
Synergistic Effect of Propidium Iodide and Small Molecule Antibiotics with the Antimicrobial Peptide Dendrimer G3KL against Gram-Negative Bacteria


Abstract
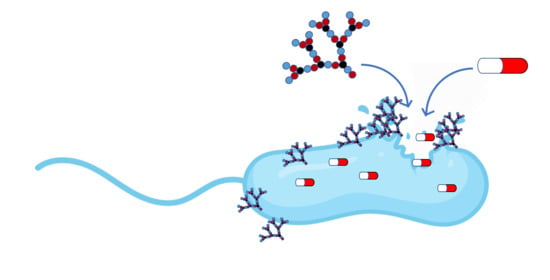
:
1. Introduction
2. Results and Discussion
2.1. Membrane Permeabilization and Synergy with Propidium Iodide
2.2. G3KL Synergizes with Small Molecule Antibiotics against K. pneumoniae
3. Materials and Methods
3.1. Compounds
3.2. Broth Microdilution Method
3.3. Checkerboard Assay
3.4. Time-Kill Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Li, X.Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria: An update. Drugs 2009, 69, 1555–1623. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Mojsoska, B.; Jenssen, H. Peptides and Peptidomimetics for Antimicrobial Drug Design. Pharmaceuticals 2015, 8, 366–415. [Google Scholar] [CrossRef] [PubMed]
- Czaplewski, L.; Bax, R.; Clokie, M.; Dawson, M.; Fairhead, H.; Fischetti, V.A.; Foster, S.; Gilmore, B.F.; Hancock, R.E.W.; Harper, D.; et al. Alternatives to antibiotics-a pipeline portfolio review. Lancet Infect. Dis. 2016, 16, 239–251. [Google Scholar] [CrossRef] [Green Version]
- Acedo, J.Z.; Chiorean, S.; Vederas, J.C.; van Belkum, M.J. The expanding structural variety among bacteriocins from Gram-positive bacteria. FEMS Microbiol. Rev. 2018, 42, 805–828. [Google Scholar] [CrossRef]
- Amso, Z.; Hayouka, Z. Antimicrobial random peptide cocktails: A new approach to fight pathogenic bacteria. Chem. Commun. 2019, 55, 2007–2014. [Google Scholar] [CrossRef]
- Torres, M.D.T.; Sothiselvam, S.; Lu, T.K.; de la Fuente-Nunez, C. Peptide Design Principles for Antimicrobial Applications. J. Mol. Biol. 2019, 431, 3547–3567. [Google Scholar] [CrossRef]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef]
- Shinohara, D.R.; Menegucci, T.C.; Fedrigo, N.H.; Migliorini, L.B.; Carrara-Marroni, F.E.; Maria Dos Anjos, M.; Cardoso, C.L.; Nishiyama, S.A.B.; Tognim, M.C.B. Synergistic activity of polymyxin B combined with vancomycin against carbapenem-resistant and polymyxin-resistant Acinetobacter baumannii: First in vitro study. J. Med. Microbiol. 2019, 68, 309–315. [Google Scholar] [CrossRef]
- Ofek, I.; Cohen, S.; Rahmani, R.; Kabha, K.; Tamarkin, D.; Herzig, Y.; Rubinstein, E. Antibacterial synergism of polymyxin B nonapeptide and hydrophobic antibiotics in experimental gram-negative infections in mice. Antimicrob. Agents Chemother. 1994, 38, 374–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandler, M.D.; Baidin, V.; Lee, J.; Pahil, K.S.; Owens, T.W.; Kahne, D. Novobiocin enhances polymyxin activity by stimulating lipopolysaccharide transport. J. Am. Chem. Soc. 2018, 140, 6749–6753. [Google Scholar] [CrossRef] [PubMed]
- Simmons, N.A. Colistin, sulphamethoxazole, and trimethoprim in synergy against Gram-negative bacteria. J. Clin. Pathol. 1970, 23, 757–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenblatt, J.E.; Stewart, P.R. Combined Activity of Sulfamethoxazole, Trimethoprim, and Polymyxin B against Gram-Negative Bacilli. Antimicrob. Agents Chemother. 1974, 6, 84–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nageeb, W.; Metwally, L.; Kamel, M.; Zakaria, S. In vitro antimicrobial synergy studies of carbapenem-resistant Acinetobacter baumannii isolated from intensive care units of a tertiary care hospital in Egypt. J. Infect. Public Health 2015, 8, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Abdul Rahim, N.; Cheah, S.-E.; Johnson, M.D.; Yu, H.; Sidjabat, H.E.; Boyce, J.; Butler, M.S.; Cooper, M.A.; Fu, J.; Paterson, D.L.; et al. Synergistic killing of NDM-producing MDR Klebsiella pneumoniae by two ‘old’ antibiotics—Polymyxin B and chloramphenicol. J. Antimicrob. Chemother. 2015, 70, 2589–2597. [Google Scholar] [CrossRef] [Green Version]
- Elemam, A.; Rahimian, J.; Doymaz, M. In Vitro Evaluation of Antibiotic Synergy for Polymyxin B-Resistant Carbapenemase-Producing Klebsiella pneumoniae. J. Clin. Microbiol. 2010, 48, 3558–3562. [Google Scholar] [CrossRef] [Green Version]
- Pletzer, D.; Mansour, S.C.; Hancock, R.E.W. Synergy between conventional antibiotics and anti-biofilm peptides in a murine, sub-cutaneous abscess model caused by recalcitrant ESKAPE pathogens. PLoS Pathog. 2018, 14, e1007084. [Google Scholar] [CrossRef]
- Ulvatne, H.; Karoliussen, S.; Stiberg, T.; Rekdal, Ø.; Svendsen, J.S. Short antibacterial peptides and erythromycin act synergically against Escherichia coli. J. Antimicrob. Chemother. 2001, 48, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Giacometti, A.; Cirioni, O.; Del Prete, M.S.; Paggi, A.M.; D’Errico, M.M.; Scalise, G. Combination studies between polycationic peptides and clinically used antibiotics against Gram-positive and Gram-negative bacteria. Peptides 2000, 21, 1155–1160. [Google Scholar] [CrossRef]
- Grassi, L.; Maisetta, G.; Esin, S.; Batoni, G. Combination Strategies to Enhance the Efficacy of Antimicrobial Peptides against Bacterial Biofilms. Front. Microbiol. 2017, 8, 2409–2417. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Li, Z.; Li, X.; Tian, Y.; Fan, Y.; Yu, C.; Zhou, B.; Liu, Y.; Xiang, R.; Yang, L. Synergistic effects of antimicrobial peptide DP7 combined with antibiotics against multidrug-resistant bacteria. Drug. Des. Dev. Ther. 2017, 11, 939–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollmann, A.; Martinez, M.; Maturana, P.; Semorile, L.C.; Maffia, P.C. Antimicrobial peptides: Interaction with model and biological membranes and synergism with chemical antibiotics. Front. Chem. 2018, 6, 204–217. [Google Scholar] [CrossRef] [Green Version]
- Shurko, J.F.; Galega, R.S.; Li, C.; Lee, G.C. Evaluation of LL-37 antimicrobial peptide derivatives alone and in combination with vancomycin against S. aureus. J. Antibiot. 2018, 71, 971–974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otvos, L., Jr.; Ostorhazi, E.; Szabo, D.; Zumbrun, S.D.; Miller, L.L.; Halasohoris, S.A.; Desai, P.D.; Int Veldt, S.M.; Kraus, C.N. Synergy Between Proline-Rich Antimicrobial Peptides and Small Molecule Antibiotics Against Selected Gram-Negative Pathogens in vitro and in vivo. Front. Chem. 2018, 6, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi Azad, Z.; Moravej, H.; Fasihi-Ramandi, M.; Masjedian, F.; Nazari, R.; Mirnejad, R.; Moosazadeh Moghaddam, M. In vitro synergistic effects of a short cationic peptide and clinically used antibiotics against drug-resistant isolates of Brucella melitensis. J. Med. Microbiol. 2017, 66, 919–926. [Google Scholar] [CrossRef]
- Kampshoff, F.; Willcox, M.D.P.; Dutta, D. A Pilot Study of the Synergy between Two Antimicrobial Peptides and Two Common Antibiotics. Antibiotics 2019, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, M.F.; Abdelkhalek, A.; Seleem, M.N. Evaluation of short synthetic antimicrobial peptides for treatment of drug-resistant and intracellular Staphylococcus aureus. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Zheng, W.; Sun, W.; Simeonov, A. Drug repurposing screens and synergistic drug-combinations for infectious diseases. Br. J. Pharm. 2018, 175, 181–191. [Google Scholar] [CrossRef]
- Li, Y.; Lin, X.; Yao, X.; Huang, Y.; Liu, W.; Ma, T.; Fang, B. Synergistic Antimicrobial Activity of Colistin in Combination with Rifampin and Azithromycin against Escherichia coli Producing MCR-1. Antimicrob. Agents Chemother. 2018, 62, 18–28. [Google Scholar] [CrossRef] [Green Version]
- Stach, M.; Maillard, N.; Kadam, R.U.; Kalbermatter, D.; Meury, M.; Page, M.G.P.; Fotiadis, D.; Darbre, T.; Reymond, J.-L. Membrane disrupting antimicrobial peptide dendrimers with multiple amino termini. MedChemComm 2012, 3, 86–89. [Google Scholar] [CrossRef]
- Stach, M.; Siriwardena, T.N.; Köhler, T.; van Delden, C.; Darbre, T.; Reymond, J.-L. Combining Topology and Sequence Design for the Discovery of Potent Antimicrobial Peptide Dendrimers against Multidrug-Resistant Pseudomonas aeruginosa. Angew. Chem. Int. Ed. 2014, 53, 12827–12831. [Google Scholar] [CrossRef] [PubMed]
- Pires, J.; Siriwardena, T.N.; Stach, M.; Tinguely, R.; Kasraian, S.; Luzzaro, F.; Leib, S.L.; Darbre, T.; Reymond, J.-L.; Endimiani, A. In Vitro Activity of the Novel Antimicrobial Peptide Dendrimer G3KL against Multidrug-Resistant Acinetobacter baumannii and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2015, 59, 7915–7918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Sayed, P.; Kaeppeli, A.; Siriwardena, T.; Darbre, T.; Perron, K.; Jafari, P.; Reymond, J.-L.; Pioletti, D.P.; Applegate, L.A. Anti-Microbial Dendrimers against Multidrug-Resistant P. aeruginosa Enhance the Angiogenic Effect of Biological Burn-wound Bandages. Sci. Rep. 2016, 6, 23872–23882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pompilio, A.; Geminiani, C.; Mantini, P.; Siriwardena, T.N.; Bonaventura, I.D.; Reymond, J.L.; Bonaventura, G.D. Peptide dendrimers as “lead compounds” for the treatment of chronic lung infections by Pseudomonas aeruginosa in cystic fibrosis patients: In vitro and in vivo studies. Infect. Drug. Resist. 2018, 11, 1767–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Liu, Y.; Ma, Y.; Zhang, M.; He, Z.; Siriwardena, T.N.; Xu, H.; Bai, Y.; Zhang, X.; Reymond, J.-L.; et al. Peptide dendrimers G3KL and TNS18 inhibit Pseudomonas aeruginosa biofilms. Appl. Microbiol. Biotechnol. 2019, 103, 5821–5830. [Google Scholar] [CrossRef]
- Gan, B.-H.; Siriwardena, T.N.; Javor, S.; Darbre, T.; Reymond, J.-L. Fluorescence Imaging of Bacterial Killing by Antimicrobial Peptide Dendrimer G3KL. ACS Infect. Dis. 2019, 5, 2164–2173. [Google Scholar] [CrossRef]
- Jeddou, F.B.; Falconnet, L.; Luscher, A.; Siriwardena, T.; Reymond, J.-L.; Delden, C.; van Köhler, T. Adaptive and Mutational Responses to Peptide Dendrimer Antimicrobials in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2020, 64, e02040-19. [Google Scholar]
- Doern, C.D. When Does 2 Plus 2 Equal 5? A Review of Antimicrobial Synergy Testing. J. Clin. Microbiol. 2014, 52, 4124–4128. [Google Scholar] [CrossRef] [Green Version]
- Grela, E.; Kozłowska, J.; Grabowiecka, A. Current methodology of MTT assay in bacteria—A review. Acta Histochem. 2018, 120, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Renneberg, J. Definitions of antibacterial interactions in animal infection models. J. Antimicrob. Chemother. 1993, 31, 167–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roell, K.R.; Reif, D.M.; Motsinger-Reif, A.A. An Introduction to Terminology and Methodology of Chemical Synergy—Perspectives from Across Disciplines. Front. Pharm. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Gopal, R.; Kim, Y.G.; Lee, J.H.; Lee, S.K.; Chae, J.D.; Son, B.K.; Seo, C.H.; Park, Y. Synergistic Effects and Antibiofilm Properties of Chimeric Peptides against Multidrug-Resistant Acinetobacter baumannii Strains. Antimicrob. Agents Chemother. 2014, 58, 1622–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Netuschil, L.; Auschill, T.M.; Sculean, A.; Arweiler, N.B. Confusion over live/dead stainings for the detection of vital microorganisms in oral biofilms—Which stain is suitable? BMC Oral Health 2014, 14, 2–14. [Google Scholar] [CrossRef] [Green Version]
- Deris, Z.Z.; Swarbrick, J.D.; Roberts, K.D.; Azad, M.A.K.; Akter, J.; Horne, A.S.; Nation, R.L.; Rogers, K.L.; Thompson, P.E.; Velkov, T.; et al. Probing the Penetration of Antimicrobial Polymyxin Lipopeptides into Gram-Negative Bacteria. Bioconjugate Chem. 2014, 25, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Velkov, T.; Thompson, P.E.; Nation, R.L.; Li, J. Structure-Activity Relationships of Polymyxin Antibiotics. J. Med. Chem. 2010, 53, 1898–1916. [Google Scholar] [CrossRef] [Green Version]
- Watanakunakorn, C. Mode of action and in-vitro activity of vancomycin. J. Antimicrob. Chemother. 1984, 14, 7–18. [Google Scholar] [CrossRef]
- Mao, J.C.-H.; Putterman, M. Accumulation in Gram-positive and Gram-negative Bacteria as a Mechanism of Resistance to Erythromycin. J. Bacteriol. 1968, 95, 1111–1117. [Google Scholar] [CrossRef] [Green Version]
- Salter, A.J. Overview. Trimethoprim-Sulfamethoxazole: An Assessment of More Than 12 Years of Use. Rev. Infect. Dis. 1982, 4, 196–236. [Google Scholar] [CrossRef]
- Smith, D.H.; Davis, B.D. Mode of Action of Novobiocin in Escherichia coli. J. Bacteriol. 1967, 93, 71–79. [Google Scholar] [CrossRef] [Green Version]
- May, J.M.; Owens, T.W.; Mandler, M.D.; Simpson, B.W.; Lazarus, M.B.; Sherman, D.J.; Davis, R.M.; Okuda, S.; Massefski, W.; Ruiz, N.; et al. The antibiotic novobiocin binds and activates the ATPase that powers lipopolysaccharide transport. J. Am. Chem. Soc. 2017, 139, 17221–17224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hooper, D.C.; Wolfson, J.S.; Ng, E.Y.; Swartz, M.N. Mechanisms of action of and resistance to ciprofloxacin. Am. J. Med. 1987, 82, 12–20. [Google Scholar] [PubMed]
- Bergmann, E.D.; Sicher, S. Mode of Action of Chloramphenicol. Nature 1952, 170, 931–932. [Google Scholar] [CrossRef] [PubMed]
- Hahn, F.E.; Sarre, S.G. Mechanism of Action of Gentamicin. J. Infect. Dis. 1969, 119, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Retsema, J.; Girard, A.; Schelkly, W.; Manousos, M.; Anderson, M.; Bright, G.; Borovoy, R.; Brennan, L.; Mason, R. Spectrum and mode of action of azithromycin (CP-62,993), a new 15-membered-ring macrolide with improved potency against gram-negative organisms. Antimicrob. Agents Chemother. 1987, 31, 1939–1947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tipper, D.J. Mode of action of β-lactam antibiotics. Pharm. Ther. 1985, 27, 1–35. [Google Scholar] [CrossRef]
- Okamoto, K.; Gotoh, N.; Nishino, T. Pseudomonas aeruginosa Reveals High Intrinsic Resistance to Penem Antibiotics: Penem Resistance Mechanisms and Their Interplay. Antimicrob. Agents Chemother. 2001, 45, 1964–1971. [Google Scholar] [CrossRef] [Green Version]
- Siriwardena, T.N.; Stach, M.; He, R.; Gan, B.; Javor, S.; Heitz, M.; Cai, X.; Chen, P.; Wei, D.; Li, H.; et al. Lipidated Peptide Dendrimers Killing Multidrug-Resistant Bacteria. J. Am. Chem. Soc. 2018, 140, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Siriwardena, T.N.; Capecchi, A.; Gan, B.-H.; Jin, X.; He, R.; Wei, D.; Ma, L.; Köhler, T.; van Delden, C.; Javor, S.; et al. Optimizing Antimicrobial Peptide Dendrimers in Chemical Space. Angew. Chem. Int. Ed. 2018, 57, 8483–8487. [Google Scholar] [CrossRef]
- Siriwardena, T.N.; Lüscher, A.; Köhler, T.; van Delden, C.; Javor, S.; Reymond, J.-L. Antimicrobial Peptide Dendrimer Chimera. Helv. Chim. Acta 2019, 102, e1900034. [Google Scholar] [CrossRef]
- Lenhard, J.R.; Nation, R.L.; Tsuji, B.T. Synergistic combinations of polymyxins. Int. J. Antimicrob. Agents 2016, 48, 607–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaara, M. Polymyxin Derivatives that Sensitize Gram-Negative Bacteria to Other Antibiotics. Molecules 2019, 24, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, A.G.; Butterworth, D.; Cole, M.; Hanscomb, G.; Hood, J.D.; Reading, C.; Rolinson, G.N. Naturally-Occurring β-Lactamase Inhibitors with Antibacterial Activity. J. Antibiot. 1976, 29, 668–669. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.K.; Sheehan, J.P.; Bratton, B.P.; Moore, G.M.; Mateus, A.; Li, S.H.-J.; Kim, H.; Rabinowitz, J.D.; Typas, A.; Savitski, M.M.; et al. A Dual-Mechanism Antibiotic Kills Gram-Negative Bacteria and Avoids Drug Resistance. Cell 2020, 181, 1518–1532. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. aeruginosa PAO1 | E. coli W3110 | A. baumannii ATCC 19606 | K. pneumoniae NCTC 418 | MRSA COL | |
|---|---|---|---|---|---|
| G3KLa | 4 | 4 | 8 | >64 | >64 |
| PIa | ≥500 | 62.5–250 | >500 | ≥500 | 62.5 |
| PMBa | 0.25 | 0.25 | 0.125 | 0.125 | 64 |
| G3KLcomb/PIcomb (FICi) b | 2/8 (0.5) | 2/31.3 (0.6) | 4/15.6 (<0.53) | >64/>500 (-) | >64/62.5 (>2) |
| PMBcomb/PIcomb (FICi) b | 0.125/62.5 (0.6) | 0.125/31.25 (0.6) | 0.063/125 (<0.8) | 2/16 (>4) | 16/31.25 (0.8) |
| P. aeruginosa PAO1 | K. pneumoniae NCTC418 | MRSA COL | |
|---|---|---|---|
| G3KL | 4–8 | >64 | >64 |
| Vancomycin | 256 | 256 | 0.5 |
| G3KLcomb/Vancomycincomb (FICi) b | 1/32 (0.6) | 16/16 (<0.3) | >64/0.5 (>2) |
| Erythromycin | 128 | 64 | 0.5 |
| G3KLcomb/Erythromycincomb (FICi) b | 2/64 (1) | 8/8 (<0.3) | 32/0.25 (<1) |
| Ampicillin | >256 | >256 | 128 |
| G3KLcomb/Ampicillincomb (FICi) b | 4/>256 (>2) | >64/>256 (-) | 32/64 (<1) |
| Novobiocin | >256 | 16 | 0.31 |
| G3KLcomb/Novobiocincomb (FICi) b | 2/256 (<1.5) | 8/1 (<0.2) | >64/0.31 (>2) |
| Ciprofloxacin | 0.125 | 0.031 | 0.25 |
| G3KLcomb/Ciprofloxacincomb (FICi) b | 4/0.125 (2) | >32/0.031 (>2) | >64/0.25 (>2) |
| Chloramphenicol | 8 | 8 | 8 |
| G3KLcomb/Chloramphenicolcomb (FICi) b | 2/1 (0.6) | 16/2 (<0.5) | >64/8 (>2) |
| Gentamicin | 1 | 2 | 0.5 |
| G3KLcomb/Gentamicincomb (FICi) b | 4/1 (2) | 32/0.25 (<0.6) | 64/0.25 (<1.3) |
| Azithromycin | 64 | 4 | 4 |
| G3KLcomb/Azithromycincomb (FICi) b | 8/0.5 (0.5) | 8/1 (<0.4) | 16/2 (<0.8) |
| Sulfamethoxazole | 256 | >256 | >32 |
| G3KLcomb/Sulfamethoxazolecomb (FICi) b | 1/128 (1) | >64/>256 (-) | >64/>32 (-) |
| Trimethoprim | 128 | >256 | >32 |
| G3KLcomb/Trimethoprimcomb (FICi) b | 4/128 (2) | 32/8 (<0.5) | 64/32 (<1.5) |
Sample Availability: Samples for the compounds are accessible as described in the methods Section 3.1. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gan, B.-H.; Cai, X.; Javor, S.; Köhler, T.; Reymond, J.-L. Synergistic Effect of Propidium Iodide and Small Molecule Antibiotics with the Antimicrobial Peptide Dendrimer G3KL against Gram-Negative Bacteria. Molecules 2020, 25, 5643. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25235643
Gan B-H, Cai X, Javor S, Köhler T, Reymond J-L. Synergistic Effect of Propidium Iodide and Small Molecule Antibiotics with the Antimicrobial Peptide Dendrimer G3KL against Gram-Negative Bacteria. Molecules. 2020; 25(23):5643. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25235643
Chicago/Turabian StyleGan, Bee-Ha, Xingguang Cai, Sacha Javor, Thilo Köhler, and Jean-Louis Reymond. 2020. "Synergistic Effect of Propidium Iodide and Small Molecule Antibiotics with the Antimicrobial Peptide Dendrimer G3KL against Gram-Negative Bacteria" Molecules 25, no. 23: 5643. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25235643