
Preconditioning of Adipose-Derived Mesenchymal Stem-Like Cells with Eugenol Potentiates Their Migration and Proliferation In Vitro and Therapeutic Abilities in Rat Hepatic Fibrosis

 ,
, 
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Characterization of Markers Expression in the Isolated ASCs
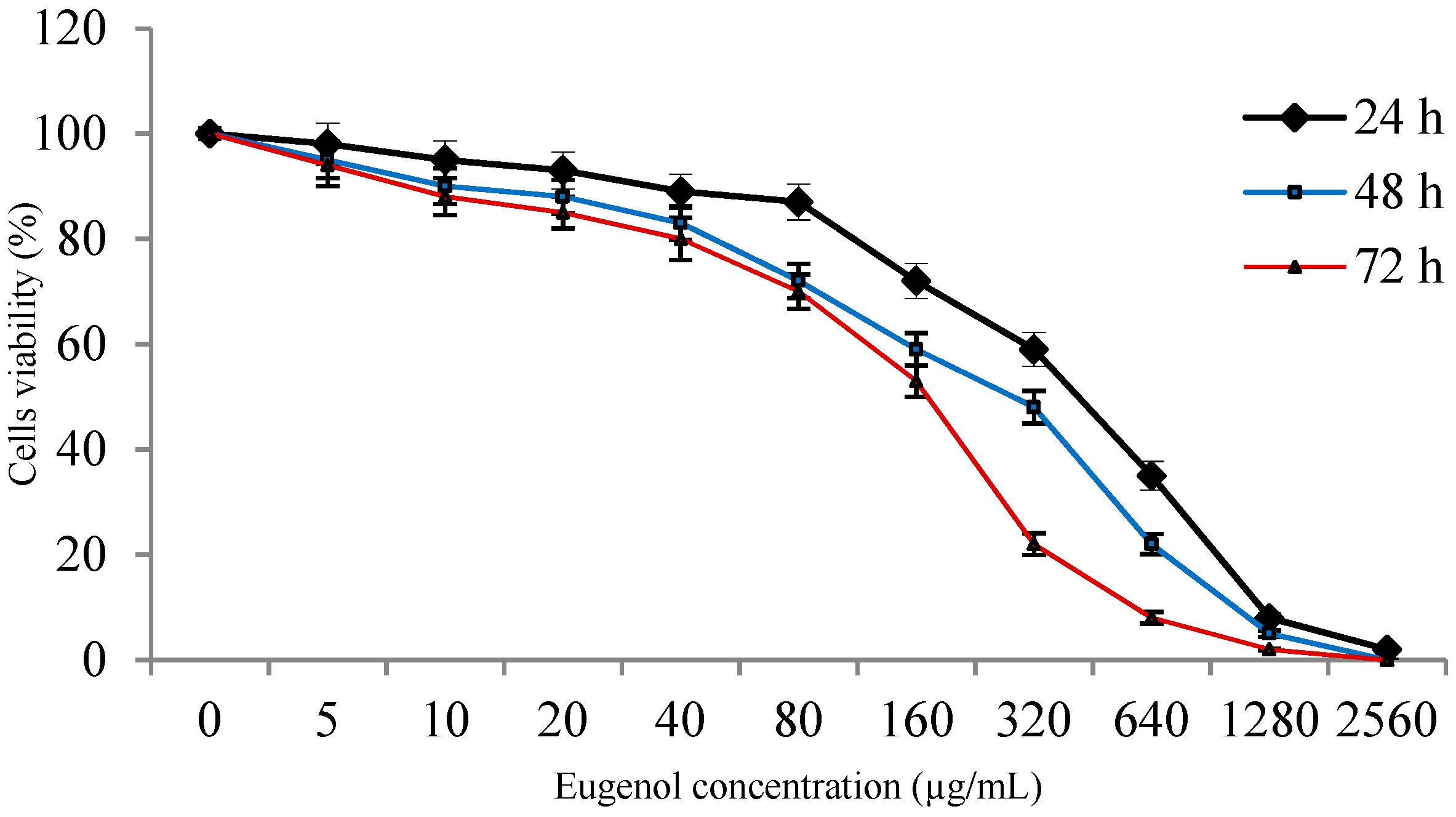
2.2. Viability of ASCs with Eugenol
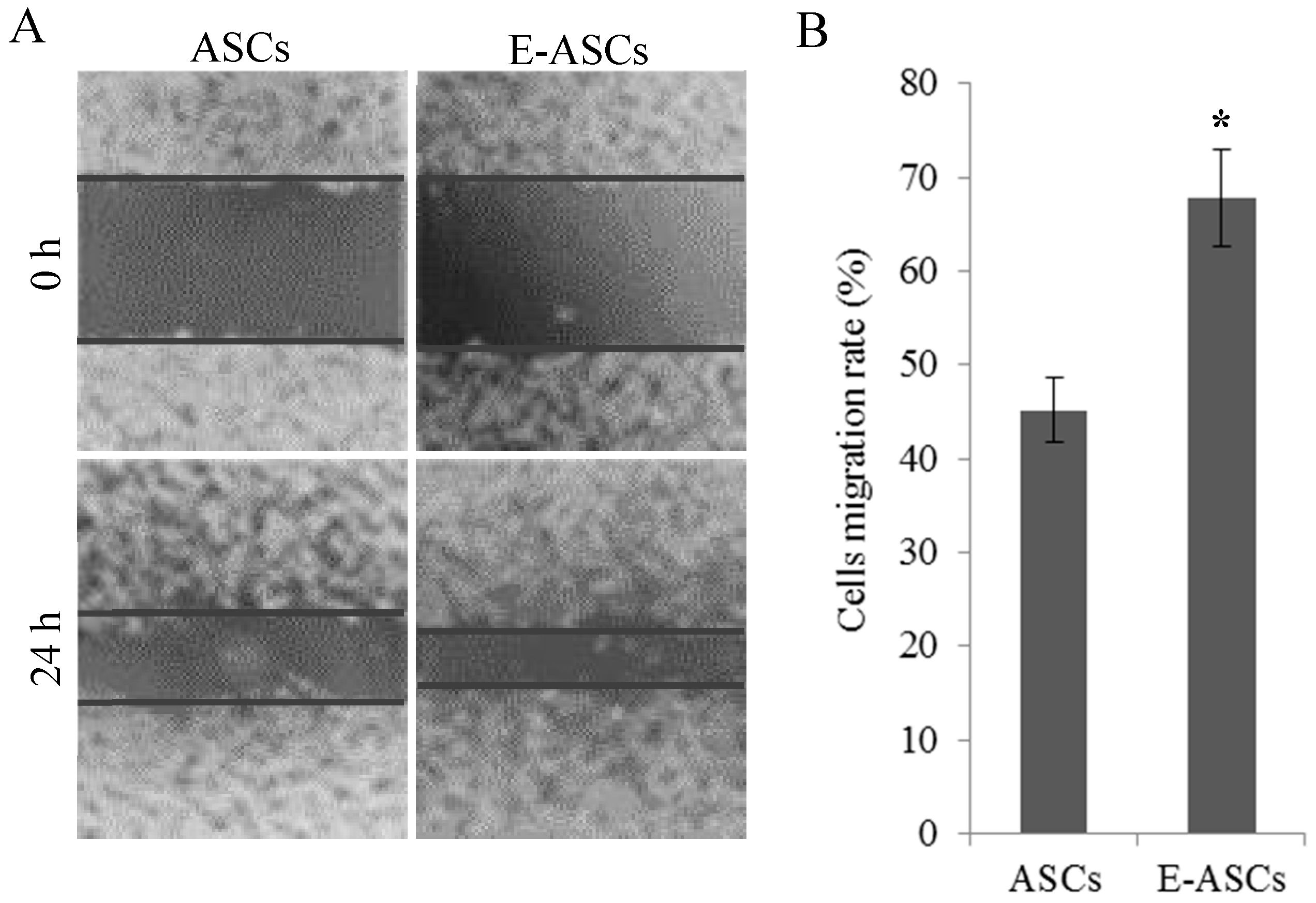
2.3. The In Vitro Migration Ability of ASCs is Improved by Eugenol
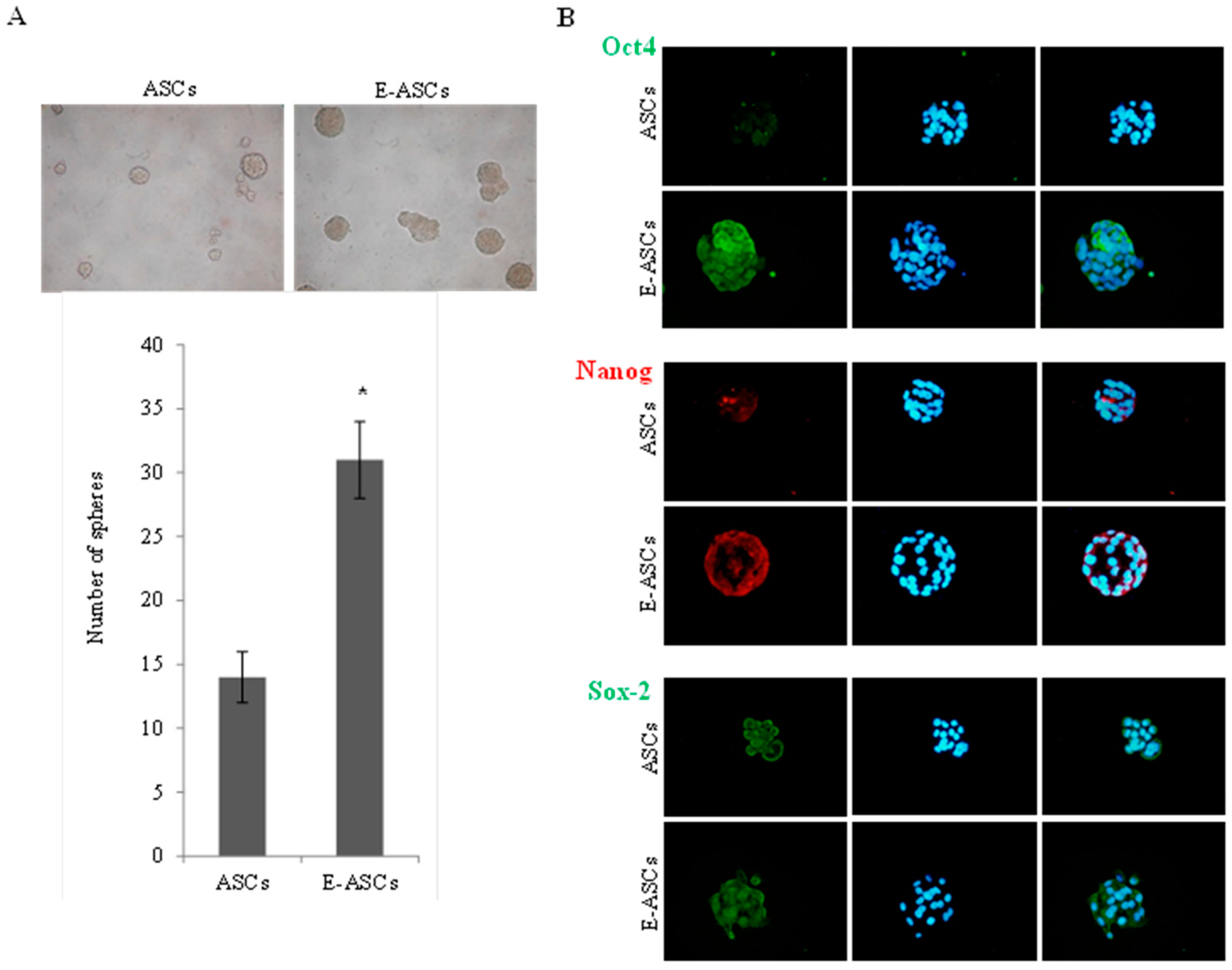
2.4. Effect of Eugenol on Self-Renewal Characteristics of ASCs
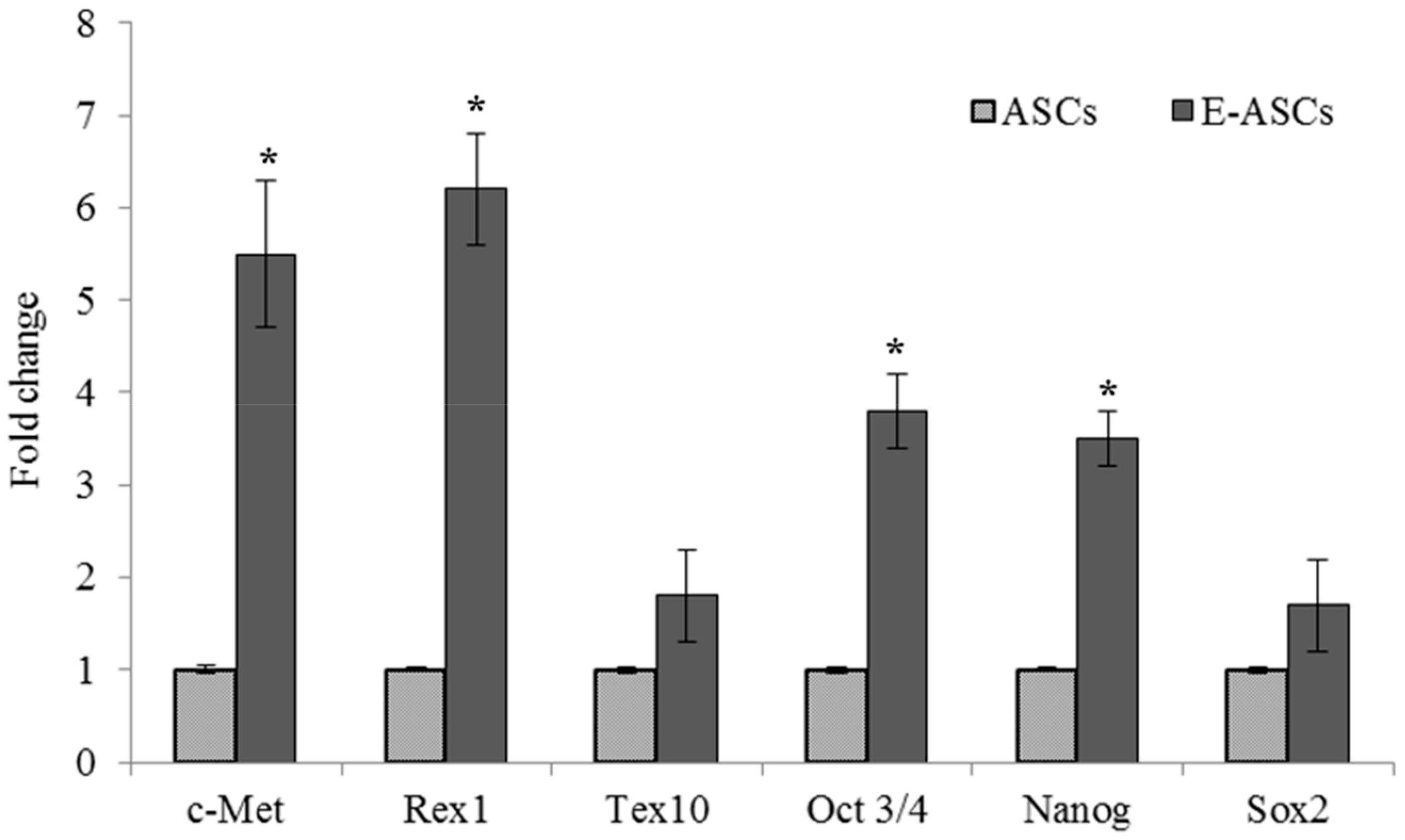
2.5. Gene Expression of ASCs Treated with Eugenol
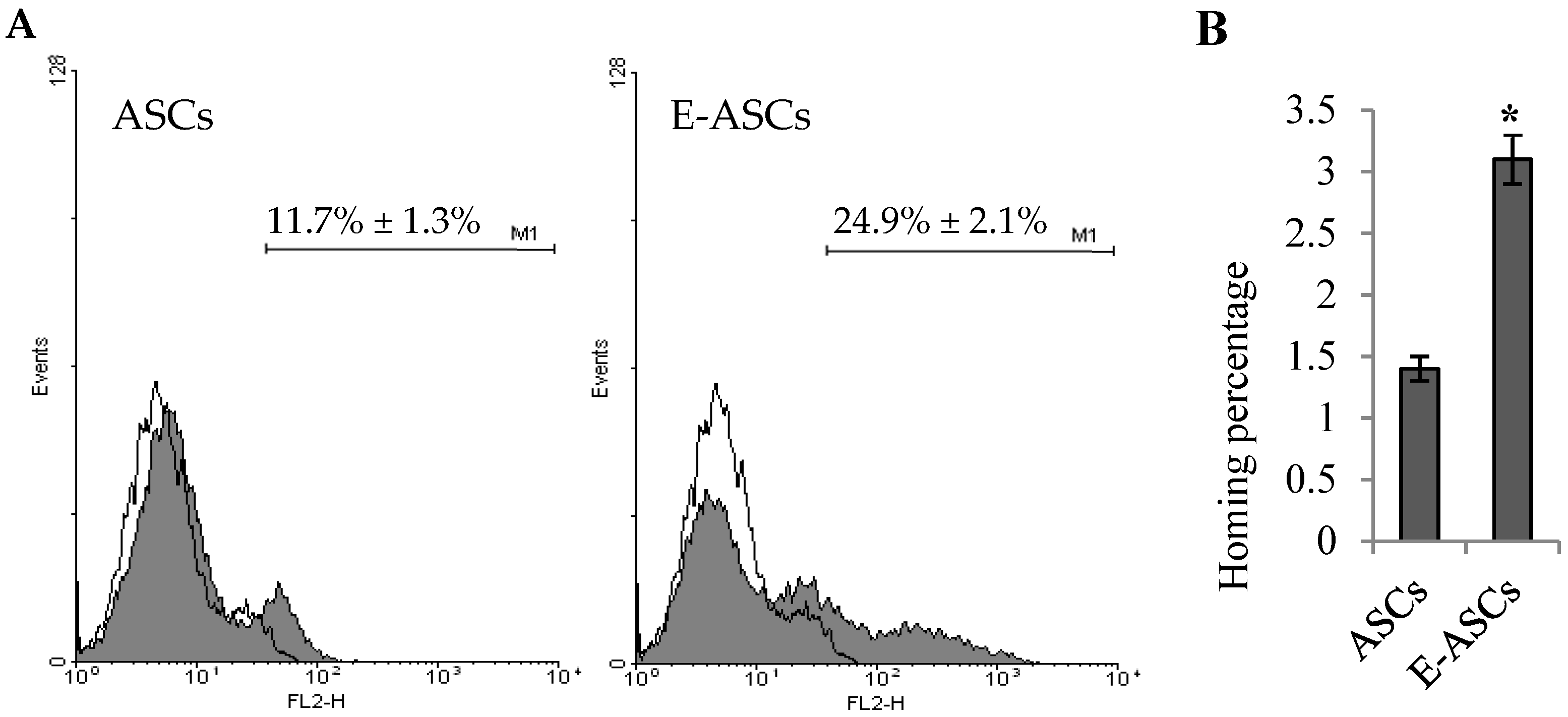
2.6. Homing Ability of ASCs is Enhanced by Eugenol
2.7. Serum Biochemical Investigations
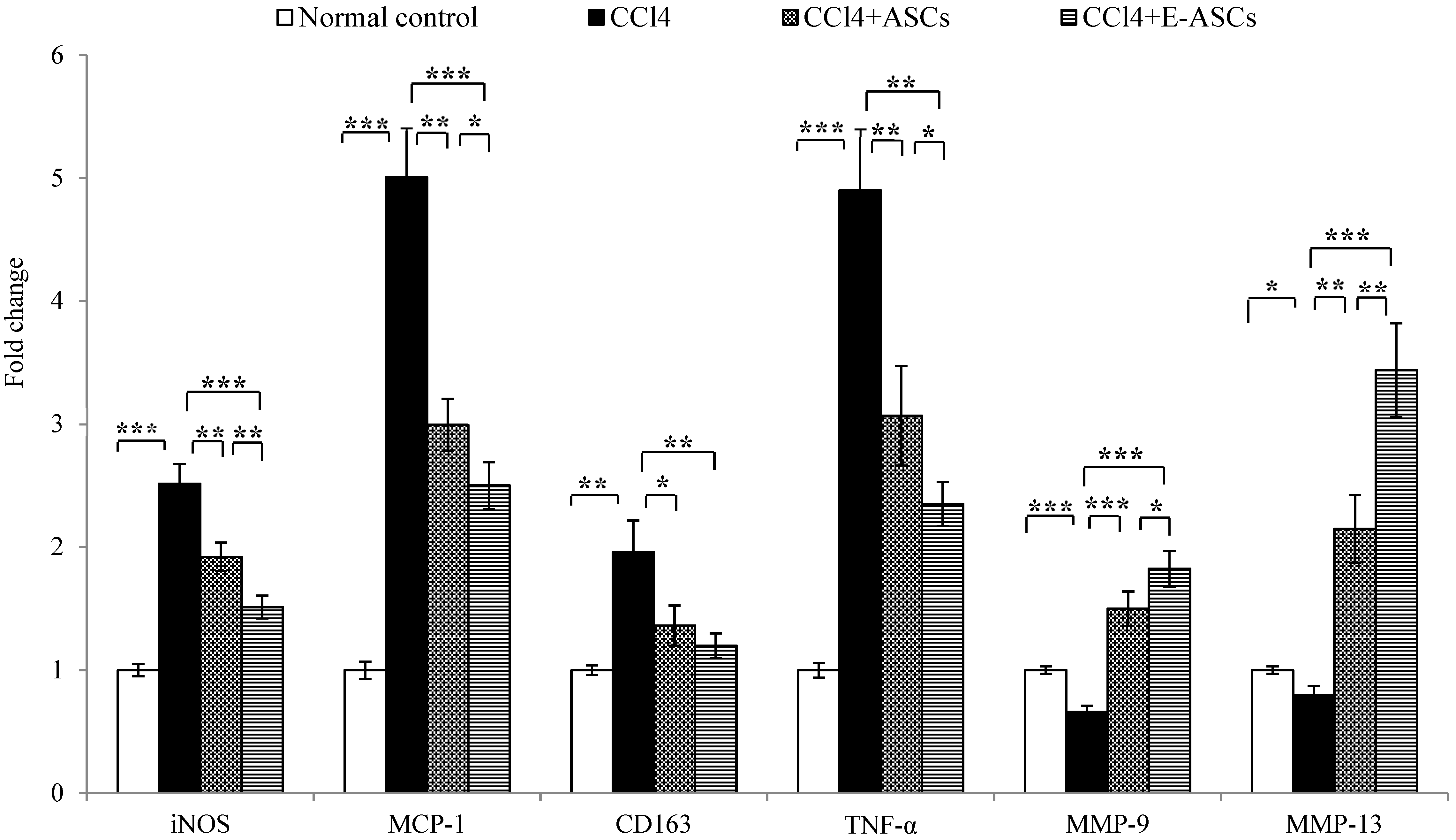
2.8. Gene Expression for Rats of Different Groups
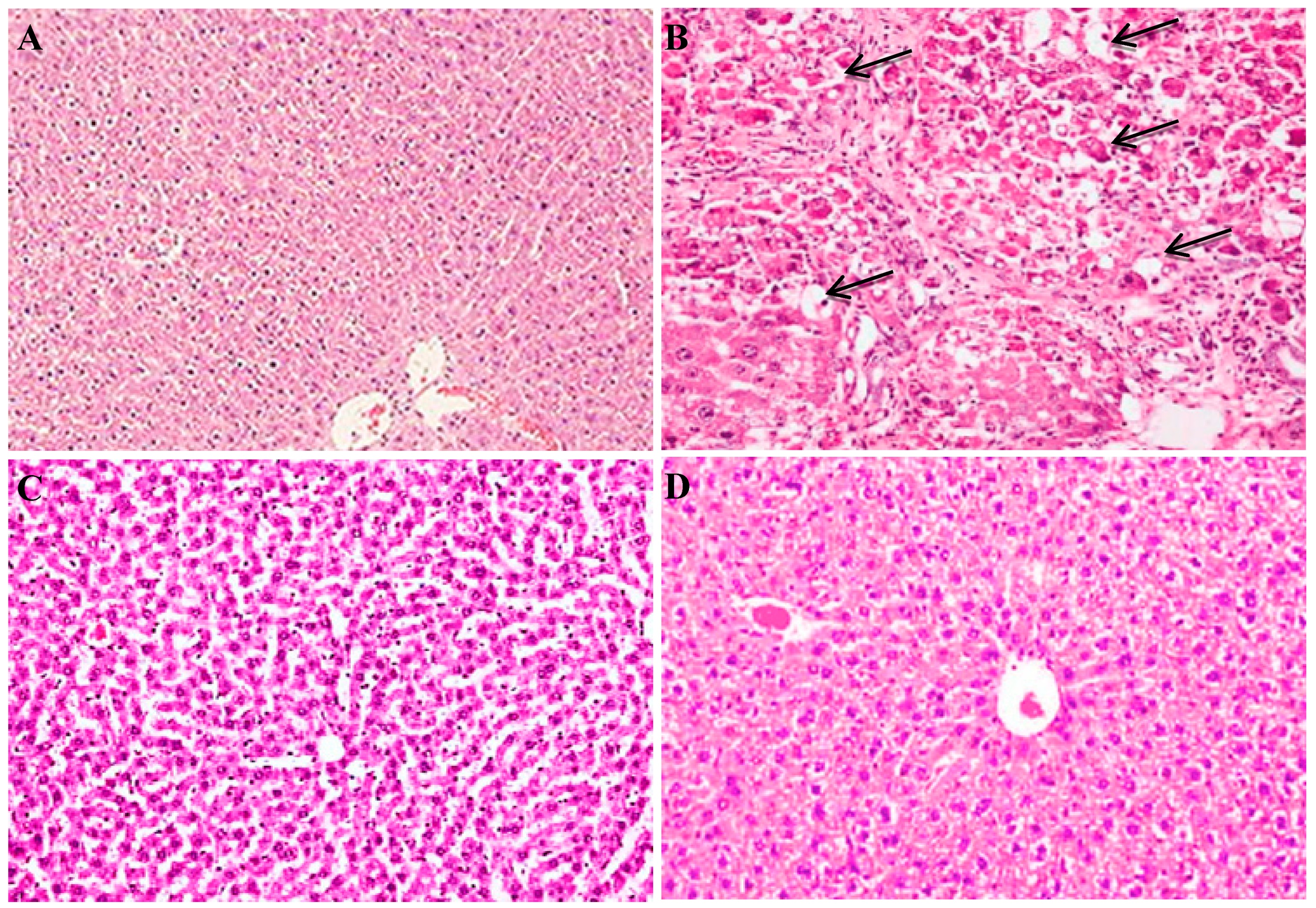
2.9. Histopathological Examination for Rats of Different Groups
3. Discussion
4. Materials and Methods
4.1. Cells Isolation and Characterization
4.2. Cell Viability Assay
4.3. Scratch Migration Assay
4.4. Sphere Formation Assay and Immunofluorescence Staining
4.5. Animals and the Experimental Design
4.6. Flow Cytometry for Homing Assessment
4.7. Biochemical Investigations
4.8. Gene Expression Analysis using Quantitative Real-Time PCR
4.9. Histological Examination
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mortezaee, K.; Minaii, B.; Sabbaghziarani, F.; Ragerdi Kashani, I.; Hassanzadeh, G.; Pasbakhsh, P.; Barbarestani, M.; Latifpour, M. Retinoic Acid as the Stimulating Factor for Differentiation of Wharton’s Jelly-Mesenchymal Stem Cells into Hepatocyte-like Cells. Avicenna J. Med. Biotechnol. 2015, 7, 106–112. [Google Scholar] [PubMed]
- Motawi, T.M.; Atta, H.M.; Sadik, N.A.; Azzam, M. The therapeutic effects of bone marrow-derived mesenchymal stem cells and simvastatin in a rat model of liver fibrosis. Cell Biochem. Biophys. 2014, 68, 111–125. [Google Scholar] [CrossRef] [PubMed]
- Tanimoto, H.; Terai, S.; Taro, T.; Murata, Y.; Fujisawa, K.; Yamamoto, N.; Sakaida, I. Improvement of liver fibrosis by infusion of cultured cells derived from human bone marrow. Cell Tissue Res. 2013, 354, 717–728. [Google Scholar] [CrossRef]
- De Luna-Saldivar, M.M.; Marino-Martinez, I.A.; Franco-Molina, M.A.; Rivera-Morales, L.G.; Alarcon-Galvan, G.; Cordero-Perez, P.; Rojas-Martinez, A.; Rodriguez-Padilla, C.; Munoz-Espinosa, L.E. Advantages of adipose tissue stem cells over CD34(+) mobilization to decrease hepatic fibrosis in Wistar rats. Ann. Hepatol. 2019, 18, 620–626. [Google Scholar] [CrossRef] [PubMed]
- Gugjoo, M.B.; Amarpal; Chandra, V.; Wani, M.Y.; Dhama, K.; Sharma, G.T. Mesenchymal Stem Cell Research in Veterinary Medicine. Curr. Stem Cell Res. Ther. 2018, 13, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Que, Y.; Lv, D.; Bi, S.; Xu, Z.; Wang, D.; Zhang, Z. Overexpression of TG2 enhances the differentiation of ectomesenchymal stem cells into neuronlike cells and promotes functional recovery in adult rats following spinal cord injury. Mol. Med. Rep. 2019, 20, 2763–2773. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.T.; Xie, Y.; Li, D.S.; Zhu, Y.Y.; Zhang, X.L.; Feng, Y.L.; Chen, Y.P.; Xu, L.J.; Liao, P.F.; Wang, G. Comparison of biological characteristics of mesenchymal stem cells derived from the human umbilical cord and decidua parietalis. Mol. Med. Rep. 2019, 20, 633–639. [Google Scholar] [CrossRef]
- Li, Q.; Zhou, X.; Shi, Y.; Li, J.; Zheng, L.; Cui, L.; Zhang, J.; Wang, L.; Han, Z.; Han, Y.; et al. In vivo tracking and comparison of the therapeutic effects of MSCs and HSCs for liver injury. PLoS ONE 2013, 8, e62363. [Google Scholar] [CrossRef]
- Shi, Y.; Hu, G.; Su, J.; Li, W.; Chen, Q.; Shou, P.; Xu, C.; Chen, X.; Huang, Y.; Zhu, Z.; et al. Mesenchymal stem cells: A new strategy for immunosuppression and tissue repair. Cell Res. 2010, 20, 510–518. [Google Scholar] [CrossRef]
- Koike, C.; Zhou, K.; Takeda, Y.; Fathy, M.; Okabe, M.; Yoshida, T.; Nakamura, Y.; Kato, Y.; Nikaido, T. Characterization of amniotic stem cells. Cell Reprogram. 2014, 16, 298–305. [Google Scholar] [CrossRef] [Green Version]
- Wolbank, S.; Peterbauer, A.; Fahrner, M.; Hennerbichler, S.; van Griensven, M.; Stadler, G.; Redl, H.; Gabriel, C. Dose-dependent immunomodulatory effect of human stem cells from amniotic membrane: A comparison with human mesenchymal stem cells from adipose tissue. Tissue Eng. 2007, 13, 1173–1183. [Google Scholar] [CrossRef]
- Saad Eldien, H.M.; Abdel-Aziz, H.O.; Sayed, D.; Mubarak, W.; Hareedy, H.H.G.; Mansor, S.G.; Yoshida, T.; Fathy, M. Periostin expression and characters of human adipose tissue-derived mesenchymal stromal cells were aberrantly affected by in vitro cultivation. Stem Cell Investig. 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Mortezaee, K.; Pasbakhsh, P.; Ragerdi Kashani, I.; Sabbaghziarani, F.; Omidi, A.; Zendedel, A.; Ghasemi, S.; Dehpour, A.R. Melatonin Pretreatment Enhances the Homing of Bone Marrow-derived Mesenchymal Stem Cells Following Transplantation in a Rat Model of Liver Fibrosis. Iran Biomed. J. 2016, 20, 207–216. [Google Scholar] [CrossRef]
- Tang, Y.; Cai, B.; Yuan, F.; He, X.; Lin, X.; Wang, J.; Wang, Y.; Yang, G.Y. Melatonin Pretreatment Improves the Survival and Function of Transplanted Mesenchymal Stem Cells after Focal Cerebral Ischemia. Cell Transplant. 2014, 23, 1279–1291. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L. Hepatic stellate cells: Protean, multifunctional, and enigmatic cells of the liver. Physiol. Rev. 2008, 88, 125–172. [Google Scholar] [CrossRef] [PubMed]
- Fathy, M.; Nikaido, T. In vivo modulation of iNOS pathway in hepatocellular carcinoma by Nigella sativa. Environ. Health Prev. Med. 2013, 18, 377–385. [Google Scholar] [CrossRef]
- Fathy, M.; Nikaido, T. In vivo attenuation of angiogenesis in hepatocellular carcinoma by Nigella sativa. Turk. J. Med. Sci. 2018, 48, 178–186. [Google Scholar] [CrossRef]
- Wang, J.; Cen, P.; Chen, J.; Fan, L.; Li, J.; Cao, H.; Li, L. Role of mesenchymal stem cells, their derived factors, and extracellular vesicles in liver failure. Stem Cell Res. Ther. 2017, 8, 137. [Google Scholar] [CrossRef] [Green Version]
- Sisakhtnezhad, S.; Heidari, M.; Bidmeshkipour, A. Eugenol enhances proliferation and migration of mouse bone marrow-derived mesenchymal stem cells in vitro. Environ. Toxicol. Pharmacol. 2018, 57, 166–174. [Google Scholar] [CrossRef]
- Islam, S.S.; Al-Sharif, I.; Sultan, A.; Al-Mazrou, A.; Remmal, A.; Aboussekhra, A. Eugenol potentiates cisplatin anti-cancer activity through inhibition of ALDH-positive breast cancer stem cells and the NF-kappaB signaling pathway. Mol. Carcinog. 2018, 57, 333–346. [Google Scholar] [CrossRef]
- Yi, J.L.; Shi, S.; Shen, Y.L.; Wang, L.; Chen, H.Y.; Zhu, J.; Ding, Y. Myricetin and methyl eugenol combination enhances the anticancer activity, cell cycle arrest and apoptosis induction of cis-platin against HeLa cervical cancer cell lines. Int. J. Clin. Exp. Pathol. 2015, 8, 1116–1127. [Google Scholar] [PubMed]
- Shaban, S.; El-Husseny, M.W.A.; Abushouk, A.I.; Salem, A.M.A.; Mamdouh, M.; Abdel-Daim, M.M. Effects of Antioxidant Supplements on the Survival and Differentiation of Stem Cells. Oxid. Med. Cell. Longev. 2017, 2017, 5032102. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, O.; Okabe, M.; Yoshida, T.; Fathy, M.; Saito, S.; Miyawaki, T.; Nikaido, T. Stemness of human Wharton’s jelly mesenchymal cells is maintained by floating cultivation. Cell Reprogram. 2012, 14, 448–455. [Google Scholar] [CrossRef]
- Zhou, K.; Koike, C.; Yoshida, T.; Okabe, M.; Fathy, M.; Kyo, S.; Kiyono, T.; Saito, S.; Nikaido, T. Establishment and characterization of immortalized human amniotic epithelial cells. Cell Reprogram. 2013, 15, 55–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathy, M.; Okabe, M.; Saad Eldien, H.M.; Yoshida, T. AT-MSCs Antifibrotic Activity is Improved by Eugenol through Modulation of TGF-beta/Smad Signaling Pathway in Rats. Molecules 2020, 25, 348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortezaee, K.; Khanlarkhani, N.; Sabbaghziarani, F.; Nekoonam, S.; Majidpoor, J.; Hosseini, A.; Pasbakhsh, P.; Kashani, I.R.; Zendedel, A. Preconditioning with melatonin improves therapeutic outcomes of bone marrow-derived mesenchymal stem cells in targeting liver fibrosis induced by CCl4. Cell Tissue Res. 2017, 369, 303–312. [Google Scholar] [CrossRef]
- Fathy, M.; Khalifa, E.; Fawzy, M.A. Modulation of inducible nitric oxide synthase pathway by eugenol and telmisartan in carbon tetrachloride-induced liver injury in rats. Life Sci. 2019, 216, 207–214. [Google Scholar] [CrossRef]
- Fathy, M.; Fawzy, M.A.; Hintzsche, H.; Nikaido, T.; Dandekar, T.; Othman, E.M. Eugenol Exerts Apoptotic Effect and Modulates the Sensitivity of HeLa Cells to Cisplatin and Radiation. Molecules 2019, 24, 3979. [Google Scholar] [CrossRef] [Green Version]
- Kabuto, H.; Yamanushi, T.T. Effects of zingerone [4-(4-hydroxy-3-methoxyphenyl)-2-butanone] and eugenol [2-methoxy-4-(2-propenyl)phenol] on the pathological progress in the 6-hydroxydopamine-induced Parkinson’s disease mouse model. Neurochem. Res. 2011, 36, 2244–2249. [Google Scholar] [CrossRef]
- Kamatou, G.P.; Vermaak, I.; Viljoen, A.M. Eugenol--from the remote Maluku Islands to the international market place: A review of a remarkable and versatile molecule. Molecules 2012, 17, 6953–6981. [Google Scholar] [CrossRef]
- Nagura, S.; Otaka, S.; Koike, C.; Okabe, M.; Yoshida, T.; Fathy, M.; Fukahara, K.; Yoshimura, N.; Misaki, T.; Nikaido, T. Effect of exogenous Oct4 overexpression on cardiomyocyte differentiation of human amniotic mesenchymal cells. Cell Reprogram. 2013, 15, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Otaka, S.; Nagura, S.; Koike, C.; Okabe, M.; Yoshida, T.; Fathy, M.; Yanagi, K.; Misaki, T.; Nikaido, T. Selective isolation of nanog-positive human amniotic mesenchymal cells and differentiation into cardiomyocytes. Cell Reprogram. 2013, 15, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Wang, H.; Pan, G.; Geng, Y.; Guo, Y.; Pei, D. Regulation of the pluripotency marker Rex-1 by Nanog and Sox2. J. Biol. Chem. 2006, 281, 23319–23325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, M.Y.; Choi, H.; Han, Y.M.; Cho, Y.S. Unveiling the critical role of REX1 in the regulation of human stem cell pluripotency. Stem Cells 2013, 31, 2374–2387. [Google Scholar] [CrossRef]
- Heo, J.S.; Choi, Y.; Kim, H.S.; Kim, H.O. Comparison of molecular profiles of human mesenchymal stem cells derived from bone marrow, umbilical cord blood, placenta and adipose tissue. Int. J. Mol. Med. 2016, 37, 115–125. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, D.R.; Seo, K.W.; Roh, K.H.; Jung, J.W.; Kang, S.K.; Kang, K.S. REX-1 expression and p38 MAPK activation status can determine proliferation/differentiation fates in human mesenchymal stem cells. PLoS ONE 2010, 5, e10493. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.J.; Deng, J.S.; Chiu, C.S.; Liao, J.C.; Hsieh, W.T.; Sheu, M.J.; Wu, C.H. Hispolon Protects against Acute Liver Damage in the Rat by Inhibiting Lipid Peroxidation, Proinflammatory Cytokine, and Oxidative Stress and Downregulating the Expressions of iNOS, COX-2, and MMP-9. Evid. Based Complement. Alternat. Med. 2011, 2012, 480714. [Google Scholar] [CrossRef]
- Calleja, M.A.; Vieites, J.M.; Montero-Melendez, T.; Torres, M.I.; Faus, M.J.; Gil, A.; Suarez, A. The antioxidant effect of beta-caryophyllene protects rat liver from carbon tetrachloride-induced fibrosis by inhibiting hepatic stellate cell activation. Br. J. Nutr. 2013, 109, 394–401. [Google Scholar] [CrossRef]
- Abdel-Salam, O.M.; Sleem, A.A.; Morsy, F.A. Effects of biphenyldimethyl-dicarboxylate administration alone or combined with silymarin in the CCL4 model of liver fibrosis in rats. Sci. World J. 2007, 7, 1242–1255. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Yu, P.; Wang, W.; Zhang, P.; Yang, H.; Li, S.; Zhang, L. Gene expression profiles of various cytokines in mesenchymal stem cells derived from umbilical cord tissue and bone marrow following infection with human cytomegalovirus. Cell. Mol. Biol. Lett. 2014, 19, 140–157. [Google Scholar] [CrossRef]
- Ren, G.; Zhao, X.; Zhang, L.; Zhang, J.; L’Huillier, A.; Ling, W.; Roberts, A.I.; Le, A.D.; Shi, S.; Shao, C.; et al. Inflammatory cytokine-induced intercellular adhesion molecule-1 and vascular cell adhesion molecule-1 in mesenchymal stem cells are critical for immunosuppression. J. Immunol. 2010, 184, 2321–2328. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Sui, Z. Recent developments in CCR2 antagonists. Expert. Opin. Ther. Pat. 2009, 19, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Foresti, M.L.; Arisi, G.M.; Katki, K.; Montanez, A.; Sanchez, R.M.; Shapiro, L.A. Chemokine CCL2 and its receptor CCR2 are increased in the hippocampus following pilocarpine-induced status epilepticus. J. Neuroinflammation 2009, 6, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semple, B.D.; Bye, N.; Rancan, M.; Ziebell, J.M.; Morganti-Kossmann, M.C. Role of CCL2 (MCP-1) in traumatic brain injury (TBI): Evidence from severe TBI patients and CCL2-/- mice. J. Cereb. Blood Flow Metab. 2010, 30, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.; Vari, F.; Keane, C.; Crooks, P.; Nourse, J.P.; Seymour, L.A.; Gottlieb, D.; Ritchie, D.; Gill, D.; Gandhi, M.K. Serum CD163 and TARC as disease response biomarkers in classical Hodgkin lymphoma. Clin. Cancer Res. 2013, 19, 731–742. [Google Scholar] [CrossRef] [Green Version]
- Moller, H.J. Soluble CD163. Scand. J. Clin. Lab. Invest. 2012, 72, 1–13. [Google Scholar] [CrossRef]
- Buisson, A.C.; Zahm, J.M.; Polette, M.; Pierrot, D.; Bellon, G.; Puchelle, E.; Birembaut, P.; Tournier, J.M. Gelatinase B is involved in the in vitro wound repair of human respiratory epithelium. J. Cell. Physiol. 1996, 166, 413–426. [Google Scholar] [CrossRef]
- Mohan, R.; Chintala, S.K.; Jung, J.C.; Villar, W.V.; McCabe, F.; Russo, L.A.; Lee, Y.; McCarthy, B.E.; Wollenberg, K.R.; Jester, J.V.; et al. Matrix metalloproteinase gelatinase B (MMP-9) coordinates and effects epithelial regeneration. J. Biol. Chem. 2002, 277, 2065–2072. [Google Scholar] [CrossRef] [Green Version]
- Johansson, N.; Ahonen, M.; Kahari, V.M. Matrix metalloproteinases in tumor invasion. Cell. Mol. Life Sci. 2000, 57, 5–15. [Google Scholar] [CrossRef]
- Sakaida, I.; Terai, S.; Yamamoto, N.; Aoyama, K.; Ishikawa, T.; Nishina, H.; Okita, K. Transplantation of bone marrow cells reduces CCl4-induced liver fibrosis in mice. Hepatology 2004, 40, 1304–1311. [Google Scholar] [CrossRef]
- Watanabe, T.; Niioka, M.; Hozawa, S.; Kameyama, K.; Hayashi, T.; Arai, M.; Ishikawa, A.; Maruyama, K.; Okazaki, I. Gene expression of interstitial collagenase in both progressive and recovery phase of rat liver fibrosis induced by carbon tetrachloride. J. Hepatol. 2000, 33, 224–235. [Google Scholar] [CrossRef]
- Hardjo, M.; Miyazaki, M.; Sakaguchi, M.; Masaka, T.; Ibrahim, S.; Kataoka, K.; Huh, N.H. Suppression of carbon tetrachloride-induced liver fibrosis by transplantation of a clonal mesenchymal stem cell line derived from rat bone marrow. Cell Transplant. 2009, 18, 89–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szilvassy, S.J.; Meyerrose, T.E.; Ragland, P.L.; Grimes, B. Differential homing and engraftment properties of hematopoietic progenitor cells from murine bone marrow, mobilized peripheral blood, and fetal liver. Blood 2001, 98, 2108–2115. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | ALT (U/L) | AST (U/L) | T. Bilirubin (mg/dL) | Hyaluronic Acid (µg/L) | Albumin (g/dL) |
|---|---|---|---|---|---|
| Normal control | 46.1 ± 3.5 | 57 ± 4.1 | 0.6 ± 0.03 | 140 ± 11 | 4.1 ± 0.29 |
| CCl4 | 99 ± 7.2 ᶲ | 110.3 ± 8.2 ᶲ | 2.3 ± 0.23 ᶲ | 310 ± 25 ᶲ | 2.5 ± 0.09 ᶲ |
| CCl4+ASCs | 81 ± 4.3 † | 93.5 ± 5.9 † | 1.4 ± 0.1 †† | 209 ± 10 †† | 3.3 ± 0.2 †† |
| CCl4+E-ASCs | 67.2 ± 3.7 ††,§ | 77 ± 4.9 ††,§ | 0.9 ± 0.08 †††,§§ | 189 ± 9 ††,§ | 3.8 ± 0.19 †††,§ |
| Primer | Sequence of the Primer |
|---|---|
| CD105 | Forward: 5′-TGGTCTTTTCGAACGAGAATG-3′ |
| Reverse: 5′-AGCCGGAGGACAATGCTTTTGG-3′. | |
| CD73 | Forward: 5′-ATAGTCACCTCTGACGATGG-3′ |
| Reverse: 5′-ATTTCATCTGGGTGTCTGAG-3′. | |
| CD90 | Forward: 5′-AAGGAGAAACAGGAAACCTC-3′ |
| Reverse: 5′-ACAGACACAGTCCAACTTCC-3′. | |
| CD34 | Forward: 5′-AAGATCTTGGGAGCCACCAGAG-3′ |
| Reverse: 5′-TAGCCCTGGCCTCCACCATTC-3′. | |
| CD45 | Forward: 5′-TGAACATACGGATTGTGAAA-3′ |
| Reverse: 5′-TTTGTTCGGACTGTAAGGTT-3′. | |
| c-Met | Forward: 5′-GAGGACAAGACCACCGAGGATG-3′ |
| Reverse: 5′-GGACAACAGCAGCACCAGAATG-3′ | |
| Oct4 | Forward: 5′-CCCAGCGCCGTGAAGTTGGA-3′ |
| Reverse: 5′-AGAACGCCCAGGGTGAGCCC-3′ | |
| Nanog | Forward: 5′-CCCTTGCCGTTGGGCTGACA-3′ |
| Reverse: 5′-AAGGCGGAGGAGAGGCAGTCT-3′ | |
| Sox2 | Forward: 5′-TAGAGCTAGACTCCGGGCGATGA-3′ |
| Reverse: 5′-TTGCCTTAAACAAGACCACGAAA-3′ | |
| Rex1 | Forward: 5′-ACCCATATCCGCATCCACACC-3′ |
| Reverse: 5′-GTCTCCACCTTCAGCATTTCTTCC-3′ | |
| Tex10 | Forward: 5′-GCTTGACGGTCGCTCGCTCTC-3′ |
| Reverse: 5′-TGATGGGTCGGTCGGTCTGTCTC-3′ | |
| iNOS | Forward: 5′-CACCACCCTCCTTGTTCAAC-3′ |
| Reverse: 5′-CAATCCACAACTCGCTCCAA-3′ | |
| MCP-1 | Forward: 5′-CTGTCTCAGCCAGATGCAGTTAA-3′ |
| Reverse: 5′-AGCCGACTCATTGGGATCAT-3′ | |
| CD163 | Forward: 5′-TGTAGTTCATCATCTTCGGTCC-3′ |
| Reverse: 5′-CACCTACCAAGCGGAGTTGAC-3′ | |
| TNF-α | Forward: 5′-GGCTCCCTCTCATCAGTTCCA-3′ |
| Reverse: 5′-CGCTTGGTGGTTTGCTACGA-3′ | |
| MMP-9 | Forward: 5′-CCACTAAAGGTCGCTCGGAT-3′ |
| Reverse: 5′-GAGTTGCCCCCAGTTACAGT-3′ | |
| MMP-13 | Forward: 5′-TTCTGGTCTTCTGGCACACG-3′ |
| Reverse: 5′-ATGGGAAACATCAGGGCTCC-3′ | |
| GAPDH | Forward: 5′-AGACAGCCGCATCTTCTTGT-3′ |
| Reverse: 5′-TGATGGCAACAATGTCCACT-3′ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fathy, M.; Okabe, M.; M. Othman, E.; Saad Eldien, H.M.; Yoshida, T. Preconditioning of Adipose-Derived Mesenchymal Stem-Like Cells with Eugenol Potentiates Their Migration and Proliferation In Vitro and Therapeutic Abilities in Rat Hepatic Fibrosis. Molecules 2020, 25, 2020. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25092020
Fathy M, Okabe M, M. Othman E, Saad Eldien HM, Yoshida T. Preconditioning of Adipose-Derived Mesenchymal Stem-Like Cells with Eugenol Potentiates Their Migration and Proliferation In Vitro and Therapeutic Abilities in Rat Hepatic Fibrosis. Molecules. 2020; 25(9):2020. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25092020
Chicago/Turabian StyleFathy, Moustafa, Motonori Okabe, Eman M. Othman, Heba M. Saad Eldien, and Toshiko Yoshida. 2020. "Preconditioning of Adipose-Derived Mesenchymal Stem-Like Cells with Eugenol Potentiates Their Migration and Proliferation In Vitro and Therapeutic Abilities in Rat Hepatic Fibrosis" Molecules 25, no. 9: 2020. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules25092020