
AEDG Peptide (Epitalon) Stimulates Gene Expression and Protein Synthesis during Neurogenesis: Possible Epigenetic Mechanism

 , , , ,
, , , , 
Abstract
:1. Introduction
2. Results
2.1. Cell Characterization
2.2. Immunofluorescence Analysis
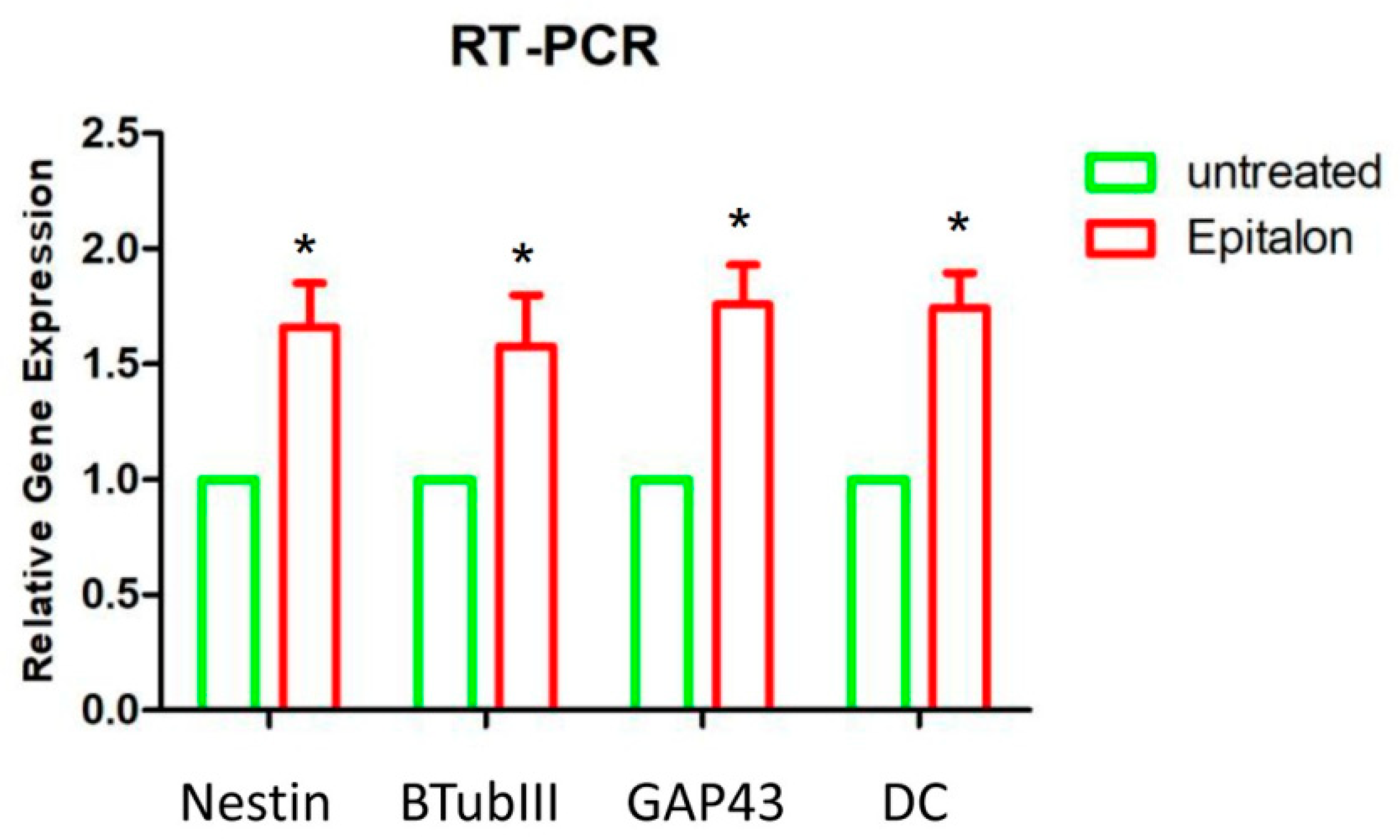
2.3. Gene Expression
2.4. Histone-Peptide Interaction Analysis
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Experimental Design
4.3. Immunofluorescence Analysis
4.4. Real Time PCR
4.5. Statistical Analysis of the Experimental Data
4.6. Molecular Modelling of AEDG Peptide—Histone Interactions
Author Contributions
Funding
Conflicts of Interest
References
- Khavinson, V.K. Peptides and ageing. Neuroendocr. Lett. 2002, 23, 11–144. [Google Scholar]
- Khavinson, V.K.; Linkova, N.S.; Tarnovskaya, S.I. Short Peptides Regulate Gene Expression. Bull. Exp. Biol. Med. 2016, 162, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Anisimov, V.N.; Khavinson, V.K. Peptide bioregulation of aging: Results and prospects. Biogerontology 2010, 11, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Khavinson, V.K.; Bondarev, I.E.; Butyugov, A.A. Epithalon Peptide Induces Telomerase Activity and Telomere Elongation in Human Somatic Cells. Bull. Exp. Biol. Med. 2003, 135, 590–592. [Google Scholar] [CrossRef]
- Ashapkin, V.V.; Linkova, N.S.; Khavinson, V.K.; Vanyushin, B.F. Epigenetic Mechanisms of Peptidergic Regulation of Gene Expression during aging of Human Cells. Biochemistry 2015, 80, 374–388. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Kuznik, B.I.; Ryzhak, G.A. Peptide bioregulators: The new class of geroprotectors. Communication 1. Results of experimental studies. Adv. Gerontol. 2012, 25, 696–708. [Google Scholar]
- Khavinson, V.K.; Kuznik, B.I.; Ryzhak, G.A. Peptide bioregulators: The new class of geroprotectors. Message 2. Clinical studies results. Adv. Gerontol. 2013, 26, 20–37. [Google Scholar]
- Sokolova, I.B.; Sergeev, I.V.; Anisimov, S.V.; Puzanov, M.V.; Dvoretsky, D.P. Effect of transplantation of mesenchymal stem cells on the density of pial microvascular network in rats of different age. Bull. Exp. Biol. Med. 2013, 154, 548–552. [Google Scholar] [CrossRef]
- Sokolova, I.B.; Sergeev, I.V.; Bilibina, A.A.; Dvoretskiy, D.P. Arteriogenesis in the pia matter of the rat brain cortex after intracerebral injection of mesenchymal stem cells. Bull. Exp. Biol. Med. 2012, 154, 177–179. [Google Scholar] [CrossRef]
- Kim, S.U.; Lee, H.J.; Kim, Y.B. Neural stem cell-based treatment for neurodegenerative diseases. Neuropathology 2013, 33, 491–504. [Google Scholar] [CrossRef]
- Kang, J.M.; Yeon, B.K.; Cho, S.J.; Suh, Y.H. Stem Cell Therapy for Alzheimer’s Disease: A Review of Recent Clinical Trials. J. Alzheimers Dis 2016, 54, 879–889. [Google Scholar] [CrossRef]
- Pizzicannella, J.; Diomede, F.; Merciaro, I.; Caputi, S.; Tartaro, A.; Guarnieri, S.; Trubiani, O. Endothelial committed oral stem cells as modelling in the relationship between periodontal and cardiovascular disease. J. Cell. Physiol. 2018, 233, 6734–6747. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, M.F.; Maria, D.A.; de Isla, N.; Pinto Leal-Junior, E.C.P.; Joensen, J.; Bjordal, J.M.; Lopes-Martins, R.; Diomede, F.; Trubiani, O.; Frigo, L. Evaluation of the Proliferative Effects Induced by Low-Level Laser Therapy in Bone Marrow Stem Cell Culture. Photomed. Laser Surg. 2015, 33, 610–616. [Google Scholar] [CrossRef]
- Diomede, F.; Gugliandolo, A.; Scionti, D.; Merciaro, I.; Cavalcanti, M.F.; Mazzon, E.; Trubiani, O. Biotherapeutic Effect of Gingival Stem Cells Conditioned Medium in Bone Tissue Restoration. Int. J. Mol. Sci. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gugliandolo, A.; Diomede, F.; Cardelli, P.; Bramanti, A.; Scionti, D.; Bramanti, P.; Trubiani, O.; Mazzon, E. Transcriptomic analysis of gingival mesenchymal stem cells cultured on 3D bioprinted scaffold: A promising strategy for neuroregeneration. J. Biomed. Mater. Res. Part. A 2018, 106, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Gugliandolo, A.; Cardelli, P.; Merciaro, I.; Ettorre, V.; Traini, T.; Bedini, R.; Scionti, D.; Bramanti, A.; Nanci, A.; et al. Three-dimensional printed PLA scaffold and human gingival stem cell-derived extracellular vesicles: A new tool for bone defect repair. Stem Cell Res. Ther. 2018, 9, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diomede, F.; Zini, N.; Pizzicannella, J.; Merciaro, I.; Pizzicannella, G.; D’Orazio, M.; Piattelli, A.; Trubiani, O. 5-Aza Exposure Improves Reprogramming Process through Embryoid Body Formation in Human Gingival Stem Cells. Front. Genet. 2018. [Google Scholar] [CrossRef] [Green Version]
- Trubiani, O.; Toniato, E.; Di Iorio, D.; Diomede, F.; Merciaro, I.; Arcangelo, C.D.; Caputi, S. Morphological analysis and interleukin release in human gingival fibroblasts seeded on different denture base acrylic resins. Int. J. Immunopathol. Pharmacol. 2012, 25, 637–643. [Google Scholar] [CrossRef]
- Libro, R.; Scionti, D.; Diomede, F.; Marchisio, M.; Grassi, G.; Pollastro, F.; Piattelli, A.; Bramanti, P.; Mazzon, E.; Trubiani, O. Cannabidiol Modulates the Immunophenotype and Inhibits the Activation of the Inflammasome in Human Gingival Mesenchymal Stem Cells. Front. Physiol. 2016, 7, 559. [Google Scholar] [CrossRef] [Green Version]
- Rajan, T.S.; Scionti, D.; Diomede, F.; Grassi, G.; Pollastro, F.; Piattelli, A.; Cocco, L.; Bramanti, P.; Mazzon, E.; Trubiani, O. Gingival stromal cells as an in vitro model: Cannabidiol modulates genes linked with amyotrophic lateral sclerosis. J. Cell. Biochem. 2017, 118, 819–828. [Google Scholar] [CrossRef]
- Trubiani, O.; Guarnieri, S.; Diomede, F.; Mariggio, M.A.; Merciaro, I.; Morabito, C.; Cavalcanti, M.F.X.B.; Cocco, L.; Ramazzotti, G. Nuclear translocation of PKCalpha isoenzyme is involved in neurogenic commitment of human neural crest-derived periodontal ligament stem cells. Cell. Signal. 2016, 28, 1631–1641. [Google Scholar] [CrossRef]
- Fedoreyeva, L.I.; Kireev, I.I.; Khavinson, V.K.; Vanyushin, B.F. Penetration of Short Fluorescence-Labeled Peptides into the Nucleus in HeLa Cells and in vitro Specific Interaction of the Peptides with Deoxyribooligonucleotides and DNA. Biochemistry 2011, 76, 1210–1219. [Google Scholar] [CrossRef] [PubMed]
- Kuznik, B.I.; Davydov, S.O.; Popravka, E.S.; Linkova, N.S.; Kozina, L.S.; Khavinson, V.K. Epigenetic Mechanism of Peptide-Driven Regulation and Neuroprotective Protein FKBP1b. Mol. Biol. 2019, 53, 299–307. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Tendler, S.M.; Kasyanenko, N.A.; Tarnovskaya, S.I.; Linkova, N.S.; Ashapkin, V.V.; Yakutseni, P.P.; Vanyushin, B.F. Tetrapeptide KEDW Interacts with DNA and Regulates Gene Expression. Am. J. Biomed. Sci. 2015, 7, 156–169. [Google Scholar] [CrossRef]
- Kolchina, N.; Khavinson, V.; Linkova, N.; Yakimov, A.; Baitin, D.; Afanasyeva, A.; Petukhov, M. Systematic search for structural motifs of peptide binding to double-stranded DNA. Nucleic Acids Res. 2019, 47, 10553–10563. [Google Scholar] [CrossRef] [PubMed]
- Fedoreyeva, L.I.; Smirnova, T.A.; Kolomijtseva, G.Y.; Khavinson, V.K.; Vanyushin, B.F. Interaction of Short Peptides with FITC-Labeled Wheat Histones and Their Complexes with Deoxyribooligonucleotides. Biochemistry 2013, 78, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Caputi, S.; Trubiani, O.; Sinjari, B.; Trofimova, S.; Diomede, F.; Linkova, N.; Diatlova, A.; Khavinson, V. Effect of short peptides on neuronal differentiation of stem cells. Int. J. Immunopathol. Pharmacol. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khavinson, V.K.; Bondarev, I.E.; Butyugov, A.A.; Smirnova, T.D. Peptide Promotes Overcoming of the Division Limit in Human Somatic Cell. Bull. Exp. Biol. Med. 2004, 137, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Khavinson, V.K.; Linkova, N.S.; Kvetnoy, I.M.; Kvetnaia, T.V.; Polyakova, V.O.; Korf, H.-W. Molecular Cellular Mechanisms of Peptide Regulation of Melatonin Synthesis in Pinealocyte Culture. Bull. Exp. Biol. Med. 2012, 153, 255–258. [Google Scholar] [CrossRef]
- Djeridane, Y.; Khavinson, V.K.; Anisimov, V.N.; Touitou, Y. Effect of synthetic pineal tetrapeptide (Ala-Glu-Asp-Gly) on melatonin secretion by the pineal gland of young and old rats. J. Endocrinol. Investig. 2003, 26, 211–215. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Pronyaeva, V.E.; Linkova, N.S.; Trofimova, S.V.; Umnov, R.S. Molecular-Physiological Aspects of Peptide Regulation of the Function of the Retina in Retinitis Pigmentosa. Hum. Physiol. 2014, 40, 153–158. [Google Scholar] [CrossRef]
- Khavinson, V.K.; Pronyaeva, V.E.; Linkova, N.S.; Trofimova, S.V. Peptidergic Regulation of Differentiation of Embryonic Retinal Cells. Bull. Exp. Biol. Med. 2013, 155, 172–174. [Google Scholar] [CrossRef] [PubMed]
- Zamorskii, I.I.; Shchudrova, T.S.; Linkova, N.S.; Nichik, T.E.; Khavinson, V.K. Peptides Restore Functional State of the Kidneys during Cisplatin-Induced Acute Renal Failure. Bull. Exp. Biol. Med. 2015, 159, 736–739. [Google Scholar] [CrossRef] [PubMed]
- Manescu, A.; Giuliani, A.; Mohammadi, S.; Tromba, G.; Mazzoni, S.; Diomede, F.; Zini, N.; Piattelli, A.; Trubiani, O. Osteogenic potential of dualblocks cultured with human periodontal ligament stem cells: In vitro and synchrotron microtomography study. J. Periodontal Res. 2016, 51, 112–124. [Google Scholar] [CrossRef]
- Diomede, F.; Zini, N.; Gatta, V.; Fulle, S.; Merciaro, I.; D’Aurora, M.; La Rovere, R.M.; Traini, T.; Pizzicannella, J.; Ballerini, P.; et al. Human periodontal ligament stem cells cultured onto cortico-cancellous scaffold drive bone regenerative process. Eur. Cells Mater. 2016, 32, 181–201. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Merciaro, I.; Martinotti, S.; Cavalcanti, M.F.; Caputi, S.; Mazzon, E.; Trubiani, O. miR-2861 is involved in osteogenic commitment of human periodontal ligament stem cells grown onto 3D scaffold. J. Biol. Regul. Homeost. Agents 2016, 30, 1009–1018. [Google Scholar] [PubMed]
- Mammana, S.; Gugliandolo, A.; Cavalli, E.; Diomede, F.; Iori, R.; Zappacosta, R.; Bramanti, P.; Conti, P.; Fontana, A.; Pizzicannella, J.; et al. Human gingival mesenchymal stem cells pretreated with vesicular moringin nanostructures as a new therapeutic approach in a mouse model of spinal cord injury. J. Tissue Eng. Regen. Med. 2019. [Google Scholar] [CrossRef] [Green Version]
- Furuhashi, M.; Hotamisligil, G.S. Fatty acid-binding proteins: Role in metabolic diseases and potential as drug targets. Nat. Rev. Drug Discov. 2008, 7, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Furuhashi, M.; Ishimura, S.; Ota, H.; Miura, T. Lipid chaperones and metabolic inflammation. Int. J. Inflamm. 2011. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.X. PPARs: Diverse regulators in energy metabolism and metabolic diseases. Cell Res. 2010, 20, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Pizzicannella, J.; Gugliandolo, A.; Orsini, T.; Fontana, A.; Ventrella, A.; Mazzon, E.; Bramanti, P.; Diomede, F.; Trubiani, O. Engineered Extracellular Vesicles from Human Periodontal-Ligament Stem Cells Increase VEGF/VEGFR2 Expression during Bone Regeneration. Front. Physiol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Diomede, F.; Rajan, T.S.; Gatta, V.; D’Aurora, M.; Merciaro, I.; Marchisio, M.; Muttini, A.; Caputi, S.; Bramanti, P.; Mazzon, E.; et al. Stemness Maintenance Properties in Human Oral Stem Cells after Long-Term Passage. Stem Cells Int. 2017. [Google Scholar] [CrossRef]
- Pizzicannella, J.; Cavalcanti, M.; Trubiani, O.; Diomede, F. MicroRNA 210 Mediates VEGF Upregulation in Human Periodontal Ligament Stem Cells Cultured on 3D Hydroxyapatite Ceramic Scaffold. Int. J. Mol. Sci. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalczyk, K.; Ziman, M. Nestin structure and predicted function in cellular cytoskeletal organisation. Histol. Histopathol. 2005, 20, 665–671. [Google Scholar] [PubMed]
- Yan, S.X.; Li, P.L.; Wang, Y.; Yu, W.; Qin, A.; Liu, M.; Xiang, A.P.; Zhang, W.; Li, W. Nestin regulates neural stem cell migration via controlling the cell contractility. Int. J. Biochem. Cell B 2016, 78, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Quick, Q.; Paul, M.; Skalli, O. Roles and Potential Clinical Applications of Intermediate Filament Proteins in Brain Tumors. Semin. Pediatr. Neurol. 2015, 22, 40–48. [Google Scholar] [CrossRef]
- Zhao, J.C.; Zhang, L.X.; Zhang, Y.; Shen, Y.F. The differential regulation of Gap43 gene in the neuronal differentiation of P19 cells. J. Cell. Physiol. 2012, 227, 2645–2653. [Google Scholar] [CrossRef]
- Gagliardini, V.; Dusart, I.; Fankhauser, C. Absence of GAP43 can protect neurons from death. Mol. Cell. Neurosci. 2000, 16, 27–33. [Google Scholar] [CrossRef]
- Tong, J.; Nguyen, L.; Vidal, A.; Simon, S.A.; Skene, J.H.P.; McIntosh, T.J. Role of GAP43 in sequestering phosphatidylinositol 4,5-bisphosphate to raft bilayers. Biophys. J. 2008, 94, 125–133. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.; Gupta, S.K.; Meiri, K.F.; Fong, M.; Thostrup, P.; Juncker, D.; Mani, S. GAP-43 is key to mitotic spindle control and centrosome-based polarization in neurons. Cell Cycle 2008, 7, 348–357. [Google Scholar] [CrossRef] [Green Version]
- Kamande, J.W.; Nagendran, T.; Harris, J.; Taylor, A.M. Multi-compartment Microfluidic Device Geometry and Covalently Bound Poly-D-Lysine Influence Neuronal Maturation. Front. Bioeng. Biotechnol. 2019, 7, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niwa, S.H.; Takahashi, H.; Hirokawa, N. β-Tubulin mutations that cause severe neuropathies disrupt axonal transport. EMBO J. 2013, 32, 1352–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burger, D.; Stihle, M.; Sharma, A.; Di Lello, P.; Benz, J.; D’Arcy, B.; Debulpaep, M.; Fry, D.; Huber, W.; Kremer, T.; et al. Crystal Structures of the Human Doublecortin C- and N-terminal Domains in Complex with Specific Antibodies. J. Biol. Chem 2016, 291, 16292–16306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labute, P. LowModeMD: Implicit low-mode velocity filtering applied to conformational search of macrocycles and protein loops. J. Chem. Inf. Model. 2010, 50, 792–800. [Google Scholar] [CrossRef] [PubMed]
- Case, D.A.; Darden, T.A.; Cheatham, T.E. AMBER 12; University of California Press: San Francisco, CA, USA, 2012. [Google Scholar]
- Edelsbrunner, H.; Facello, M.; Fu, R.; Liang, J. Measuring proteins and voids in proteins. In Proceedings of the Twenty-Eighth Annual Hawaii International Conference on System Sciences, Wailea, HI, USA, 3–6 January 1995; pp. 256–264. [Google Scholar]
- Soga, S.; Shirai, H.; Kobori, M.; Hirayama, N. Use of amino acid composition to predict ligand-binding sites. J. Chem. Inf. Model. 2007, 47, 400–406. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | hGMSCs | |
|---|---|---|
| Phenotype | MFI Ratio ± SD | |
| CD13 | +++ | 173.4 ± 21.7 |
| CD29 | +++ | 195.2 ± 22.3 |
| CD44 | +++ | 167.8 ± 27.3 |
| CD73 | ++ | 29.4 ± 2.9 |
| CD90 | +++ | 396.4 ± 28.2 |
| CD105 | + | 9.1 ± 1.3 |
| CD14 | − | ND |
| CD34 | − | ND |
| CD45 | − | ND |
| Histone of Wheat Triticum aestivum, FASTA | PDB Structure and Its Amino Acid Sequence, Used to Construct Homologous Histone Models, FASTA | Identity of Original Sequence and Template, % |
|---|---|---|
| >H1.1 MSTDVVADVPAPEVAAAADPVVETTAEPAAGDANAAKETKAKAAKAKKPSAPRKPRAAPAHPTYAEMVSEAITALKERTGSSPYAIAKFVEDKHKAHLPANFRKILSVQLKKLVASGKLTKVKASYKLSAAAAKPKPAAKKKPAAKKKAPAKKTATKTKAKAPAKKSAAKPKAKAPAKTKAAAKPKAAAKPKAKAPAKTKAAAKPKAAAKPKGPPAKAAKTSAKDAPGKNAGAAAPKKPAARKPPTKRSTPVKKAAPAKKAAPAKKAPAAKKAKK | >5NL0_Z HPKYSDMILAAVQAEKSRSGSSRQSIQKYIKNHYKVGENADSQIKLSIKRLVTSGALKQTKGVGASGSFRLAK | 34 |
| >H1.3 MSTEVAAADIPVPQVEVAADAAVDTPAANAKAPKAAKAKKSTGPKKPRVTPAHPSYAEMVSEAIAALKERSGSSTIAIGKFIEDKHKAHLPANFRKILLTQIKKLVAAGKLTKVKGSYKLAKAPAAVKPKTATKKKPAAKPKAKAPAKKTAAKSPAKKAAAKPKAKAPAKAKAVAKPKAAAKPKAAAKPKAKAAAKKAPAAATPKKPAARKPPTKRATPVKKAAPAKKPAAKKAKK | >5NL0_Z HPKYSDMILAAVQAEKSRSGSSRQSIQKYIKNHYKVGENADSQIKLSIKRLVTSGALKQTKGVGASGSFRLAK | 36 |
| >H1.6 PVPQVEVAADAAVDTPAASAKAPKAAKAKKSTGPKKPRVTPAHPSYAEMVSEAIAALKERSGSSTIAIAKFIEDKHKAHLPANFRKILLTQIKKLVAAGKLTKVKGSYKLAKAPAAVKPKTATKKKPAAKPKAKAPAKKTAAKSPAKKAAAKPKAKAPAKAKAVAKPKAASKPKAAAKPKAKAAAKKAPAAATPKKPAAARKPPTKRATPVKKAAPAKKPAAKKAKK | >5NL0_Z HPKYSDMILAAVQAEKSRSGSSRQSIQKYIKNHYKVGENADSQIKLSIKRLVTSGALKQTKGVGASGSFRLAK | 36 |
| >H2b MAPKAAKKPAAKKPAEEEPAAEKAEKTPAGKKPKAERRIPAGKSAAKAGGDKKGKKKAKKSVETYKKYIFKVIKQVHPDIGISSKAMSIMNSFINDIFEKLAGEAAKIARYNKKPYITSREIQTSVRLVLPGELAKHAVSEGTKAVTKFTSAS | >1KX5_H PEPAKSAPAPKKGSKKAVTKTQKKDGKKRRKTRKESYAIYVYKVLKQVHPDTGISSKAMSIMNSFVNDVFERIAGEASRLAHYNKRSTITSREIQTAVRLLLPGELAKHAVSEGTKAVTKYTSAK | 65 |
| >H3 ARTKQTARKSTGGKAPRKQLATKAARKSAPATGGVKKPHRERPGTVALREIRKYQKSTELLIRKLPFQRLVREIAQDFKTDLRFQSSAVAALQEAAEAYLVGLFEDTNLCAIHAKRVTIMPKDIQLARRIRGERA | >1KX5_A ARTKQTARKSTGGKAPRKQLATKAARKSAPATGGVKKPHRYRPGTVALREIRRYQKSTELLIRKLPFQRLVREIAQDFKTDLRFQSSAVMALQEASEAYLVALFEDTNLCAIHAKRVTIMPKDIQLARRIRGERA | 96 |
| >H4 SGRGKGGKGLGKGGAKRHRKVLRDNIQGITKPAIRRLARRGGVKRISGLIYEETRGVLKIFLENVIRDAVTYTEHARRKTVTANDVVYALKRQGRTLYGFGG | >1EQZ_D GAKRHRKVLRDNIQGITKPAIRRLARRGGVKRISGLIYEETRGVLKVFLENVIRDAVTYTEHAKRKTVTAMDVVYALKRQGRTLYGFGG | 97 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khavinson, V.; Diomede, F.; Mironova, E.; Linkova, N.; Trofimova, S.; Trubiani, O.; Caputi, S.; Sinjari, B. AEDG Peptide (Epitalon) Stimulates Gene Expression and Protein Synthesis during Neurogenesis: Possible Epigenetic Mechanism. Molecules 2020, 25, 609. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25030609
Khavinson V, Diomede F, Mironova E, Linkova N, Trofimova S, Trubiani O, Caputi S, Sinjari B. AEDG Peptide (Epitalon) Stimulates Gene Expression and Protein Synthesis during Neurogenesis: Possible Epigenetic Mechanism. Molecules. 2020; 25(3):609. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25030609
Chicago/Turabian StyleKhavinson, Vladimir, Francesca Diomede, Ekaterina Mironova, Natalia Linkova, Svetlana Trofimova, Oriana Trubiani, Sergio Caputi, and Bruna Sinjari. 2020. "AEDG Peptide (Epitalon) Stimulates Gene Expression and Protein Synthesis during Neurogenesis: Possible Epigenetic Mechanism" Molecules 25, no. 3: 609. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25030609