
Antioxidant Fusion Protein SOD1-Tat Increases the Engraftment Efficiency of Total Bone Marrow Cells in Irradiated Mice

 ,
,
Abstract
:1. Introduction
2. Results
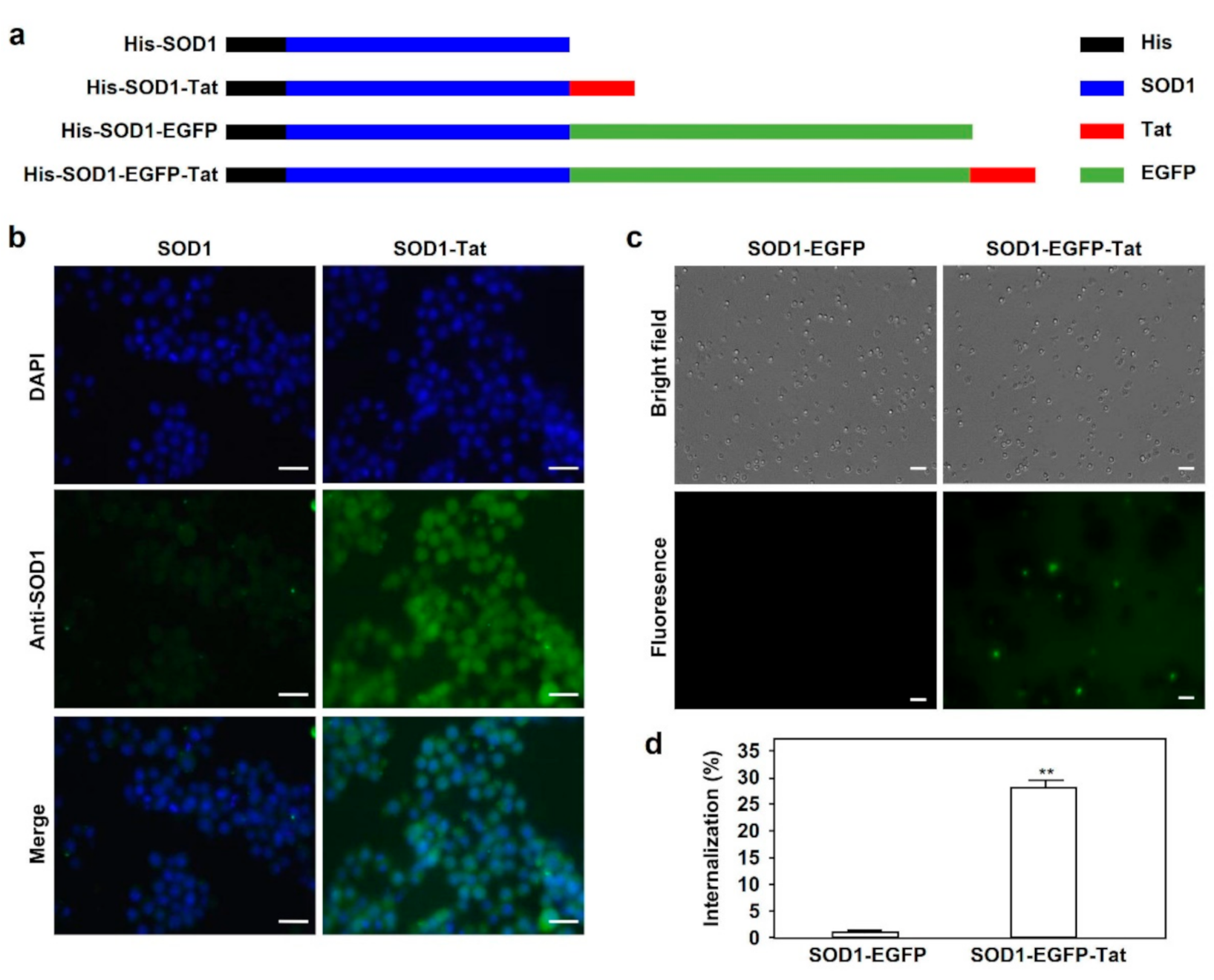
2.1. SOD1-Tat Shows Penetrating Ability in RAW264.7 Cells and Bone Marrow Cells
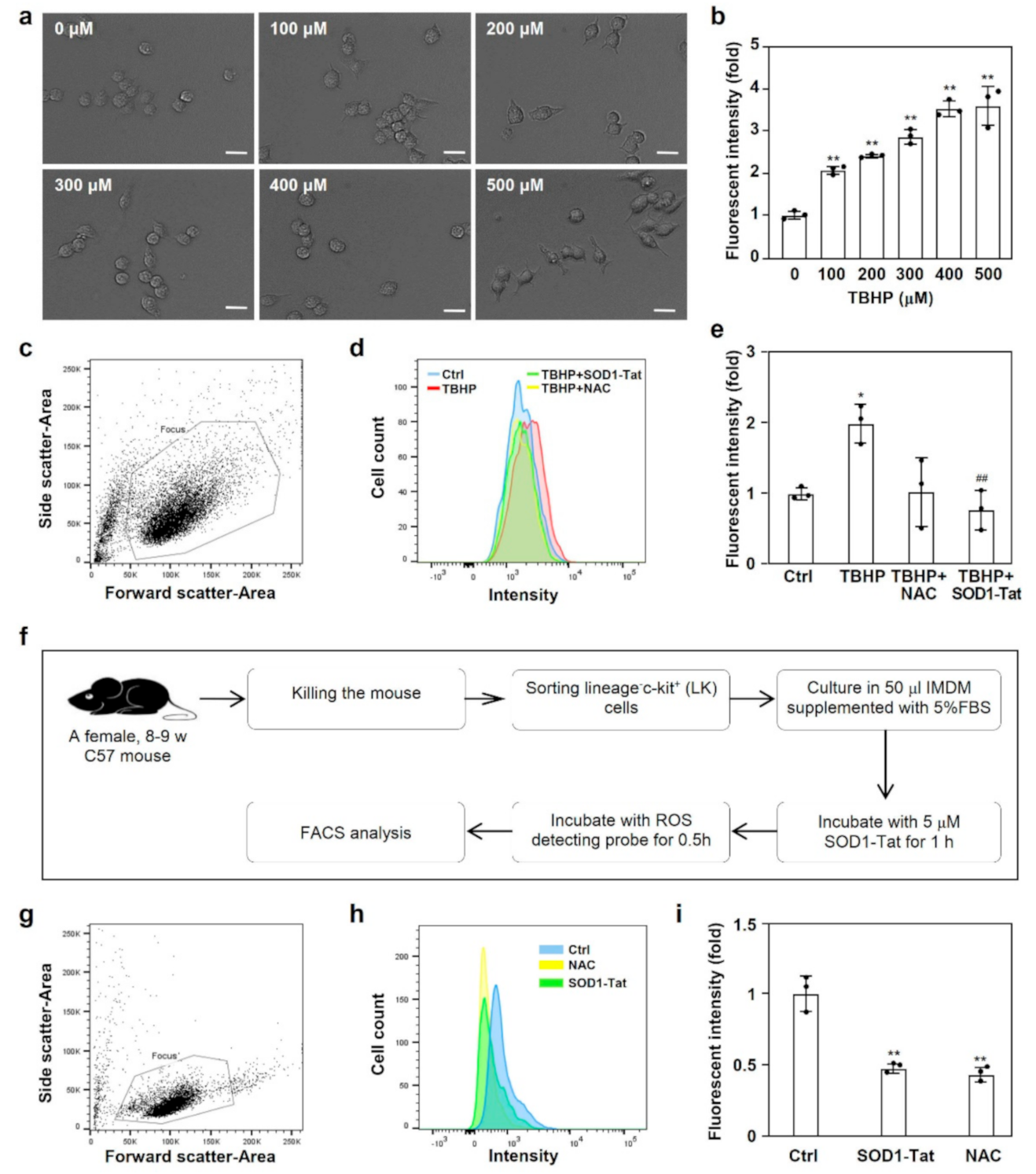
2.2. SOD1-Tat Reduces ROS in RAW264.7 Cells and Lineage-c-Kit+ (LK) Cells
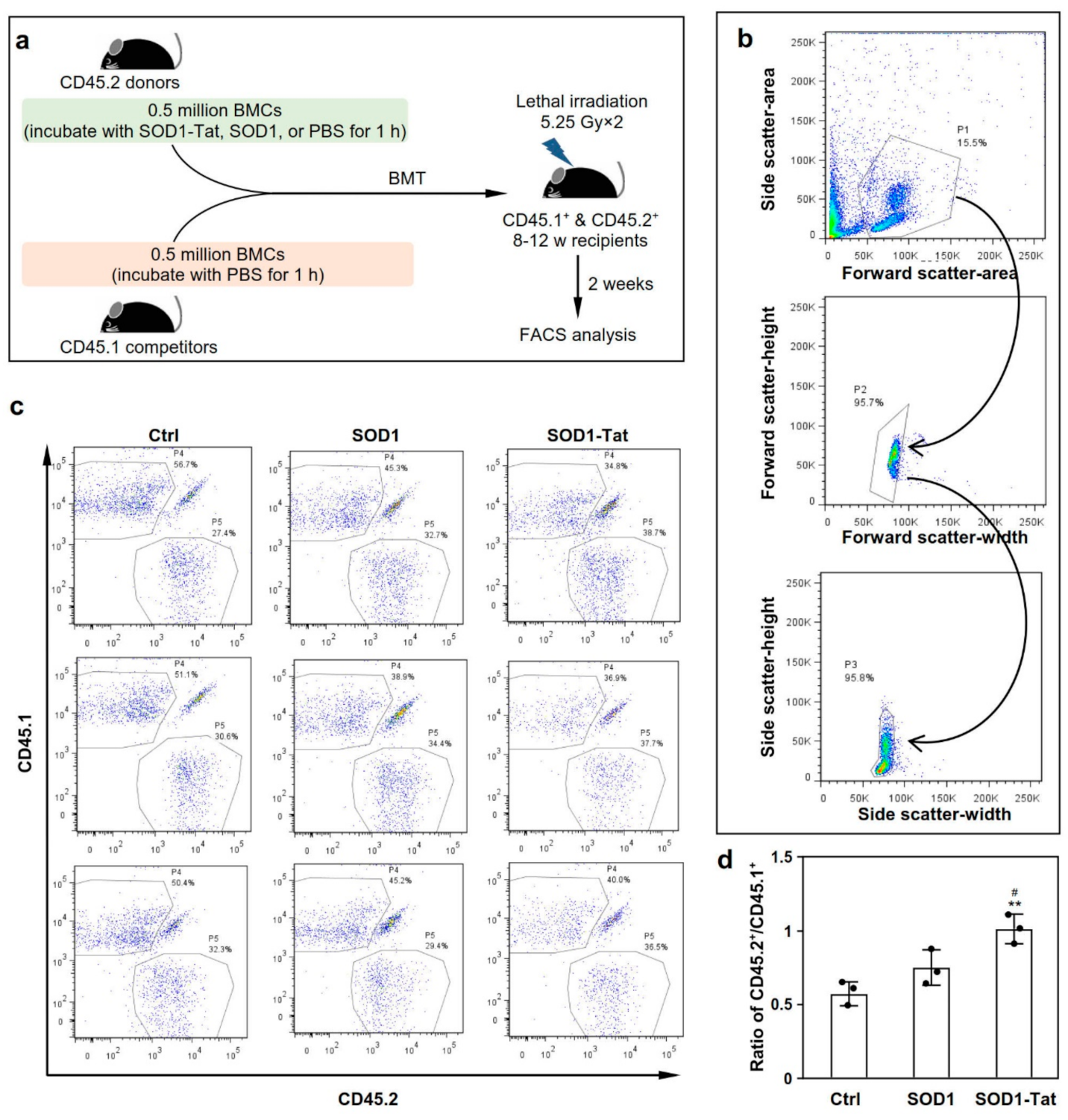
2.3. SOD1-Tat Increases the Engraftment Ability of Total Bone Marrow Cells in Irradiated Mice
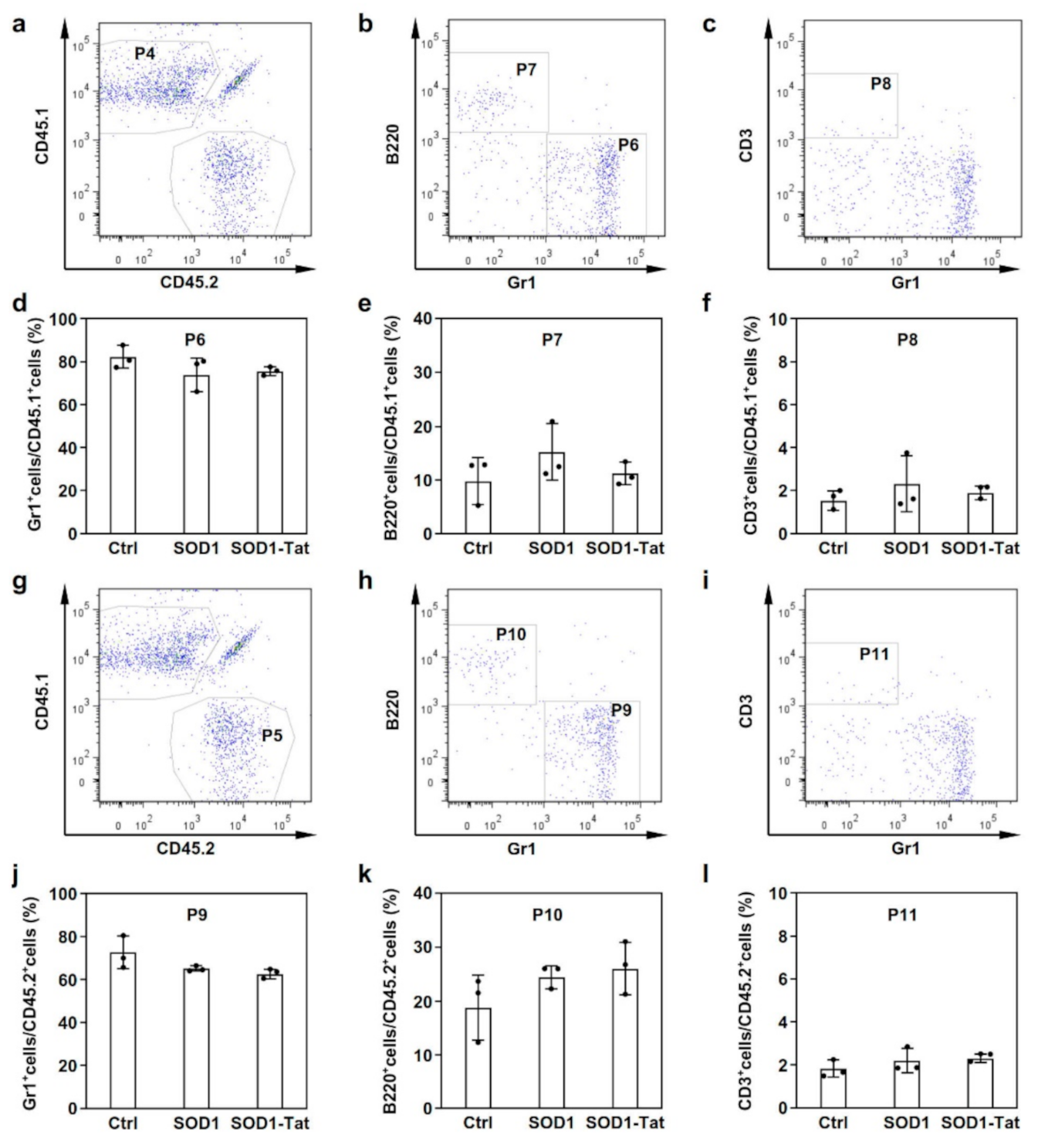
2.4. SOD1-Tat Has No Significant Effect on Myeloid Cell Differentiation in Mice With BMT
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Construction of Expression Vectors
4.3. Preparation of Recombinant Fusion Proteins
4.4. Internalization and Imaging
4.5. Detection of Hydrogen Peroxide
4.6. LK Cell Sorting
4.7. Competitive Transplantation
4.8. FACS Analysis for Hematopoietic Chimerism
4.9. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Shen, H.; Yu, H.; Liang, P.H.; Cheng, H.; XuFeng, R.; Yuan, Y.; Zhang, P.; Smith, C.A.; Cheng, T. An acute negative bystander effect of gamma-irradiated recipients on transplanted hematopoietic stem cells. Blood 2012, 119, 3629–3637. [Google Scholar] [CrossRef]
- Winston, D.J.; Gale, R.P.; Meyer, D.V.; Young, L.S. Infectious Complications of Human Bone Marrow Transplantation. Medicine 1979, 58, 1–31. [Google Scholar] [CrossRef]
- Kusne, S.; Krystofiak, S. Infection control issues after bone marrow transplantation. Curr. Opin. Infect. Dis. 2001, 14, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Cheng, H.; Gao, Y.; Shi, M.; Liu, Y.; Hu, Z.; Xu, J.; Qiu, L.; Yuan, W.; Leung, A.Y.-H.; et al. Antioxidant N-acetyl-l-cysteine increases engraftment of human hematopoietic stem cells in immune-deficient mice. Blood 2014, 124, e45–e48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, W.; XuFeng, R.; Park, M.-R.; Gu, H.; Hu, L.; Kang, J.W.; Ma, S.; Liang, P.H.; Li, Y.; Cheng, H.; et al. Hematopoietic Stem Cell Regeneration Enhanced by Ectopic Expression of ROS-detoxifying Enzymes in Transplant Mice. Mol. Ther. 2013, 21, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Fonseca, S.B.; Pereira, M.P.; Kelley, S.O. Recent advances in the use of cell-penetrating peptides for medical and biological applications. Adv. Drug Deliv. Rev. 2009, 61, 953–964. [Google Scholar] [CrossRef]
- Green, M.; Loewenstein, P.M. Autonomous functional domains of chemically synthesized human immunodeficiency virus tat trans-activator protein. Cell 1988, 55, 1179–1188. [Google Scholar] [CrossRef]
- Vivès, E.; Brodin, P.; Lebleu, B. A Truncated HIV-1 Tat Protein Basic Domain Rapidly Translocates through the Plasma Membrane and Accumulates in the Cell Nucleus. J. Biol. Chem. 1997, 272, 16010–16017. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, M.A. Peptide-mediated cell delivery: Application in protein target validation. Curr. Opin. Pharmacol. 2002, 2, 587–594. [Google Scholar] [CrossRef]
- Wadia, J.S.; Dowdy, S.F. Modulation of Cellular Function by TAT Mediated Transduction of Full Length Proteins. Curr. Protein Pept. Sci. 2003, 4, 97–104. [Google Scholar] [CrossRef]
- Lewin, M.; Carlesso, N.; Tung, C.-H.; Tang, X.-W.; Cory, D.G.; Scadden, D.T.; Weissleder, R. Tat peptide-derivatized magnetic nanoparticles allow in vivo tracking and recovery of progenitor cells. Nat. Biotechnol. 2000, 18, 410–414. [Google Scholar] [CrossRef]
- Song, Y.; Su, M.; Zhu, J.; Di, W.; Liu, Y.; Hu, R.; Rood, D.; Lai, L. FOXN1 recombinant protein enhances T-cell regeneration after hematopoietic stem cell transplantation in mice. Eur. J. Immunol. 2016, 46, 1518–1528. [Google Scholar] [CrossRef] [PubMed]
- Fridovich, I. Superoxide Radical and Superoxide Dismutases. Stress Health Soc. Environ. 1981, 64, 250–272. [Google Scholar] [CrossRef]
- Sheng, Y.; Abreu, I.A.; Cabelli, D.E.; Maroney, M.J.; Miller, A.-F.; Teixeira, M.; Valentine, J.S. Superoxide Dismutases and Superoxide Reductases. Chem. Rev. 2014, 114, 3854–3918. [Google Scholar] [CrossRef]
- Nilsson, S.K.; Dooner, M.S.; Weier, H.U.; Frenkel, B.; Lian, J.B.; Stein, G.S.; Quesenberry, P.J. Cells capable of bone production engraft from whole bone marrow transplants in nonablated mice. J. Exp. Med. 1999, 189, 729–734. [Google Scholar] [CrossRef]
- Madani, F.; Lindberg, S.; Langel, Ü.; Futaki, S.; Gräslund, A. Mechanisms of Cellular Uptake of Cell-Penetrating Peptides. J. Biophys. 2011, 2011, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tünnemann, G.; Martin, R.M.; Haupt, S.; Patsch, C.; Edenhofer, F.; Cardoso, M.C. Cargo-dependent mode of uptake and bioavailability of TAT-containing proteins and peptides in living cells. FASEB J. 2006, 20, 1775–1784. [Google Scholar] [CrossRef] [Green Version]
- Kwon, H.Y.; Eum, W.S.; Jang, H.W.; Kang, J.H.; Ryu, J.; Lee, B.R.; Jin, L.H.; Park, J.; Choi, S.Y. Transduction of Cu,Zn-superoxide dismutase mediated by an HIV-1 Tat protein basic domain into mammalian cells. FEBS Lett. 2000, 485, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Luangwattananun, P.; Eiamphungporn, W.; Songtawee, N.; Bulow, L.; Ayudhya, C.I.N.; Prachayasittikul, V.; Yainoy, S. Improving enzymatic activities and thermostability of a tri-functional enzyme with SOD, catalase and cell-permeable activities. J. Biotechnol. 2017, 247, 50–59. [Google Scholar] [CrossRef]
- Min, D.; Kim, H.; Park, L.; Kim, T.H.; Hwang, S.; Kim, M.J.; Jang, S.; Park, Y. Amelioration of diabetic neuropathy by TAT-mediated enhanced delivery of metallothionein and SOD. Endocrinology 2012, 153, 81–91. [Google Scholar] [CrossRef]
- Zhang, X.; Karatepe, K.; Chiewchengchol, D.; Zhu, H.; Guo, R.; Liu, P.; Yu, H.; Ren, Q.; Luo, X.; Cheng, T.; et al. Bacteria-Induced Acute Inflammation Does Not Reduce the Long-Term Reconstitution Capacity of Bone Marrow Hematopoietic Stem Cells. Front. Immunol. 2020, 11, 626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karatepe, K.; Zhu, H.; Zhang, X.; Guo, R.; Kambara, H.; Loison, F.; Liu, P.; Yu, H.; Ren, Q.; Luo, X.; et al. Proteinase 3 Limits the Number of Hematopoietic Stem and Progenitor Cells in Murine Bone Marrow. Stem Cell Rep. 2018, 11, 1092–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vector Name | Primer Name | Primer Sequence | Enzyme Site |
|---|---|---|---|
| pET14b | SOD1 | Forward: 5′ ATAGATACATATGATGGCGATGAAAGCGGTGTGCG 3′ | Nde I |
| Reverse: 5′ TAGATGGTACCCTGCGCAATCCCAATCACTCCAC 3′ | Kpn I | ||
| pET14b | EGFP | Forward: 5′ ATTAGGTACCGTGAGCAAGGGCGAGGAGCTGTTC 3′ | Kpn I |
| Reverse: 5′ ATATCTCGAGCTTGTACAGCTCGTCCATGCCGAGA 3′ | Xho I | ||
| pET14b | Tat | Forward: 5′ GATCCTATGGCCGCAAGAAACGCCGTCAGCGTCGCCGTG 3′ | BamH I |
| Reverse: 5′ GATCCACGGCGACGCTGACGGCGTTTCTTGCGGCCATAG 3′ | BamH I |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bei, T.; Cao, X.; Liu, Y.; Li, J.; Luo, H.; Huang, L.; Tian, T.; Li, L.; Jiang, Y. Antioxidant Fusion Protein SOD1-Tat Increases the Engraftment Efficiency of Total Bone Marrow Cells in Irradiated Mice. Molecules 2021, 26, 3395. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26113395
Bei T, Cao X, Liu Y, Li J, Luo H, Huang L, Tian T, Li L, Jiang Y. Antioxidant Fusion Protein SOD1-Tat Increases the Engraftment Efficiency of Total Bone Marrow Cells in Irradiated Mice. Molecules. 2021; 26(11):3395. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26113395
Chicago/Turabian StyleBei, Ting, Xusong Cao, Yun Liu, Jinmei Li, Haihua Luo, Lin Huang, Tian Tian, Lei Li, and Yong Jiang. 2021. "Antioxidant Fusion Protein SOD1-Tat Increases the Engraftment Efficiency of Total Bone Marrow Cells in Irradiated Mice" Molecules 26, no. 11: 3395. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26113395