
A Comparative Analysis of In Vitro Toxicity of Synthetic Zeolites on IMR-90 Human Lung Fibroblast Cells



Abstract
:1. Introduction
2. Results
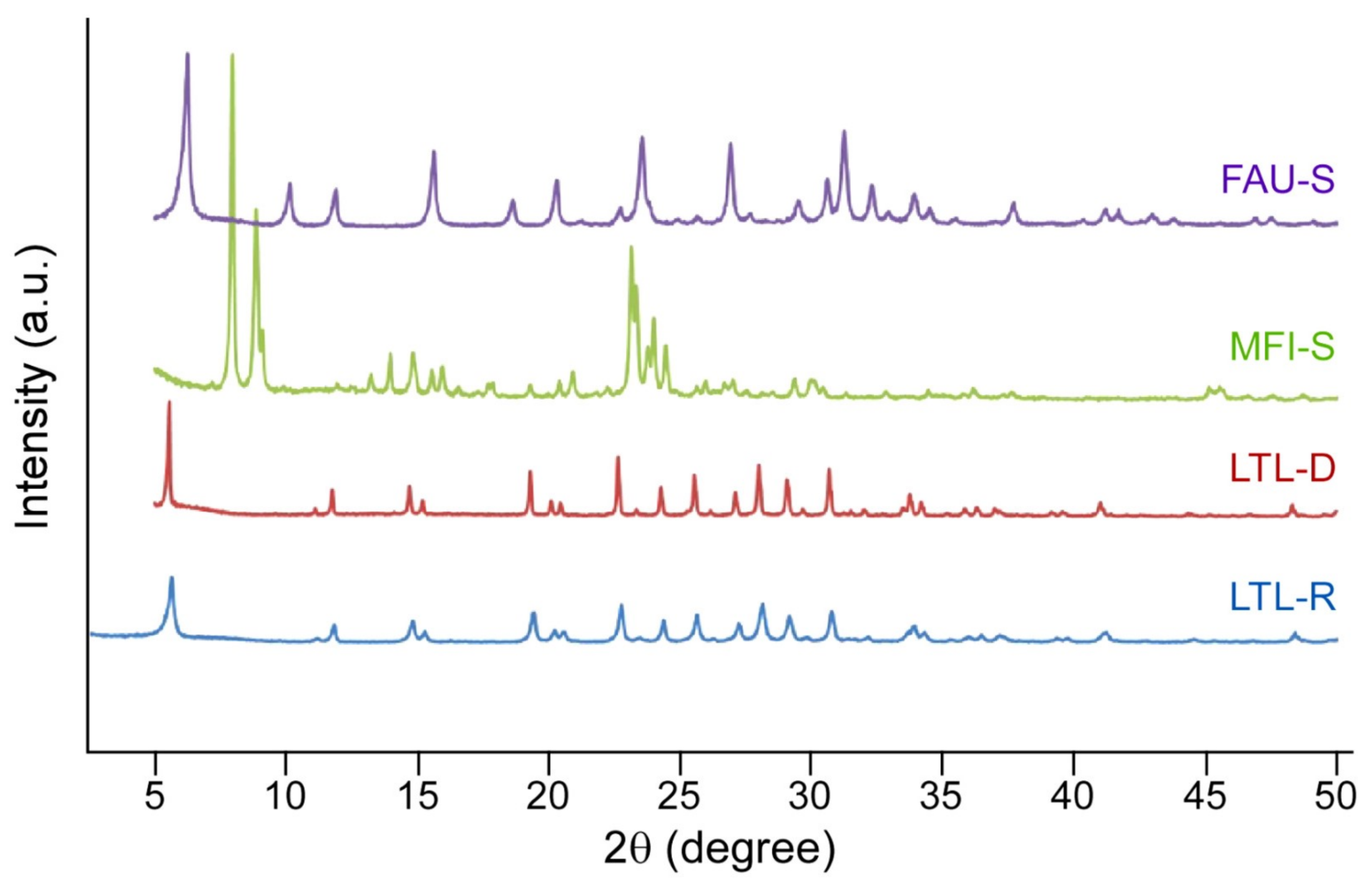
2.1. Physicochemical Properties of Zeolites
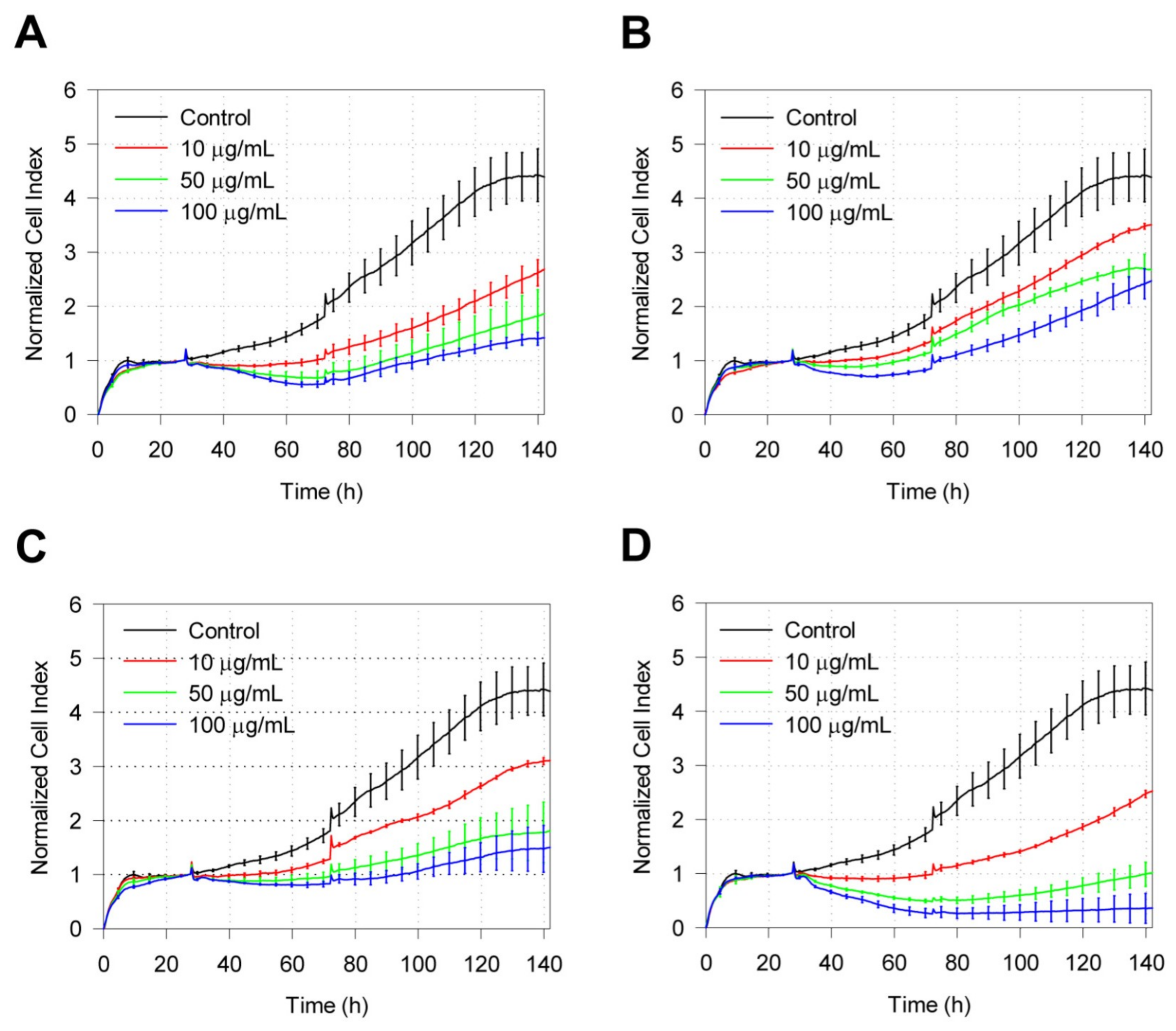
2.2. Real-Time Cell Response of IMR-90 to Zeolite Particles
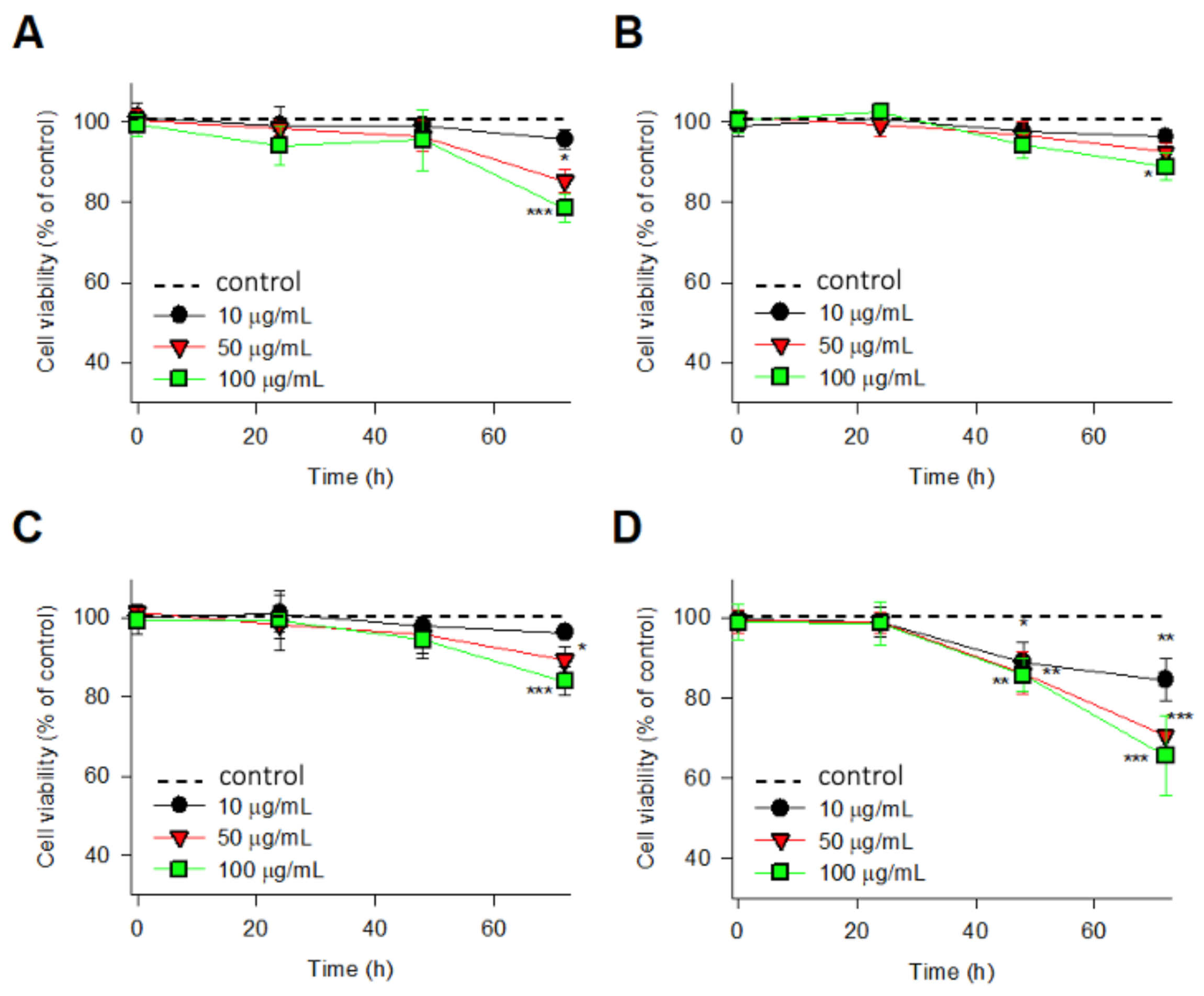
2.3. Cell Viability Study
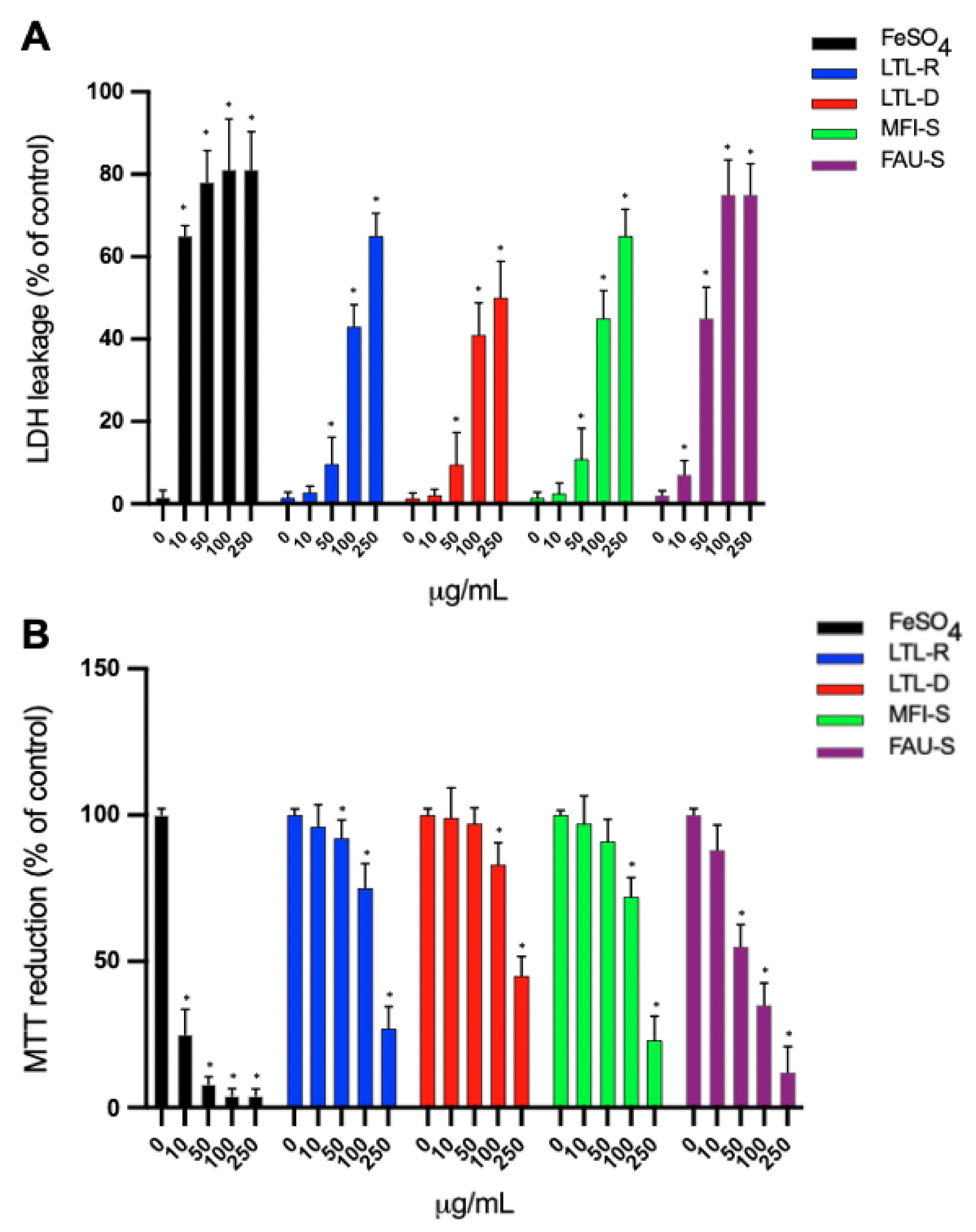
2.4. Cytotoxicity Study
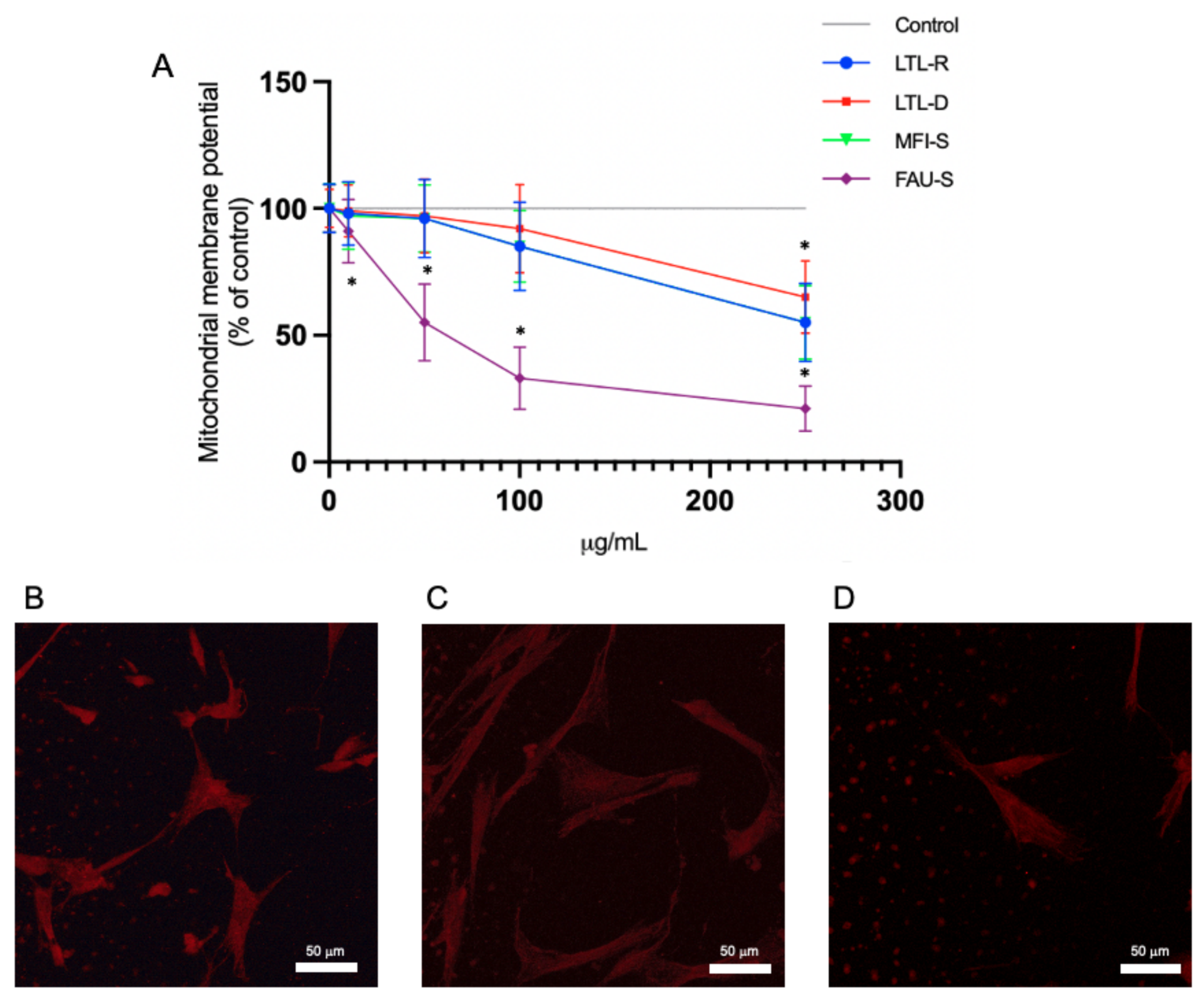
2.5. Mitochondrial Membrane Function Disruption
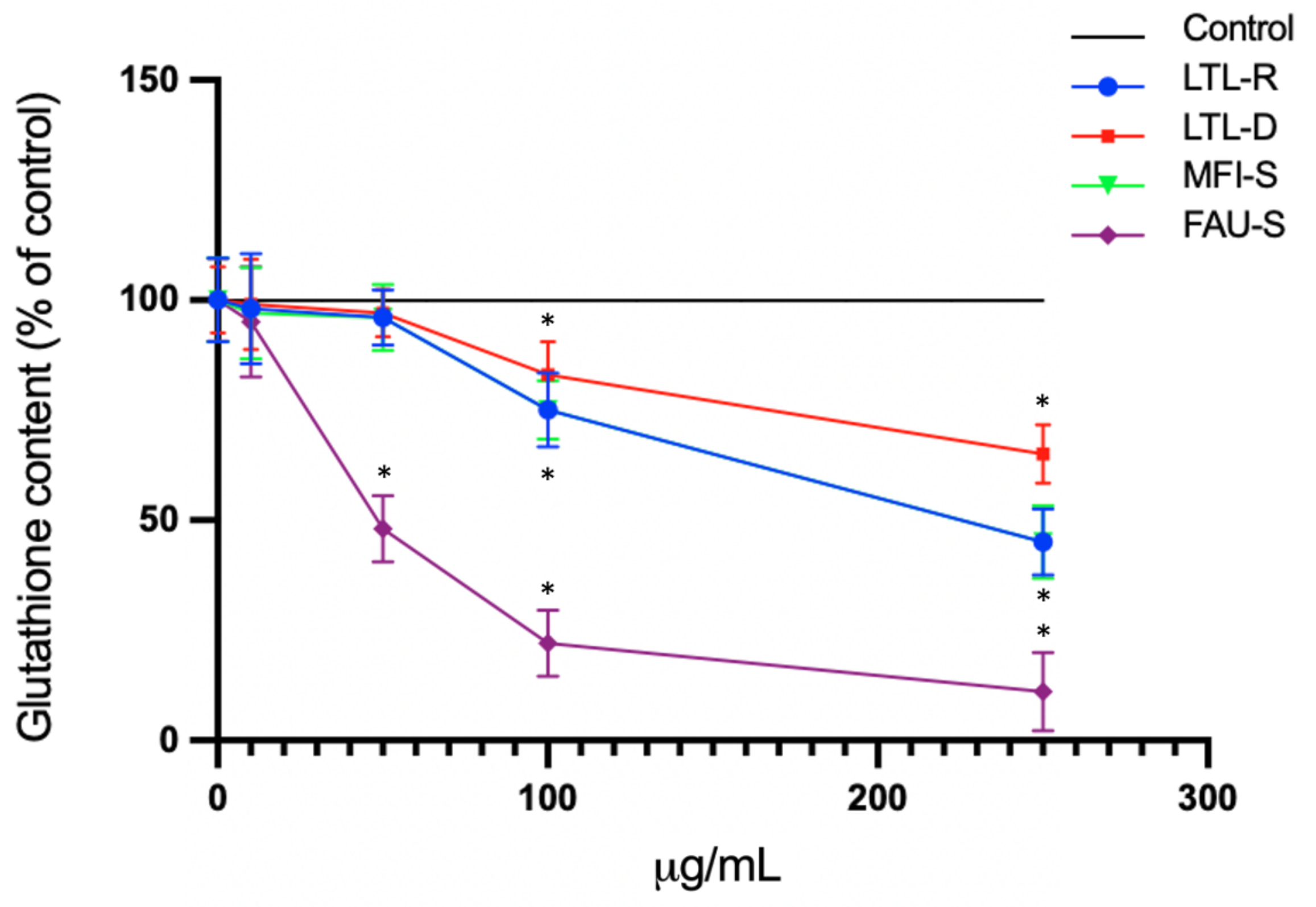
2.6. Total Glutathione Levels
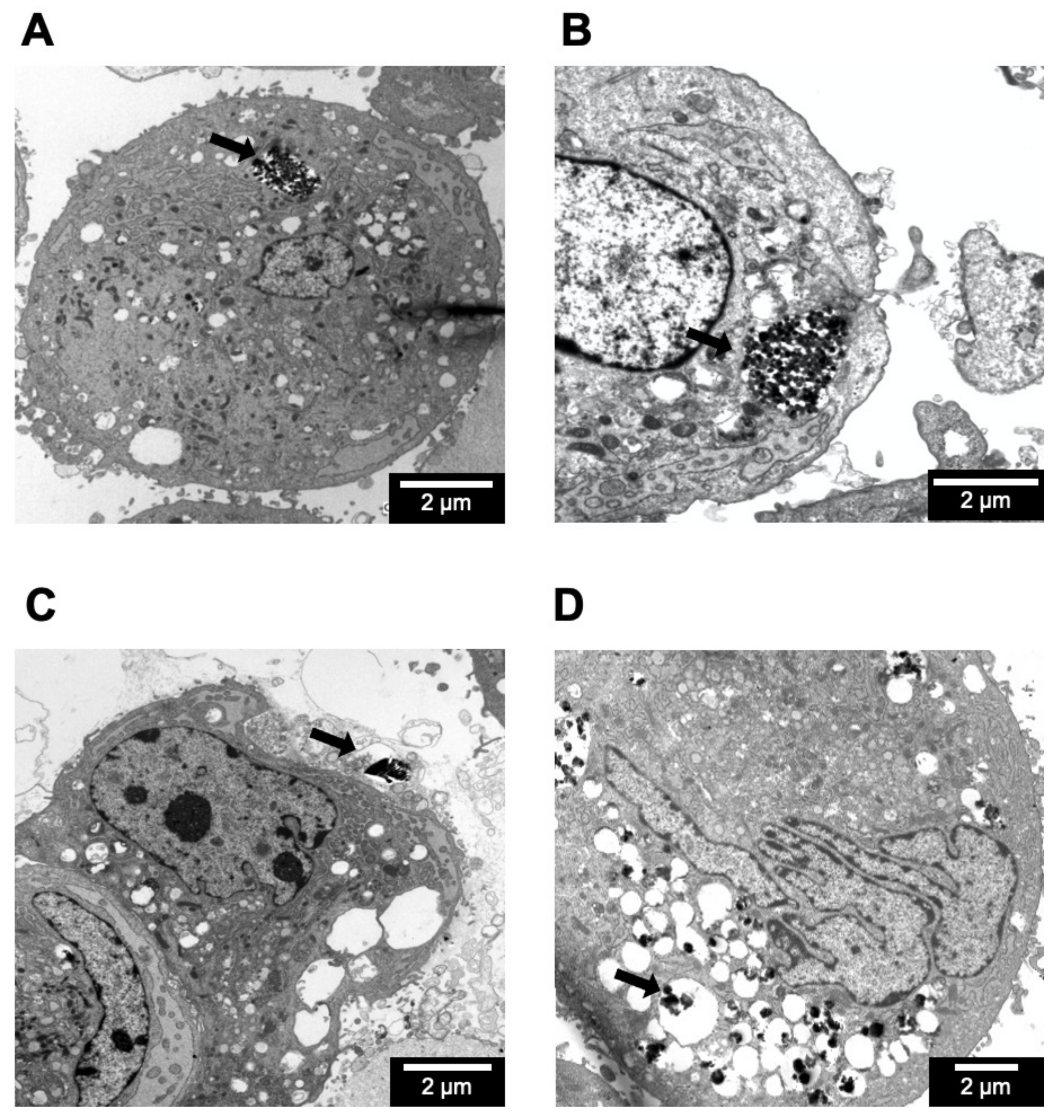
2.7. Cellular Internalization of Zeolite Particles
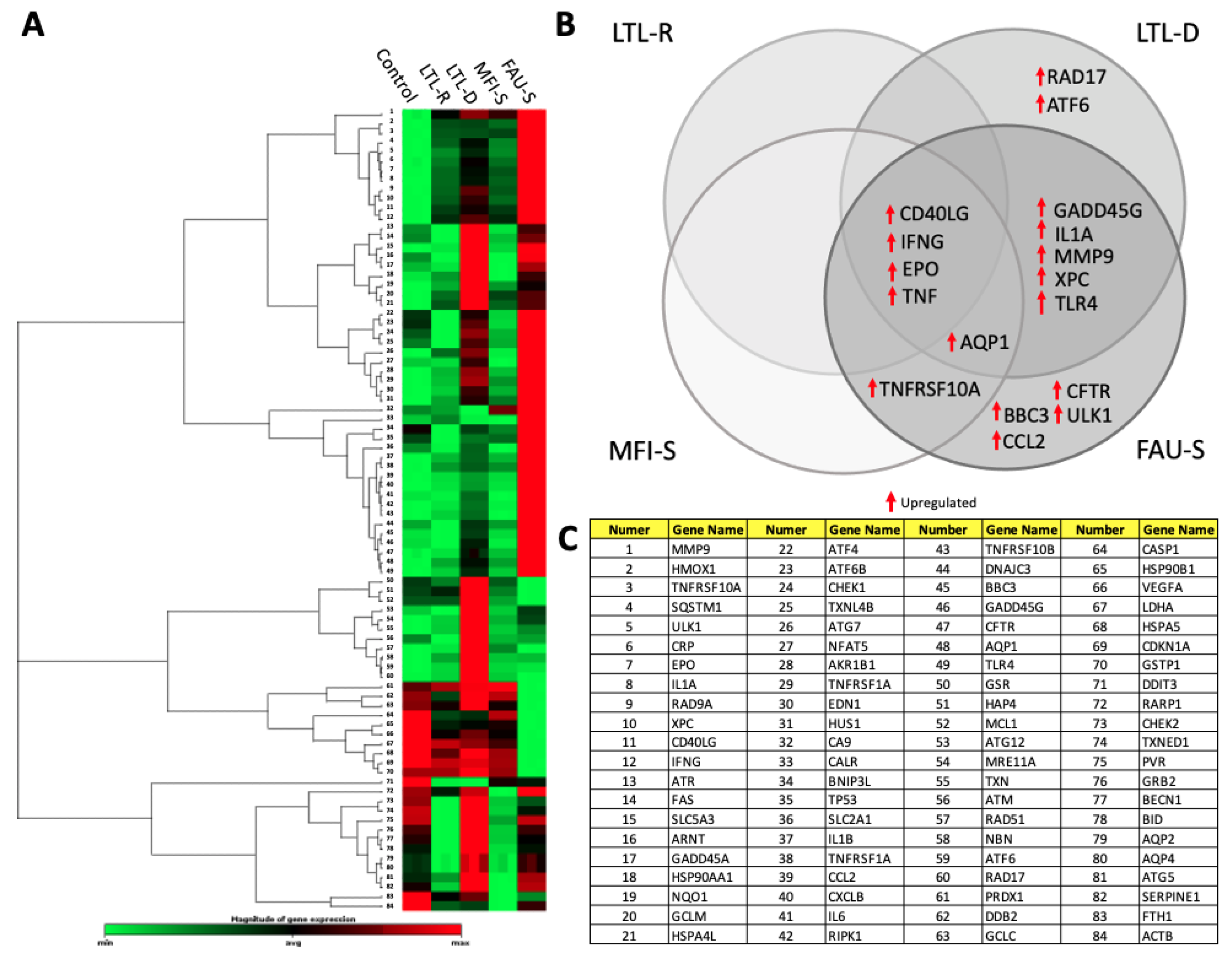
2.8. Toxicity Related Gene Expression
3. Discussion
4. Materials and Methods
4.1. Preparation of Zeolite Particles
4.1.1. Materials
4.1.2. Synthesis of Zeolite L (LTL)
4.1.3. Synthesis of ZSM-5 (MFI-S)
4.1.4. Synthesis of Faujasite (FAU-S)
4.2. Dispersion and Characterization of Materials
4.3. Zeta Potential Measurements
4.4. Cell Cultures
4.5. Real-Time Cell Monitoring
4.6. WST Cell Viability Assay
4.7. Cytotoxicity Assay
4.8. Mitotracker
4.9. Mitochondrial Membrane Potential
4.10. Quantification of Reduced (GSH) and Oxidized Glutathione (GSSG)
4.11. Transmission Electron Microscopy
4.12. RNA Isolation, PCR Arrays, and qRT-PCR Assays
4.13. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Rimer, J.D.; Chawla, A.; Le, T.T. Crystal Engineering for Catalysis. Annu. Rev. Chem. Biomol. Eng. 2018, 9, 283–309. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, M.; Oleksiak, M.D.; Chinta, S.; Rimer, J.D. Controlling crystal polymorphism in organic-free synthesis of Na-zeolites. J. Am. Chem. Soc. 2013, 135, 2641–2652. [Google Scholar] [CrossRef]
- Kihara, T.; Zhang, Y.; Hu, Y.; Mao, Q.; Tang, Y.; Miyake, J. Effect of composition, morphology and size of nanozeolite on its in vitro cytotoxicity. J. Biosci. Bioeng. 2011, 111, 725–730. [Google Scholar] [CrossRef]
- Wang, A.; Pu, K.; Dong, B.; Liu, Y.; Zhang, L.; Zhang, Z.; Duan, W.; Zhu, Y. Role of surface charge and oxidative stress in cytotoxicity and genotoxicity of graphene oxide towards human lung fibroblast cells. J. Appl. Toxicol. 2013, 33, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Laurent, S.; Ng, E.P.; Thirifays, C.; Lakiss, L.; Goupil, G.M.; Mintova, S.; Burtea, C.; Oveisi, E.; Hébert, C.; De Vries, M.; et al. Corona protein composition and cytotoxicity evaluation of ultra-small zeolites synthesized from template free precursor suspensions. Toxicol. Res. 2013, 2, 270–279. [Google Scholar] [CrossRef]
- Li, Z.; Hüve, J.; Krampe, C.; Luppi, G.; Tsotsalas, M.; Klingauf, J.; De Cola, L.; Riehemann, K. Internalization Pathways of Anisotropic Disc-Shaped Zeolite L Nanocrystals with Different Surface Properties in HeLa Cancer Cells. Small 2013, 9, 1809–1820. [Google Scholar] [CrossRef]
- Martin-Kleiner, I.; Flegar-Meštrić, Z.; Zadro, R.; Breljak, D.; Janda, S.S.; Stojković, R.; Marušić, M.; Radačić, M.; Boranić, M. The effect of the zeolite clinoptilolite on serum chemistry and hematopoiesis in mice. Food Chem. Toxicol. 2001, 39, 717–727. [Google Scholar] [CrossRef]
- Gratton, S.E.; Ropp, P.A.; Pohlhaus, P.D.; Luft, J.C.; Madden, V.J.; Napier, M.E.; DeSimone, J.M. The effect of particle design on cellular internalization pathways. Proc. Natl. Acad. Sci. USA 2008, 105, 11613–11618. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Shi, W.; Freund, L.B. Mechanics of receptor-mediated endocytosis. Proc. Natl. Acad. Sci. USA 2005, 102, 9469–9474. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Kim, B.Y.; Rutka, J.T.; Chan, W.C. Nanoparticle-mediated cellular response is size-dependent. Nat. Nanotechnol. 2008, 3, 145–150. [Google Scholar] [CrossRef]
- Bhattacharya, K.; Naha, P.C.; Naydenova, I.; Mintova, S.; Byrne, H.J. Reactive oxygen species mediated DNA damage in human lung alveolar epithelial (A549) cells from exposure to non-cytotoxic MFI-type zeolite nanoparticles. Toxicol. Lett. 2012, 215, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Carbone, M.; Emri, S.; Dogan, A.U.; Steele, I.; Tuncer, M.; Pass, H.I.; Baris, Y.I. A mesothelioma epidemic in Cappadocia: Scientific developments and unexpected social outcomes. Nat. Rev. Cancer 2007, 7, 147–154. [Google Scholar] [CrossRef]
- Lalmanach, G.; Diot, E.; Godat, E.; Lecaille, F.; Hervé-Grépinet, V. Cysteine cathepsins and caspases in silicosis. Biol. Chem. 2006, 387, 863–870. [Google Scholar] [CrossRef]
- Murray, D.K.; Harrison, J.C.; Wallace, W.E. A 13C CP/MAS and 31P NMR study of the interactions of dipalmitoylphosphatidylcholine with respirable silica and kaolin. J. Colloid Interface Sci. 2005, 288, 166–170. [Google Scholar] [CrossRef]
- Rimal, B.; Greenberg, A.K.; Rom, W.N. Basic pathogenetic mechanisms in silicosis: Current understanding. Curr. Opin. Pulm. Med. 2005, 11, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Feigin, D.S. Misconceptions regarding the pathogenicity of silicas and silicates. J. Thorac. Imaging 1989, 4, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Rohl, A.; Langer, A.; Moncure, G.; Selikoff, I.; Fischbein, A. Endemic pleural disease associated with exposure to mixed fibrous dust in Turkey. Science 1982, 216, 518–520. [Google Scholar] [CrossRef] [PubMed]
- Brody, A.R.; Hill, L. Interstitial accumulation of inhaled chrysotile asbestos fibers and consequent formation of microcalcifications. Am. J. Pathol. 1982, 109, 107–114. [Google Scholar]
- Bagnato, G.; Harari, S. Cellular interactions in the pathogenesis of interstitial lung diseases. Eur. Respir. Rev. 2015, 24, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Choi, H.H.; Park, G.; Kim, I.W.; Kim, T.J. Fibrogenic effects of crocidolite, amosite, and chrysotile asbestos fibers on lung fibroblasts. Toxicol. Environ. Chem. 2019, 101, 148–164. [Google Scholar] [CrossRef]
- Yu, S.; Choi, H.H.; Kim, I.W.; Kim, T.J. Conditioned medium from asbestos-exposed fibroblasts affects proliferation and invasion of lung cancer cell lines. PLoS ONE 2019, 14, e0222160. [Google Scholar] [CrossRef]
- Yang, H.; Bocchetta, M.; Kroczynska, B.; Elmishad, A.G.; Chen, Y.; Liu, Z.; Bubici, C.; Mossman, B.T.; Pass, H.I.; Testa, J.R.; et al. TNF-α inhibits asbestos-induced cytotoxicity via a NF-κB-dependent pathway, a possible mechanism for asbestos-induced oncogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 10397–10402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olafson, K.N.; Li, R.; Alamani, B.G.; Rimer, J.D. Engineering crystal modifiers: Bridging classical and nonclassical crystallization. Chem. Mater. 2016, 28, 8453–8465. [Google Scholar] [CrossRef]
- Lupulescu, A.I.; Kumar, M.; Rimer, J.D. A facile strategy to design zeolite L crystals with tunable morphology and surface architecture. J. Am. Chem. Soc. 2013, 135, 6608–6617. [Google Scholar] [CrossRef]
- Gies, H.; Van Koningsveld, H. Catalog of Disorder in Zeolite Frameworks. 2002. Available online: http://www.iza-structure.org/databases/ (accessed on 20 May 2018).
- Malhotra, N.; Lee, J.S.; Liman, R.A.D.; Ruallo, J.M.S.; Villaflores, O.B.; Ger, T.R.; Hsiao, C.D. Potential toxicity of iron oxide magnetic nanoparticles: A review. Molecules 2020, 25, 3159. [Google Scholar] [CrossRef] [PubMed]
- Lehman, S.E.; Larsen, S.C. Zeolite and mesoporous silica nanomaterials: Greener syntheses, environmental applications and biological toxicity. Environ. Sci. Nano 2014, 1, 200–213. [Google Scholar] [CrossRef]
- Larlus, O.; Valtchev, V.P. Crystal morphology control of LTL-type zeolite crystals. Chem. Mater. 2004, 16, 3381–3389. [Google Scholar] [CrossRef]
- Baerlocher, C.; McCusker, L.B.; Olson, D.H. Atlas of Zeolite Framework Types; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Díaz, I.; Kokkoli, E.; Terasaki, O.; Tsapatsis, M. Surface structure of zeolite (MFI) crystals. Chem. Mater. 2004, 16, 5226–5232. [Google Scholar] [CrossRef]
- Męczyńska-Wielgosz, S.; Piotrowska, A.; Majkowska-Pilip, A.; Bilewicz, A.; Kruszewski, M. Effect of surface functionalization on the cellular uptake and toxicity of nanozeolite A. Nanoscale Res. Lett. 2016, 11, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albanese, A.; Tang, P.S.; Chan, W.C. The effect of nanoparticle size, shape, and surface chemistry on biological systems. Annu. Rev. Biomed. Eng. 2012, 14, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Calatayud, M.P.; Sanz, B.; Raffa, V.; Riggio, C.; Ibarra, M.R.; Goya, G.F. The effect of surface charge of functionalized Fe3O4 nanoparticles on protein adsorption and cell uptake. Biomaterials 2014, 35, 6389–6399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saptarshi, S.R.; Duschl, A.; Lopata, A.L. Interaction of nanoparticles with proteins: Relation to bio-reactivity of the nanoparticle. J. Nanobiotechnol. 2013, 11, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Saikia, J.; Yazdimamaghani, M.; Hadipour Moghaddam, S.P.; Ghandehari, H. Differential protein adsorption and cellular uptake of silica nanoparticles based on size and porosity. ACS Appl. Mater. Interfaces 2016, 8, 34820–34832. [Google Scholar] [CrossRef] [Green Version]
- Moore, T.L.; Rodriguez-Lorenzo, L.; Hirsch, V.; Balog, S.; Urban, D.; Jud, C.; Rothen-Rutishauser, B.; Lattuada, M.; Petri-Fink, A. Nanoparticle colloidal stability in cell culture media and impact on cellular interactions. Chem. Soc. Rev. 2015, 44, 6287–6305. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Neuss, S.; Leifert, A.; Fischler, M.; Wen, F.; Simon, U.; Schmid, G.; Brandau, W.; Jahnen-Dechent, W. Size-dependent cytotoxicity of gold nanoparticles. Small 2007, 3, 1941–1949. [Google Scholar] [CrossRef]
- Sukhanova, A.; Bozrova, S.; Sokolov, P.; Berestovoy, M.; Karaulov, A.; Nabiev, I. Dependence of nanoparticle toxicity on their physical and chemical properties. Nanoscale Res. Lett. 2018, 13, 1–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Li, M.; Ji, J.; Gao, F.; Bai, R.; Chen, C.; Wang, Z.; Zhang, C.; Niu, Q. In vivo toxicity of nano-alumina on mice neurobehavioral profiles and the potential mechanisms. Int. J. Immunopathol. Pharmacol. 2011, 24, 23S–29S. [Google Scholar] [PubMed]
- Troulinaki, K.; Tavernarakis, N. Endocytosis and intracellular trafficking contribute to necrotic neurodegeneration in C. elegans. Embo J. 2012, 31, 654–666. [Google Scholar] [CrossRef] [Green Version]
- AshaRani, P.; Low Kah Mun, G.; Hande, M.P.; Valiyaveettil, S. Cytotoxicity and genotoxicity of silver nanoparticles in human cells. ACS Nano 2009, 3, 279–290. [Google Scholar] [CrossRef]
- Huang, X.; Teng, X.; Chen, D.; Tang, F.; He, J. The effect of the shape of mesoporous silica nanoparticles on cellular uptake and cell function. Biomaterials 2010, 31, 438–448. [Google Scholar] [CrossRef]
- Khan, I.; Saeed, K.; Khan, I. Nanoparticles: Properties, applications and toxicities. Arab. J. Chem. 2019, 12, 908–931. [Google Scholar] [CrossRef]
- Das, R.; Ghosh, S.; Naskar, M.K. Synthesis of single crystal zeolite L rods with high aspect ratio using rice husk ash as silica source. Indian J. Chem. Sect. Inorg. Bio-Inorg. Phys. Theor. Anal. Chem. 2014, 53, 816–819. [Google Scholar]
- Trakarnroek, S.; Jongpatiwut, S.; Rirksomboon, T.; Osuwan, S.; Resasco, D.E. n-Octane aromatization over Pt/KL of varying morphology and channel lengths. Appl. Catal. A Gen. 2006, 313, 189–199. [Google Scholar] [CrossRef]
- Jentoft, R.; Tsapatsis, M.; Davis, M.; Gates, B. Platinum clusters supported in zeolite LTL: Influence of catalyst morphology on performance inn-hexane reforming. J. Catal. 1998, 179, 565–580. [Google Scholar] [CrossRef]
- Rivero, L.G.; Bañuelos, J.; Bizkarra, K.; Izquierdo, U.; Barrio, V.L.; Cambra, J.F.; Arbeloa, I.L. Linde Type L Zeolite: A Privileged Porous Support to Develop Photoactive and Catalytic Nanomaterials. In Zeolites and Their Applications; IntechOpen: London, UK, 2018; p. 13. [Google Scholar]
- Saravanan, V.; Waijers, D.; Ziari, M.; Noordermeer, M. Recovery of 1-butanol from aqueous solutions using zeolite ZSM-5 with a high Si/Al ratio; suitability of a column process for industrial applications. Biochem. Eng. J. 2010, 49, 33–39. [Google Scholar] [CrossRef]
- Shen, C.; Ma, J.; Liu, W.; Wen, Y.; Rashid, S. Selective conversion of organic pollutant p-chlorophenol to formic acid using zeolite Fenton catalyst. Chemosphere 2016, 161, 446–453. [Google Scholar] [CrossRef]
- Shevade, S.; Ford, R.G. Use of synthetic zeolites for arsenate removal from pollutant water. Water Res. 2004, 38, 3197–3204. [Google Scholar] [CrossRef] [PubMed]
- Seigner, J.; Basilio, J.; Resch, U.; de Martin, R. CD40L and TNF both activate the classical NF-κB pathway, which is not required for the CD40L induced alternative pathway in endothelial cells. Biochem. Biophys. Res. Commun. 2018, 495, 1389–1394. [Google Scholar] [CrossRef]
- Ke, N.; Xi, B.; Ye, P.; Xu, W.; Zheng, M.; Mao, L.; Wu, M.J.; Zhu, J.; Wu, J.; Zhang, W.; et al. Screening and identification of small molecule compounds perturbing mitosis using time-dependent cellular response profiles. Anal. Chem. 2010, 82, 6495–6503. [Google Scholar] [CrossRef]
- Al-Hallak, K.M.; Azarmi, S.; Anwar-Mohamed, A.; Roa, W.H.; Löbenberg, R. Secondary cytotoxicity mediated by alveolar macrophages: A contribution to the total efficacy of nanoparticles in lung cancer therapy? Eur. J. Pharm. Biopharm. 2010, 76, 112–119. [Google Scholar] [CrossRef]
- Organisation for Economic Co-operation and Development. Crystaline, non-fibrous zeolites. In Screening Information Dataset Initial Assessment Report for SIAM 23; OECD: Paris, France, 2006; pp. 17–20. [Google Scholar]
- Dalbey, W.; Pulkowski, C. Comparison of synthetic zeolite catalysts and alumina binders administered intratracheally to rats. J. Toxicol. Environ. Health Part A 2000, 60, 355–374. [Google Scholar]
- Gloxhuber, C.; Potokar, M.; Pittermann, W.; Wallat, S.; Bartnik, F.; Reuter, H.; Braig, S. Zeolithe A—A phosphate substitute for detergents: Toxicological investigation. Food Chem. Toxicol. 1983, 21, 209–220. [Google Scholar] [CrossRef]
- Hudson, S.P.; Padera, R.F.; Langer, R.; Kohane, D.S. The biocompatibility of mesoporous silicates. Biomaterials 2008, 29, 4045–4055. [Google Scholar] [CrossRef] [Green Version]
- Ghorbanpour, A.; Gumidyala, A.; Grabow, L.C.; Crossley, S.P.; Rimer, J.D. Epitaxial growth of ZSM-5@ Silicalite-1: A core–shell zeolite designed with passivated surface acidity. ACS Nano 2015, 9, 4006–4016. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, J.; DeGraff, W.G.; Gazdar, A.F.; Minna, J.D.; Mitchell, J.B. Evaluation of a tetrazolium-based semiautomated colorimetric assay: Assessment of chemosensitivity testing. Cancer Res. 1987, 47, 936–942. [Google Scholar] [PubMed]
- Hussain, S.M.; Frazier, J.M. Cellular toxicity of hydrazine in primary rat hepatocytes. Toxicol. Sci. 2002, 69, 424–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, E.Y.; Smith, M.T.; Bellomo, G.; Di Monte, D. Relationships between the mitochondrial transmembrane potential, ATP concentration, and cytotoxicity in isolated rat hepatocytes. Arch. Biochem. Biophys. 1990, 282, 358–362. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LTL-R | LTL-D | MFI-S | FAU-S | |
|---|---|---|---|---|
| Si/Al | 3 | 3 | 50 | 1.5 |
| Zeta potential (mV) | ||||
| DI water | −52 ± 1 | −68 ± 4 | −39.3 ± 0.8 | −43.4 ± 0.5 |
| DPBS | −28.9 ± 0.2 | −22.6 ± 0.8 | −29 ± 1 | −28.4 ± 0.2 |
| EMEM-N | −28 ± 2 | −14.1 ± 0.4 | −27 ± 1 | −21 ± 1 |
| EMEM-F e | −10.5 ± 0.4 | −9.3 ± 0.5 | −10.3 ± 0.2 | −11.8 ± 0.6 |
| Zeolite | LDH EC, μg/mL | MTT EC, μg/mL |
|---|---|---|
| LTL-R | 148.8 ± 22.3 | 161.0 ± 18.6 |
| LTL-D | 214.9 ± 55.6 | 225.5 ± 28.3 |
| MFI-S | 144.5 ± 24.2 | 148.7 ± 17.7 |
| FAU-S | 57.0 ± 13.2 | 57.8 ± 10.6 |
| TNF Related Genes Mean Fold Change ± SD | LTL-R | LTL-D | MFI-S | FAU-S |
|---|---|---|---|---|
| CD40LG | 1.12 ± 0.23 | 1.19 ± 0.32 | 1.56 ± 0.14 | 6.33 ± 0.55 |
| p = 0.4679 | p = 0.3901 | p = 0.006 | p < 0.001 | |
| TNF-alpha | 0.97 ± 0.33 | 1.42 ± 0.34 | 1.83 ± 0.32 | 4.32 ± 0.33 |
| p = 0.888 | p = 0.075 | p = 0.012 | p < 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, S.-H.; Kumar, M.; Kim, I.W.; Rimer, J.D.; Kim, T.-J. A Comparative Analysis of In Vitro Toxicity of Synthetic Zeolites on IMR-90 Human Lung Fibroblast Cells. Molecules 2021, 26, 3194. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26113194
Yu S-H, Kumar M, Kim IW, Rimer JD, Kim T-J. A Comparative Analysis of In Vitro Toxicity of Synthetic Zeolites on IMR-90 Human Lung Fibroblast Cells. Molecules. 2021; 26(11):3194. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26113194
Chicago/Turabian StyleYu, Seung-Hye, Manjesh Kumar, Il Won Kim, Jeffrey D. Rimer, and Tae-Jung Kim. 2021. "A Comparative Analysis of In Vitro Toxicity of Synthetic Zeolites on IMR-90 Human Lung Fibroblast Cells" Molecules 26, no. 11: 3194. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26113194