
Glossogyne tenuifolia Attenuates Proliferation and Migration of Vascular Smooth Muscle Cells

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
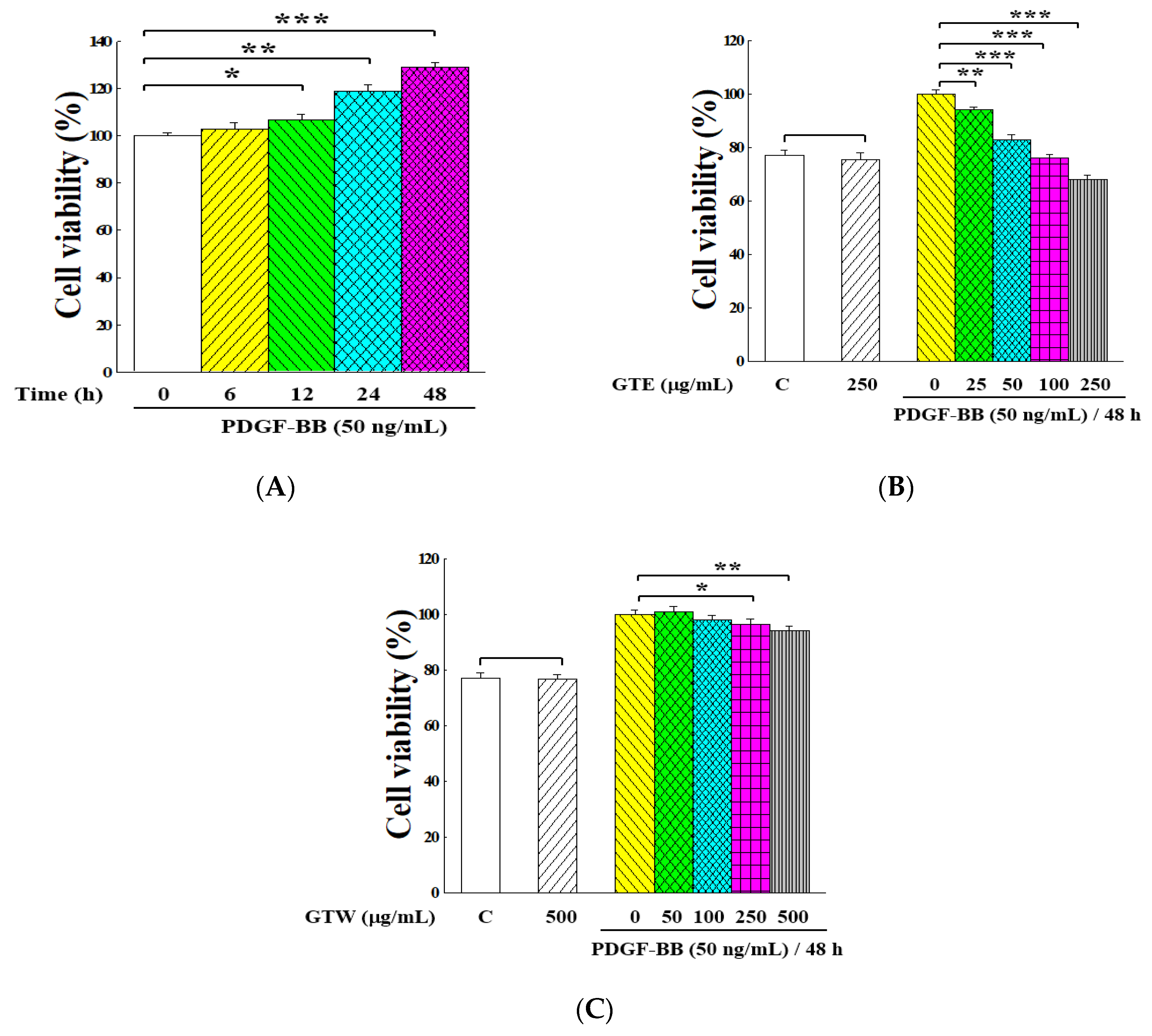
2.1. Inhibitory Effects of GT Extracts on Proliferation of PDGF-BB-Stimulated VSMCs
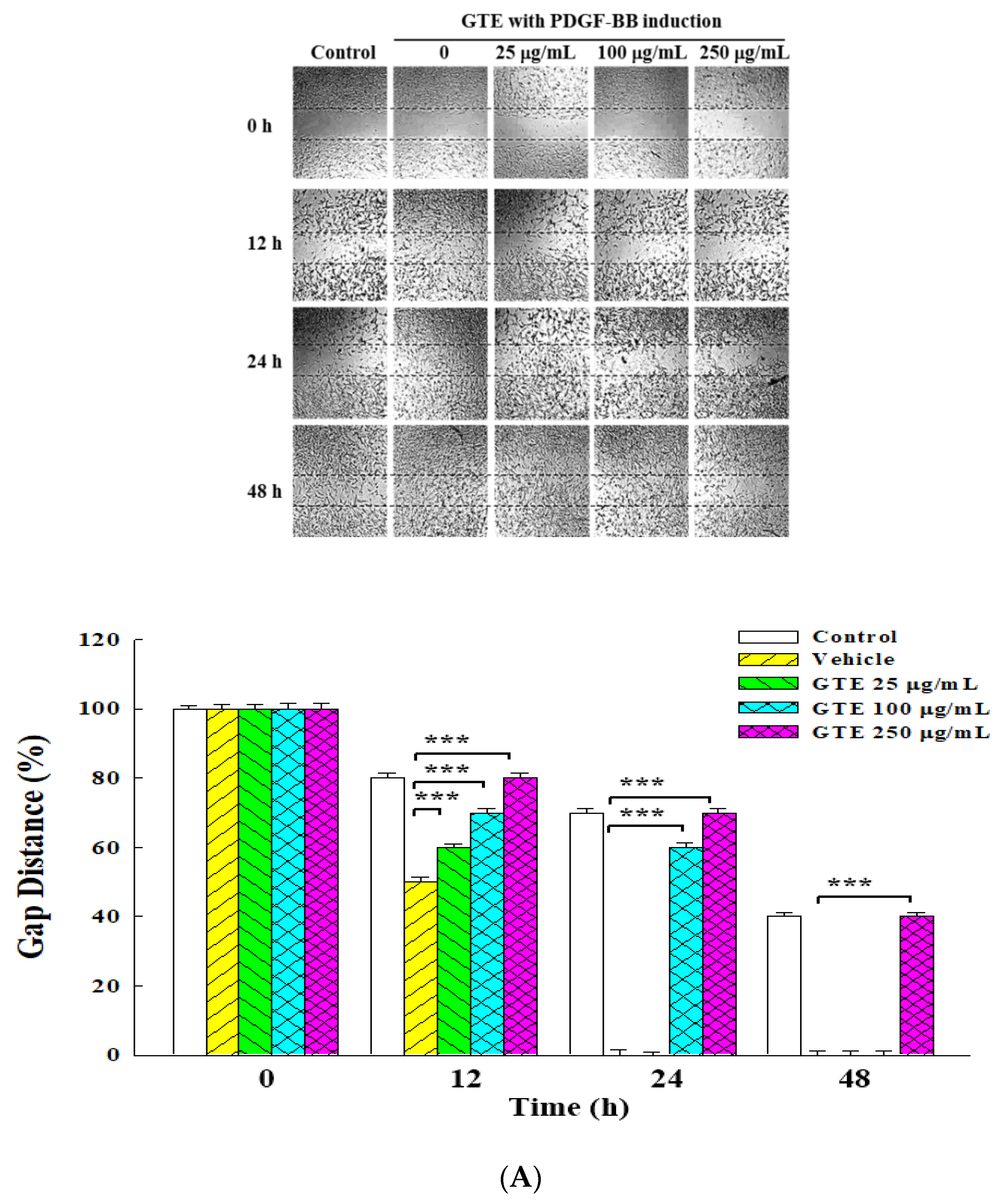
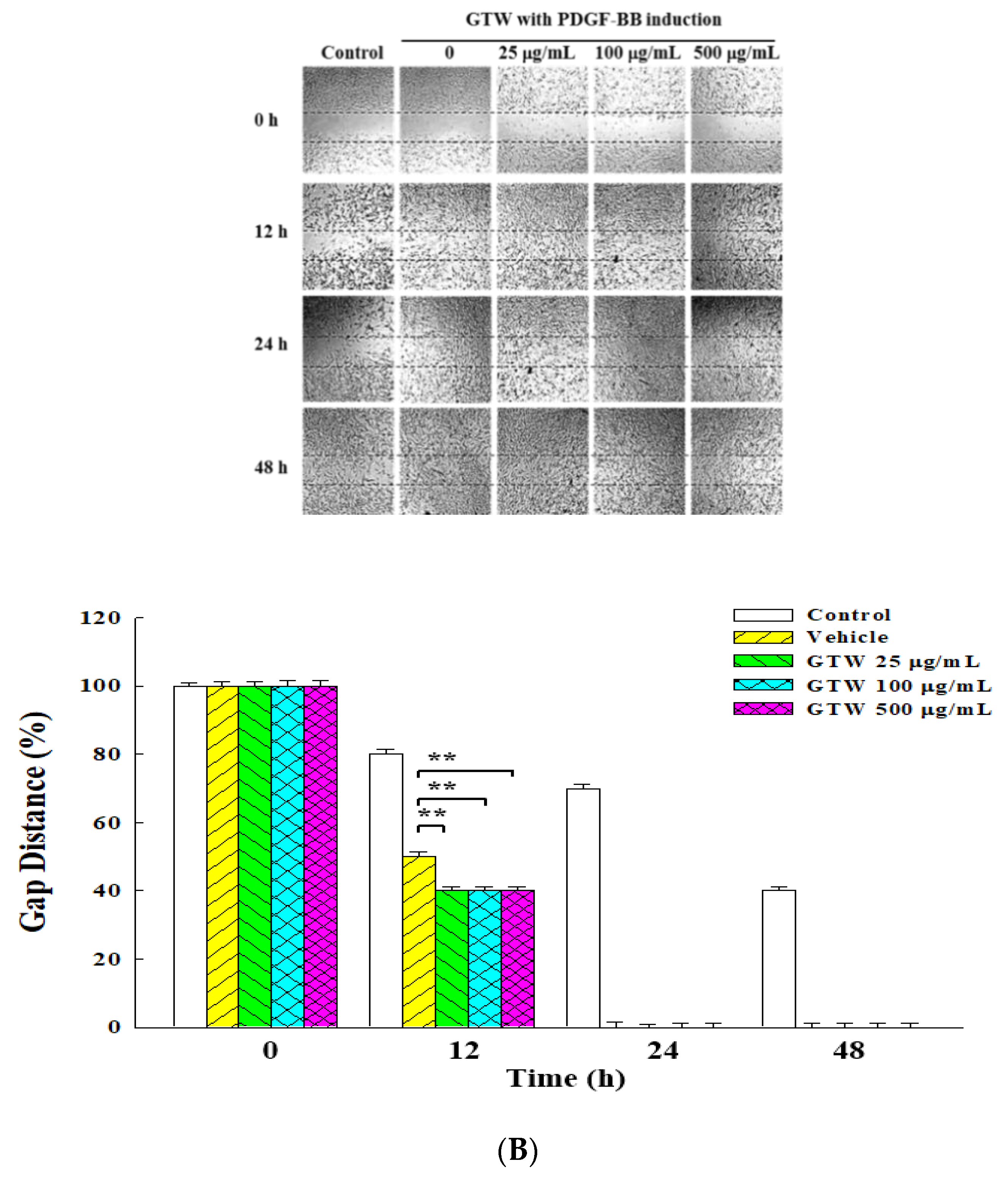
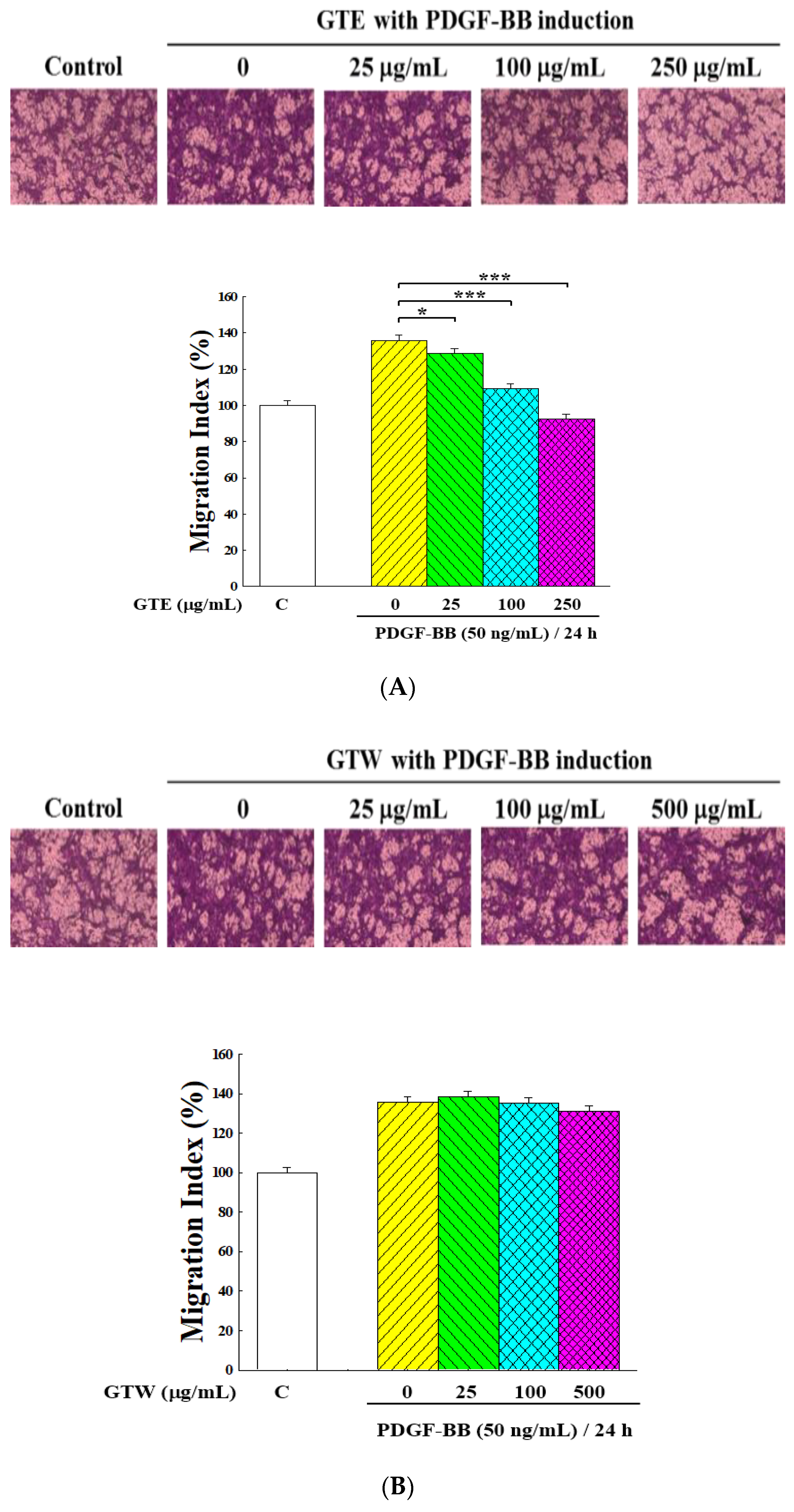
2.2. Inhibitory Effects of GT Extracts on Migration of PDGF-BB-Stimulated VSMCs
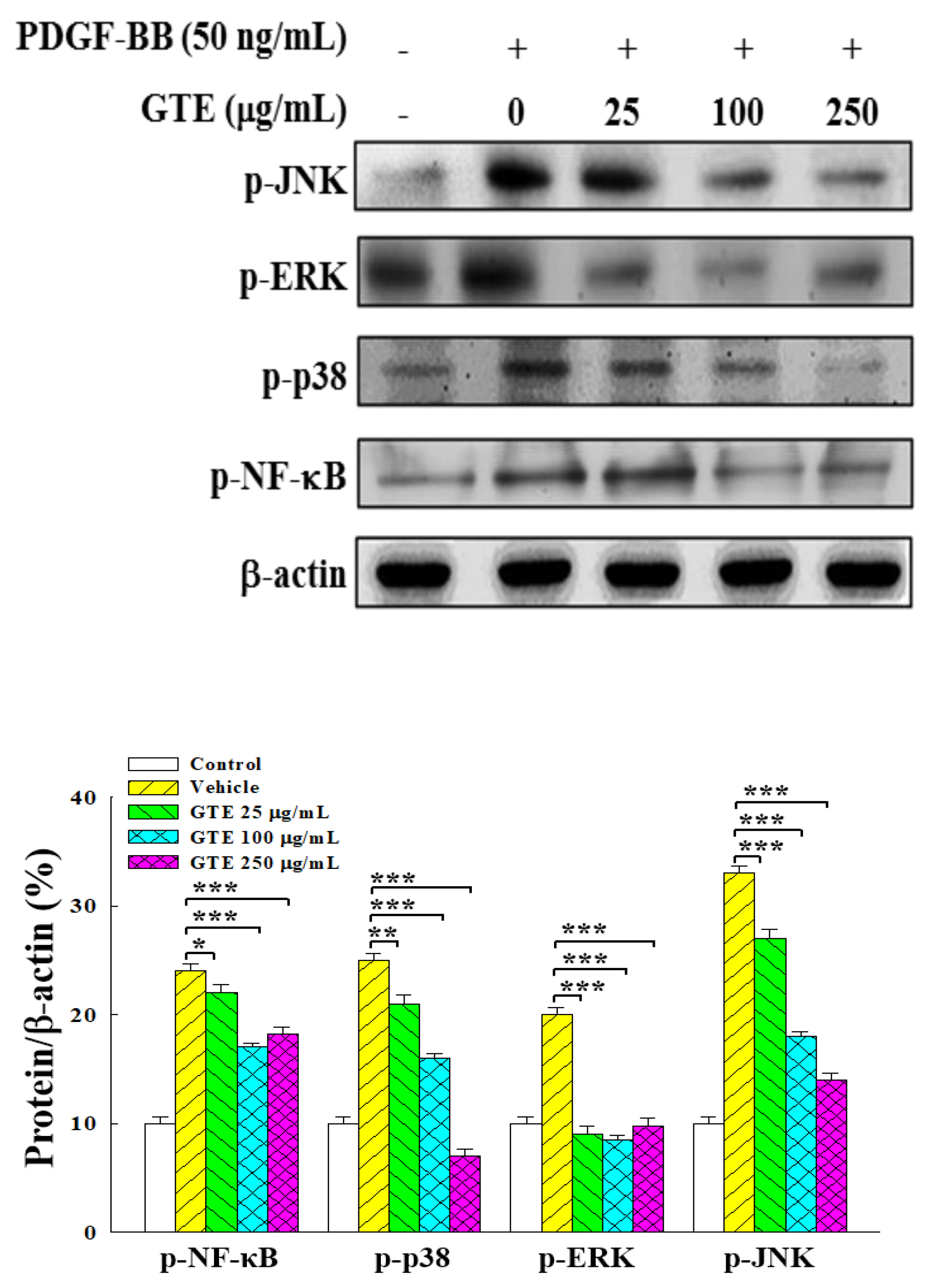
2.3. GTE Suppresses NF-κB and MAPK Signaling Pathways in PDGF-BB-Stimulated VSMCs
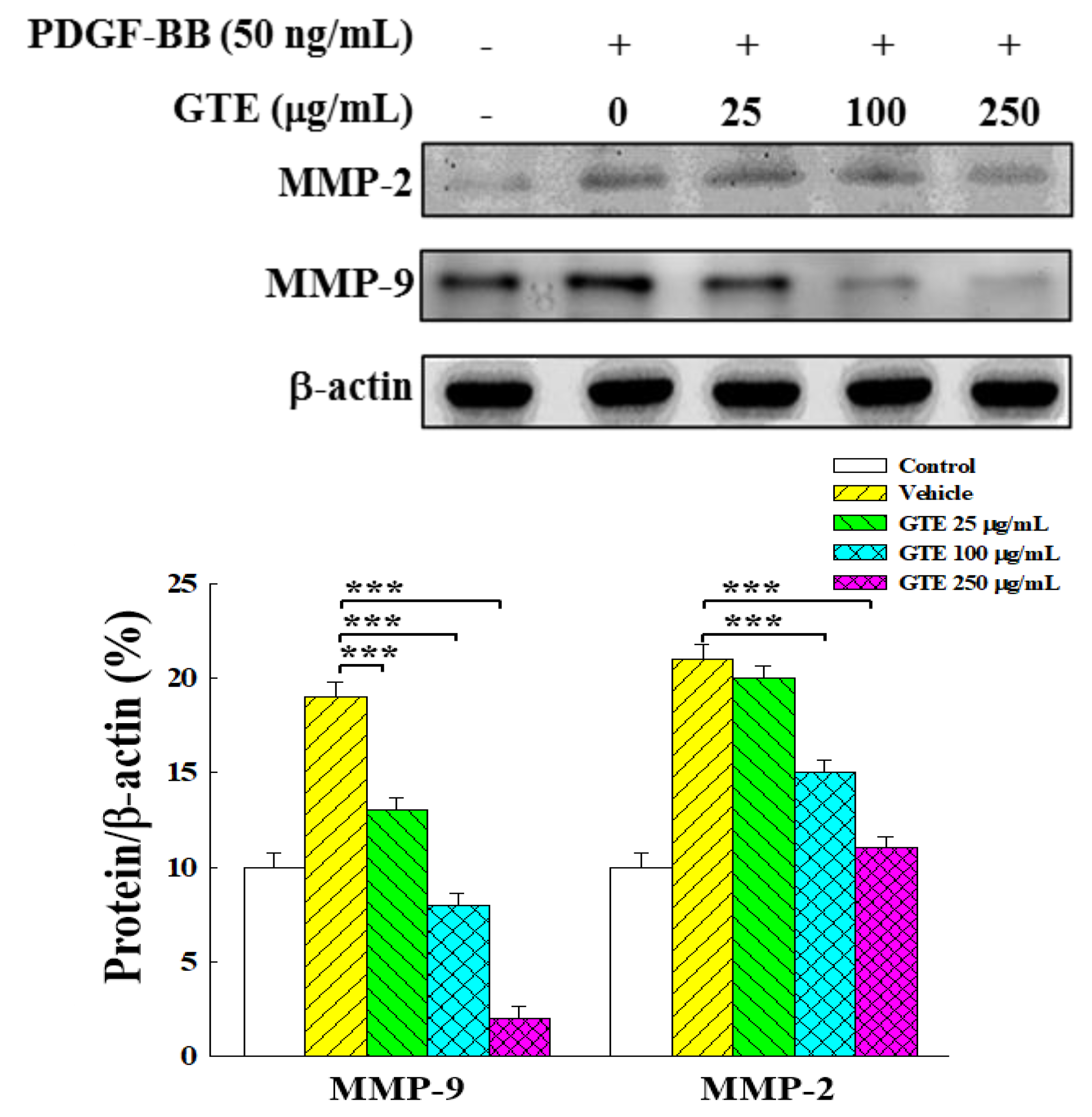
2.4. GTE Inhibits VSMC Migration by Suppressing the Expression of MMP-2 and MMP-9
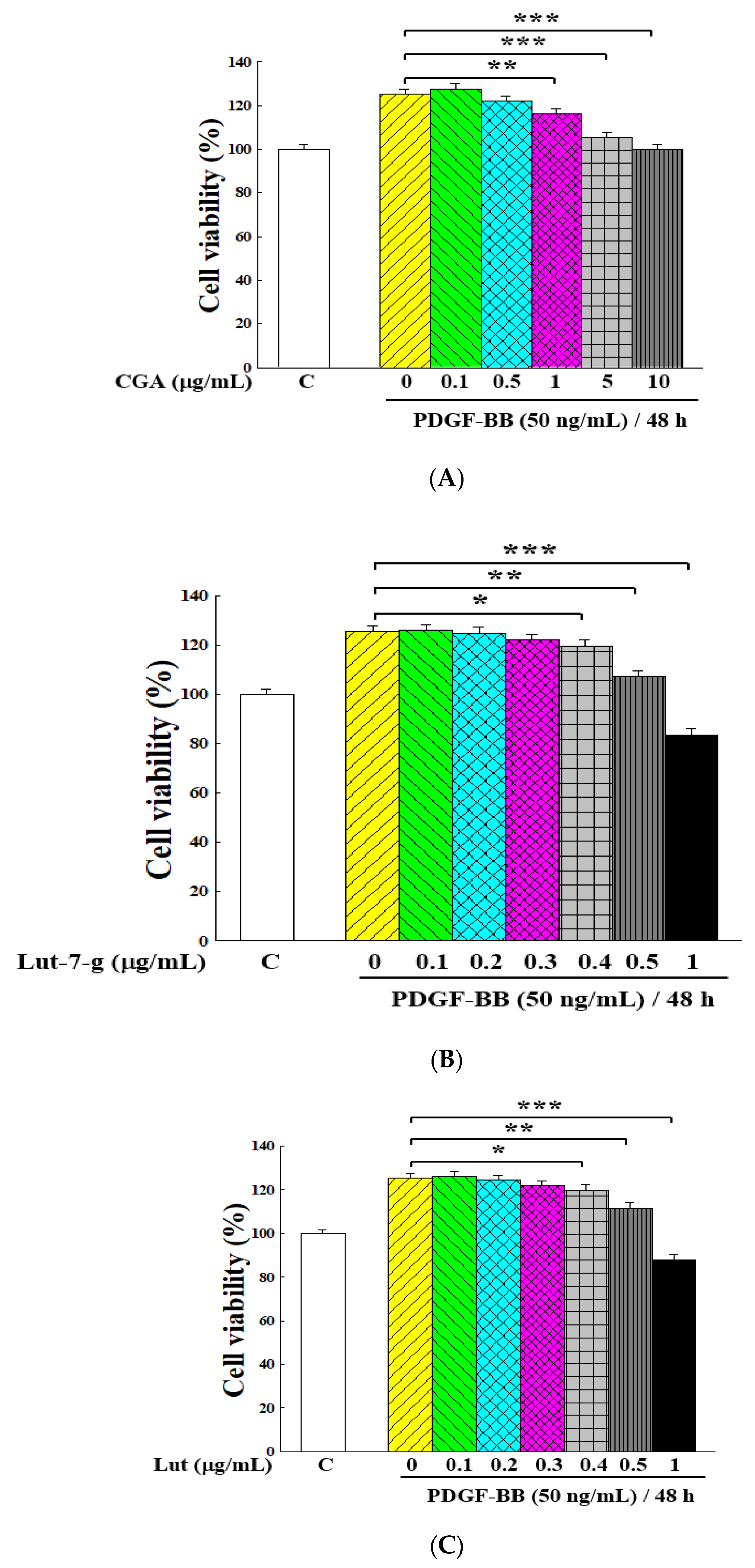
2.5. Inhibitory Effects of Main Ingredients on Proliferation of PDGF-BB-Stimulated VSMCs
3. Discussion
4. Materials and Methods
4.1. Preparation of GT Extracts
4.2. Cell Culture
4.3. Cell Migration by Wound Healing Assay
4.4. Cell Migration Assay by Boyden Chamber
4.5. Western Blot Analysis
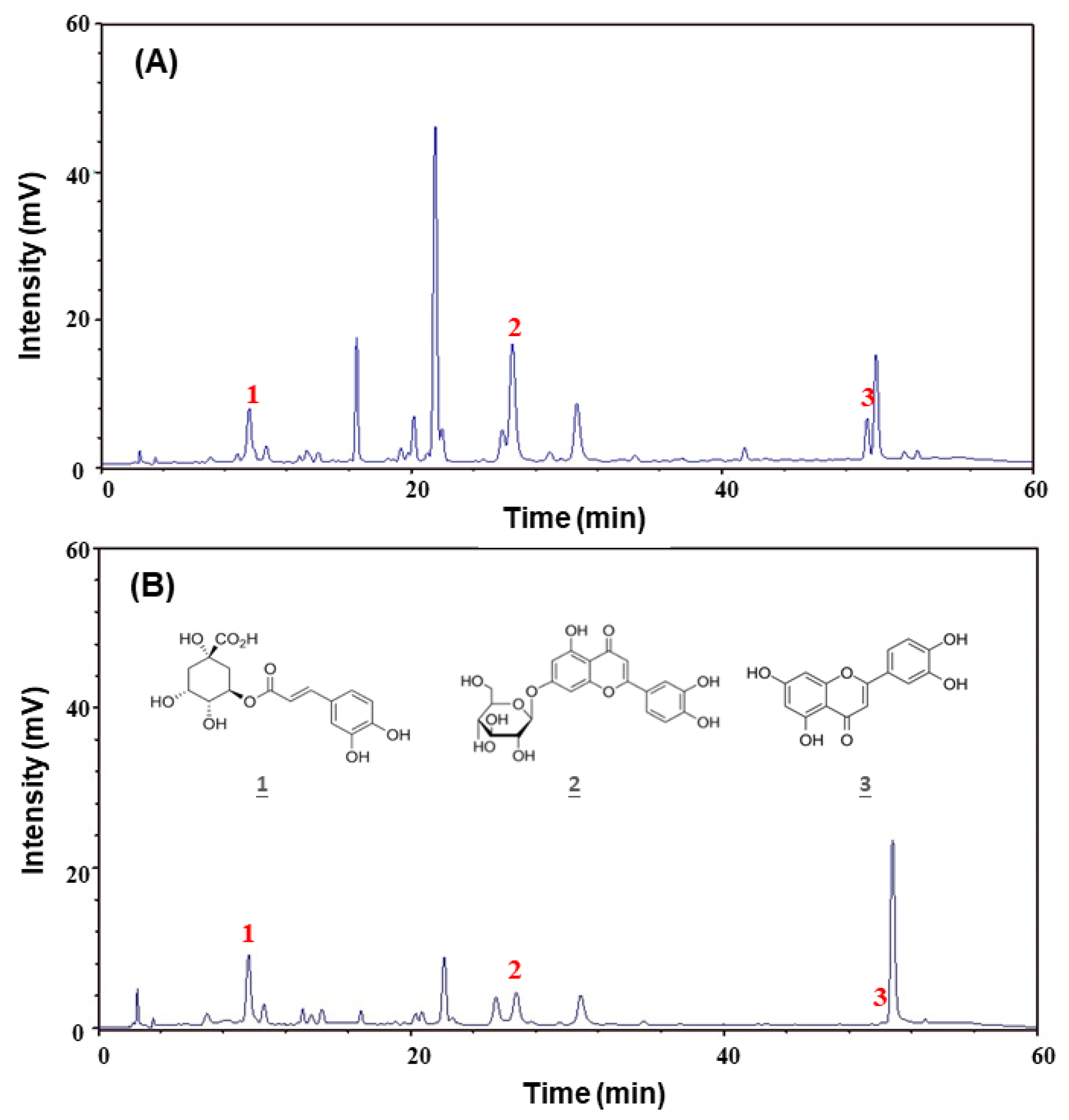
4.6. Determination of the Main Ingredients Amounts by HPLC
4.7. Determination of Total Polyphenols Content and Total Flavonoids Content
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chen, Y.; Su, X.; Qin, Q.; Yu, Y.; Jia, M.; Zhang, H.; Li, H.; Pei, L. New insights into phenotypic switching of VSMCs induced by hyperhomocysteinemia: Role of endothelin-1 signaling. Biomed. Pharmacother. 2020, 123, 109758. [Google Scholar] [CrossRef] [PubMed]
- Hansson, G.K.; Hermansson, A. The immune system in atherosclerosis. Nat. Immunol. 2011, 12, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and challenges in translating the biology of atherosclerosis. Nature 2011, 473, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Basatemur, G.L.; Jørgensen, H.F.; Clarke, M.C.H.; Bennett, M.R.; Mallat, Z. Vascular smooth muscle cells in atherosclerosis. Nat. Rev. Cardiol. 2019, 16, 727–744. [Google Scholar] [CrossRef]
- Kang, H.; Ahn, D.H.; Pak, J.H.; Seo, K.H.; Baek, N.I.; Jang, S.W. Magnobovatol inhibits smooth muscle cell migration by suppressing PDGF-Rβ phosphorylation and inhibiting matrix metalloproteinase-2 expression. Int. J. Mol. Med. 2016, 37, 1239–1246. [Google Scholar] [CrossRef] [Green Version]
- Rudijanto, A. The role of vascular smooth muscle cells on the pathogenesis of atherosclerosis. Acta Med. Indones. 2007, 39, 86–93. [Google Scholar]
- Newby, A.C.; Zaltsman, A.B. Molecular mechanisms in intimal hyperplasia. J. Pathol. 2000, 190, 300–309. [Google Scholar] [CrossRef]
- Raines, E.W. PDGF and cardiovascular disease. Cytokine Growth Factor Rev. 2004, 15, 237–254. [Google Scholar] [CrossRef]
- Rubin, K.; Tingström, A.; Hansson, G.K.; Larsson, E.; Rönnstrand, L.; Klareskog, L.; Claesson-Welsh, L.; Heldin, C.H.; Fellström, B.; Terracio, L. Induction of B-type receptors for platelet-derived growth factor in vascular inflammation: Possible implications for development of vascular proliferative lesions. Lancet 1988, 1, 1353–1356. [Google Scholar] [CrossRef]
- Hellstrom, M.; Kalen, M.; Lindahl, P.; Abramsson, A.; Betsholtz, C. Role of PDGF-B and PDGFR-β in recruitment of vascular smooth muscle cells and pericytes during embryonic blood vessel formation in the mouse. Development 1999, 126, 3047–3055. [Google Scholar]
- Lee, C.K.; Lee, H.M.; Kim, H.J.; Park, H.J.; Won, K.J.; Roh, H.Y.; Choi, W.S.; Jeon, B.H.; Park, T.K.; Kim, B. Syk contributes to PDGF-BB-mediated migration of rat aortic smooth muscle cells via MAPK pathways. Cardiovasc. Res. 2007, 74, 159–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heldin, C.H.; Westermark, B. Mechanism of action and in vivo role of platelet-derived growth factor. Physiol. Rev. 1999, 79, 1283–1316. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.; Kim, S.; Izumi, Y.; Izumiya, Y.; Nakao, T.; Miyazaki, H.; Iwao, H. Role of JNK, p38, and ERK in platelet-derived growth factor-induced vascular proliferation, migration, and gene expression. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 795–801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, Y.; Dolence, J.; Ramanan, S.; Ren, J.; Nair, S. Bisdemethoxycurcumin inhibits PDGF-induced vascular smooth muscle cell motility and proliferation. Mol. Nutr. Food Res. 2013, 57, 1611–1618. [Google Scholar] [CrossRef] [PubMed]
- Song, I.S.; Jeong, Y.J.; Park, J.H.; Shim, S.; Jang, S.W. Chebulinic acid inhibits smooth muscle cell migration by suppressing PDGF-Rβ phosphorylation and inhibiting matrix metalloproteinase-2 expression. Sci. Rep. 2017, 7, 11797. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Li, Y.; Xie, H.; Chen, J.; Liu, S. Curcumin inhibits proliferation, migration and neointimal formation of vascular smooth muscle via activating miR-22. Pharm. Biol. 2020, 58, 610–619. [Google Scholar] [CrossRef] [PubMed]
- Nelson, A.R.; Fingleton, B.; Rothenberg, M.L.; Matrisian, L.M. Matrix metalloproteinases: Biologic activity and clinical implications. J. Clin. Oncol. 2000, 18, 1135–1149. [Google Scholar] [CrossRef]
- Newby, A.C. Matrix metalloproteinases regulate migration, proliferation, and death of vascular smooth muscle cells by degrading matrix and non-matrix substrates. Cardiovasc. Res. 2006, 69, 614–624. [Google Scholar] [CrossRef]
- Hsieh, W.C.; Hsieh, M.T.; Chiu, N.Y.; Lin, J.K. Investigation and study on the resources of Chinese crude drugs in Taiwan (VI): Investigation and study on the medicinal plants in Penghu County. Annu. Rep. Chin. Med. 1999, 17, 245–407. [Google Scholar]
- Hsu, H.F.; Houng, J.Y.; Chang, C.L.; Wu, C.C.; Chang, F.R.; Wu, Y.C. Antioxidant activity, cytotoxicity, and DNA information of Glossogyne tenuifolia. J. Agric. Food Chem. 2005, 53, 6117–6125. [Google Scholar] [CrossRef]
- Wu, M.J.; Huang, C.L.; Lian, T.W.; Kou, M.C.; Wang, L. Antioxidant activity of Glossogyne tenuifolia. J. Agric. Food Chem. 2005, 53, 6305–6312. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.J.; Weng, C.Y.; Ding, H.Y.; Wu, P.J. Anti-inflammatory and antiviral effects of Glossogyne tenuifolia. Life Sci. 2005, 76, 1135–1146. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.W.; Kuo, H.C.; Hsu, H.F.; Tu, Y.K.; Cheng, T.T.; Houng, J.Y. Inhibitory activity on RANKL-mediated osteoclastogenesis of Glossogyne tenuifolia extract. J. Funct. Foods 2014, 6, 215–223. [Google Scholar] [CrossRef]
- Ha, C.L.; Weng, C.Y.; Wang, L.; Lian, T.W.; Wu, M.J. Immunomodulatory effect of Glossogyne tenuifolia in murine peritoneal macrophages and splenocytes. J. Ethnopharmacol. 2006, 107, 116–125. [Google Scholar] [CrossRef]
- Wang, C.P.; Houng, J.Y.; Hsu, H.F.; Chen, H.J.; Huang, B.; Hung, W.C.; Yu, T.H.; Chiu, C.N.; Lu, L.F.; Hsu, C.C. Glossogyne tenuifolia enhances posttranslational S-nitrosylation of proteins in vascular endothelial cells. Taiwania 2011, 56, 97–104. [Google Scholar]
- Wu, M.J.; Wang, L.; Ding, H.Y.; Weng, C.Y.; Yen, J.H. Glossogyne tenuifolia acts to inhibit inflammatory mediator production in a macrophage cell line by downregulating LPS-induced NF-κB. J. Biomed. Sci. 2004, 11, 186–199. [Google Scholar] [CrossRef]
- Hsuan, C.F.; Hsu, H.F.; Tseng, W.K.; Lee, T.L.; Wei, Y.F.; Hsu, K.L.; Wu, C.C.; Houng, J.Y. Glossogyne tenuifolia extract inhibits TNF-α-induced expression of adhesion molecules in human umbilical vein endothelial cells via blocking the NF-κB signaling pathway. Molecules 2015, 20, 16908–16923. [Google Scholar] [CrossRef] [Green Version]
- Yu, Q.; Zeng, K.; Ma, X.; Song, F.; Jiang, Y.; Tu, P.; Wang, X. Resokaempferol-mediated anti-inflammatory effects on activated macrophages via the inhibition of JAK2/STAT3, NF-κB and JNK/p38 MAPK signaling pathways. Int. Immunopharmacol. 2016, 38, 104–114. [Google Scholar] [CrossRef]
- Pan, J.X. LncRNA H19 promotes atherosclerosis by regulating MAPK and NF-kB signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 322–328. [Google Scholar]
- Tsai, Y.D.; Hsu, H.F.; Chen, Z.H.; Wang, Y.T.; Huang, S.H.; Chen, H.J.; Wang, P.W.; Wang, S.W.; Chang, C.C.; Houng, J.Y. Antioxidant, anti-inflammatory, and anti-proliferative activities of extracts from different parts of farmed and wild Glossogyne tenuifolia. Ind. Crop. Prod. 2014, 57, 98–105. [Google Scholar] [CrossRef]
- Johnson, J.L.; Dwivedi, A.; Somerville, M.; George, S.J.; Newby, A.C. Matrix metalloproteinase (MMP)-3 activates MMP-9 mediated vascular smooth muscle cell migration and neointima formation in mice. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 35–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, M.R.; Sinha, S.; Owens, G.K. Vascular smooth muscle cells in atherosclerosis. Circ. Res. 2016, 118, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Kim, J.E.; Kim, H.; Chang, H.R.; Lee, D.W.; Park, W.H. Bo-Gan-Whan regulates proliferation and migration of vascular smooth muscle cells. BMC Complement. Altern. Med. 2016, 16, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Ding, Y.; Tao, W.; Zhang, W.; Liang, T.; Liu, C. Naringenin inhibits TNF-α induced VSMC proliferation and migration via induction of HO-1. Food Chem. Toxicol. 2012, 50, 3025–3031. [Google Scholar] [CrossRef] [PubMed]
- Bargou, R.C.; Emmerich, F.; Krappmann, D.; Bommert, K.; Mapara, M.Y.; Arnold, W.; Royer, H.D.; Grinstein, E.; Greiner, A.; Scheidereit, B.; et al. Constitutive nuclear factor-κB-RelA activation is required for proliferation and survival of Hodgkin’s disease tumor cells. J. Clin. Investig. 1997, 100, 2961–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landry, D.B.; Couper, L.L.; Bryant, S.R.; Lindner, V. Activation of the NF-κB and IκB system in smooth muscle cells after rat arterial injury. Induction of vascular cell adhesion molecule-1 and monocyte chemoattractant protein-1. Am. J. Pathol. 1997, 151, 1085–1095. [Google Scholar]
- Mehrhof, F.B.; Schmidt-Ullrich, R.; Dietz, R.; Scheidereit, C. Regulation of vascular smooth muscle cell proliferation: Role of NF-κB revisited. Circ. Res. 2005, 96, 958–964. [Google Scholar] [CrossRef] [Green Version]
- Xia, Z.; Dickens, M.; Raingeaud, J.; Davis, R.J.; Greenberg, M.E. Opposing effects of ERK and JNK-p38 MAP kinases on apoptosis. Science 1995, 270, 1326–1331. [Google Scholar] [CrossRef]
- Graf, K.; Xi, X.P.; Yang, D.; Fleck, E.; Hsueh, W.A.; Law, R.E. Mitogen-activated protein kinase activation is involved in platelet-derived growth factor directed migration by vascular smooth muscle cells. Hypertension 1997, 29, 334–339. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Al-Ani, M.K.; Pan, X.; Chi, Q.; Dong, N.; Qiu, X. Plant-derived products for treatment of vascular intima hyperplasia selectively inhibit vascular smooth muscle cell functions. Evid. Based Complement. Alternat. Med. 2018, 3549312. [Google Scholar] [CrossRef] [Green Version]
- Siasos, G.; Tousoulis, D.; Tsigkou, V.; Kokkou, E.; Oikonomou, E.; Vavuranakis, M.; Basdra, E.K.; Papavassiliou, A.G.; Stefanadis, C. Flavonoids in atherosclerosis: An overview of their mechanisms of action. Curr. Med. Chem. 2013, 20, 2641–2660. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, R.M.; Naidoo, N.; Landberg, R. Dietary flavonoids and the development of type 2 diabetes and cardiovascular diseases: Review of recent findings. Curr. Opin. Lipidol. 2013, 24, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Y.; Zhu, Y.F.; Zhang, M.; Chen, L.; Zhang, Z.; Du, Y.L.; Ren, G.Q.; Tang, J.M.; Zhong, M.K.; Shi, X.J. Chlorogenic acid inhibits hypoxia-induced pulmonary artery smooth muscle cells proliferation via c-Src and Shc/Grb2/ERK2 signaling pathway. Eur. J. Pharmacol. 2015, 751, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.J.; Kim, J.H.; Jin, Y.R.; Yun, Y.P. The inhibitory effect and mechanism of luteolin 7-glucoside on rat aortic vascular smooth muscle cell proliferation. Arch. Pharm. Res. 2006, 29, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Lang, Y.; Chen, D.; Li, D.; Zhu, M.; Xu, T.; Zhang, T.; Qian, W.; Luo, Y. Luteolin inhibited hydrogen peroxide-induced vascular smooth muscle cells proliferation and migration by suppressing the Src and Akt signalling pathways. J. Pharm. Pharmacol. 2012, 64, 597–603. [Google Scholar] [CrossRef]
- Ding, H.; Li, D.; Zhang, Y.; Zhang, T.; Zhu, H.; Xu, T.; Luo, Y.; Wang, C. Luteolin inhibits smooth muscle cell migration and proliferation by attenuating the production of Nox4, p-Akt and VEGF in endothelial cells. Curr. Pharm. Biotechnol. 2014, 14, 1009–1015. [Google Scholar] [CrossRef]
- Xu, T.; Zhu, H.; Li, D.; Lang, Y.; Cao, L.; Liu, Y.; Wu, W.; Chen, D. Luteolin inhibits Angiotensin II-stimulated VSMC proliferation and migration through downregulation of Akt phosphorylation. Evid. Based Complement. Alternat. Med. 2015, 2015, 931782. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Xu, H.T.; Yu, J.J.; Gao, J.L.; Lei, J.; Yin, Q.S.; Li, B.; Pang, M.X.; Su, M.X.; Mi, W.J.; et al. Luteolin ameliorates hypertensive vascular remodeling through inhibiting the proliferation and migration of vascular smooth muscle cells. Evid. Based Complement. Alternat. Med. 2015, 2015, 364876. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.C.; Hsu, H.F.; Huang, K.H.; Wu, J.M.; Kuo, S.M.; Ling, X.H.; Houng, J.Y. Anti-proliferative effects of Siegesbeckia orientalis ethanol extract on human endometrial RL-95 cancer cells. Molecules 2014, 19, 19980–19994. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds are not available from the authors. |









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Concentration (mg/g Extract) | |
|---|---|---|
| GTE | GTW | |
| Chlorogenic acid (1) | 9.03 ± 0.25 | 9.45 ± 0.34 |
| Luteolin-7-glucoside (2) | 46.23 ± 1.42 | 15.40 ± 0.85 |
| Luteolin (3) | 3.71 ± 0.11 | 1.70 ± 0.06 |
| Total polyphenols content | 64.04 ± 1.68 | 26.15 ± 0.96 |
| Total flavonoids content | 52.97 ± 1.14 | 17.49 ± 0.67 |
| Antibody | Company | Identical Number |
|---|---|---|
| p-JNK | Sigma-Aldrich | SAB4504450 |
| p-ERK | Sigma-Aldrich | SAB4301578 |
| p-p38 | Sigma-Aldrich | SAB4301534 |
| p-NF-κB | Sigma-Aldrich | SAB4502609 |
| MMP-2 | Sigma-Aldrich | SAB5700824 |
| MMP-9 | Sigma-Aldrich | SAB5700152 |
| β-Actin | Sigma-Aldrich | SAB3500350 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsuan, C.-F.; Lu, Y.-C.; Tsai, I.-T.; Houng, J.-Y.; Wang, S.-W.; Chang, T.-H.; Chen, Y.-L.; Chang, C.-C. Glossogyne tenuifolia Attenuates Proliferation and Migration of Vascular Smooth Muscle Cells. Molecules 2020, 25, 5832. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25245832
Hsuan C-F, Lu Y-C, Tsai I-T, Houng J-Y, Wang S-W, Chang T-H, Chen Y-L, Chang C-C. Glossogyne tenuifolia Attenuates Proliferation and Migration of Vascular Smooth Muscle Cells. Molecules. 2020; 25(24):5832. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25245832
Chicago/Turabian StyleHsuan, Chin-Feng, Yung-Chuan Lu, I-Ting Tsai, Jer-Yiing Houng, Shih-Wei Wang, Tzu-Hsien Chang, Ya-Ling Chen, and Chi-Chang Chang. 2020. "Glossogyne tenuifolia Attenuates Proliferation and Migration of Vascular Smooth Muscle Cells" Molecules 25, no. 24: 5832. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25245832