
Phytochemical Characterization of Olea europaea L. Cultivars of Cilento National Park (South Italy) through NMR-Based Metabolomics

 ,
,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
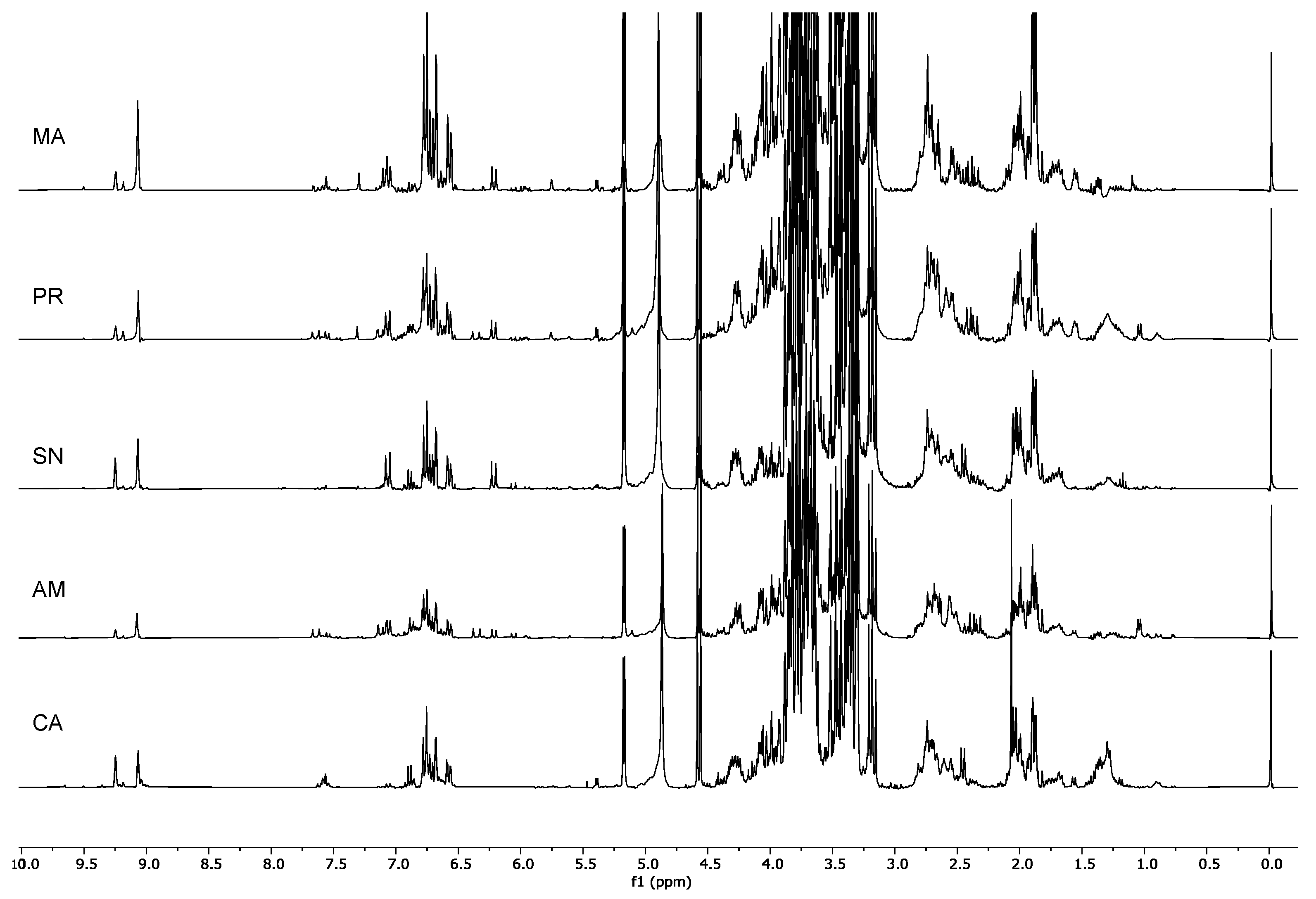
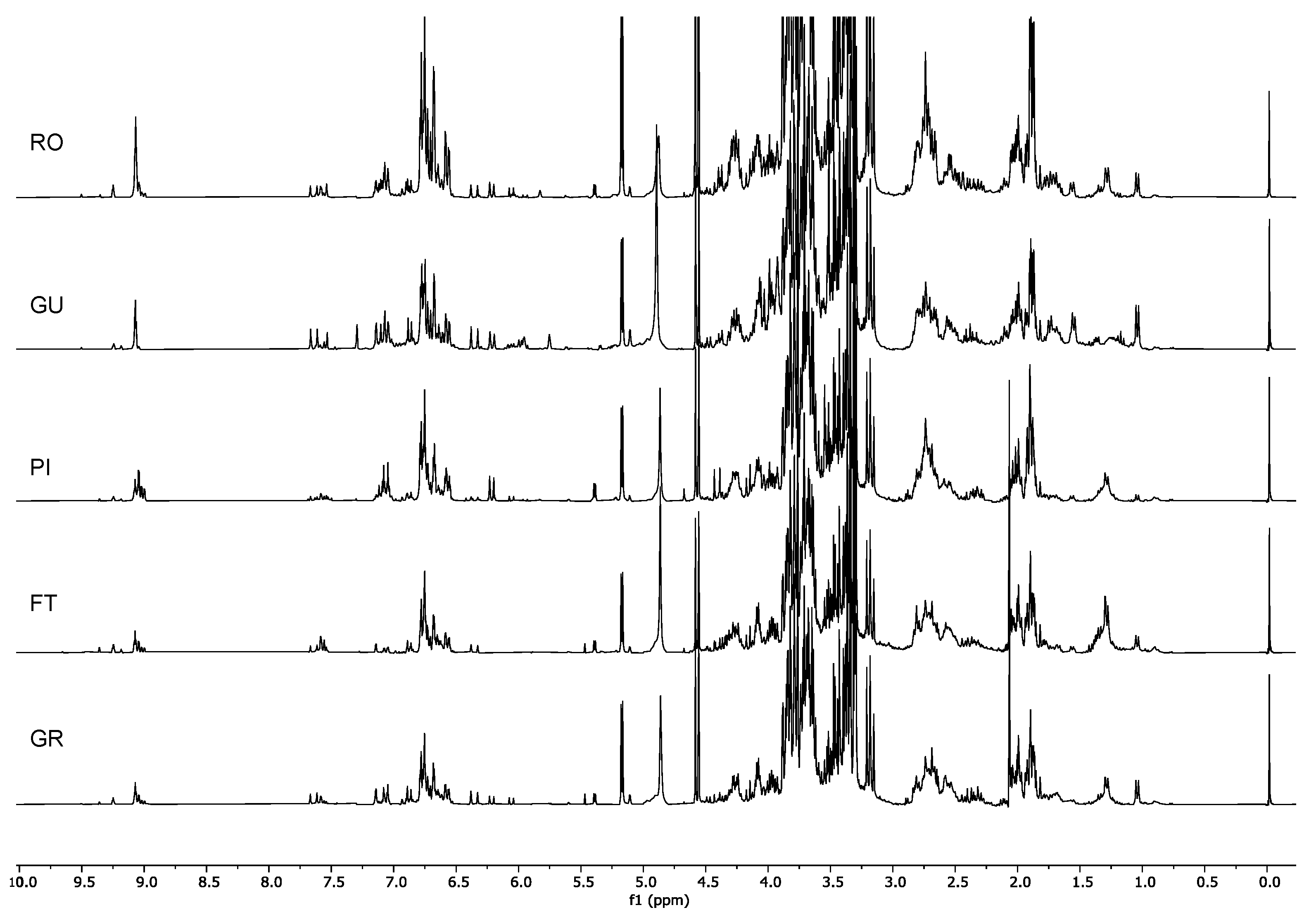
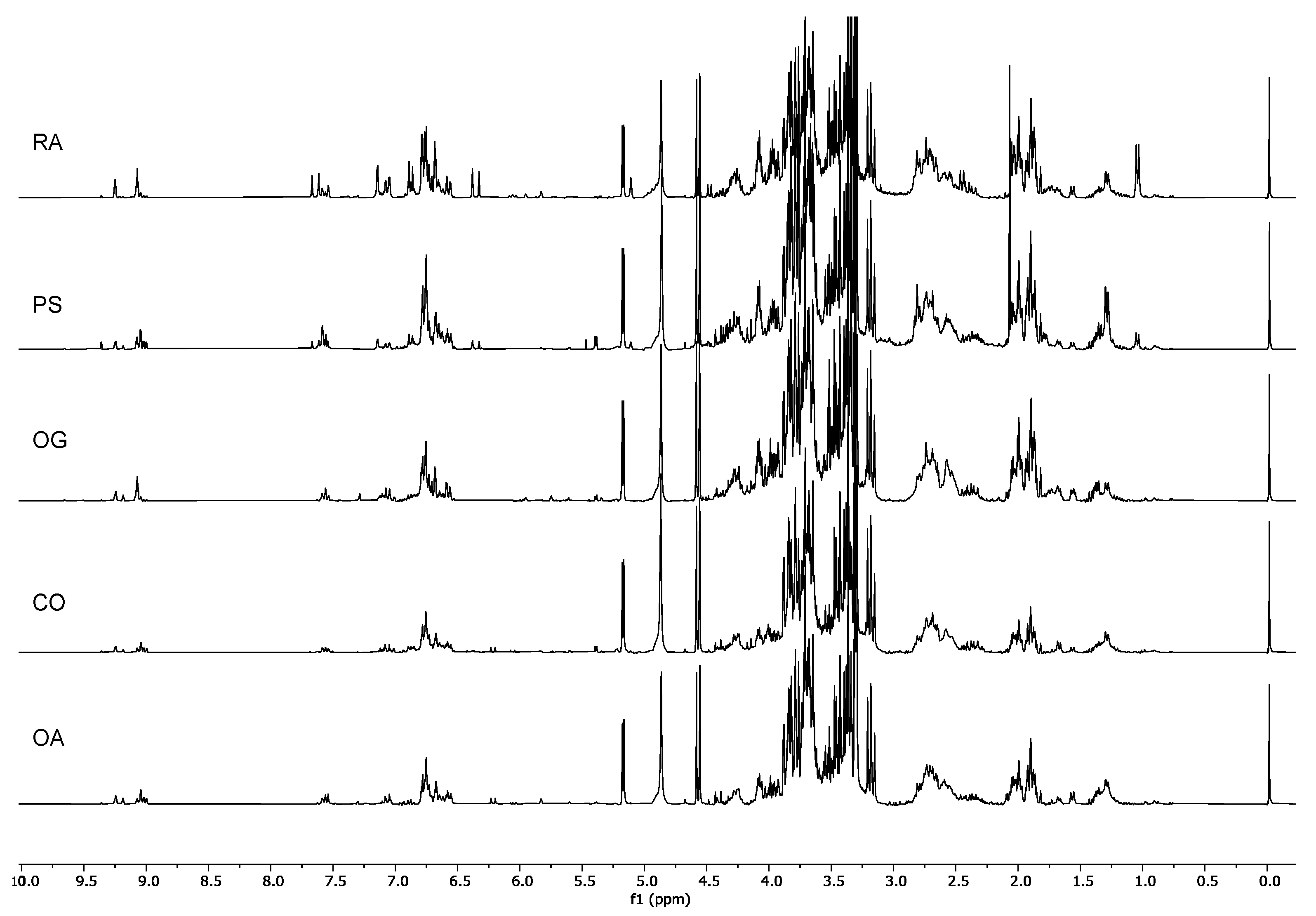
2.1. H-NMR Analysis
2.1.1. Multivariate Data Analysis: PCA on NMR Data
2.1.2. Identification of the Metabolites in the Extracts
2.2. Classifications of the Analyzed Cultivars Based on Their Metabolite Content
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Metabolomics Analysis
4.2.1. Extraction
4.2.2. NMR Analysis
4.2.3. Multivariate Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Bartolini, G.; Prevost, G.; Messeri, C.; Carignani, G.; Menini, U. Olive Germplasm: Cultivars and World-Wide Collections; FAO: Rome, Italy, 1998. [Google Scholar]
- Di Donna, L.; Mazzotti, F.; Naccarato, A.; Salerno, R.; Tagarelli, A.; Taverna, D.; Sindona, G. Secondary metabolites of Olea europaea leaves as markers for the discrimination of cultivars and cultivation zones by multivariate analysis. Food Chem. 2010, 121, 492–496. [Google Scholar] [CrossRef]
- Scognamiglio, M.; D’Abrosca, B.; Pacifico, S.; Fiumano, V.; De Luca, P.F.; Monaco, P.; Fiorentino, A. Polyphenol characterization and antioxidant evaluation of Olea europaea varieties cultivated in Cilento National Park (Italy). Food Res. Int. 2012, 46, 294–303. [Google Scholar] [CrossRef]
- Ben Mohamed, M.; Guasmi, F.; Ben Ali, S.; Radhouani, F.; Faghim, J.; Triki, T.; Kammoun, N.G.; Baffi, C.; Lucini, L.; Benincasa, C. The LC-MS/MS characterization of phenolic compounds in leaves allows classifying olive cultivars grown in South Tunisia. Biochem. Syst. Ecol. 2018, 78, 84–90. [Google Scholar] [CrossRef]
- Ganino, T.; Bartolini, G.; Fabbri, A. The classification of olive germplasm. J. Hortic. Sci. Biotechnol. 2006, 81, 319–334. [Google Scholar] [CrossRef]
- Muzzalupo, I.; Vendramin, G.G.; Chiappetta, A. Genetic biodiversity of Italian olives (Olea europaea) germplasm analyzed by SSR markers. Sci. World J. 2014, 2014, 296590. [Google Scholar] [CrossRef] [Green Version]
- Muzzalupo, I.; Chiappetta, A.; Benincasa, C.; Perri, E. Intra-cultivar variability of three major olive cultivars grown in different areas of central-southern Italy and studied using microsatellite markers. Sci. Hortic. 2010, 126, 324–329. [Google Scholar] [CrossRef]
- Olmo-García, L.; Kessler, N.; Neuweger, H.; Wendt, K.; Olmo-Peinado, J.M.; Fernández-Gutiérrez, A.; Baessmann, C.; Carrasco-Pancorbo, A. Unravelling the distribution of secondary metabolites in Olea europaea L.: Exhaustive characterization of eight olive-tree derived matrices by complementary platforms (LC-ESI/APCI-MS and GC-APCI-MS). Molecules 2018, 23, 2419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Termentzi, A.; Halabalaki, M.; Skaltsounis, A.L. 6—From drupes to olive oil: An exploration of olive key metabolites. In Olive and Olive Oil Bioactive Constituents; Boskou, D., Ed.; AOCS Press: Urbana, IL, USA, 2015; pp. 147–177. [Google Scholar]
- Uccella, N. Olive biophenols: Biomolecular characterization, distribution and phytoalexin histochemical localization in the drupes. Trends Food Sci. Technol. 2000, 11, 315–327. [Google Scholar] [CrossRef]
- Kanakis, P.; Termentzi, A.; Michel, T.; Gikas, E.; Halabalaki, M.; Skaltsounis, A. From olive drupes to olive oil. An HPLC-orbitrap-based qualitative and quantitative exploration of olive key metabolites. Planta Med. 2013, 79, 1576–1587. [Google Scholar] [CrossRef] [Green Version]
- Perona, J.S.; Botham, K.M. Olive oil as a functional food: Nutritional and health benefits. In Handbook of Olive Oil; Springer: Boston, MA, USA, 2013; pp. 677–714. [Google Scholar]
- Speroni, E.; Guerra, M.; Minghetti, A.; Crespi-Perellino, N.; Pasini, P.; Piazza, F.; Roda, A. Oleuropein evaluated in vitro and in vivo as an antioxidant. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 1998, 12, S98–S100. [Google Scholar] [CrossRef]
- Rizzo, M.; Ventrice, D.; Giannetto, F.; Cirinnà, S.; Santagati, N.A.; Procopio, A.; Mollace, V.; Muscoli, C. Antioxidant activity of oleuropein and semisynthetic acetyl-derivatives determined by measuring malondialdehyde in rat brain. J. Pharm. Pharmacol. 2017, 69, 1502–1512. [Google Scholar] [CrossRef]
- Hassen, I.; Casabianca, H.; Hosni, K. Biological activities of the natural antioxidant oleuropein: Exceeding the expectation—A mini-review. J. Funct. Foods 2015, 18, 926–940. [Google Scholar] [CrossRef]
- Pang, K.-L.; Chin, K.-Y. The biological activities of oleocanthal from a molecular perspective. Nutrients 2018, 10, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Impellizzeri, J.; Lin, J. A simple high-performance liquid chromatography method for the determination of throat-burning oleocanthal with probated antiinflammatory activity in extra virgin olive oils. J. Agric. Food Chem. 2006, 54, 3204–3208. [Google Scholar] [CrossRef] [PubMed]
- Muzzalupo, I. Olive Germplasm: The Olive Cultivation, Table Olive and Olive Oil Industry in Italy; BoD–Books on Demand: Norderstedt, Germany, 2012. [Google Scholar]
- Cicatelli, A.; Fortunati, T.; De Feis, I.; Castiglione, S. Oil composition and genetic biodiversity of ancient and new olive (Olea europea L.) varieties and accessions of southern Italy. Plant Sci. 2013, 210, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Scognamiglio, M.; D’Abrosca, B.; Fiumano, V.; Golino, M.; Esposito, A.; Fiorentino, A. Seasonal phytochemical changes in Phillyrea angustifolia L.: Metabolomic analysis and phytotoxicity assessment. Phytochem. Lett. 2014, 8, 163–170. [Google Scholar] [CrossRef]
- Starec, M.; Calabretti, A.; Berti, F.; Forzato, C. Oleocanthal quantification ising 1H NMR spectroscopy and polyphenols HPLC analysis of olive oil from the Bianchera/Belica cultivar. Molecules 2021, 26, 242. [Google Scholar] [CrossRef]
- Karkoula, E.; Skantzari, A.; Melliou, E.; Magiatis, P. Direct measurement of oleocanthal and oleacein levels in olive oil by quantitative 1H NMR. Establishment of a new index for the characterization of extra virgin olive oils. J. Agric. Food Chem. 2012, 60, 11696–11703. [Google Scholar] [CrossRef]
- Servili, M.; Baldioli, M.; Selvaggini, R.; Macchioni, A.; Montedoro, G. Phenolic compounds of olive fruit: One- and two-dimensional nuclear magnetic resonance characterization of Nüzhenide and its distribution in the constitutive parts of fruit. J. Agric. Food Chem. 1999, 47, 12–18. [Google Scholar] [CrossRef]
- Bianco, A.D.; Piperno, A.; Romeo, G.; Uccella, N. NMR Experiments of oleuropein biomimetic hydrolysis. J. Agric. Food Chem. 1999, 47, 3665–3668. [Google Scholar] [CrossRef]
- Scognamiglio, M.; Fiumano, V.; D’Abrosca, B.; Esposito, A.; Choi, Y.H.; Verpoorte, R.; Fiorentino, A. Chemical interactions between plants in Mediterranean vegetation: The influence of selected plant extracts on Aegilops geniculata metabolome. Phytochemistry 2014, 106, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Valentino, G.; Graziani, V.; D’Abrosca, B.; Pacifico, S.; Fiorentino, A.; Scognamiglio, M. NMR-based plant metabolomics in nutraceutical research: An Overview. Molecules 2020, 25, 1444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhatia, A.; Tripathi, T.; Singh, S.; Bisht, H.; Behl, H.M.; Roy, R.; Sidhu, O.P. Comprehensive metabolite profiling in distinct chemotypes of Commiphora wightii. Nat. Prod. Res. 2019, 33, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.; Robards, K. Critical Review. Phenolic compounds in olives. Analyst 1998, 123, 31R–44R. [Google Scholar] [CrossRef]
- Nicolì, F.; Negro, C.; Vergine, M.; Aprile, A.; Nutricati, E.; Sabella, E.; Miceli, A.; Luvisi, A.; De Bellis, L. Evaluation of phytochemical and antioxidant properties of 15 Italian Olea europaea L. cultivar leaves. Molecules 2019, 24, 1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, E.; Dal Toso, R.; Pressi, G.; Bramanti, P.; Meli, R.; Cuzzocrea, S. Protective effect of verbascoside in activated C6 glioma cells: Possible molecular mechanisms. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2010, 381, 93–105. [Google Scholar] [CrossRef]
- Andreadou, I.; Iliodromitis, E.K.; Mikros, E.; Constantinou, M.; Agalias, A.; Magiatis, P.; Skaltsounis, A.L.; Kamber, E.; Tsantili-Kakoulidou, A.; Kremastinos, D.T. The olive constituent oleuropein exhibits anti-ischemic, antioxidative, and hypolipidemic effects in anesthetized rabbits. J. Nutr. 2006, 136, 2213–2219. [Google Scholar] [CrossRef] [Green Version]
- Antoniou, C.; Hull, J. The Anti-cancer Effect of Olea europaea L. Products: A Review. Curr. Nutr. Rep. 2021, 10, 99–124. [Google Scholar] [CrossRef]
- Caturla, N.; Estepa, A.; Micol, V. Chapter 150—The Relationship between oleuropein antimicrobial activity and its effects on biological membranes. In Olives and Olive Oil in Health and Disease Prevention; Preedy, V.R., Watson, R.R., Eds.; Academic Press: San Diego, CA, USA, 2010; pp. 1345–1354. [Google Scholar]
- Cherubini, C.; Migliorini, M.; Mugelli, M.; Viti, P.; Berti, A.; Cini, E.; Zanoni, B. Towards a technological ripening index for olive oil fruits. J. Sci. Food Agric. 2009, 89, 671–682. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | Abbreviation | Collection Site |
|---|---|---|
| Oliva amara | OA | Morigerati |
| Cammarotana | CA | Orria |
| Carolea | CO | Vallo della Lucania |
| Femminella | FT | Torraca |
| Grossale | GR | Torraca |
| Guglia | GU | Morigerati |
| Marinella | MA | Morigerati |
| Nostrale | NO | Castel S. Lorenzo |
| Ogliara | OA | Agropoli |
| Ogliastro | OG | Ogliastro |
| Pizzulella | PI | Orria |
| Provenzale | PR | Torre Orsaia |
| Pisciottana | PS | Pisiotta |
| Racioppa | RA | Vallo della Lucania |
| Ricippudda | RI | Caselle in Pittari |
| Racioppella | RO | Vallo della Lucania |
| Rotondella | RT | Giungano |
| Salella | SA | Salento |
| Sanginara | SN | Stella Cilento |
| Position | 1H | 13C | HMBC Correlations | |
|---|---|---|---|---|
| Compound 1 | 1 | 9.24 | 198.2 | C5, C9 |
| 3 | 4.36 | 99.8 | C5 | |
| 4 | 1.72/2.05 | 37.5 | C3, C5, C6, C9 | |
| 5 | 3.26 | 30.2 | C1, C3, C4, C8, C9 | |
| 6 | 2.54/2.68 | 37.5 | C4, C5, C7, C9 | |
| 7 | 178.9 | |||
| 8 | 6.92 | 157.3 | C1, C5, C9, C10 | |
| 9 | 146.3 | |||
| 10 | 2.05 | 14.8 | C1, C8, C9 | |
| DHPEA-EDA (2) | 1 | 9.09 | 200.6 | C5, C8, C9 |
| 3 | 4.33 | 99.8 | ||
| 4 | 1.72/2.04 | 37.4 | C3, C5, C6, C9 | |
| 5 | 3.18 | 32.5 | C1, C3, C4, C8, C9 | |
| 6 | 2.54/2.72 | 38.4 | C4, C5, C7, C9 | |
| 7 | 174.3 | |||
| 8 | 6.72 | 160.7 | C1, C5, C9, C10 | |
| 9 | 145.1 | |||
| 10 | 1.90 | 17.3 | C1, C8, C9 | |
| 1′ | 130.6 | |||
| 2′ | 6.70 | 116.1 | C1′, C6′, C7′ | |
| 3′ | 144.3 | |||
| 4′ | 143.6 | |||
| 5′ | 6.79 | 115.9 | C1′, C3′, C4′, C6′ | |
| 6′ | 6.58 | 120.7 | C1′, C2′, C5′, C7′ | |
| 7′ | 2.77/2.80 | 33.6 | C7 | |
| 8′ | 4.17/4.28 | 65.6 | C1′, C7, C7′ | |
| Oleomissional (3) | 1 | 9.04 | 197.5 | C4, C8, C9 |
| 3 | 9.00 | 197.1 | C4, C9 | |
| 4 | 3.47 | 32.5 | C1, C3, C7, C8, C9 | |
| 7 | 173.5 | |||
| 8 | 6.78 | 157.9 | C1, C5, C9, C10 | |
| 9 | 140.9 | |||
| 10 | 1.90 | 15.5 | C1, C8, C9 | |
| 1′ | 130.6 | |||
| 2′ | 6.70 | 116.1 | C1′, C6′, C7′ | |
| 3′ | 144.3 | |||
| 4′ | 143.6 | |||
| 5′ | 6.79 | 115.9 | C1′, C3′, C4′, C6′ | |
| 6′ | 6.58 | 120.7 | C1′, C2′, C5′, C7′ | |
| 7′ | 2.75/2.80 | 33.6 | C7 | |
| 8′ | 4.17/4.28 | 65.6 | C1′, C7, C7′ | |
| Oleocanthal (4) | 1 | 9.10 | 198.4 | C5, C8, C9 |
| 3 | 4.28 | 96.9 | ||
| 4 | 1.77/2.01 | 38.5 | C3, C5, C6, C9 | |
| 5 | 3.2 | 30.3 | C1, C3, C4, C8, C9 | |
| 6 | 2.56/2.75 | 37.5 | C4, C5, C7, C9 | |
| 7 | - | 175.3 | ||
| 8 | 6.76 | 157 | C1, C5, C9, C10 | |
| 9 | - | 142.9 | ||
| 10 | 1.91 | 14.7 | C1, C8, C9 | |
| 1′ | 130.6 | |||
| 2′/6′ | 7.05 | 129.7 | C1′, C6′, C7′ | |
| 3′/5′ | 6.72 | 115.7 | ||
| 4′ | - | 154.6 | ||
| 7′ | 2.76 | 33.9 | ||
| 8′ | 4.09/4.22 | 65.6 | C1′, C7, C7′ | |
| Cornoside (9) | 1 | 70.8 | ||
| 2/6 | 7.06 | 156.2 | C2, C3, C4, C6 | |
| 3/5 | 6.23 | 129.5 | C1, C2, C4, C5 | |
| 4 | 189.8 | |||
| Halleridone (10) | 1 | 77.5 | ||
| 2 | 83.0 | |||
| 3 | 39.0 | |||
| 4 | 6.08 | 199.9 | ||
| 5 | 6.93 | 130.4 | C1, C3, C4 | |
| 6 | 157.6 | C2, C4, C5 | ||
| Verbascoside (11) | 1′ | 125.1 | ||
| 2′ | 7.12 | 113.3 | C3′, C4′, C6′ | |
| 3′ | 148.4 | |||
| 4′ | 145.2 | |||
| 5′ | 6.89 | 112.3 | C1′, C3′, C4′ | |
| 6′ | 7.08 | 121.4 | C2′, C4′ | |
| 7′ | 7.67 | 147.6 | C2′. C3′, C6′, C9′ | |
| 8′ | 6.36 | 113.9 | C1′, C9′ | |
| Glc-1 | 4.49 | 102.3 | ||
| Rha-1 | 5.13 | 101.8 | ||
| Rha-6 | 1.06 | 17.3 |
| Metabolites | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cultivar | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 |
| RO | ● | ● | ● | ● | ● | ● | ● | ● | |||
| GU | ● | ● | ● | ● | ● | ● | ● | ||||
| PI | ● | ● | ● | ● | ● | ● | ● | ||||
| MA | ● | ● | ● | ● | ● | ● | |||||
| RA | ● | ● | ● | ● | ● | ● | ● | ● | |||
| PS | ● | ● | ● | ● | ● | ● | ● | ||||
| PR | ● | ● | ● | ● | ● | ● | ● | ||||
| GR | ● | ● | ● | ● | ● | ● | ● | ||||
| FT | ● | ● | ● | ● | ● | ● | |||||
| AM | ● | ● | ● | ● | ● | ● | |||||
| OG | ● | ● | ● | ● | ● | ● | |||||
| SN | ● | ● | ● | ● | ● | ||||||
| CA | ● | ● | |||||||||
| OA | ● | ● | ● | ● | ● | ||||||
| CO | ● | ● | ● | ● | ● | ||||||
| NO | ● | ● | ● | ● | ● | ● | ● | ● | ● | ● | |
| SA | ● | ● | ● | ● | |||||||
| RT | ● | ● | ● | ||||||||
| RI | ● | ● | |||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Esposito, A.; De Luca, P.F.; Graziani, V.; D’Abrosca, B.; Fiorentino, A.; Scognamiglio, M. Phytochemical Characterization of Olea europaea L. Cultivars of Cilento National Park (South Italy) through NMR-Based Metabolomics. Molecules 2021, 26, 3845. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26133845
Esposito A, De Luca PF, Graziani V, D’Abrosca B, Fiorentino A, Scognamiglio M. Phytochemical Characterization of Olea europaea L. Cultivars of Cilento National Park (South Italy) through NMR-Based Metabolomics. Molecules. 2021; 26(13):3845. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26133845
Chicago/Turabian StyleEsposito, Assunta, Pietro Filippo De Luca, Vittoria Graziani, Brigida D’Abrosca, Antonio Fiorentino, and Monica Scognamiglio. 2021. "Phytochemical Characterization of Olea europaea L. Cultivars of Cilento National Park (South Italy) through NMR-Based Metabolomics" Molecules 26, no. 13: 3845. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26133845