of severe COVID-19 pneumonia with hyperinflammatory syndrome and acute respiratory failure: A single center study of 100 patients in Brescia, Italy. Autoimmun. Rev. 2020, 19, 102568. [Google Scholar] [CrossRef]
- Sciascia, S.; Aprà, F.; Baffa, A.; Baldovino, S.; Boaro, D.; Boero, R.; Bonora, S.; Calcagno, A.; Cecchi, I.; Cinnirella, G.; et al. Pilot prospective open, single-arm multicentre study on off-label use of tocilizumab in patients with severe COVID-19. Clin. Exp. Rheumatol. 2020, 38, 529–532. [Google Scholar]
- Berlot, G.; Tomasini, A.; Roman Pognuz, E.; Randino, A.; Chiella, F.; La Fata, C.; Piva, M.; Amato, P.; di Maso, V.; Bianco, F.; et al. The Combined Use of Tocilizumab and Hemoadsorption in a Patient with SARS-COV-2-19-Associated Pneumonia: A Case Report. Nephron 2020, 144, 459–462. [Google Scholar] [CrossRef]
- Della-Torre, E.; Campochiaro, C.; Cavalli, G.; De Luca, G.; Napolitano, A.; La Marca, S.; Boffini, N.; Da Prat, V.; Di Terlizzi, G.; Lanzillotta, M.; et al. Interleukin-6 blockade with sarilumab in severe COVID-19 pneumonia with systemic hyperinflammation: An open-label cohort study. Ann. Rheum. Dis. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Benucci, M.; Giannasi, G.; Cecchini, P.; Gobbi, F.L.; Damiani, A.; Grossi, V.; Infantino, M.; Manfredi, M. COVID-19 pneumonia treated with Sarilumab: A clinical series of eight patients. J. Med. Virol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Filocamo, G.; Mangioni, D.; Tagliabue, P.; Aliberti, S.; Costantino, G.; Minoia, F.; Bandera, A. Use of anakinra in severe COVID-19: A case report. Int. J. Infect. Dis. 2020, 96, 607–609. [Google Scholar] [CrossRef] [PubMed]
- Dimopoulos, G.; de Mast, Q.; Markou, N.; Theodorakopoulou, M.; Komnos, A.; Mouktaroudi, M.; Netea, M.G.; Spyridopoulos, T.; Verheggen, R.J.; Hoogerwerf, J.; et al. Favorable Anakinra Responses in Severe Covid-19 Patients with Secondary Hemophagocytic Lymphohistiocytosis. Cell Host Microbe 2020, 28, 117–123.e1. [Google Scholar] [CrossRef] [PubMed]
- Cavalli, G.; De Luca, G.; Campochiaro, C.; Della-Torre, E.; Ripa, M.; Canetti, D.; Oltolini, C.; Castiglioni, B.; Tassan Din, C.; Boffini, N.; et al. Interleukin-1 blockade with high-dose anakinra in patients with COVID-19, acute respiratory distress syndrome, and hyperinflammation: A retrospective cohort study. Lancet Rheumatol. 2020, 2, e325–e331. [Google Scholar] [CrossRef]
- Dolinger, M.T.; Person, H.; Smith, R.; Jarchin, L.; Pittman, N.; Dubinsky, M.C.; Lai, J. Pediatric Crohn Disease and Multisystem Inflammatory Syndrome in Children (MIS-C) and COVID-19 Treated With Infliximab. J. Pediatr. Gastroenterol. Nutr. 2020, 71, 153–155. [Google Scholar] [CrossRef]
- Diurno, F.; Numis, F.G.; Porta, G.; Cirillo, F.; Maddaluno, S.; Ragozzino, A.; Negri, P.D.E.; Gennaro, C.D.I.; Pagano, A.; Allegorico, E.; et al. Eculizumab treatment in patients with COVID-19: Preliminary results from real life ASL Napoli 2 Nord experience. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 4040–4047. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
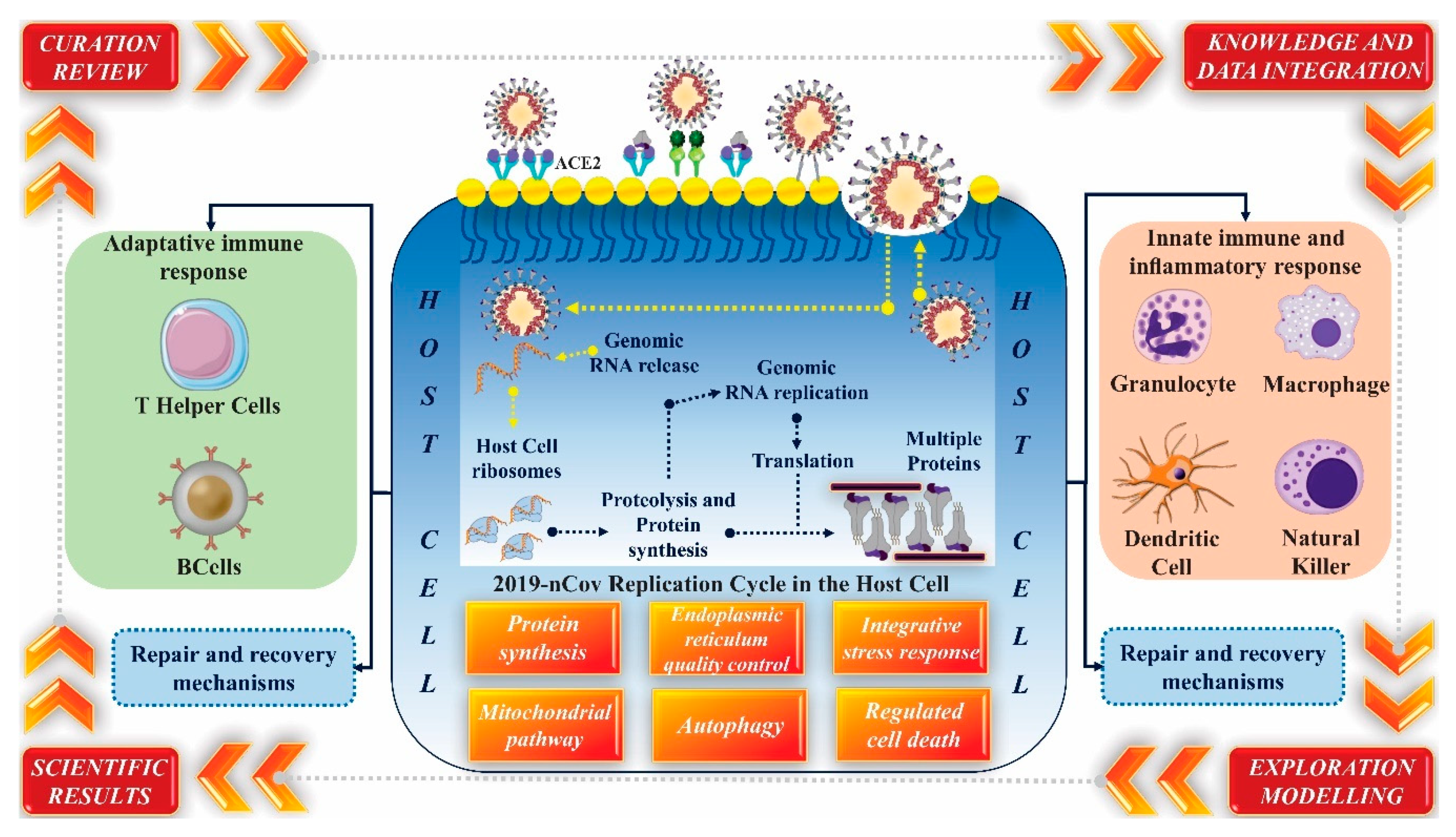{kind=link}
| Origin | Species | Compounds | Molecular Structures | Model Used | Doses and Route of Administration | Duration | Results | Ref. |
|---|---|---|---|---|---|---|---|---|
| Plant | Nerium oleander | Oleandrin |  | Vero cells | 0.05 μg/mL | 48 h | Viral replication reduction up to 78 times | [146] |
| 0.1 μg/mL | Viral replication reduction up to 100 times | |||||||
| Plant | Lonicera japonica | Chlorogenic acid |  | VeroE6 cells | 76.4 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | [147] |
| 39.48 µM | IC50 for 3CLpro of SARS-CoV-2 | |||||||
| 20.2 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Neochlorogenic acid |  | VeroE6 cells | 49.2 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 10.4 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Isochlorogenic acid A |  | VeroE6 cells | 77.0 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 18.9 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Isochlorogenic acid B |  | VeroE6 cells | 52.4 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 26.3 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Isochlorogenic acid C |  | VeroE6 cells | 78.2 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 18.4 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| 1,3-Dicaffeoylquinic acid |  | VeroE6 cells | 87.3 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 27.8 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Luteoloside |  | VeroE6 cells | 65.4 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 14.8 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Plant | Scutellaria baicalensis | Baicalin |  | VeroE6 cells | 97.6 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | [147] |
| 6.41 µM | IC50 for 3CLpro of SARS-CoV-2 | |||||||
| 68.9 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Baicalein |  | VeroE6 cells | 99.4 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 0.94 µM | IC50 for 3CLpro of SARS-CoV-2 | |||||||
| 87.0 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Scutellarein |  | VeroE6 cells | 101.6 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 3.02 µM | IC50 for 3CLpro of SARS-CoV-2 | |||||||
| 90.7 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Scutellarin |  | VeroE6 cells | 76.8 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 18.9 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Chrysin-7-O-β-D-glucuronide |  | VeroE6 cells | 50.6 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 24.2 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Plant | Forsythia suspensa | Forsythoside A |  | VeroE6 cells | 95.3 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | [147] |
| 3.18 µM | IC50 for 3CLpro of SARS-CoV-2 | |||||||
| 70.5 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Forsythoside B |  | VeroE6 cells | 101.4 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 2.88 µM | IC50 for 3CLpro of SARS-CoV-2 | |||||||
| 80.9 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Forsythoside E |  | VeroE6 cells | 96.6 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 6.68 µM | IC50 for 3CLpro of SARS-CoV-2 | |||||||
| 41.9 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Forsythoside H |  | VeroE6 cells | 99.3 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 10.17 µM | IC50 for 3CLpro of SARS-CoV-2 | |||||||
| 61.7 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Forsythoside I |  | VeroE6 cells | 95.9 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 5.47 µM | IC50 for 3CLpro of SARS-CoV-2 | |||||||
| 46.3 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Isoforsythiaside |  | VeroE6 cells | 94.4 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 5.85 µM | IC50 for 3CLpro of SARS-CoV-2 | |||||||
| 46.8 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Acteoside |  | VeroE6 cells | 97.0 µM | 48 h | 100% inhibition of SARS-CoV-2 3CLpro | |||
| 34.6 µM | 10% inhibition of SARS-CoV-2 3CLpro | |||||||
| Fungus | Colletotrichum gloeosporioides | Phillyrin |  | VeroE6 cells | 10 µM | 48 h | 7,8% inhibition of SARS-CoV-2 3CLpro |
| Compounds | Molecular Structures | Model Used | Doses and Route of Administration | Duration | Results | References |
|---|---|---|---|---|---|---|
| Colchicine |  | Humans | 1.5 mg followed by 0.5 mg after 60 min and maintenance doses of 0.5 mg twice daily | 3 weeks | It had significantly improved in the time of clinical deterioration | [148] |
| 1 mg/day | 21 days | Best survival rate | [149] | |||
| Remdesivir |  | Vero E6 cells | 1.65 μM | 48 h | Reduced inhibitory activity on viral replication | [150] |
| Calu3 2B4 human lung cells | 0.28 μM | 72 h | Strong inhibition of SARS-CoV-2 replication | |||
| Human airway epithelial cells | 0.01 μM | 48 h | Strong inhibition of SARS-CoV-2 replication | |||
| Mice | 25 mg/kg | 5 days | Drastic reduction of viral load in the lung | |||
| Humans | 200 mg on day 1 followed by 100 mg on days 2 to 10, intravenously | 10 days | No statistically significant clinical benefits were observed | [151] | ||
| Humans | Clinical improvement and fewer adverse effects | [152] | ||||
| Lopinavir + Ritonavir * |  | Humans | 400 mg + 100 mg | 14 days | No benefit was seen | [153] |
| Humans | 800 mg per day | 6 days | Absence of viral clearance | [154] | ||
| IFN-β-1b + Lopinavir + Ritonavir + Ribavirin * |  | Humans | 8 million international units + 400 mg + 100 mg + 400 mg | 14 days | Relief of symptoms and reduction of hospital stay | [155] |
| IFN-β-1a | * | Humans | 44 µg subcutaneously every two days | 10 days | Improved viral clearance and faster recovery speed | [156] |
| Hydroxychloroquine |  | Humans | 1200 mg daily for three days and a maintenance dose of 800 mg daily | Two or three weeks depending on the severity of the condition | No virus elimination benefits were observed | [157] |
| Humans | 400 mg per day | 5 days | Improved patient prognosis | [158] | ||
| Humans | 600 mg per day | 6 days | Reduction/disappearance of viral load | [159] | ||
| Humans | 400 mg HCQ twice daily on day 1, followed by 200 mg twice daily on days 2 to 5 | 5 days | Significant reduction in mortality | [160] | ||
| Vero cells | 0.72 μM | 48 h | Significant antiviral activity | [161] | ||
| 6.14 μM | 24 h | Insignificant antiviral activity | ||||
| VeroE6 cells | 4.06 μM | 48 h | Significant antiviral activity | [162] | ||
| Azithromycin |  | Humans | 500 mg of AZTM once daily on day 1, followed by 250 mg once daily for the next 4 days | 5 days | No benefit was seen | [160] |
| Hydroxychloroquine + Azithromycin * |  | Vero E6 | 1 μM of HCQ + 5 μM of AZTM or + 10 μM of AZTM | 60h | No significant results were observed | [163] |
| 2 μM of HCQ + 5 μM of AZTM or + 10 μM of AZTM | ||||||
| 5 μM of HCQ + 5 μM of AZTM | Relative viral inhibition of 97.5% and 99.1% | |||||
| 5 μM of HCQ + 10 μM of AZTM | ||||||
| Humans | 400 mg HCQ × 2 on day 1, followed by 200 mg × 2 on days 2 to 5 + 500 mg of AZTM on day 1, followed by 250 mg the next 4 days | 5 days | Significant reduction in mortality | [160] | ||
| Humans | 200 mg HCQ × 3/day + 500 mg of AZTM on day 1 followed by 250 mg/day the next four days | HCQ for ten days + AZTM for 5 days | Administration prior to the occurrence of complications was associated with a reduction in the mortality rate | [164] | ||
| Humans | 600 mg HCQ + 500 mg AZTM, followed by 250 mg AZTM on the following days | 6 days | Absence of viral clearance | [154] | ||
| Humans | 400 mg HCQ, twice on day 1, followed by 200 mg twice for the next 4 days + 500 mg of AZTM for 3 days | HCQ for 5 days + AZTM for 3 days | No security problems were found using the combination | [165] | ||
| Humans | 200 mg HCQ × 3/day + 500 mg AZTM on day 1, followed by 250 mg for the next 4 days orally | HCQ for 10 days + AZTM for 5 days | Improvement in clinical results and faster reduction of viral load | [166] | ||
| Chloroquine Diphosphate |  | Humans | 600 mg twice daily | 10 days | Higher lethality rate | [167] |
| 450 mg twice daily | 4 days | No benefit was seen | ||||
| Vero cells | 5.47 μM | 48 h | Insignificant antiviral activity | [161] | ||
| 23.90 μM | 24 h | |||||
| VeroE6 cells | 2.71 μM | 48 h | Insignificant antiviral activity | [162] | ||
| Ivermectin |  | Vero-hSLAM cells | 5 μM | 48 h | Reduction of viral replication approximately 5000 times | [168] |
| Nafamostat mesylate |  | Calu-3 cells | 10 nM | 5 days | Potent inhibition of SARS-CoV-2 fusion mediated by protein S and consequent inhibition of infection | [169] |
| VeroE6/TMPRSS2 cells | 30 μM | 3 days | ||||
| Cenicriviroc |  | VeroE6/TMPRSS2 cells | 40 μM | 3 days | Inhibition of viral replication and control of excessive immune response | [170] |
| Baricitinib |  | Humans | 4 mg per day | 2 weeks | Reduction of inflammatory indices and clinical improvement | [171] |
| Compounds | Model Used | Doses and Route of Administration | Duration | Results | References |
|---|---|---|---|---|---|
| Tocilizumab | Humans | 80 to 600 mg, depending on the severity of the condition | 38 days | Reduces risk of cytokine storms in patients with severe COVID-19 | [172] |
| 400 mg intravenously | 3 days | Cytokine release syndrome even after treatment | [173] | ||
| 560 mg for 2 days, followed by 700 mg the next day intravenously | |||||
| 400 mg | _ | Clinical improvement and lower mortality | [174] | ||
| 400 mg to 800 mg according to the severity of the condition, intravenously | Single dose, 1 h infusion | Immediate improvement of symptoms and normalization of inflammatory indices after 5 days | [175] | ||
| 8 mg/kg intravenously every 12 h and a third infusion 24 h later | 2 days | Improvement of clinical and respiratory condition | [176] | ||
| 8 mg/kg intravenously | 6 days | Improvement in respiratory and laboratory parameters | [177] | ||
| 8 mg/kg once daily | 2 days | The treatment associated with hemoadsorption, improved gas exchange and reduced levels of inflammatory mediators | [178] | ||
| Sarilumab | Humans | 400 mg intravenously | 10 days | Treatment was associated with faster recovery | [179] |
| 5 days | Reduction of inflammation and rapid recovery | [180] | |||
| 200 mg | _ | Clinical improvement and lower mortality | [174] | ||
| Anakinra | Humans | 200 mg intravenously, followed by 100 mg every 6 h subcutaneously | _ | Progressive improvement in respiratory function and Marked reduction in inflammatory markers | [181] |
| 200 mg every 8 h intravenously | 7 days | Improved respiratory function | [182] | ||
| 300 mg once daily intravenously, followed by 100 mg once daily subcutaneously | 300 mg for 4 days + 100 mg until hospital discharge | ||||
| 5 mg/kg twice a day intravenously | 10 days | Reduction of systemic inflammation and progressive improvement in respiratory function | [183] | ||
| 100 mg twice daily subcutaneously | 7 days | Absence of significant clinical or anti-inflammatory effects | |||
| Infliximab | Humans | 10 mg/kg | 2 days | Interruption of the systemic inflammatory response in a patient with Crohn’s disease and COVID-19 | [184] |
| Eculizumab | Humans | 4 weekly infusions of 900 mg | 4 weeks | Clinical improvement in the first 48 h after the first administration | [185] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villas-Boas, G.R.; Rescia, V.C.; Paes, M.M.; Lavorato, S.N.; Magalhães-Filho, M.F.d.; Cunha, M.S.; Simões, R.d.C.; Lacerda, R.B.d.; Freitas-Júnior, R.S.d.; Ramos, B.H.d.S.; et al. The New Coronavirus (SARS-CoV-2): A Comprehensive Review on Immunity and the Application of Bioinformatics and Molecular Modeling to the Discovery of Potential Anti-SARS-CoV-2 Agents. Molecules 2020, 25, 4086. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25184086
Villas-Boas GR, Rescia VC, Paes MM, Lavorato SN, Magalhães-Filho MFd, Cunha MS, Simões RdC, Lacerda RBd, Freitas-Júnior RSd, Ramos BHdS, et al. The New Coronavirus (SARS-CoV-2): A Comprehensive Review on Immunity and the Application of Bioinformatics and Molecular Modeling to the Discovery of Potential Anti-SARS-CoV-2 Agents. Molecules. 2020; 25(18):4086. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25184086
Chicago/Turabian StyleVillas-Boas, Gustavo R., Vanessa C. Rescia, Marina M. Paes, Stefânia N. Lavorato, Manoel F. de Magalhães-Filho, Mila S. Cunha, Rafael da C. Simões, Roseli B. de Lacerda, Renilson S. de Freitas-Júnior, Bruno H. da S. Ramos, and et al. 2020. "The New Coronavirus (SARS-CoV-2): A Comprehensive Review on Immunity and the Application of Bioinformatics and Molecular Modeling to the Discovery of Potential Anti-SARS-CoV-2 Agents" Molecules 25, no. 18: 4086. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25184086