
Inhibition of Staphylococcus aureus LC 554891 by Moringa oleifera Seed Extract either Singly or in Combination with Antibiotics

 ,
, 
 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Isolation and Identification of Presumptive Staphylococcus aureus Strains from some Egyptian Foods
2.2. Antibiotic Sensitivity Test
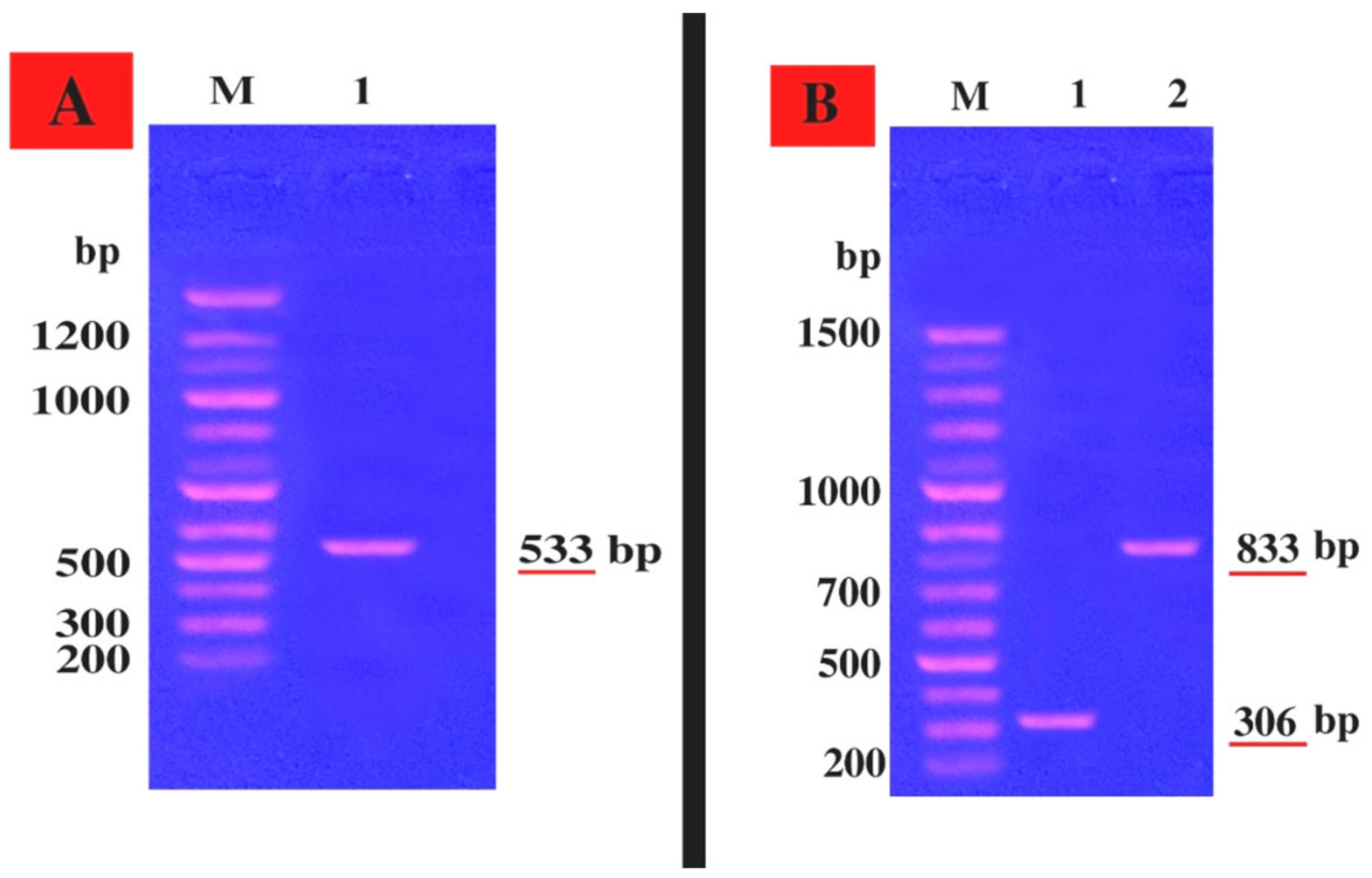
2.3. Molecular Identification of the B3 Strain by Sequencing of the 16S rRNA Gene
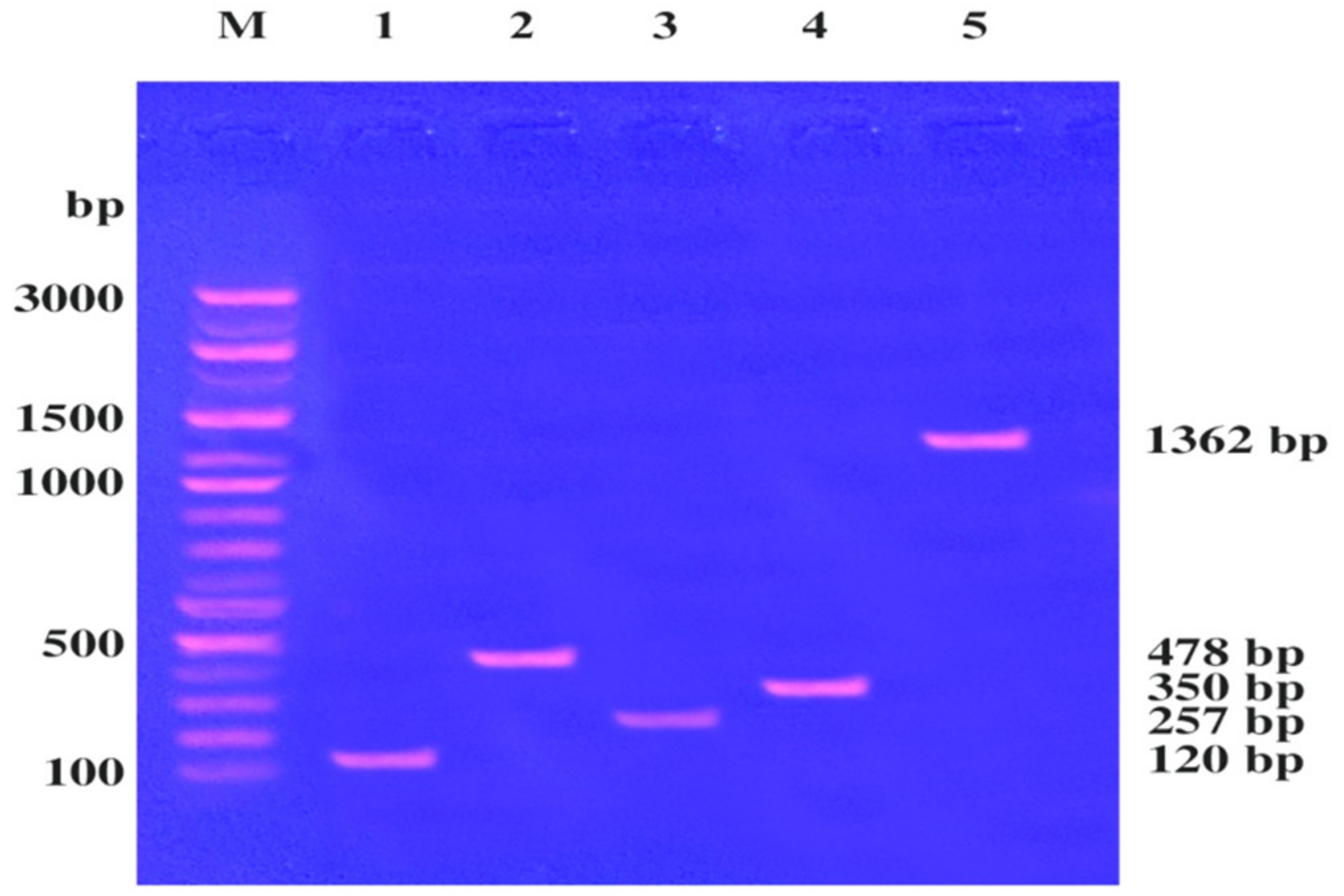
2.4. Detection of Virulence Factors within an S. aureus LC 554891 Genome
2.5. Genetic Linkage of the mecA Gene and Hemolysin Toxin hla and hlb Genes within the S. aureus LC 554891 Strain
2.6. Antibacterial Activity of some Essential Oils against S. aureus LC 554891 by the Disc Diffusion Assay
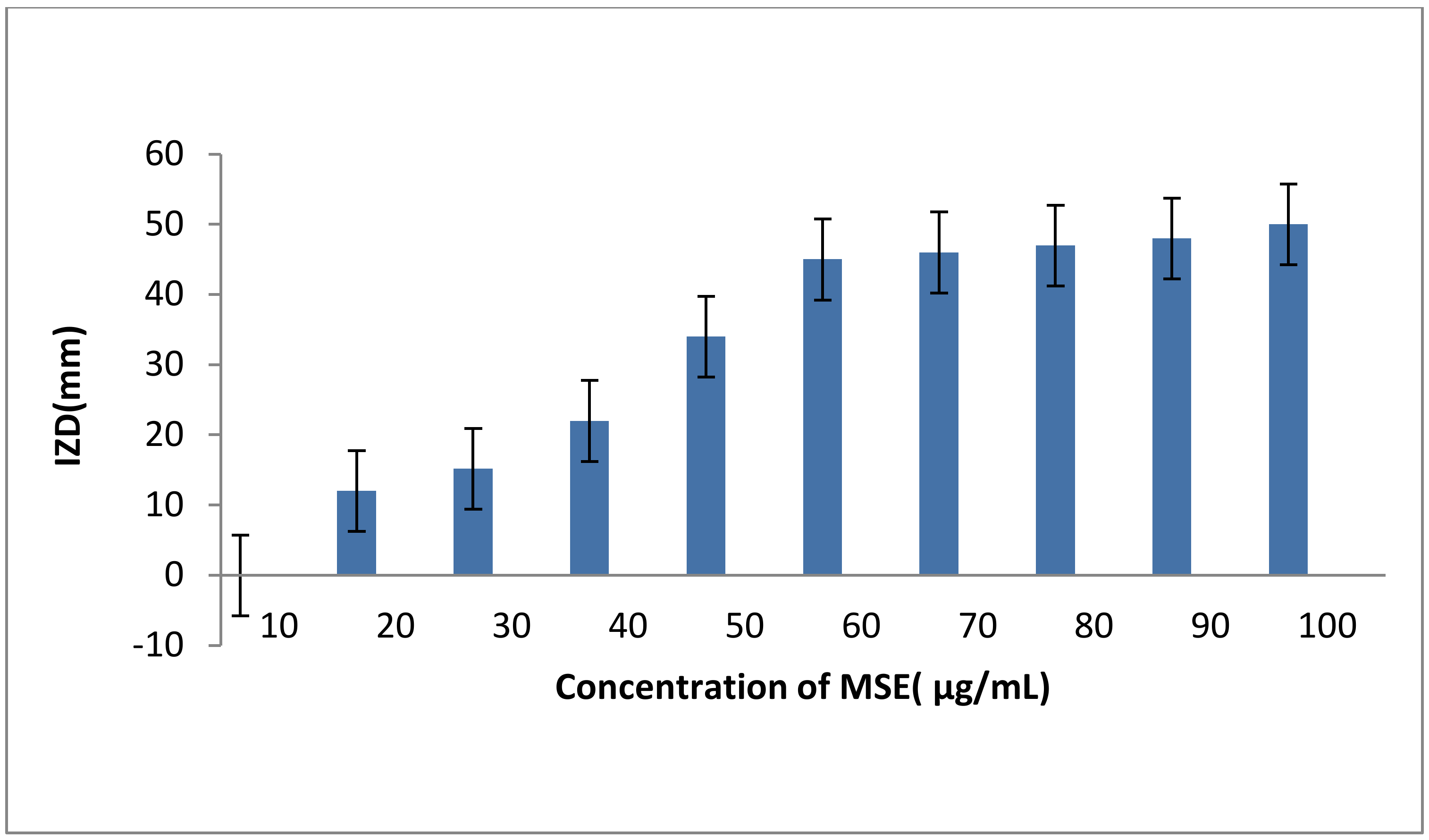
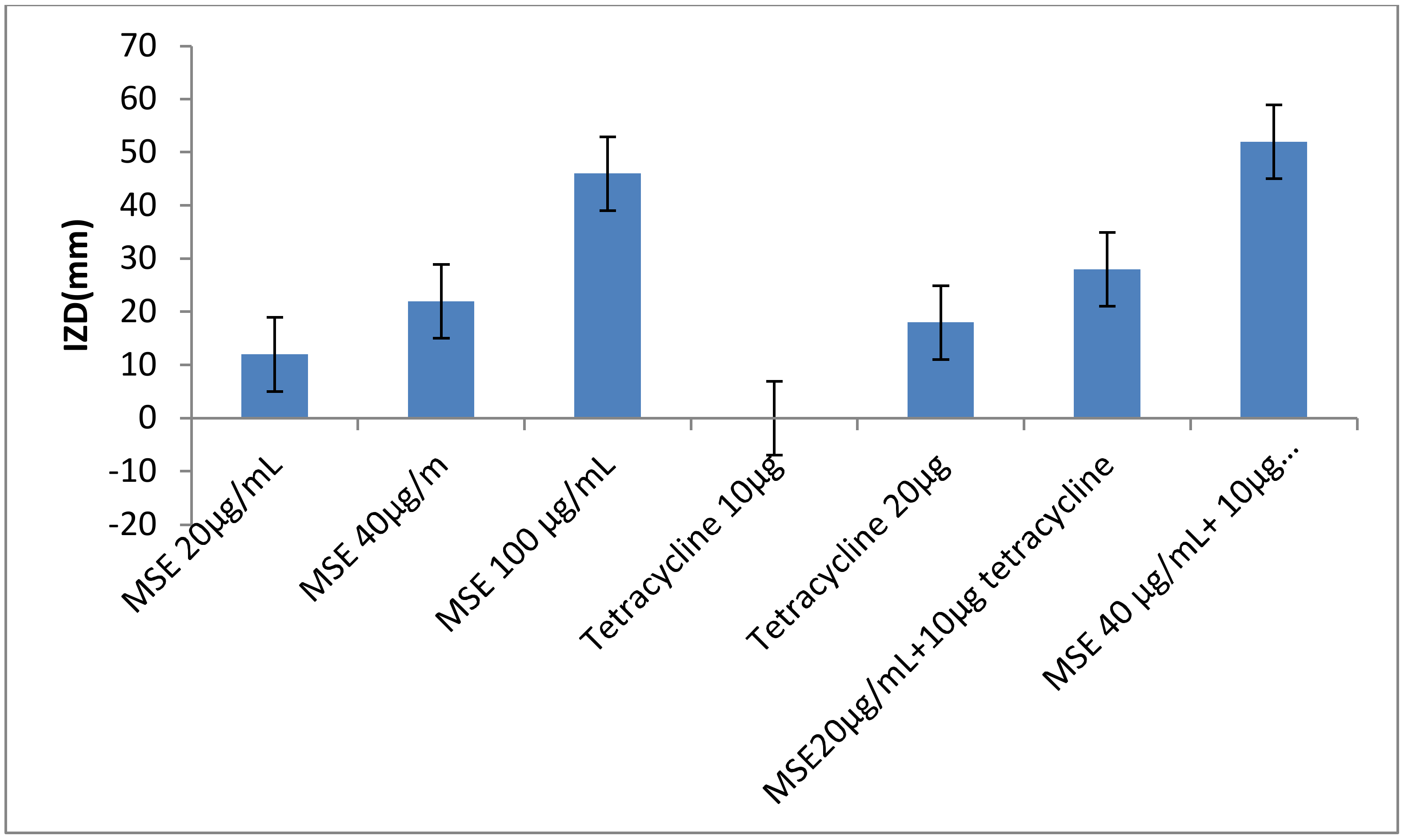
2.7. The Antibacterial Activity of M. oleifera Leaves Extract (MLE), M. oleifera Seeds Extract (MSE) and Honey Bee (HB) either Singly or in Combinations of MSE Plus Tetracycline
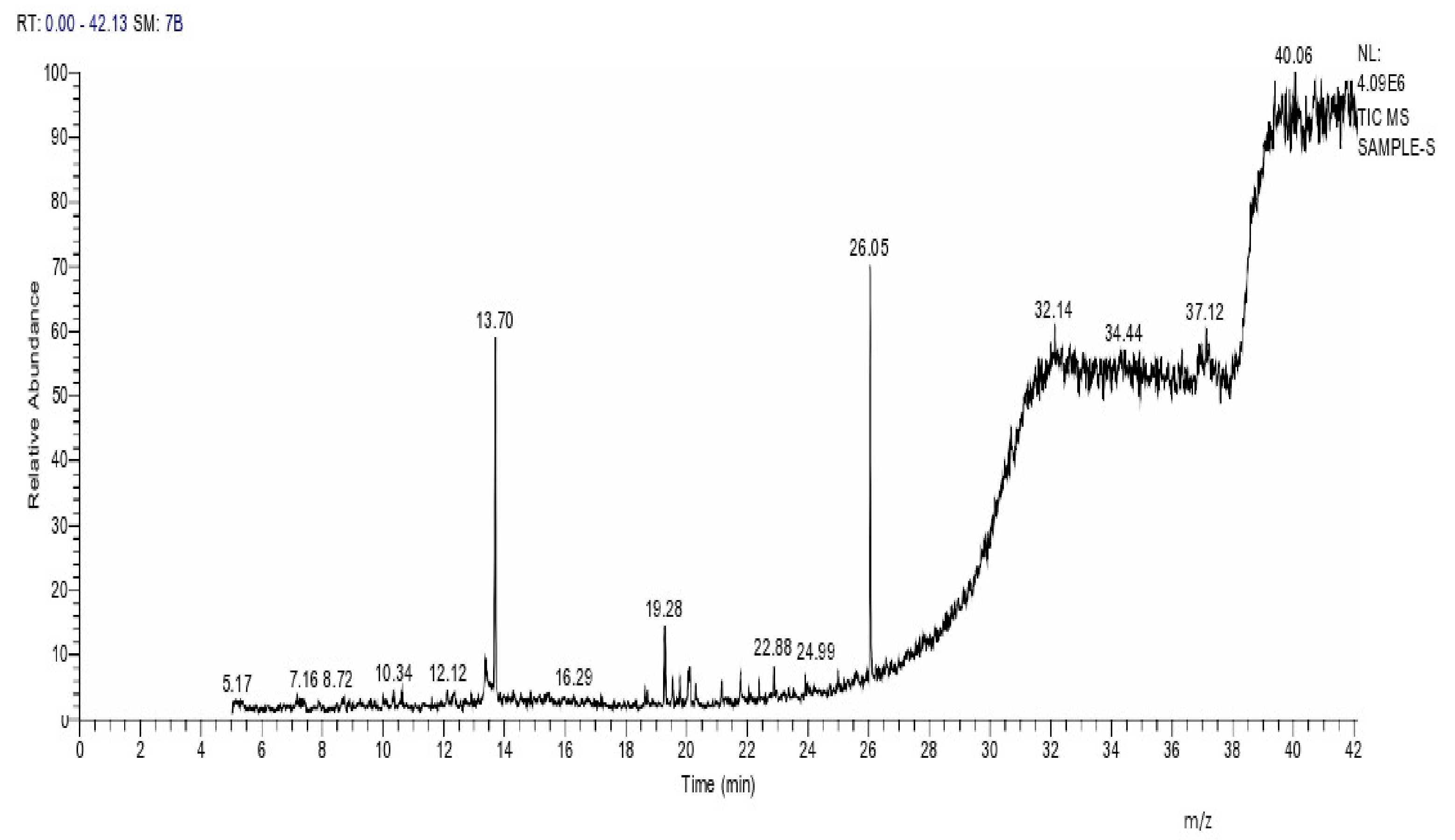
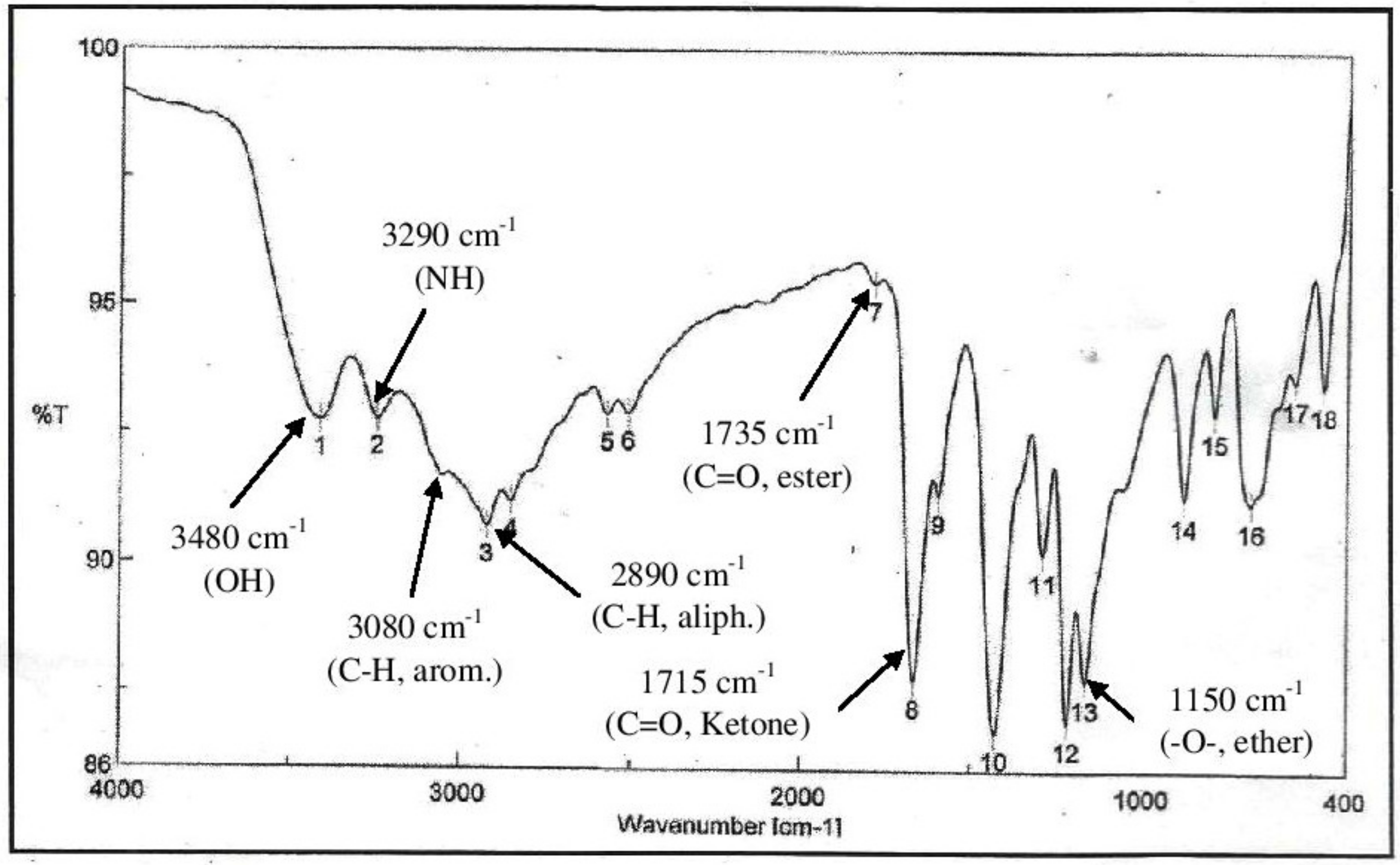
2.8. Instrumental Analysis of MSE by GC- MS
3. Discussion
4. Materials and Methods
4.1. Food Sampling
4.2. Isolation of Bacteria Suspected to Be S. aureus
4.3. Antibiotics Susceptibility Test
4.4. Molecular Identification of S. aureus No. B3
4.5. Detection of Virulence Factors (sea, seb, sec, tsst-1 and fnbA) of the Strain
4.6. Genetic Linkage of mecA, hla, and hlb Genes
4.7. Screening of the Antibacterial Activity of Essential Oils of Moringa olifera, Allium sativum, and Syzygium aromaticum against S. aureus LC 554891
4.8. Preparation of the M. oleifera Leaves (MLE) and Seeds (MSE)
4.9. Preparation of Honey Bee (HB) Solutions
4.10. Bioassay of the Antibacterial Activity of MLE, MSE, and HB
4.11. Minimum Inhibitory Concentration (MIC) of the MSE Extract
4.12. Antibacterial Activity of Combination of Antibiotics and MSE
4.13. Instrumental Analysis of MSE
4.14. Statistical Analysis
4.15. Ethical Approval
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Colombari, V.; Mayer, M.D.; Laicini, Z.M.; Mamizuka, E.; Franco, B.D.; Destro, M.T.; Landgraf, M. Food borne outbreak caused by Staphylococcus aureus: Phenotypic and genotypic characterization of strains of food and human sources. J. Food Prot. 2007, 70, 489–493. [Google Scholar] [CrossRef]
- Strommenge, B.; Layer, F.; Werner, G. Methicillin-Resistant Staphylococcus aureus in Workers in the Food Industry; Academic Press: Cambridge, MA, USA, 2018; pp. 163–188. [Google Scholar]
- Castro, A.; Silva, J.; Teixeira, P. Staphylococcus aureus, a Food Pathogen: Virulence Factors and Antibiotic Resistance. Foodborne Dis. 2018, 1085, 213–238. [Google Scholar]
- Ge, B.; Mukherjee, S.; Hsu, C.-H.; Davis, J.A.; Thuy, T.; Tran, Q.; Yang, J.W.; Abbott, S.L.; Ayers, S.R.; Young, E.T.; et al. MRSA and multidrug-resistant Staphylococcus aureus in U.S. retail meats, 2010-2011. Food Microbiol. 2017, 62, 289–297. [Google Scholar] [CrossRef]
- Sulley, M.S. The Hygienic Standard of Meat Handling in the Tamale Metropolis. Bachelor’s Thesis, University for Development Studies, Tamale, Ghana, 2006. [Google Scholar]
- Aung, K.T.; Hsu, L.Y.; Koh, T.H.; Hapuarachchi, H.C.; Chau, M.L.; Gutiérrez, R.A.; Ng, L.C. Prevalence of methicillin-resistant Staphylococcus aureus (MRSA) in retail food in Singapore. Antimicrob Resist Infect Control. 2017, 6, 94. [Google Scholar] [CrossRef] [Green Version]
- Osman, A.; El-Daidamony, G.; Sitohy, M.; Khalifa, M.; Enan, G. Soybean glycinin basic subunit inhibits methicillin resistant-vancomycin intermediate Staphylococcus aureus (MRSA-VISA) in vitro. Int. J. Appl. Res. Nat. Prod. 2016, 9, 17–26. [Google Scholar]
- Abdel-Shafi, S.; Al-Mohammadi, A.R.; Osman, A.; Enan, G.; Abdel-Hameid, S.; Sitohy, M. Characterization and Antibacterial Activity of 7S and 11S Globulins Isolated from Cowpea Seed Protein. Molecules 2019, 24, 1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Shafi, S.; Al-Mohammadi, A.; Hamdi, S.; Moustafa, A.H.; Enan, G. Biological characterization and inhibition of Streptococcus pyogenesZUH1 causing chronic cystitis by both Crocus sativus methanol extract; bee honey singly or in combination with antibiotics: An in vitro study. Molecules 2019, 24, 2903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Shafi, S.; Al-Mohammadi, A.R.; Sitohy, M.; Mousa, B.; Ismaiel, A.; Enan, G.S.; Osman, A. Antimicrobial Activity and Chemical Constitution of the Crude, Phenolic-Rich Extracts of Hibiscus sabdariffa, Brassica oleracea and Beta vulgaris. Molecules 2019, 24, 4280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abd Rani, N.Z.; Hussain, K.; Kumolosasi, E. Moringa Genus: A review of phytochemistry and pharmacology. Front. Pharmacol. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, F.; Martins, N.; Barros, L.; Rodrigues, M.E.; Oliveira, M.B.; Henriques, M.; Ferreira, I.C. Plant phenolic extracts as an effective strategy to control Staphylococcus aureus, the dairy industry pathogen. Ind. Crops Prod. 2018, 112, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Maddocks, S.E.; Lopez, R.S.; Rowlands, R.S.; Cooper, R.A. Manuka honey inhibits the development of Streptococcus pyogenes biofilms and causes reduced expression of two fibronectin binding proteins. Microbiology 2012, 158, 781–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, R.; Burton, N.; Cooper, R. Manuka honey inhibits cell division in methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2011, 66, 2536–2542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramosa, O.Y.; Salomón, V.; Libonattic, C.; Cepedad, R.; Maldonadob, L.; Basualdoc, M. Effect of botanical and physicochemical composition of Argentinean honeys on the inhibitory action against food pathogens. LWT Food Sci. Technol. 2018, 87, 457–463. [Google Scholar] [CrossRef]
- Enan, G.; El-Essawy, A.A.; Uyttendael, M.; Debevere, J. Antibacterial activity of Lactobacillus planetarium UG1 isolated from dry sausage: Characterization, production and bactericidal action of plantaricin UG1. Int. J. Food Microbiol. 1996, 30, 189–215. [Google Scholar] [CrossRef]
- Enan, G.; Abdel-Shafi, S.; Ouda, S.; Negm, S. Novel antibacterial activity of LactococcusLactis subspecies lactis Z11 isolated from Zabady. Int. J. Biomed. Sci. 2013, 9, 144–180. [Google Scholar]
- Enan, G.; Seham, A.S.; Abdel-Halem, M.F.; Negm, S. Characterization of probiotic lactic acid bacteria to be used as starter and protective cultures for dairy fermentations. Int. J. Probiotics Prebiotics 2013, 8, 157–163. [Google Scholar]
- El-Gazzar, N.; Ismail, A.M. The potential use of Titanium, Silver and Selenium nanoparticles in controlling leaf blight of tomato caused by Alternaria alternata. Biocatal. Agric. Biotechnol. 2020, 27, 101708. [Google Scholar] [CrossRef]
- Enan, G.; Abdel-Haliem, M.E.F.; Tartour, E. Evaluation of the antimicrobial activity, starter capability and technological properties of some probiotic bacteria isolated from some Egyptian Pickles. Life Sci. J. 2014, 11, 976–985. [Google Scholar]
- Abdel-Shafi, S.; Osman, A.; Enan, G.; Sitohy, M.Z. Antibacterial activity of methylated egg white proteins against pathogenic G+ and G− bacteria matching antibiotics. Springerplus 2016, 5, 983–996. [Google Scholar] [CrossRef] [Green Version]
- FDA. Revised guidelines for the assessment of microbiological quality of processed food. Retrieved from 2013. Available online: http://www.fda.gov.ph/attachments/article/17218/FC2013-010 (accessed on 9 September 2020).
- Ebert, M. Hygiene principles to avoid contamination/cross-contamination in the kitchen and during food processing. In Staphylococcus aureus; Academic Press: Cambridge, MA, USA, 2018; Chapter 11; pp. 217–234. [Google Scholar]
- Ulusoy, B.H.; Sancar, B.C.; Öztürk, M. Prevalence of Staphylococcal Enterotoxins in Ready-to-Eat Foods Sold in Istanbul. J. Food Prot. 2017, 80, 1734–1736. [Google Scholar] [CrossRef]
- Ezeamagu, C.; Imanatue, I.; Dosunmu, M.; Odeseye, A.; Baysah, G.; Aina, D.; Odutayo, F.; Mensah-Agyei, G. Detection of methicillin resistant and toxin-associated genes in Staphylococcus aureus. Beni Suef Univ. J. Basic Appl. Sci. 2018, 7, 92–97. [Google Scholar] [CrossRef]
- Liu, J.; Wang, Z.; Ma, H.; Wang, S. Probing and quantifying the food-borne pathogens and toxins: From in Vitro to in Vivo. J. Agric. Food Chem. 2018, 66, 1061–1066. [Google Scholar] [CrossRef] [PubMed]
- Rajendhran, J.; Gunasekaran, P. Microbial phylogeny and diversity: Small sub unit ribosomal RNA sequence analysis and beyond. Microbol. Res. 2010, 166, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Eilert, U.; Wolters, B.; Nahrstedt, A. The antibiotic principle of seeds of Moringa oleifera and Moringa stenopetala. Planta Med. 1981, 42, 55–61. [Google Scholar] [CrossRef]
- Omosa, K.L.; Jacob, O.M.; Armella, M.T.; Mbaveng, M.; Tankeo, S.B.; Seukep, J.A.; Voukehg, I.K.; Dzotam, J.K.; Isemk, J.; Decrese, s.; et al. Antibacterial activities and structure–activity relationships of a panel of 48 compounds from Kenyan plants against multidrug resistant phenotypes. Springerplus 2016, 5, 901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, J.K.; Das, G.; Baek, K.H. Chemical Composition and Antioxidant and Antibacterial Activities of an essential oil Extracted from an Edible Seaweed, Laminaria japonica L. Molecules 2015, 20, 12093–12113. [Google Scholar] [CrossRef] [Green Version]
- Kumar, C.G.; Mongolla, P.; Pombala, S.; Kamle, A.J. Physicochemical characterization and antioxidant activity of melanin from a novel strain of Aspergillus bridgeri ICTF-201. Lett. Appl. Microbiol. 2011, 53, 350–358. [Google Scholar] [CrossRef]
- Dolan, N.; Gavin, D.P.; Eshwika, A.; Kavanagh, K.; McGinley, J.; Stephens, J.C. Synthesis, antibacterial and anti-MRSA activity, in vivo toxicity, and a structure–activity relationship study of a quinolinethiourea. Bioorg. Med. Chem. Lett. 2016, 26, 630–635. [Google Scholar] [CrossRef] [Green Version]
- Mukhtyar, S.; Kumar, A.; Dwivedi, J.; Singh, R. A review: Biological significance of heterocyclic compounds. Int. J. Pharm. Sci. Res. 2013, 4, 66–76. [Google Scholar]
- Anwar, F.; Latif, M.S.; Ashraf., M.; Gilani, A.H. Moringa oleifera: A food Plant with multiple medicinal uses. Phytother. Res. 2007, 21, 17–25. [Google Scholar] [CrossRef]
- Bilal, A.N.; Molan, P.C.; Sallal, A.K. Antimicrobial activity of honey on selected microorganisms: A preliminary study. Biomed. Res. 1998, 9, 51–54. [Google Scholar]
- Mama, M.; Teshome, T.; Detamo, J. Antibacterial Activity of Honey against Methicillin-Resistant Staphylococcus aureus: A Laboratory-Based Experimental Study. Int. J. Microbiol. 2019, 7686130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeleke, O.E.; Olaitan, J.O.; Okpekpe, E.I. Comparative antibacterial activity of honey and gentamicin against E. coli and S. aureus. Ann. Burn. Fire Disasters 2006, 19, 201–205. [Google Scholar]
- Moundoi, M.A.; Padila-Zakour, O.I.; Worobo, R.W. Antimicrobial activity of honey against food pathogens and food spoilage microorganisms. N. Y. State Agric. Exp. Stn. 2001, 1, 61–71. [Google Scholar]
- Abdel-Shafi, S.; Osman, A.; Al-Mohammadi, A.R.; Enan, G.; Kamal, N.; Sitohy, M. Biochemical, biological characteristics and antibacterial activity of glycoprotein extracted from the epidermal mucus of African catfish (Clariasgariepinus). Int. J. Biol. Macromol. 2019, 138, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Aiyegoro, O.A.; Okoh, A.I. Use of bioactive plant products in combinartion with standard antibiotics; implications in antimicrobial chemotherapy. J. Med. Plants Res. 2009, 3, 1147–1152. [Google Scholar]
- Oludare, T.O.; Oluduro, A.O.; Idowu, T.O. Assessment of Nephrotoxicity, Anti-inflammatory and Antioxidant properties of Epigallocatechin, Epicatechin and Stigmasterolphytosterol (synergy) Derived from ethyl acetate stem bark extract of Spondiasmombinon Wister Rats Using Molecular method of analysis. J. Mol. Microbiol. 2017, 1, 1–11. [Google Scholar]
- Arikawa, J.; Ishibashi, M.; Kawashima, M.; Takaqi, Y.; Ichikawa, Y. Decreased levels of sphingosine, a natural antimicrobial agent, may be associated with vulnerability of the stratum corneum from patients with atopic dermatitis to colonization by Staphylococcus aureus. J. Invest. Dermatol. 2002, 119, 433–439. [Google Scholar] [CrossRef]
- Drake, D.R.; Brogden, K.A.; Dawson, D.V.; Wertz, P.W. Thematic review series: Skin lipids. Antimicrobial lipids at the skin surface. J. Lipid Res. 2008, 49, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, X.; Wu, A. Mini Review on Antimicrobial Activity and Bioactive Compounds of Moringa oleifera. Med. Chem. 2016, 6, 9. [Google Scholar] [CrossRef] [Green Version]
- Tirado-Torres, D.; Chan-Keb, C.A.; Perez-Balan, R.A.; Ake-Canché, B.; Gómez-Solano, M.I.; Aragón-Gastélum, J.L.; Gómez-López, I.; Aguirre-Crespo, F.J.; López-Ramos, M.C.; Gutiérrez-Alcantara, E.J. Antimicrobial activity of Moringa oleifera against multidrug-resistant Staphylococcus aureus isolated from raw milk. Appl. Ecol. Environ. Res. 2019, 17, 587–599. [Google Scholar] [CrossRef]
- Othman, A.S. Bactericidal Efficacy of Omega-3 Fatty Acids and Esters Present in Moringa oleifera and Portulaca oleracea Fixed Oils Against Oral and Gastro Enteric Bacteria. Int. J. Pharmacol. 2017, 1811–7775. [Google Scholar] [CrossRef]
- Dzotam, J.K.; Touani, F.K.; Kuete, V. Antibacterial and antibiotic-modifying activities of three food plants (Xanthosoma mafaffa Lam., Moringa oleifera (L.) Schott and Passiflora edulis Sims) against multidrug-resistant (MDR) Gram-negative bacteria. BMC Complement. Altern Med. 2016, 16, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Othman, L.; Sleiman, A.; Abdel-Massih, R.M. Antimicrobial Activity of Polyphenols and Alkaloids in Middle Eastern Plants. Front. Microbiol. 2019, 10, 911. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing: Eighteenth Informational Supplement; CLSI: Wayne, PA, USA, 2008. [Google Scholar]
- Raja, M.M.M.; John, S.A. Multidrug resistance profile of urinary tract infected Gram positive pathogenic bacterial isolates. Int. J. Infect. 2015, 2, e22774. [Google Scholar]
- Sambrook, J.; Russel, D. Molecular Cloning: A Laboratory Manual, 3rd ed.; Cold Springs Harbour Laboratory Press: Woodbury NY, USA, 2001. [Google Scholar]
- Chénbey, D.; Philippot, L.; Hartmann, A.; Hénalut, C.; Germon, J.C. 16S rDNA analysis for characterization of denitrifying bacterial isolated from three agricultural soils. FEMS Microbiol. Ecol. 2000, 24, 121–128. [Google Scholar] [CrossRef]
- Turner, S.; Preyer, K.M.; Mias, V.P.W.; Palmer, D.J. Investigation of phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef]
- Sanger, F.; Nicklen, S.; Coulson, A.R. DNA sequencing with chain-terminating inhibitors. Proc. Nat. Acad. Sci. USA 1977, 74, 5463–5467. [Google Scholar] [CrossRef] [Green Version]
- Freeman, K.H.; Hayes, J.M.; Trendel, J.M.; Albrecht, P. Evidence from GC-MS carbon isotopic measurements for multiple origins of sedimentary hydrocarbons. Nature 1990, 353, 627–644. [Google Scholar]
- Williams, J.K.; Kubelisk, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Salam, H.A.; El-Khamisssy, T.; Enan, G.A.; Hollenberg, C.P. Expression of mouse anticreatine kinase (MAK33) monoclonal antibody in the yeast Hansenulapolymorpha. Appl. Microbiol. Biotechnol. 2001, 56, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Purrello, S.M.; Daum, R.S.; Edwards, G.F.S.; Lina, G.; Lindsay, J.; Peters, G.; Stefani, S. Meticillin-Resistant Staphylococcus Aureus (MRSA) Update: New Insights Into Bacterial Adaptation and Therapeutic Targets. J. Glob Antimicrob. Resist. 2014, 2, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.; Minamide, W.; Wada, K.; Nakmura, E.; Teraoka, H.; Watanabe, S. Identification of methicillin-resistant strains of Staphylococci by polymerase chain reaction. J. Clin. Microbiol. 1991, 29, 2240–2244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Booth, M.; Pence, L.; Mahasresthi, P.; Callegan, M.; Gilmore, M. Clonal Association among Staphylococcus aureus isolates from Various Sites of Infection. Infect Immun. 2001, 69, 345–352. [Google Scholar] [CrossRef] [Green Version]
- Goerke, C.; Flucklger, U.; Steinhuber, A.; Zimmerli, W. Impact of the regulatory loci agr, sarA and sae of Staphylococcus aureus on the induction of a-toxin during device-related infection resolved by direct quantitative transcript analysis. Mol. Microbiol. 2001, 40, 1439–1447. [Google Scholar] [CrossRef] [Green Version]
- Dub, A.M.; Dugani, A.M. Antithrombotic effect of repeated doses of ethanolic extract of local olive (Oleaeuropaea L.) leaves in Rabbits. Libyan J. Med. 2013, 8, 20947. [Google Scholar] [CrossRef]
- Patton, T.; Barrett, J.; Brennan, N.; Moran, N. Use of a spectrophotometric bioassay for determination of microbial sensitivity to manuka honey. J. Microbiol. Methods 2006, 64, 84–95. [Google Scholar] [CrossRef]
- Abdel-Shafi, S.; Al-Mohammadi, A.R.; Almanaa, T.N.; Moustafa, A.H.; Saad, T.M.M.; Ghonemy, A.; Anacarso, I.; Enan, G.; El-Gazzar, N. Identification and testing antidermatophytic oxaborole-6-benzene sulphonoamide derivative (OXBS) from Streptomyces atrovirens KM192347 isolated from soil. Antibiotics 2020, 9, 176. [Google Scholar] [CrossRef]
- Al-Rubaye, A.F.; Hamid, I.H.; Kadhvin, M.J. A Review: Uses of gas chromatography- Mass spectrometry (GC-MS) technique for analysis of bioactive material compounds of some plants. Int. J. Toxicol. Pharmacol. Res. 2017, 9, 81–85. [Google Scholar] [CrossRef]
- El-Gazzar, N.; Almaary, K.H.; Ismail, A.; Polizzi, G. Influence of Funneliformis mosseae enhanced with titanium dioxide nanoparticles (TiO2NPs) on Phaseolus vulgaris L. under salinity stress. PLoS ONE 2020, 15, e0235355. [Google Scholar] [CrossRef]
- Victoria, C.N.; Harrison, J.; Cox, J.A.G. Dissecting the antimicrobial compostion of honey. Antibiotics 2019, 8, 251. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Not available. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Diameters of the Inhibition Zone (mm) ± SD | MAR Index | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SP | DA | DO | AM | TE | CIP | N | OFX | AMC | P | OX | ME | ||
| S. aureus ATCC 6538 | 0 | 0 | 0 | 0.8 ± 0.10 | 0 | 2.1 ± 0.15 | 1± 0.07 | 0 | 0.8 ± 0.11 | 1.7 ± 0.18 | 1 ± 0.09 | 0.9 ± 0.13 | 5 (41.67%) |
| S. aureus B3 | 0 | 0 | 0 | 0 | 4± 0.36 | 3 ± 0.25 | 0 | 0 | 0 | 0 | 0 | 0 | 10 (83.3%) |
| S. aureus B7 | 1 | 0.9 ± 0.08 | 2.9 ± 0.18 | 0 | 3 ± 0.29 | 2.1 ± 0.16 | 0 | 0 | 1.2 ± 0.08 | 1.7 ± 0.15 | 0 | 0.9 ± 0.10 | 4 (33.33%) |
| S. aureus B8 | 0 | 2.9 ± 0.26 | 1.5 ± 0.15 | 0 | 0 | 3 ± 0.34 | 0.8 ± 0.10 | 0 | 1 ± 0.10 | 0 | 0 | 0 | 7 (58.33%) |
| S. aureus B14 | 0 | 0.9 ± 0.10 | 2.9 ± 0.23 | 0 | 1 | 2.1 ± 0.20 | o.8 ± 0.11 | 0 | 1.6 ± 0.21 | 0 | 0 | 0.9± 0.11 | 5(41.67%) |
| S. aureus B17 | 1.1 ± 0.09 | 0.8 ± 0.12 | 0 | 0 | 0 | 2.1 ± 0.22 | 1.1 ± 0.14 | 0 | 0 | 0 | 2.9 ± 0.32 | 1.7 ± 0.15 | 6 (50%) |
| S. aureus B18 | 2.1 ± 0.20 | 1.1 ± 0.13 | 0 | 2.9 ± 0.32 | 0 | 0 | 0 | 0 | 1.1 ± 0.15 | 1.7 ± 0.16 | 2.1 ± 0.39 | 1.1 | 5 (41.67%) |
| S. aureus B22 | 2.1 ± 0.22 | 0.8 ± 0.10 | 2.1 ± 0.20 | 0 | 0 | 2.1 ± 0.25 | 0.9 ± 0.09 | 0 | 1 ± 0.12 | 0 | 2.9 ± 0.40 | 1.7 ± 0.15 | 4 (33.33%) |
| S. aureus B24 | 2.9 ± 0.30 | 1.7 ± 0.14 | 0 | 0 | 0 | 2.1 ± 0.23 | 0.8 ± 0.10 | 0 | 1.2 ± 0.13 | 0 | 0 | 0 | 7 (58.33%) |
| S. aureusCh32 | 2.9 ± 0.31 | 0 | 2.9 ± 0.33 | 2.9 ± 0.35 | 0 | 0 | 0 | 0 | 1 ± 0.09 | 1.7 ± 0.18 | 2.1 ± 0.28 | 1 ± 0.10 | 5 (41.67%) |
| S. aureus Ch35 | 1.1 ± 0.12 | 2.1 ± 0.26 | 0 | 2.9 ± 0.32 | 0 | 2.1 ± 0.22 | 1 ± 0.13 | 0 | 0 | 1.7 ± 0.16 | 1 ± 0.10 | 1.1 ± 0.13 | 4 (33.33%) |
| S. aureus Ch40 | 0.8 ± 0.10 | 0 | 0.8 ± 0.09 | 2.9 ± 0.39 | 0 | 0 | 0 | 0 | 1 ± 0.10 | 2 ± 0.22 | 0 | 0.9 ± 0.09 | 6 (50%) |
| S. aureus Ch41 | 0 | 3.2 ± 0.27 | 0.9 ± 0.10 | 3.2 ± 0.25 | 0 | 0 | 0 | 0 | 0 | 2 ± 0.23 | 0 | 0.9 ± 0.09 | 7 (58.33%) |
| S. aureus Ch48 | 1.2 ± 0.07 | 1 ± 0.1 | 0 | 2.9 ± 0.31 | 0 | 2.1± 0.22 | 1 ± 0.11 | 0 | 0 | 1.7 ± 0.15 | 0.9 ± 0.07 | 1 ± 0.10 | 4 (33.33%) |
| S. aureus Ch50 | 1.2 ± 0.07 | 0 | 2.1 ± 0.19 | 2.9 ± 0.33 | 0 | 0 | 0 | 0 | 1 ± 0.11 | 1.7 ± 0.16 | 0 | 0 | 7 (58.33%) |
| S. aureus Ch53 | 2.7 ± 0.30 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1.2 ± 0.10 | 1.7 ± 0.15 | 2.7 ± 0.25 | 1 ± 0.10 | 7 (58.33%) |
| S. aureus Cf58 | 1.2 ± 0.11 | 0 | 0 | 3.2 ± 0.33 | 0 | 0 | 0.9 ± 0.08 | 0 | 0 | 2 ± 0.25 | 3.4 ± 0.41 | 1.3 ± 0.08 | 6 (50%) |
| S. aureus Cf66 | 3.5 ± 0.25 | 0.9 ± 0.88 | 0 | 2 ± 0.19 | 0 | 3.5 ± 0.30 | 0.9 ± 0.10 | 0 | 0 | 1.3 ± 0.11 | 0 | 0.9 ± 0.08 | 5 (41.67%) |
| S. aureus Cf69 | 1 ± 0.09 | 3.2 ± 0.29 | 2 ± 0.18 | 0 | 0 | 3.5 ± 0.33 | 0.9 ± 0.12 | 0 | 1.3 ± 0.11 | 0 | 0 | 0.9± 0.08 | 5 (41.67%) |
| No. of strains (%) | 5(19) 26.3% | 7(19) 36.8% | 10(19) 52.6% | 9(19) 47.4% | 16(19) 84.2% | 7(19) 36.8% | 8(19) 42.1% | 19(19) 100% | 8(19) 42.1% | 7(19) 36.8% | 10(19) 52.6% | 4(19) 21.1% | |
| Diameters of Inhibition Zone (mm) ± SD | |||||||
|---|---|---|---|---|---|---|---|
| Strains | Tetracycline 10 µg/disc (Positive Control) | Garlic Oil | Moringa Oil | Clove Oil | |||
| 0.25% | 0.5% | 0.25% | 0.5% | 0.25% | 0.5% | ||
| S. aureus ATCC 6538 | 16.0 ± 0.0 | 23 ± 0.2 | 31 ± 0.2 | 35 ± 0.1 | 37 ± 0.1 | 14 ± 0.1 | 23 ± 0.0 |
| S. aureus LC 554891 | 0 | 25 ± 0.1 | 32 ± 0.2 | 30 ± 0.1 | 35 ± 0.2 | 13 ± 0.3 | 24 ± 0.1 |
| The Bacteria | Diameters of Inhibition Zone (mm) ± SD | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tetracycline (10 µg) (Positive Control) | MLE (µg/mL) | MSE (µg/mL) | HB (%) | |||||||||||||||||||
| Methanolic Extract (ME) | Ethanolic Extract (EE) | Water Extract (WE) | Methanolic Extract (ME) | Ethanolic Extract (EE) | Water Extract (WE) | |||||||||||||||||
| 50 | 100 | 200 | 50 | 100 | 200 | 50 | 100 | 200 | 50 | 100 | 200 | 50 | 100 | 200 | 50 | 100 | 200 | 5 | 50 | 100 | ||
| S. aureus ATCC 6538 | 15.0 ± 0.0 | 9.5 ± 0.31 | 11 ± 0.00 | 13.0 ± 0.91 | 12.0 ± 0.7 | 13.1 ± 0.47 | 14.7 ± 0.7 | 5.6 ± 0.24 | 8.3 ± 0.91 | 9.3 ± 0.46 | 12.00 ± 0.00 | 15.2 ± 00.1 | 18.81 ± 0.61 | 8.1 ± 0.00 | 9.31 ± 0.07 | 10.00 ± 0.00 | 34 ± 0.11 | 47 ± 0.00 | 50.00 ± 0.00 | 0 | 30.1 ± 0.2 | 34.43 ± 0.00 |
| S. aureus LC 554891 | 0 | 8.3 ± 0.7 | 9.3 ± 0.46 | 11.0 ± 0.00 | 10.0 ± 0.7 | 10.0 ± 0.7 | 13.0 ± 0.00 | 2.3 ± 0.00 | 5.6 ± 0.00 | 6.4 ± 0.7 | 11.00 ± 0.98 | 14.25 ± 0.12 | 17.75 ± 0.41 | 8.0 ± 0.00 | 9.0 ± 0.07 | 10.00 ± 0.00 | 32 ± 0.11 | 45 ± 0.11 | 48.00 ± 0.00 | 0 | 30.51 ± 0.07 | 34.00 ± 0.00 |
| No. | Classification | M. Formula | M.W. | Compound Name and Structure | Area | Parent Ion (M+) | Base Peak (m/e) (100%) |
|---|---|---|---|---|---|---|---|
| 1 | Spiro ketone | C17H24 O3 | 276.0 |  7,9-Di-tert-butyl-1-oxaspiro [4,5] deca-6,9-diene-2,8-diene | 5.25 | 276.0 | 57.00 |
| 2 | Ester | C18H28O3 | 292.0 |  Methyl 3-[3,5-di-tert-butyl-4-hdroxy phenyl] propionate | 2.04 | 292.0 | 277.0 |
| 3 | Heterocyclic compounds | C14H22N2O | 234.0 | 1-Methyl-2-cyano-3-ethyl-4-pivaloyl-2-piperidine | 2.08 | 234.0 | 149.0 |
| 4 | Polynuclear ketone | C10H10O4 | 194.0 | 3,6,8-Trilydroxy-Naphtalen-1-one | 2.08 | 195.0(M+1) | 149.0 |
| 5 | Saturated fatty ester | C18H36O2 | 284.0 | Ethyl hexadecanoate | 7.12 | 284.0 | 88.0 |
| 6 | Ketone | C10H11IO 3 | 306.0 | 6-Iodoacetoveratrone | 1.77 | 308.0(M+2) | 291.0 |
| 7 | Ketone | C13H18 O2 | 206 | 2-Alyl-5-t-butyl hydroquinone | 27.82 | 207.0 (M+1) | 191 |
| 8 | Fatty Ether | C14H28O | 212.0 |  Vinyl lauryl ether | 2.04 | 212.0 | 43.0 |
| 9 | Fatty ester | C16H32O3 | 272.0 | Hydroxy ethyl myristate | 2.66 | 272.0 | 104.0&43.0 |
| 10 | Fatty ester | C18H36O3 | 300.0 | 2-Hydroxy ethyl palmitate | 2.66 | 300.0 | 104.0&43.00 |
| 11 | Dibasic fatty acid | C18H34O4 | 314.0 | Octadecanedoic | 2.66 | 314.0 | 98.0 |
| 12 | Ester | C24H38O4 | 390.0 |  Bis [2-ethyl hexyl] phlthalate | 29.30 | 391.0(M+1) | 149.0 |
| 13 | Fatty ester | C39H76O5 | 624.0 | 1-(Hydroxymethyl)-1,2-etheraneelyl ester octadecanoic acid | 625.0(M+1) | 267.0 | |
| 14 | Aromatic amines | C13H17NO | 203.0 | Formylcyclohexyl Aniline | 3.43 | 203.0 | 174.0 |
| Detected Virulence Factors | Primer Sequence (Forwarded) | Primer Sequence (Reverse) | Size of the PCR Products (bp) |
|---|---|---|---|
| Sea | TTGGAAACGGTTAAAACGAA | GAACCTTCCGATCAAAAACA | 120 |
| Seb | TCGCATCAAACTGACAAACG | GCAGGTACTCTATAAGTGCC | 478 |
| Sec | GACATAAAAGCTAGGAATTT | AAATCGGATTAACATTATCC | 257 |
| Tsst-1 | ATGGCAGCATCAGCTTGATA | TTTCCAATAACCACCCGTTT | 350 |
| fnbA | CACAACCAGCAAATATAG | CTG TGTGGTAATCAATGT | 1362 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enan, G.; Al-Mohammadi, A.-R.; Mahgoub, S.; Abdel-Shafi, S.; Askar, E.; Ghaly, M.F.; Taha, M.A.; El-Gazzar, N. Inhibition of Staphylococcus aureus LC 554891 by Moringa oleifera Seed Extract either Singly or in Combination with Antibiotics. Molecules 2020, 25, 4583. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25194583
Enan G, Al-Mohammadi A-R, Mahgoub S, Abdel-Shafi S, Askar E, Ghaly MF, Taha MA, El-Gazzar N. Inhibition of Staphylococcus aureus LC 554891 by Moringa oleifera Seed Extract either Singly or in Combination with Antibiotics. Molecules. 2020; 25(19):4583. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25194583
Chicago/Turabian StyleEnan, Gamal, Abdul-Raouf Al-Mohammadi, Samir Mahgoub, Seham Abdel-Shafi, Eman Askar, Mohamed F. Ghaly, Mohamed A. Taha, and Nashwa El-Gazzar. 2020. "Inhibition of Staphylococcus aureus LC 554891 by Moringa oleifera Seed Extract either Singly or in Combination with Antibiotics" Molecules 25, no. 19: 4583. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25194583