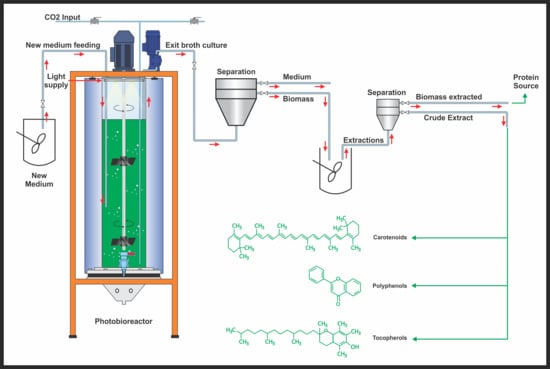
2.1. Growth Analyses in Batch and Continuous Cultivation
The growth profiles were analyzed in the batch and continuous cultures of four microalgae. The growth performance of four microalgae in batch cultivation is shown in
Figure 1. Specific growth rates were calculated by the Gompertz growth equation, which were found to be 0.53, 0.49, 0.51, and 1.1 d
−1 for
Spirulina platensis,
Isochrysis galbana,
Tetraselmis suecica, and
Porphyridium cruentum, respectively. Biomass concentration and biomass productivity values are summarized in
Table 1. The biomass concentrations in batch cultures were 3, 1.5, 1.8, 2.1 g L
−1 for
S. platensis,
I. galbana,
T. suecica, and
P. cruentum, respectively (
Table 1). The growth of microalgae depends on the culture conditions such as medium composition, hydrodynamics in the bioreactor, and light irradiation that reaches every cell. By comparing all data of the Gompertz growth parameters among all microalgae, μ (growth specific rate) and maximum biomass density were different for all strains (
Table 1).
The batch cultures reached maximum cell concentrations over nine days of cultivation; maximum cell concentration indicates the change from batch to continuous cultivation. The dilution rate, which corresponds to the growth rate in continuous culture, was fixed at 0.6 d
−1 for
S. platensis,
I. galbana,
T. suecica, and
P. cruentum. Different light intensities were supplied (16.27, 21.26, 30.42, and 39.63 kJ d
−1). The biomass concentrations of the four microalgae increased as light intensity increased in continuous culture (
Table 2). The biomass concentration under all light intensities in continuous cultures were significantly different for all microalgae used. Biomass concentration in descending order at a light intensity of 39.63 kJ d
−1 was: 3.35 > 2.63 > 1.93 > 1.58 g biomass L
−1 for
S. platensis,
P. cruentum,
T. suecica, and
I. galbana, respectively (
Table 2), in comparison with biomass in bath cultures: 3.0 > 2.1 > 1.8 > 1.5 g L
−1, respectively, for the same light irradiance (39.63 kJ d
−1).
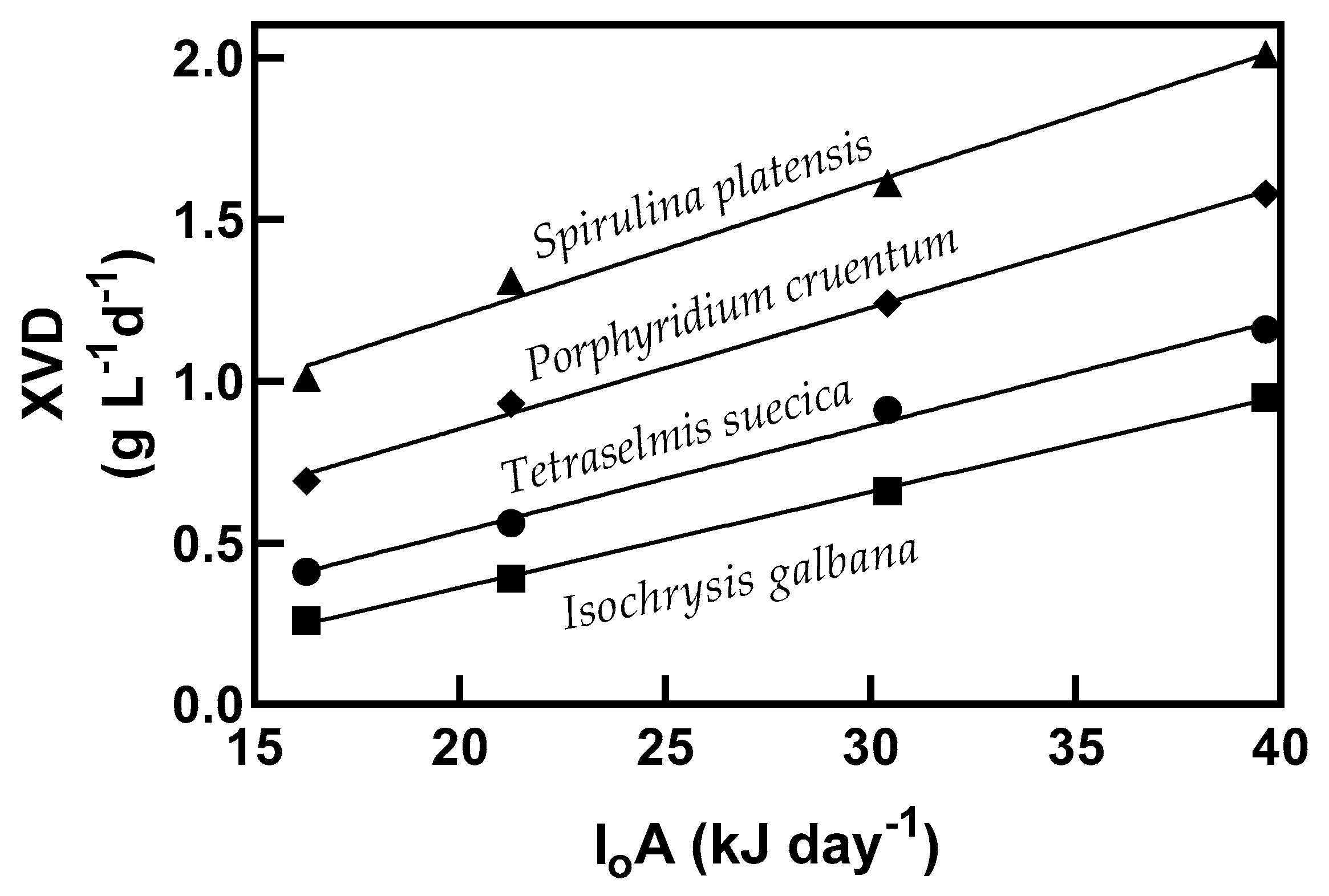
Biomass productivity (XVD (g biomass L
−1 d
−1), where X is biomass (g), V is working volume (L
−1), and D is dilution rate (d
−1)) obtained in terms of total light supplied (I
0A (kJ d
−1) energy supplied from light to a surface area per unit of time) is shown in
Figure 2. The productivity of biomass at a light intensity of 39.63 kJ d
−1 in batch culture was 0.34, 0.24, 0.2, and 0.16 g biomass L
−1 d
−1 (
Table 1) and 2.01, 1.58, 1.16, and 0.95, g biomass L
−1 d
−1, for
S. platensis,
P. cruentum,
T. suecica, and
I. galbana, respectively, in continuous cultivation (
Figure 2). Comparison of the biomass productivity (XVD) though linear regression analysis of each microalga showed statistically significant differences with increasing light intensity. The productivity increase of biomass concentration was 5.9–6.6 times higher in continuous cultures, which may support the final quantity of antioxidants produced, as shown below. The energetic yield of the produced biomass for the supplied light energy varied according to the strain: 0.041, 0.037, 0.033, and 0.030 g biomass produced per kJ for
S. platensis,
P. cruentum,
T. suecica, and
I. galbana, respectively, and a yield of conversion efficiency (Y
ce, luminous energy to chemical energy) of 1.04, 0.95, 0.83, and 0.75, respectively (–ΔHo × Y
kcal; enthalpy change for O
2 = 106 kcal mol
−1 O
2).
2.2. Antioxidant Production in Batch Culture
Total phenols, alkaloids, and terpenoids content were measured in
S. platensis,
I. galbana, and
T. suecica during batch cultivation (
Table 3). All compounds were produced by the microalgae in significantly different amounts (
p < 0.05), except for alkaloids between
I. galbana and
T. suecica, where no significant differences were observed. Phenol, alkaloids, and terpenoids were more abundant in
S. platensis than the others; these compounds accumulated during growth.
The productivities of phenols, alkaloids, and terpenoids showed significant differences among the three species (
Figure 3). The productivity of phenols in
S. platensis was four times that of
I. galbana and 1.75 times that of
T. suecica (
Figure 3). Alkaloids and terpenoids also showed significant differences among the three species of microalgae.
Tocopherols (α, δ, and γ), carotenoids, superoxide dismutase (SOD), and phycobiliproteins were recorded during batch production in the four microalgae (
Table 4). The total tocopherol amount is reported here as the sum of α, δ, and γ tocopherols in
I. galbana,
T. suecica, as well as the sum of α and γ tocopherols in
S. platensis and
P. cruentum. In
S. platensis and
P. cruentum, δ-tocopherol was not found. A large amount of tocopherol was produced in
P. cruentum, with α- and γ-tocopherol at 80.5 and 104.2 μg g
−1, respectively. Durmaz et al. reported 106.3 μg g
−1 biomass for both α- and γ-tocopherols in
P. cruentum [
21], which is different from the 185 μg g
−1 obtained in this study.
Figure 4 shows the accumulation of PC and APC in
S. platensis during growth.
The productivities of antioxidants in microalgae are shown in
Table 5. The productivity of tocopherols in
S. platensis and
P. cruentum showed no significant differences. Carotenoids, PC, APC, and SOD productivities were statistically different in all microalgae.
2.3. Antioxidant Concentration and Productivities in Continuous Culture at Different Light Intensities
The concentrations of total phenols, terpenoids, and alkaloids were analyzed in continuous cultures of
S. platensis,
I. galbana, and
T. suecica at different light intensities. Total phenols, alkaloids, and terpenoids concentrations were significantly different among the three microalgae (
p < 0.05;
Table 6). Phenols, alkaloids, and terpenoids content did not significantly change in any of the microalgae with 16.27 and 21.26 kJ d
−1 of supplied light intensity (
Table 6). By increasing light intensity to 30.42 kJ d
−1 phenols, alkaloids, and terpenoids content changed significantly in every microalga, except for phenols content in
I. galbana. Phenols, alkaloids, and terpenoids content showed significant differences at 39.63 kJ d
−1 in comparison with the contents of these compounds at 16.27 and 21.26 kJ d
−1 in every microalga. A significant difference in phenols content was found in
S. platensis and
I. galbana by changing light intensity from 30.42 to 39.63 kJ d
−1 (
Table 6).
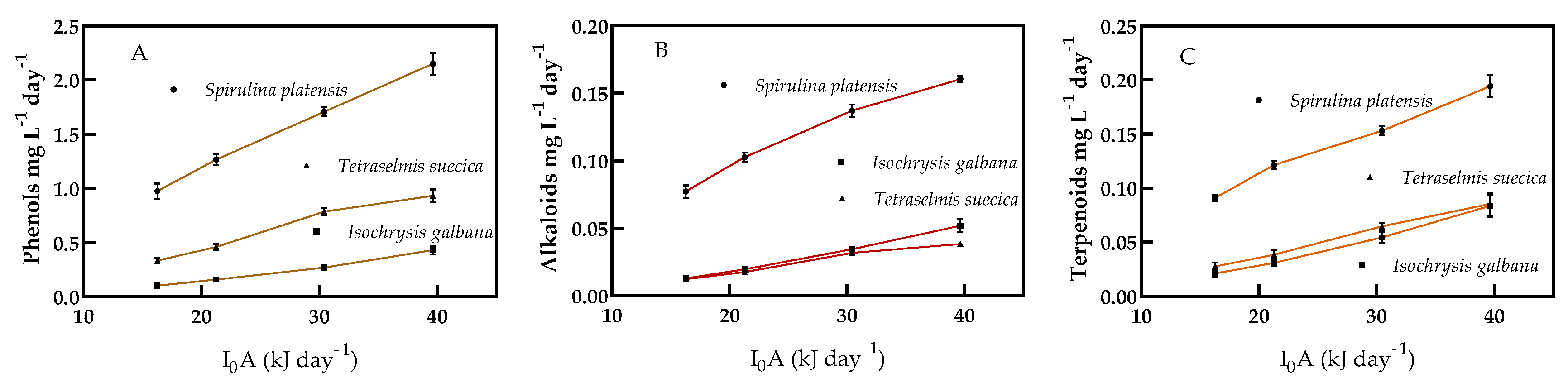
The productivity of phenols, alkaloids, and terpenoids in
S. platensis,
I. galbana, and
T. suecica under four different light intensities cultured under continuous cultivation is shown in
Figure 5A–C. The three compounds’ productivity increased significantly with increasing light intensity in every microalga. Phenol productivity in
S. platensis was five times higher than in
I. galbana, and more than twice that of
T. suecica at a light intensity of 39.63 kJ d
−1. Phenol productivity increased more than twice in
S. platensis, more than four times in
I. galbana, and three times than
T. suecica with the change in light intensity from 16.27 to 39.63 kJ d
−1 (
Figure 5A). Alkaloid productivity of
S. platensis was more than three times that of
I. galbana, and more than four times that of
T. suecica (
Figure 5B). The alkaloid productivity was 2.1 times higher in
S. platensis, 4.1 times in
I. galbana, and more than three times higher in
T. suecica with increasing the light intensity from 16.27 to 37.63 kJ d
−1 (
Figure 5B). Terpenoids productivity increased significantly with increasing light intensity. The productivity of terpenoids in
S. platensis was significantly different from the productivity in
I. galbana and
T. suecica, whereas productivity of terpenoids in
I. galbana and
T. suecica showed no significant differences (
Figure 5C).
Tocopherols content was significantly different in the four microalgae analyzed.
T. suecica and
P. cruentum showed higher amounts of tocopherols than
S. platensis and
I. galbana. In addition, tocopherols concentration increased with light intensity in the four microalgae (
Table 7). Tocopherol concentration increased 1.22 times in
P. cruentum to 1.56 times in
I. galbana by changing light intensity from 16.27 to 39.63 kJ d
−1.
Tocopherols productivity in
S. platensis and
P. cruentum was not significantly different at 16.27 and 21.26 kJ d
−1 of light intensity supplied, but significant different when light intensity increased from 21.26 to 39.63 kJ d
−1.
P. cruentum showed a productivity increase of more than 1.2 times the productivity of
S. platensis, 2.66 times the productivity of
I. galbana, and 1.57 times the productivity of
T. suecica (
Figure 6).
The carotenoids content in the microalgae changed significantly from light intensities of 16.27 to 39.63 kJ d
−1. Carotenoid content increased 1.5, 1.35, 1.56, and 1.57 times in
S. platensis,
I. galbana,
T. suecica, and
P. cruentum, respectively. At a light intensity of 39.63 kJ d
−1, carotenoids content showed no significant differences among the microalgae studied (
Table 8).
Carotenoids productivity changed with the biomass productivity for each microalga;
S. platensis had a productivity at least twice that of
I. galbana and
T. suecica and at least 1.3 times that of
P. cruentum (
Figure 7). Carotenoids productivities in
T. suecica and
I. galbana were not significantly different at light intensities of 16.27 and 21.26 kJ d
−1.
The amounts of UA SOD were significantly different among
S. platensis,
I. galbana, and
P. cruentum at different light intensities, but no difference was observed between
S. platensis and
T. suecica. SOD units increased significantly by increasing light intensity supplied in each microalga (
Table 9).
The SOD UA productivity increased in accordance with biomass productivity with increasing supplied light energy (
Figure 8). Soluble protein was found to be 65.7, 43.5, 51.3, and 77.9 mg protein g
−1 biomass for
S. platensis,
I. galbana,
T. suecica, and
P. cruentum, respectively. SOD UA productivity was significantly different in all microalgae.
Phycocyanin (PC) and allophycocyanin (APC) were found only in
S. platensis and
P. cruentum. Compared with the other antioxidants analyzed in this work, PC and APC content decreased with increasing light intensity (
Table 10). PC concentration in
S. platensis was 5.6 times the concentration in
P. cruentum, and APC concentration in
S. platensis was 7.1 times the concentration of APC in
P. cruentum at a light intensity of 39.63 kJ d
−1.
The PC production by
S. platensis was 7.0 times that by
P. cruentum; APC production of
S. platensis was 8.9 times that of
P. cruemtum (
Figure 9). Although PC and APC content decreased with increasing light intensity, the production of both pigments increased with increasing light intensity due to biomass productivity.
The extract of
S. platensis was dissolved in solvent, then the extract was analyzed by FTIR (
Table 11). Several signals appeared related to some characteristic vibrational groups. An important vibrational signal, the methylene group (2829–2848 cm
−1 [
22]), appeared at 2835 cm
−1, which is possibly found in carotenoids and phytosterols molecules such as lycopene, which is a precursor of carotene [
23]. Another vibrational signal found at 1230 cm
−1 was assigned to the phenyl group and 1517 cm
−1 to the stretching of the flavone phenyl ring; these signals correlate well with antioxidants such as flavone. The vibration band at 1740 cm
−1 corresponds to the carbonyl group of phytosterols and the signal at 1612–1652 cm
−1 to the C=O group, which are signals assigned to ketones found in flavonoids.


 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}