
Induction of Apoptosis by Gluconasturtiin-Isothiocyanate (GNST-ITC) in Human Hepatocarcinoma HepG2 Cells and Human Breast Adenocarcinoma MCF-7 Cells

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. GNST-ITC-Mediated Cytotoxicity
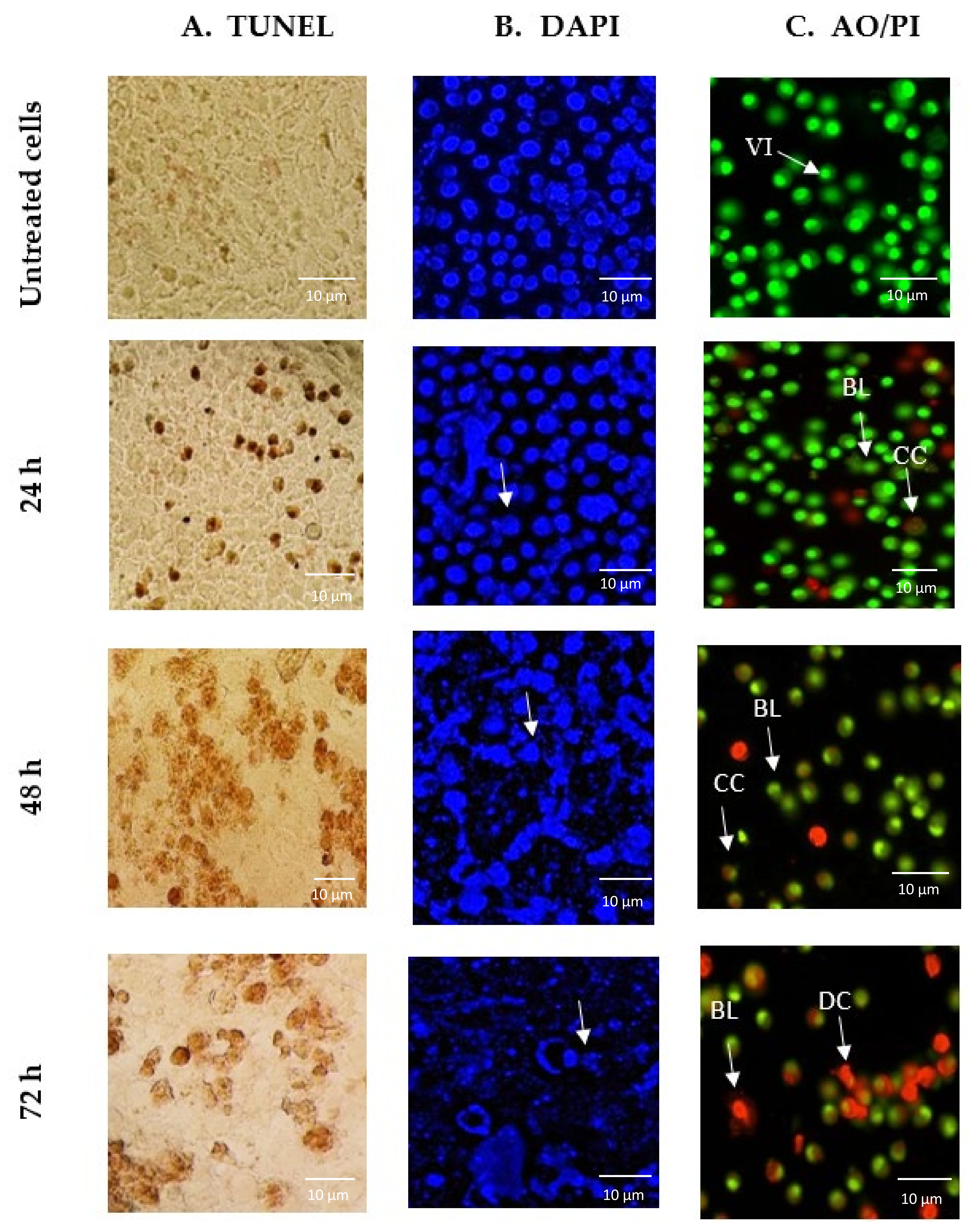
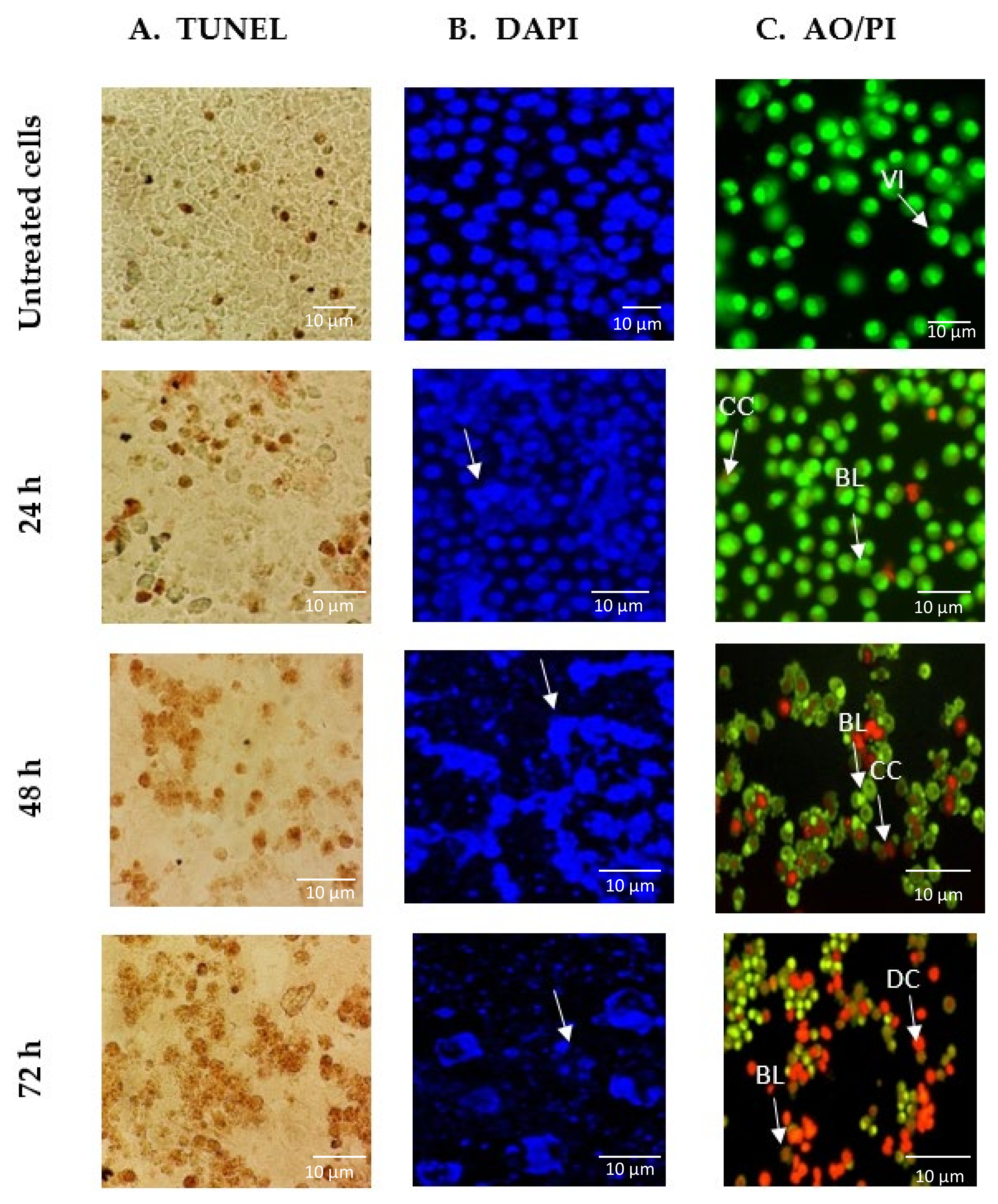
2.2. GNST-ITC-Mediated Morphological Changes in HepG2 and MCF-7 Cells
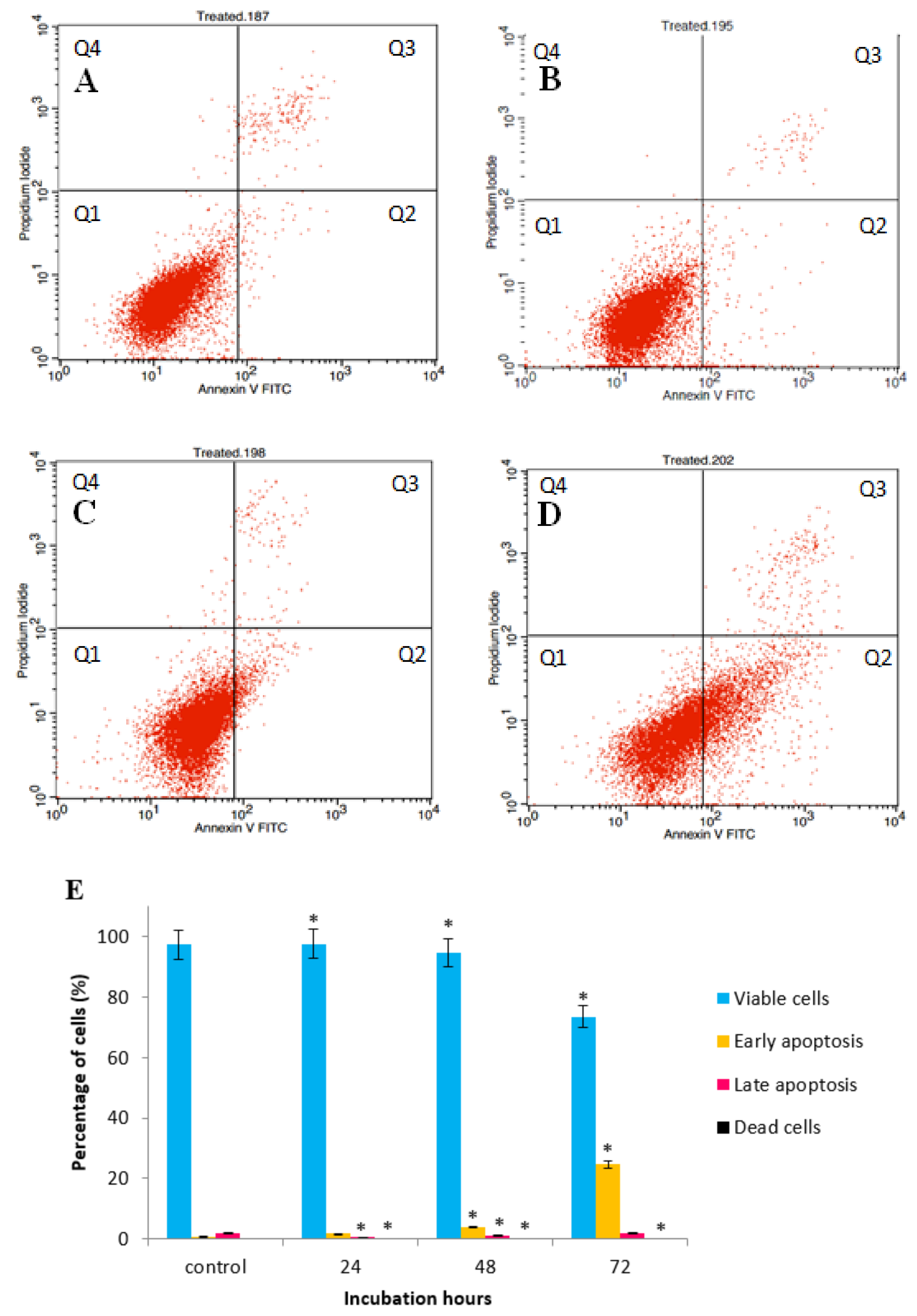
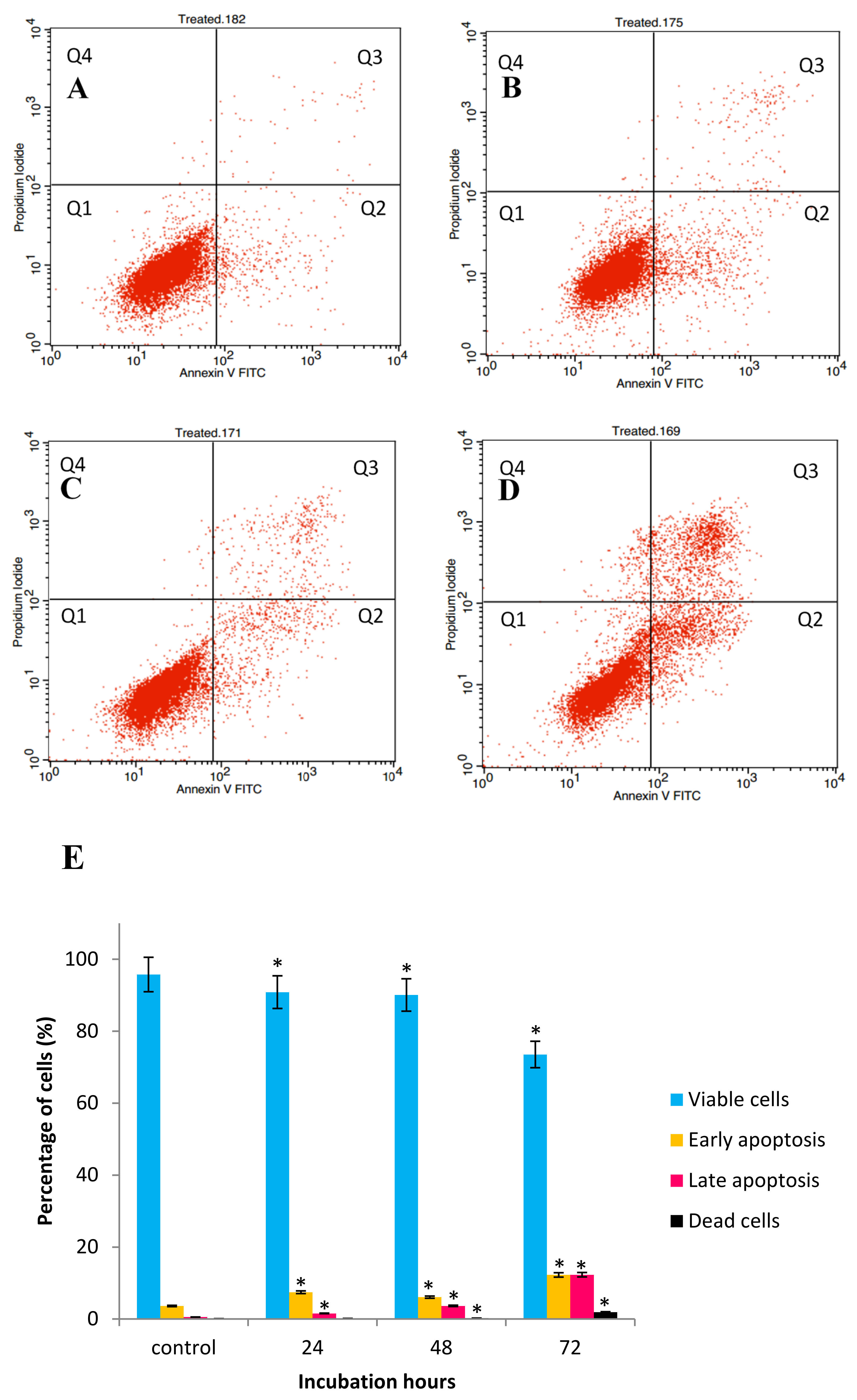
2.3. GNST-ITC-Mediated Induction of Early Apoptosis in HepG2 and MCF-7 Cell Lines
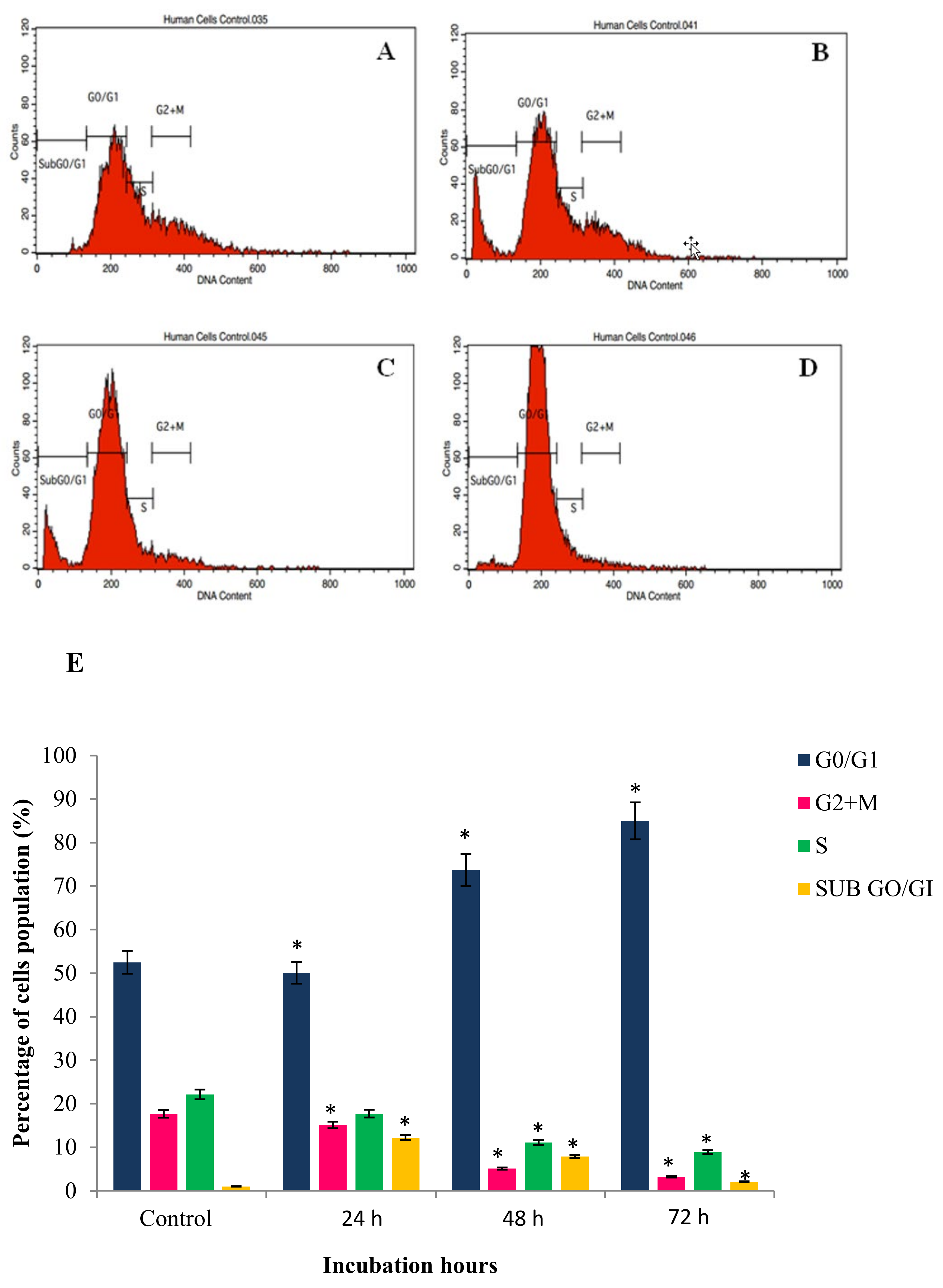
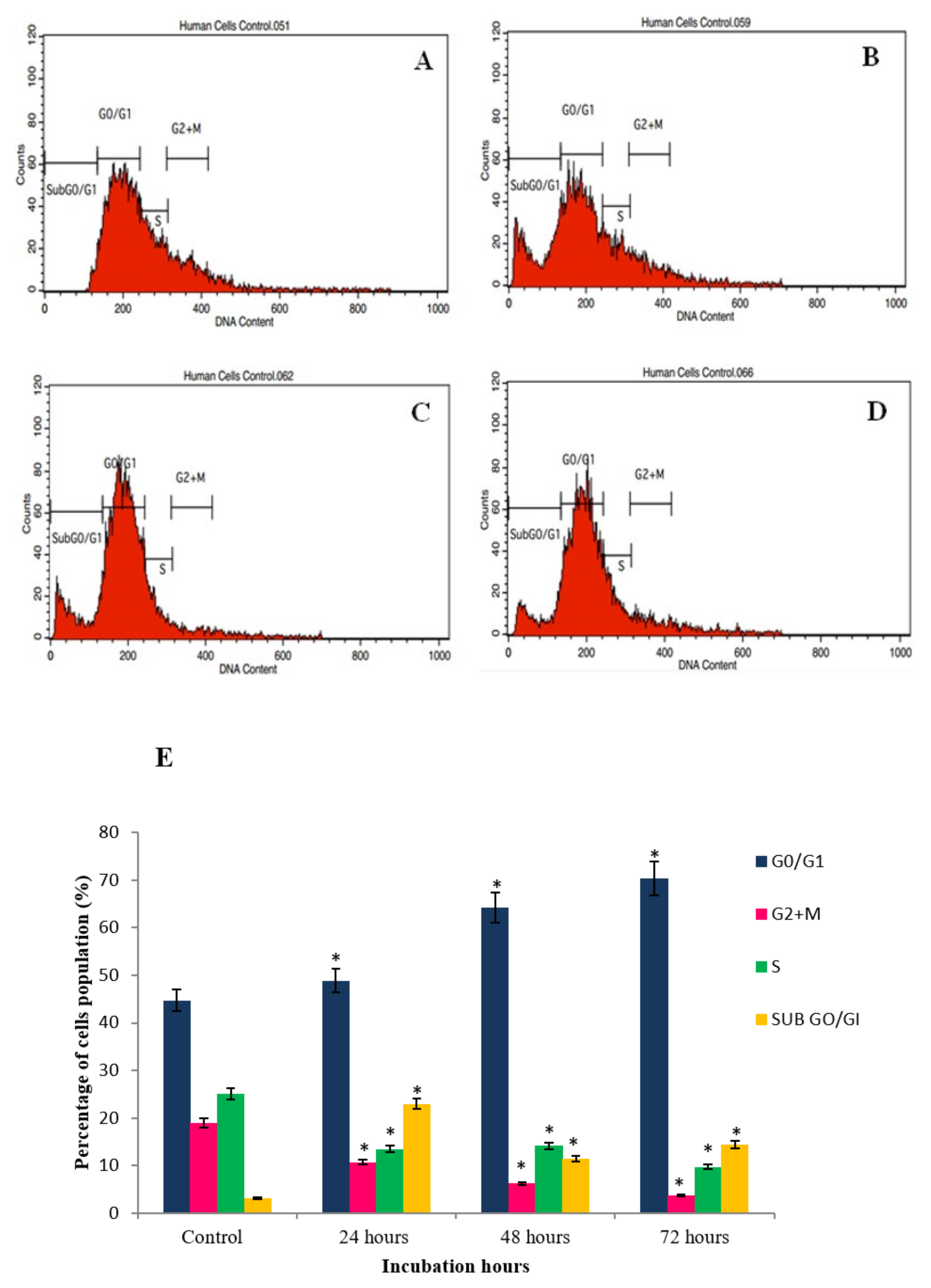
2.4. GNST-ITC-Mediated Cell Cycle Arrest
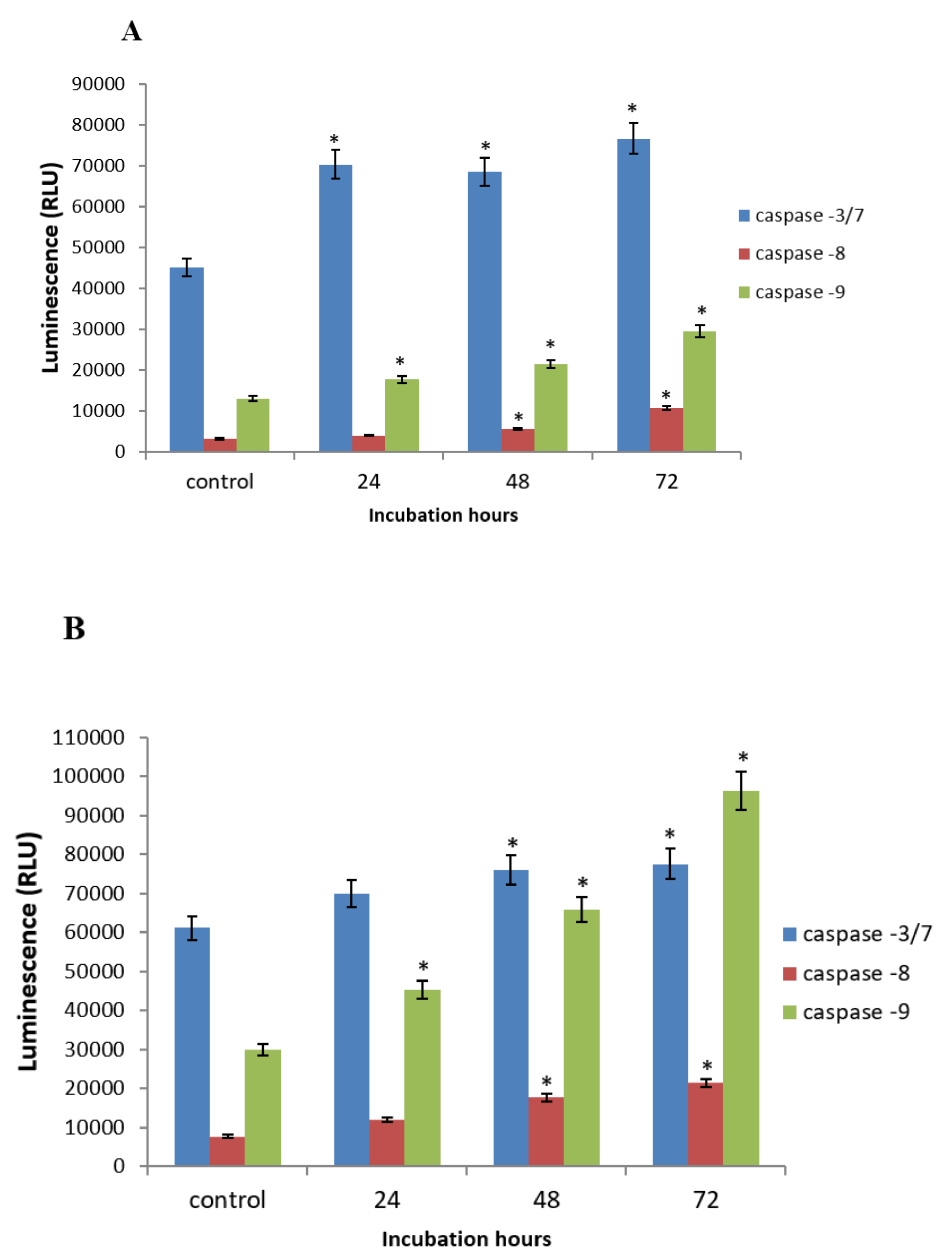
2.5. GNST-ITC-Mediated Modulation of Caspase-3/7, -8, and -9 Activities
3. Discussion
4. Materials and Methods
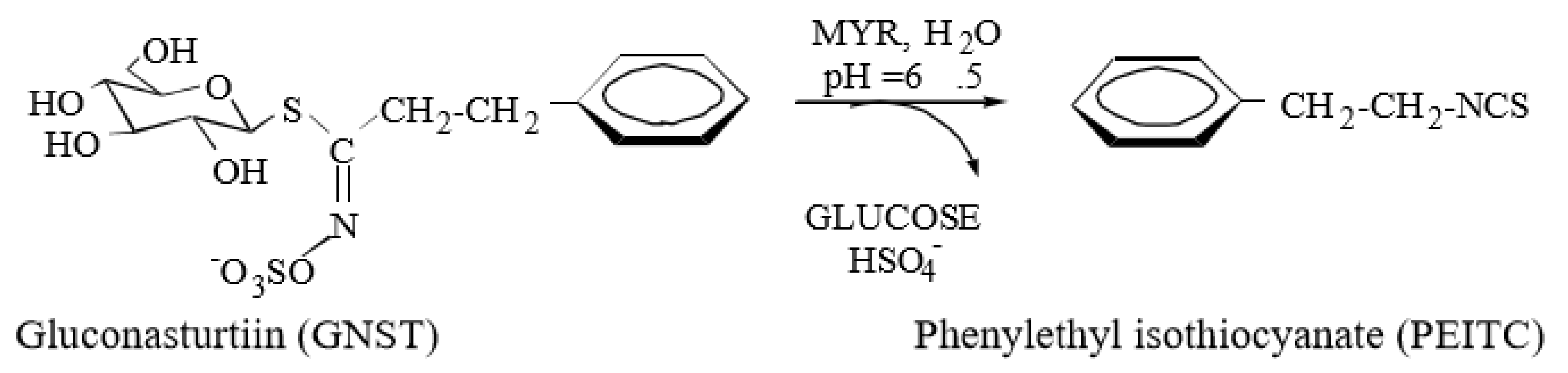
4.1. Isolation and Characterisation of Gluconasturtiin (GNST)
4.2. Cell Culture
4.3. Assessment of Cytotoxicity Using the MTT Assay
4.4. Morphology Assessment of Apoptotic Cells by TUNEL Assay
4.5. Morphology Assessment of Apoptotic Cells by DAPI Staining
4.6. Morphology Assessment of Apoptotic Cells by AO/PI Staining
4.7. Annexin V-FITC/PI by Flow Cytometry
4.8. Cell Cycle Analysis by Flow Cytometry
4.9. Determination of Caspase-3/7, -8, and -9 Activities
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vassiliki, B.; Areti, L.; Pagona, L. Chemoprevention of cancer: Current evidence and future prospects. F1000Research 2015, 4, 1–10. [Google Scholar]
- Aghajanpour, M.; Nazer, M.R.; Obeidavi, Z.; Akbari, M.; Ezati, P.; Kor, N.M. Functional foods and their role in cancer prevention and health promotion: A comprehensive review. Am. J. Cancer Res. 2017, 7, 740. [Google Scholar]
- Arends, J.; Bachmann, P.; Baracos, V.; Barthelemy, N.; Bertz, H.; Bozzetti, F.; Krznaric, Z. ESPEN guidelines on nutrition in cancer patients. Clin. Nutr. 2017, 36, 11–48. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.; Kntayya, S.; Mohd Ain, N.; Iori, R.; Ioannides, C.; Abdull Razis, A. Induction of Apoptosis and Cytotoxicity by raphasatin in human breast adenocarcinoma MCF-7 cells. Molecules 2018, 23, 3092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kntayya, S.; Ibrahim, M.; Mohd Ain, N.; Iori, R.; Ioannides, C.; Abdull Razis, A. Induction of apoptosis and cytotoxicity by isothiocyanate sulforaphene in human hepatocarcinoma HepG2 cells. Nutrients 2018, 10, 718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arumugam, A.; Razis, A.F.A. Apoptosis as a mechanism of the cancer chemopreventive activity of glucosinolates: A review. Asian Pac. J. Cancer Prev. 2018, 19, 1439. [Google Scholar] [PubMed]
- Fouad, Y.A.; Aanei, C. Revisiting the hallmarks of cancer. Am. J. Cancer Res. 2017, 7, 1016. [Google Scholar]
- Razis, A.F.A.; Noor, N.M. Cruciferous vegetables: Dietary phytochemicals for cancer prevention. Asian Pac. J. Cancer Prev. 2013, 14, 1565–1570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barba, F.J.; Nikmaram, N.; Roohinejad, S.; Khelfa, A.; Zhu, Z.; Koubaa, M. Bioavailability of glucosinolates and their breakdown products: Impact of processing. Front. Nutr. 2016, 3, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdull Razis, A.F.; Konsue, N.; Ioannides, C. Isothiocyanates and xenobiotic detoxification. Mol. Nutr. Food Res. 2018, 62, 1700916. [Google Scholar] [CrossRef]
- Razis, A.; Faizal, A.; Mohd Noor, N.; Konsue, N. Induction of epoxide hydrolase, glucuronosyl transferase, and sulfotransferase by phenethyl isothiocyanate in male Wistar albino rats. BioMed Res. Int. 2014. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, J.; Tang, L. Cancer-preventive isothiocyanates: Dichotomous modulators of oxidative stress. Radic. Biol. Med. 2005, 38, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Lee, H.J. Effects of phenylethyl isothiocyanate and its metabolite on cell-cycle arrest and apoptosis in LNCaP human prostate cancer cells. Intl. J. Food Sci. Nutr. 2010, 61, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Mitsiogianni, M.; Koutsidis, G.; Mavroudis, N.; Trafalis, D.T.; Botaitis, S.; Franco, R.; Panayiotidis, M.I. The Role of Isothiocyanates as Cancer Chemo-Preventive, Chemo-Therapeutic and Anti-Melanoma Agents. Antioxidants 2019, 8, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, H.Y.; Gong, Y.; Ye, F.G.; Ling, H.; Hu, X. Incidence and prognostic factors of patients with synchronous liver metastases upon initial diagnosis of breast cancer: A population-based study. Cancer Manag Res. 2018, 10, 5937. [Google Scholar] [CrossRef] [Green Version]
- McClay, E.F.; McClay, M.E.T.; Monroe, L.; Baron, P.L.; Cole, D.J.; O’Brien, P.H.; Maize, J.C. The effect of tamoxifen and cisplatin on the disease-free and overall survival of patients with high risk malignant melanoma. Brit. J. Cancer. 2000, 83, 16. [Google Scholar] [CrossRef] [Green Version]
- Konsue, N.; Abdul Razis, A.F. Cytotoxicity of gluconasturtiin and its derivative against MCF-7 and HepG2. KKU Sci. J. 2016, 44, 272–281. [Google Scholar]
- Tseng, E.; Scott-Ramsay, E.A.; Morris, M.E. Dietary organic isothiocyanates are cytotoxic in human breast cancer MCF-7 and mammary epithelial MCF-12A cell lines. Exp. Biol. Med. 2004, 229, 835–842. [Google Scholar] [CrossRef]
- Liu, K.; Cang, S.; Ma, Y.; Chiao, J.W. Synergistic effect of paclitaxel and epigenetic agent phenethyl isothiocyanate on growth inhibition, cell cycle arrest and apoptosis in breast cancer cells. Cancer Cell Int. 2013, 13, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hanlon, N.; Coldham, N.; Gielbert, A.; Kuhnert, N.; Sauer, M.J.; King, L.J.; Ioannides, C. Absolute bioavailability and dose-dependent pharmacokinetic behaviour of dietary doses of the chemopreventive isothiocyanate sulforaphane in the rat. Br. J. Nutr. 2008, 99, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Konsue, N.; Kirkpatrick, J.; Kuhnert, N.; King, L.J.; Ioannides, C. Repeated oral administration modulates the pharmacokinetic behaviour of the chemopreventive agent phenethyl isothiocyanate in rats. Mol. Nutr. Food Res. 2010, 54, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Sankari, L.; Masthan, K.M.K.; Aravindha Babu, N.; Bhattacharjee, T.; Elumalai, M. Apoptosis in Cancer – An Update. Asian Pac. J. Cancer Prev. 2012, 13, 4873–4878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ulukaya, E.; Acilan, C.; Yilmaz, Y. Apoptosis: Why and how does it occur in biology? Cell Biochem. Funct. 2011, 29, 468–480. [Google Scholar]
- Chiao, J.W.; Chung, F.; Krzeminski, J. Modulation of growth of human prostate cancer cells by the N-acetylcysteine conjugate of phenethyl isothiocyanate. Int. J. Oncol. 2000, 16, 1215–1219. [Google Scholar] [CrossRef] [PubMed]
- Rose, P.; Whiteman, M.; Huang, S.H.; Halliwell, B.; Ong, C.N. beta-Phenylethyl isothiocyanate-mediated apoptosis in hepatoma HepG2 cells. Cell Mol. Life Sci. 2003, 60, 1489–1503. [Google Scholar] [CrossRef]
- Elsa, N.H.; ElSherbiny, N.M.; Shebl, A.M.; Eissa, L.A.; El-Shishtawy, M.M. Phenethyl isothiocyanate potentiates anti-tumour effect of doxorubicin through Akt-dependent pathway. Cell Biochem. Funct. 2015, 33, 541–551. [Google Scholar]
- Sarkars, R.; Mukherjee, S.; Roy, M. Targeting heat shock proteins by phenethyl isothiocyanate results in cell-cycle arrest and apoptosis of human breast cancer cells. Nutr. Cancer 2013, 65, 480–493. [Google Scholar] [CrossRef]
- Hasegawa, T.; Nishino, H.; Iwashima, A. Isothiocyanates inhibit cell cycle progression of HeLa cells at G2/M phase. Anticancer Drugs. 1993, 4, 273–279. [Google Scholar] [CrossRef]
- Gupta, P.; Srivastava, S.K. Antitumor activity of phenethyl isothiocyanate in HER2-positive breast cancer models. BMC Med. 2012, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Visentin, M.; Tava, A.; Iori, R.; Palmieri, S. Isolation and identification for trans-4-(methylthio)-3-butenyl glucosinolate from radish roots (Raphanus sativus L.). J. Agr. Food Chem. 1992, 40, 1687–1691. [Google Scholar] [CrossRef]
- Lawson, A.P.; Long, M.J.; Coffey, R.T.; Qian, Y.; Weerapana, E.; El Oualid, F.; Hedstrom, L. Naturally occurring isothiocyanates exert anticancer effects by inhibiting deubiquitinating enzymes. Cancer Res. 2015, 75, 5130–5142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, Z.S.O.; Li, X.; Li, F.; Cheaito, H.A.; Patel, K.; Mosallam, E.S.M.; Dou, Q.P. Computational and biochemical studies of isothiocyanates as inhibitors of proteasomal cysteine deubiquitinases in human cancer cells. J. Cell. Biochem. 2018, 119, 9006–9016. [Google Scholar] [CrossRef] [PubMed]
- Yousaf, M.; Ahmad, M.; Bhatti, I.A.; Nasir, A.; Hasan, M.; Jian, X.; Mahmood, N. In Vivo and In Vitro Monitoring of Amyloid Aggregation via BSA@ FGQDs Multimodal Probe. ACS Sensors 2018, 4, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.; Yang, W.; Ju, Y.; Chu, X.; Wang, Y.; Deng, Y.; Hou, Y. Biocompatibility of iron carbide and detection of metals ions signaling proteomic analysis via HPLC/ESI-Orbitrap. Nano Res. 2017, 10, 1912–1923. [Google Scholar] [CrossRef]
- Barillari, J.; Gueyrard, D.; Rollin, P.; Iori, R. Barbarea verna as a source of 2-phenylethyl glucosinolate, precursor of cancer chemopreventive phenylethyl isothiocyanate. Fitoterapia 2001, 72, 760–764. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid calorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Incubation (h) | HepG2 Cells (IC50) | Compound | Incubation (h) | MCF-7 Cells (IC50) |
|---|---|---|---|---|---|
| GNST | 24 | ND | GNST | 24 | ND |
| 48 | ND | 48 | ND | ||
| 72 | ND | 72 | ND | ||
| GNST-ITC | 24 | 22.54 ± 0.08 µM | GNST-ITC | 24 | 10.67 ± 0.05 µM |
| 48 | 12.14 ± 0.13 µM | 48 | 6.19 ± 0.26 µM | ||
| 72 | 7.83 ± 0.34 µM | 72 | 5.02 ± 0.08 µM | ||
| Cisplatin | 24 | 7.78 ± 0.4 µM | Paclitaxel | 24 | 6.92 ± 0.37 nM |
| 48 | 2.56 ± 0.44 µM | 48 | 4.28 ± 0.77 nM | ||
| 72 | 1.99 ± 0.23 µM | 72 | 3.21 ± 0.47 nM |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arumugam, A.; Ibrahim, M.D.; Kntayya, S.B.; Mohd Ain, N.; Iori, R.; Galletti, S.; Ioannides, C.; Abdull Razis, A.F. Induction of Apoptosis by Gluconasturtiin-Isothiocyanate (GNST-ITC) in Human Hepatocarcinoma HepG2 Cells and Human Breast Adenocarcinoma MCF-7 Cells. Molecules 2020, 25, 1240. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25051240
Arumugam A, Ibrahim MD, Kntayya SB, Mohd Ain N, Iori R, Galletti S, Ioannides C, Abdull Razis AF. Induction of Apoptosis by Gluconasturtiin-Isothiocyanate (GNST-ITC) in Human Hepatocarcinoma HepG2 Cells and Human Breast Adenocarcinoma MCF-7 Cells. Molecules. 2020; 25(5):1240. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25051240
Chicago/Turabian StyleArumugam, Asvinidevi, Muhammad Din Ibrahim, Saie Brindha Kntayya, Nooraini Mohd Ain, Renato Iori, Stefania Galletti, Costas Ioannides, and Ahmad Faizal Abdull Razis. 2020. "Induction of Apoptosis by Gluconasturtiin-Isothiocyanate (GNST-ITC) in Human Hepatocarcinoma HepG2 Cells and Human Breast Adenocarcinoma MCF-7 Cells" Molecules 25, no. 5: 1240. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25051240