
Plants Secondary Metabolites as Blood Glucose-Lowering Molecules


Department of Pharmaceutical Sciences, School of Pharmacy, The University of Jordan, Amman 11942, Jordan
*
Author to whom correspondence should be addressed.
Molecules 2021, 26(14), 4333; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26144333
Submission received: 2 June 2021
/
Revised: 6 July 2021
/
Accepted: 7 July 2021
/
Published: 17 July 2021
(This article belongs to the Special Issue Bioactive Molecules: Isolation, Synthesis, Analysis, and Application in Medicinal Chemistry)
Abstract
:Recently, significant advances in modern medicine and therapeutic agents have been achieved. However, the search for effective antidiabetic drugs is continuous and challenging. Over the past decades, there has been an increasing body of literature related to the effects of secondary metabolites from botanical sources on diabetes. Plants-derived metabolites including alkaloids, phenols, anthocyanins, flavonoids, stilbenoids, saponins, tannins, polysaccharides, coumarins, and terpenes can target cellular and molecular mechanisms involved in carbohydrate metabolism. In addition, they can grant protection to pancreatic beta cells from damage, repairing abnormal insulin signaling, minimizing oxidative stress and inflammation, activating AMP-activated protein kinase (AMPK), and inhibiting carbohydrate digestion and absorption. Studies have highlighted many bioactive naturally occurring plants’ secondary metabolites as candidates against diabetes. This review summarizes the current knowledge compiled from the latest studies published during the past decade on the mechanism-based action of plants-derived secondary metabolites that can target various metabolic pathways in humans against diabetes. It is worth mentioning that the compiled data in this review will provide a guide for researchers in the field, to develop candidates into environment-friendly effective, yet safe antidiabetics.
1. Introduction
Millions of years ago, plants appeared on land and prevailed in almost every habitat. Nearly 400,000 species have been recognized and assorted to date. In ancient medicine systems practiced worldwide, such as Ayurveda, Chinese and Egyptian, plants played a primary role in treating a myriad of diseases [1,2].
Nature contains an astronomical chemical space providing numerous compounds that impact human health and disease states. Over history, humankind has used natural products in crude form or as an extract. After the 20th revolution, scientists started to think about using pure isolated chemicals and elucidating their structures. The modern pharmaceutical industry has partially been founded by research looking for new therapeutic agents from medicinal plants [3].
Natural products are organic compounds synthesized by living organisms, e.g., plants, animals, and micro-organisms, which have biological activities. Secondary metabolites of botanical origin are produced to support and enhance the permanence of the plant. Photosynthesis, glycolysis and the Krebs cycle are overflowing with several intermediates that manufacture these metabolites [4,5].
Phytomedicine is a plant-based medicine, and it is one of the subsets of complementary and alternative medicinal (CAM) therapies [6]. The World Health Organization (WHO) defined herbal medicine as the knowledge, skills and practices based on the views and experiences related to different cultures used in the preservation of health and the prevention, diagnosis, improvement, or treatment of illness [2,7]. Traditional herbal medicines are plant-derived substances with minimal or no industrial processing that have been used to treat illness within local or regional healing conventionalisms [2].
Diabetes mellitus is a chronic progressive metabolic disorder of endocrine origin. It is characterized by disturbances of carbohydrate metabolism associated with hyperglycemia (WHO). Insulin is the key to maintaining an average level of blood glucose. In diabetes, insulin production is either absent or decreased, leading to hyperglycemia [6]. Diabetes is swiftly increasing: in 2019, approximately 463 million people had diabetes. According to the IDF (International Diabetes Federation) estimates, the number is expected to increase to 578 million in 2030 and 700 million in 2045 [8]. Diabetes mellitus can be treated by regulating the blood sugar level with different pharmacological and non-pharmacological approaches. Exercises, diet plans and diverse types of medications including insulin, glycosidase inhibitors, glycosuric, biguanides, meglitinides, sulphonylureas, thiazolidinediones and peptide analogs [6].
Published research has proven that several secondary metabolites demonstrate hypoglycemic activity in vivo and in vitro, and usually, they affect multiple targets, proteins and enzymes. Alkaloids, phenols, anthocyanin, flavonoids, saponins, tannins, terpenes and coumarins were found to elicit a significant influence on diabetes [9,10].
Phlorizin is a natural dihydrochalcone found in a number of fruitful trees, mainly the Malus genus. It produces renal glycosuria and blocks intestinal and renal glucose reabsorption by inhibiting the sodium-glucose symporters located there [11,12]. Recently, ertugliflozin arrived as the newest synthetic GLT-2 (glucose transporter-2) inhibitor to receive FDA approval for the treatment of diabetes [13,14].
This review highlights the proposed secondary metabolites mechanism-based action targeting various metabolic pathways involved in glucose metabolism in humans. The summarized data from in vivo and in vitro bioassays of phytochemicals will lead future research towards developing effective antidiabetics with low toxicity.
2. Mechanisms Involved in Glucose Metabolism and Homeostasis
Carbohydrates comprise the major constituents of a regular human diet. Starch digestion starts in the mouth and continues through the upper gastrointestinal tract until the intestine. It is converted into smaller molecules (monosaccharides) absorbed into the bloodstream [15].
Starch is mainly converted to glucose by the action of α- amylase that hydrolyzes macromolecules into oligoglucans, and α-glucosidase further degrades oligoglucans into absorbable glucose molecules at the brush border of the small intestine and is then absorbed through the glucose transporters sodium-dependent glucose cotransporter 1 (SGLT1) and glucose transporter 2 (GLUT2) [15,16]. Several phytoconstituents are known to suppress the activity of α-amylase, α-glucosidase and to inhibit intestinal absorption of glucose, inhibiting postprandial hyperglycemia and keeping the concentration of glucose in the blood constant after a meal [15,17].
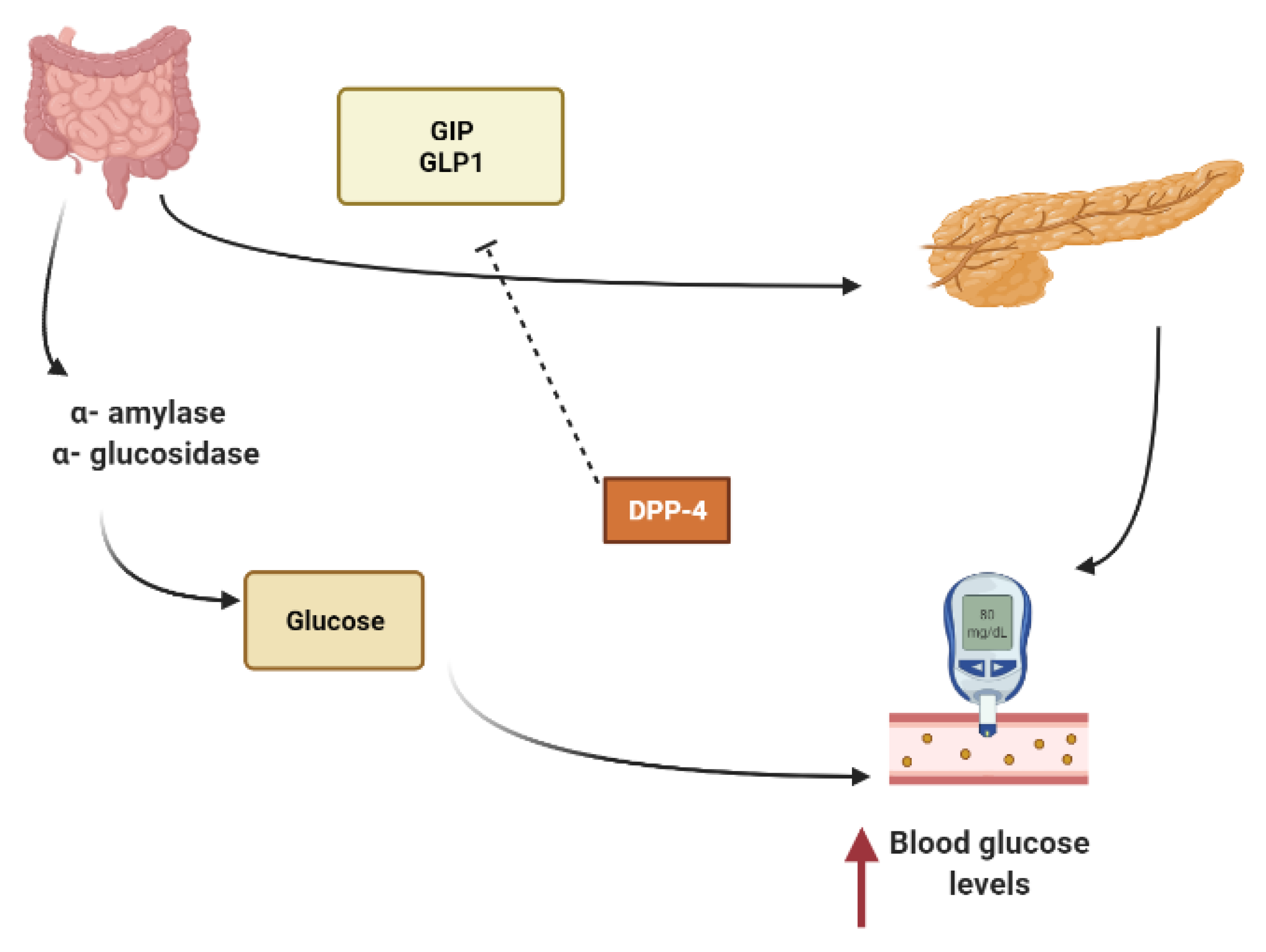
Other secondary metabolites can modulate the secretion of glucagon-like peptide-1 (GLP1), inhibiting dipeptidyl peptidase-4 (DPP4) from extending the effect of GLP1. In L-cells of the small intestine, basal serum GLP1 is released in response to nutrients loads (Figure 1). It promotes insulin secretion and stimulates the hypothalamus gland to induce postprandial satiety and to inhibit glucagon secretion. GLP1 has a very short half-life because it is hydrolyzed by dipeptidyl peptidase-4 (DPP4) [18,19,20].
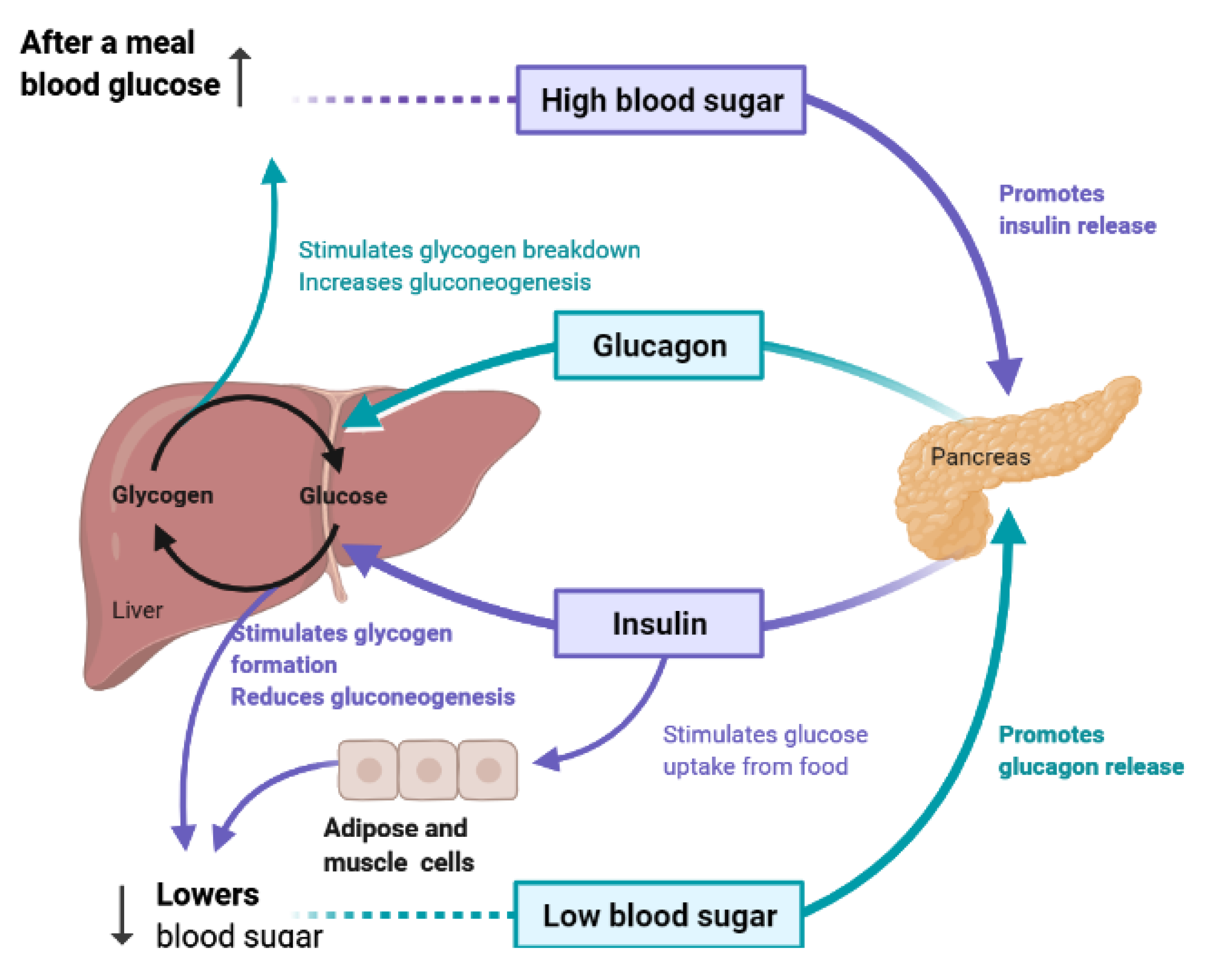
To maintain constant blood glucose levels, various body organs, including the pancreas, liver, intestine, adipose and muscle tissues with various hormones and neuropeptides, work together. The pancreas plays a crucial role in glucose homeostasis by secreting insulin and its opponent glucagon [21]. The increased circulating glucose is sensed by pancreatic β-cells and subsequently glucose influx into the cells via GLUT2, an insulin-independent transporter (Figure 2). Glucose induces insulin secretion from β-cells via the closure of ATP-gated potassium channels and activation of voltage-gated calcium channels [16,21,22,23].
3. Insulin Signal Transduction
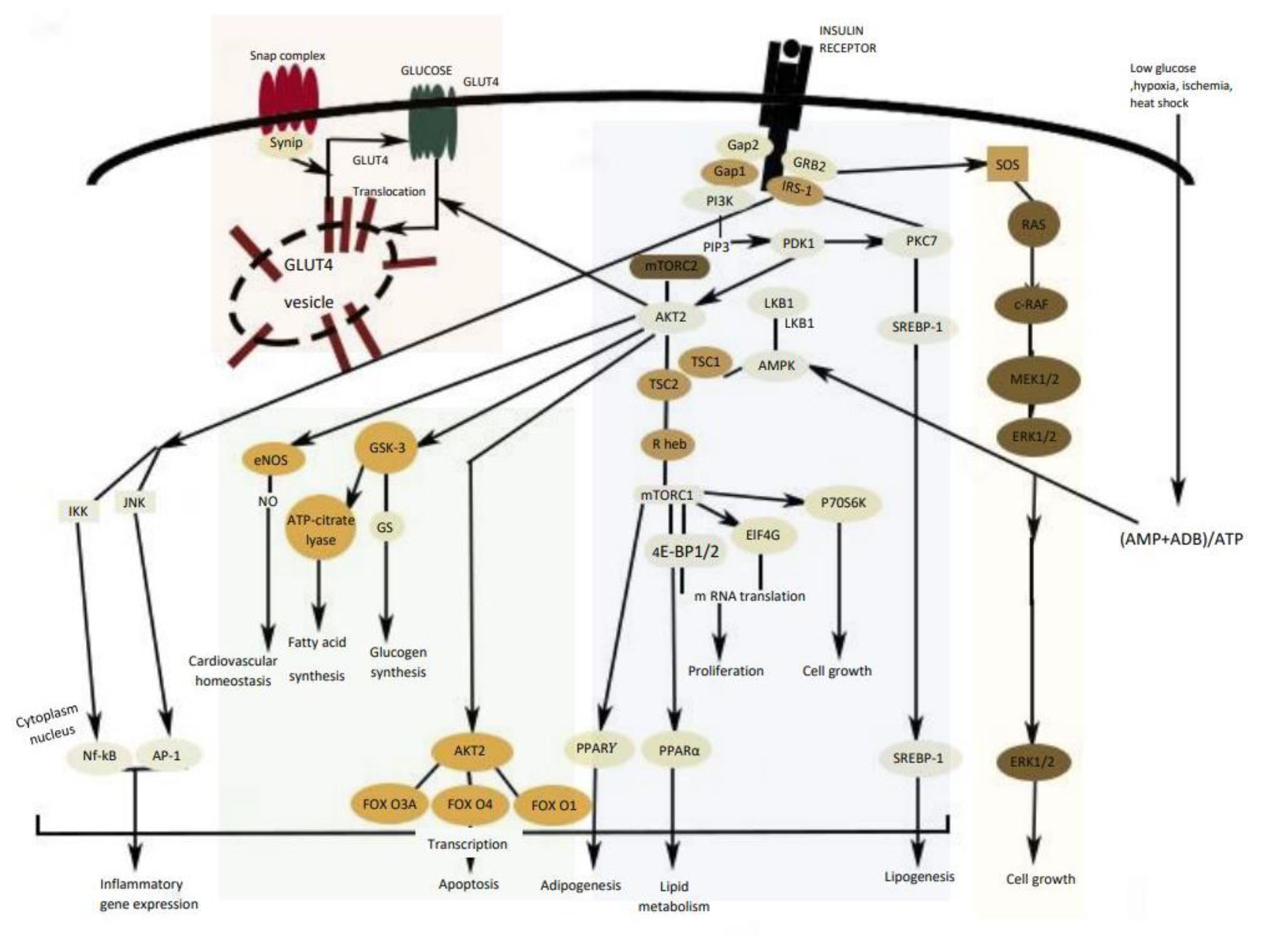
Insulin modulates several metabolic pathways through a cascade of signal transduction steps initiated when the insulin binds to the insulin receptor (IR), stimulating a receptor’s intrinsic kinase activity [21]. Insulin receptor substrates IRS1 and IRS2 are phosphorylated by several kinases such as ERK (extracellular signal-regulated kinases), JNK (c-Jun N-terminal kinase), and AMPK (AMP-activated protein kinase). Consequently, insulin substrates activate a number of kinases, including PI3K (PI3K: phosphoinositide 3-kinase)/Akt pathway as well as the MAPK (MAPK: mitogen-activated protein kinase) pathway [24,25].
The IRS protein activates phosphatidylinositol 3 kinase (PI3K) that causes the activation of Akt, the main mediator to activate the most biochemical mechanism in glucose metabolism [26]. Akt activates the glucose transporter translocation to the cellular membrane (GLUT-4) and triggers the phosphorylation of glycogen synthase kinase 3 (GSK3), which leads to stimulation of glycogen synthesis in liver and skeletal muscle and downregulation of PEPCK (phosphoenolpyruvate carboxykinase) and G6Pase (glucose-6- phosphatase) gene expression [24]. However, in a diabetic state, these pathways are not blocked because insulin is unable to adequately regulate the gene expression and function of PEPCK and G6Pase, leading to excessive hepatic glucose production through gluconeogenesis and glycogenolysis [9,16]. In addition, the activation of the Akt system leads to protein synthesis through the activation of the mechanistic target of rapamycin complex 1 (mTORC1), cell survival by the suppression of various pro-apoptotic molecules, mainly of the FOXO (forkhead box O) family of transcription factors (Figure 4) [27].
In the liver, insulin activates AMPK which leads to the inhibition of acetyl-CoA carboxylase (ACC) and sterol regulatory element-binding protein (SREPB-1) activity, thereby inhibiting fatty acid biosynthesis and increasing fatty acid oxidation [24].
MAPK is a specific protein kinase involved in various physiological and biochemical mechanisms, including cell differentiation, proliferation, apoptosis, and cell endurance. ERK1/2 (extracellular-signal-regulated kinase 1/2) and JNK (c-Jun N-terminal kinase) are other cell signaling kinases co-task with MAPK, involved in cell growth, differentiation, inflammatory response, and apoptosis. Overstimulation of MAPK generally provides the failure of insulin synthesis linked with the apoptosis process in pancreatic islet cells and downregulates GLUT4 expression [25,28].
4. Liver and Glucose Homeostasis
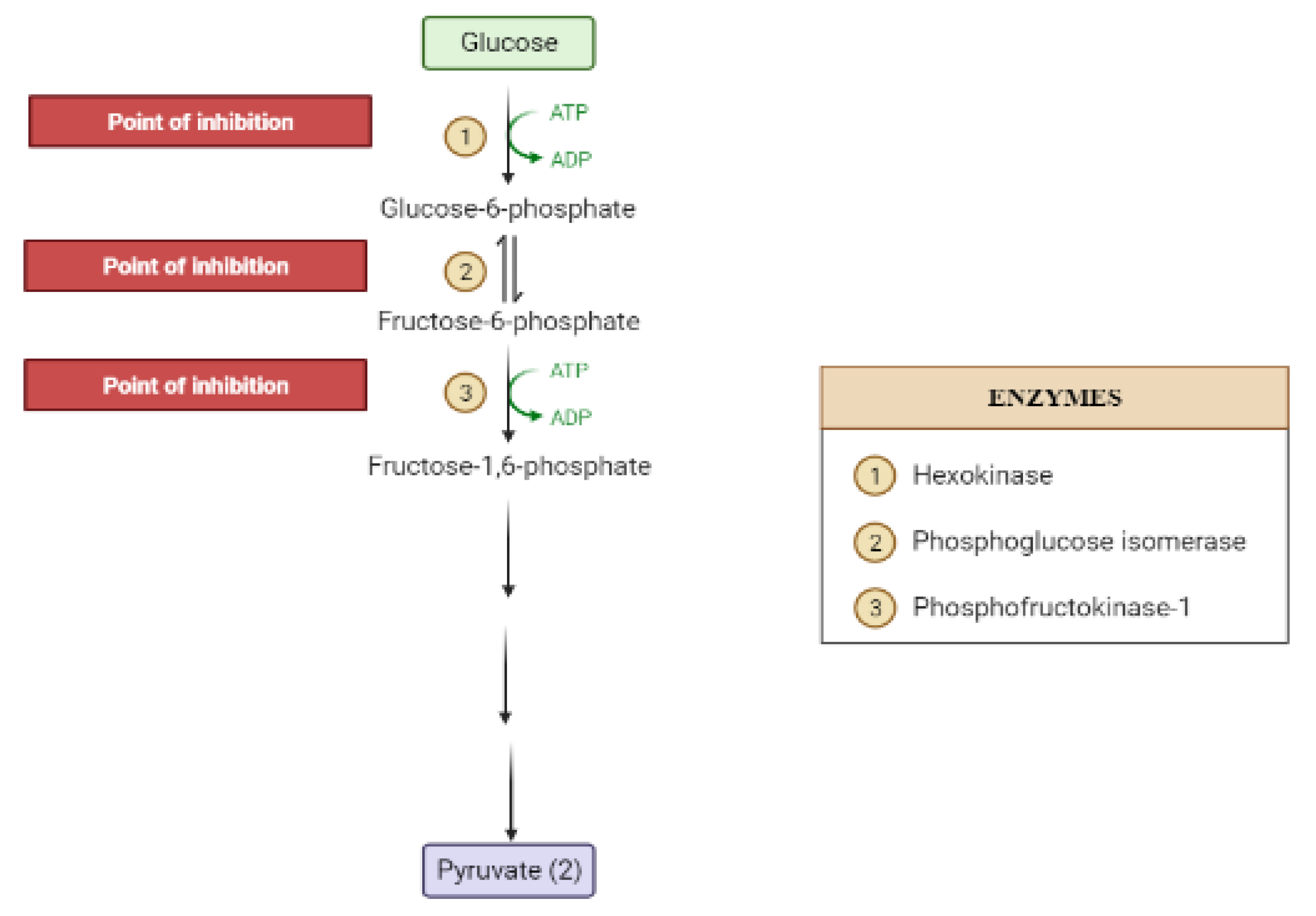
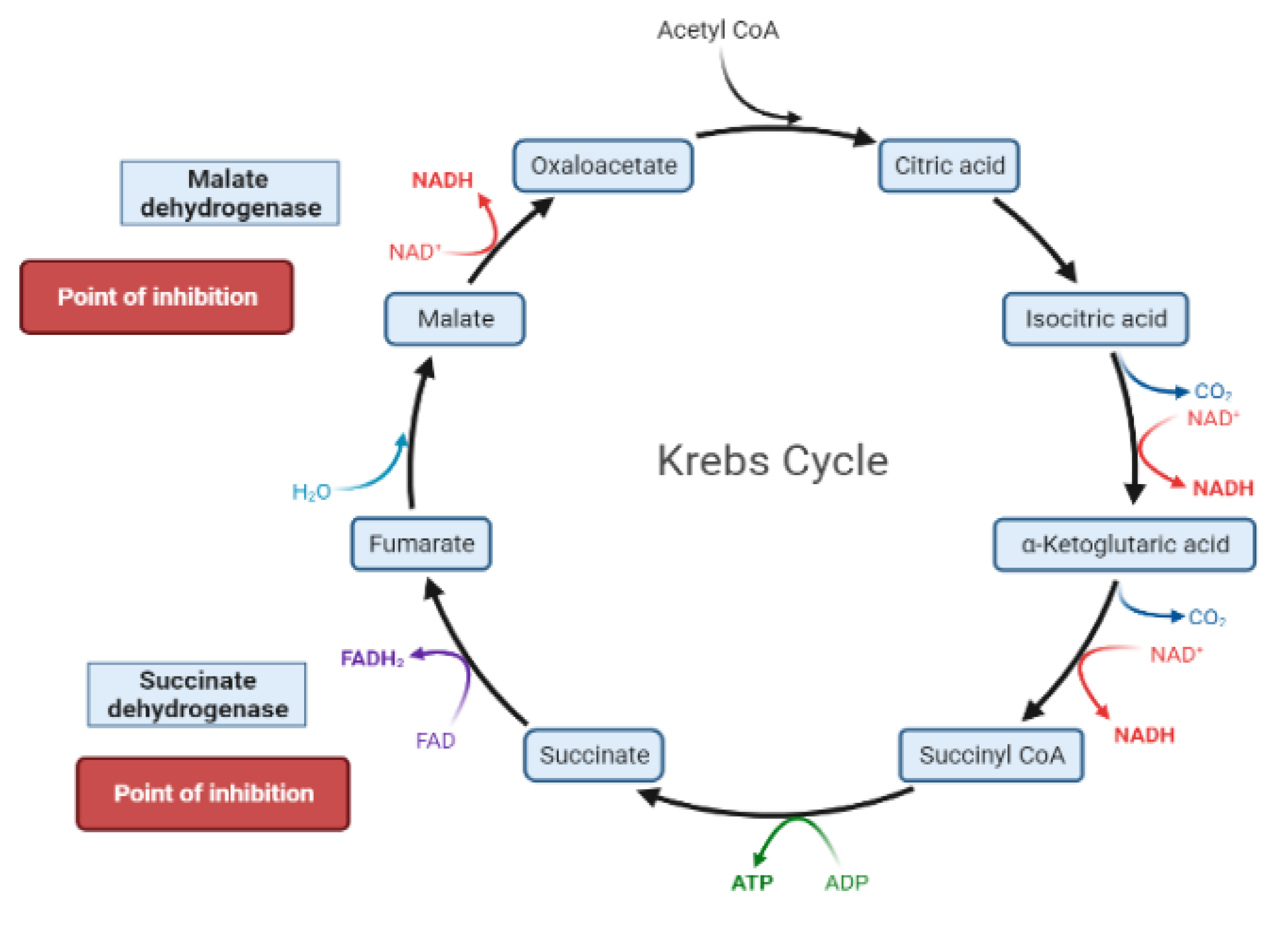
The liver is the chief organ in maintaining glucose homeostasis through controlling various pathways, including glycogenesis, glycogenolysis, glycolysis and gluconeogenesis [29]. During feeding conditions, glucose is a primary fuel source across multiple fuel sources across numerous body tissues by eliciting ATP molecules during hydrolysis and the Krebs cycle. The increase in glucose uptake in hepatocytes promotes glycolysis and lipogenesis to generate triglycerides as storage forms of fuel [22,29]. Glycolysis can be regulated through the reactions catalyzed by hexokinase, phosphofructokinase, and pyruvate kinase (Figure 5 and Figure 6). Moreover, there are seven enzymes involved in this Krebs cycle, of which only two enzymes, succinate dehydrogenase and malate synthase, can be regulated. Several plant secondary metabolites (discussed below) can regulate and affect these enzymes except for the pyruvate kinase enzyme [22,30,31].
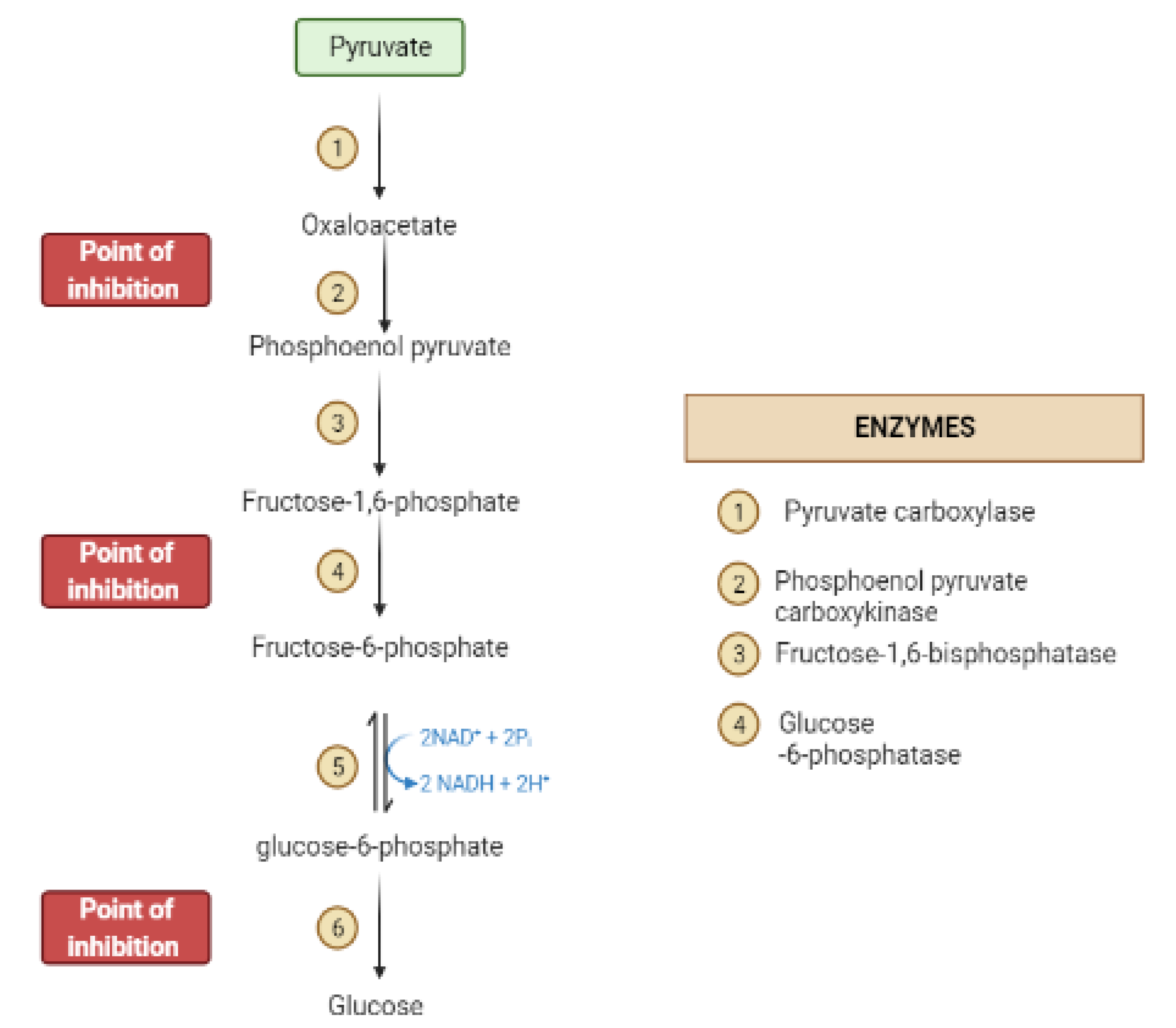
During the fasting state, the liver is the leading factory for glucose production through gluconeogenesis and glycogenolysis. The main regulatory enzymes in the gluconeogenesis pathway include glucose 6-phosphatase (G6Pase), fructose 1, 6-bisphosphatase (Fbpase1), PC (pyruvate carboxylase), and phosphoenolpyruvate carboxykinase (PEPCK), all can be inhibited by plant secondary metabolites (Figure 7) [29,30].
5. Obstacles for Insulin Signal Transduction and Insulin Effects
Several obstacles can affect the signal transduction of insulin in different tissues. Protein tyrosine phosphatase 1B (PTP1B) is a known negative regulator of the insulin-stimulated signal transduction pathway. PTP1B is localized on the cytoplasmic surface of the endoplasmic reticulum in classical insulin targeted tissues such as the liver, muscle, and fat. PTP1B catalyzes the dephosphorylation of activated insulin receptors (IR), resulting in down-regulation of insulin signaling [31,32].
The increase in the free fatty acids is connected to alteration in the diacylglycerol (DAG)/protein kinase C (PKC) pathway. The rise in DAG activates PKC-θ, -β2 and –δ that phosphorylates IRS1, which interferes with insulin-stimulated phosphorylation of IRS1, thus inhibiting insulin signaling. It is believed that targeting PKC is beneficial in treating type 2 diabetes as it will address the secretory defect promoting insulin secretion [33,34].
Inflammatory mediators (such as TNF-α, Il-6) can impair insulin and promote serine phosphorylation of IRS-1 impairing insulin signaling or can cause degradation of IRS. It can reduce GLUT-4 expression, decreasing glucose entry to cells and cause inflammation-induced nitric oxide release that suppresses PI3K–Akt pathway [23]. It is well-known that inflammation is a crucial cause for diabetes type-2. Relieving inflammation by secondary metabolites can improve diabetes [35].
Oxidative stress has been linked to diabetes. It comprises insulin secretion and insulin action. It activates NF-κB and JNK, IRS degradation, suppresses GLUT-4 expression and translocation, activates inflammatory responses [36,37]. Plant secondary metabolites can stimulate mitochondrial metabolism and/or decrease mitochondrial dysfunction through targeting sirtuin 1 activators (SIRT1) and PPAR α [37].
6. Skeletal Muscle and Adipose Tissue
Insulin works on adipose tissue and skeletal muscle tissue where it causes glucose uptake, use and storage by skeletal muscle (Figure 9). Glucose uptake is mediated by the combined influence of glucose concentrations, insulin signaling, insulin receptor substrate (IRS)-1/phosphatidylinositol kinase (PI3K)/kinase B (or Akt) pathway, and increasing membrane localization of transporters such as glucose transporter 4, GLUT4 [16,24].
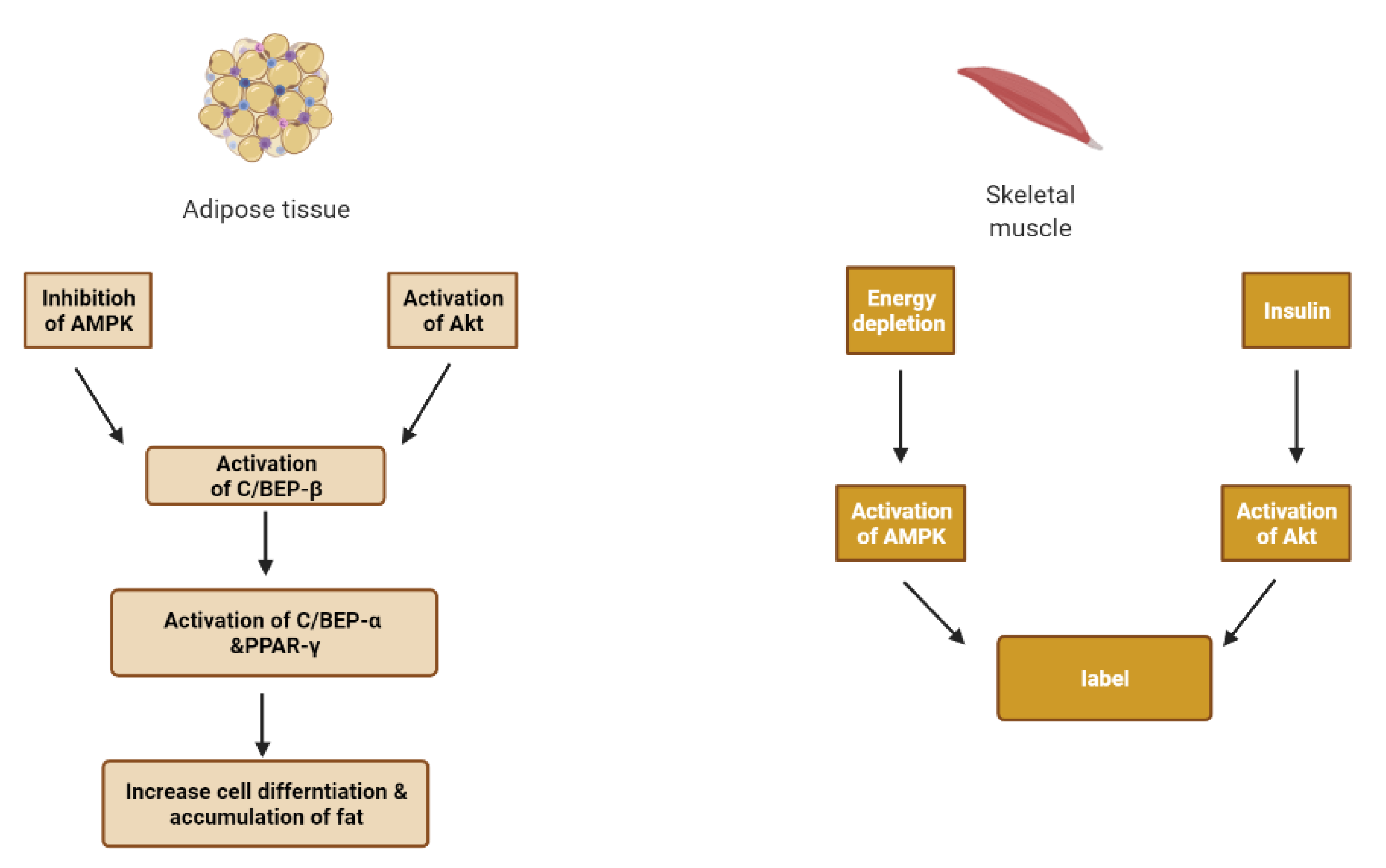
Adipose tissue plays a major role in the ability of the body to sense insulin. When nutrient intake exceeds the capacity of fat cells to store excess calories, hypoxia appears in adipose tissue. Subsequently, hypoxia-inducible factor-1 (HIF-1) is activated, and the expression of c-Jun N-terminal kinase (JNK) and inhibitor nuclear factor kappa-B kinase (IKK) is increased, producing inflammation in adipose tissue [33].
With the aggravation of inflammation, many inflammatory cytokines are released to exacerbate insulin resistance and lipolysis, reduce the activity of peroxisome proliferator-activated receptor γ (PPARγ), and accelerate the fat cell death and inflammation [33].
A vicious cycle is formed due to decreased insulin activity as an anti-lipolytic hormone and hyperinsulinemia that activates the lipoprotein lipase and causes free fatty acid (FFA) release from lipoproteins [33].
A high level of FFA causes the accumulation of diacylglycerol (DAG) and ceramide. DAG can inhibit insulin action by activating the protein kinase C isoforms (PKC) and interfering with insulin signal transduction by serine phosphorylation of insulin receptors (IRS) [33,39]. Ceramide is a potent activating agent of inflammation. It can activate JNK and NF-κB/IKK, which are closely related to insulin resistance. It can induce pancreatic β-cell apoptosis and reducing insulin gene expression [33,40].
Additionally, fat deposits in the liver and is accompanied by the build-up of insulin resistance. Kupffer cells in the liver are activated to release inflammatory cytokines. Consequently, nuclear factor κB (NF-κB) is activated and regulates the inflammatory state [33].
Lipid peroxidation and glucose autoxidation give rise to the accumulation of reactive oxygen species (ROS) and free radicals. Thereby, increased active oxygen groups result in hepatic oxidative injury and jams of antioxidant enzymes such as SOD (superoxide dismutase), CAT (catalase) and GP (glutathione peroxidase), which further aggravate the progression of insulin resistance and pancreatic β-cell dysfunction [33].
Peroxisome proliferator-activated receptor gamma (PPAR) is expressed in several tissues, especially macrophages, adipose, muscle and mainly involved in adipocyte differentiation. The PPARs and retinoid X receptors regulate some genes involved in lipid and carbohydrate metabolism. There are three human isoforms of PPAR; α, δ and γ [15].
PPARγ plays a key role in adipogenesis and glucose regulation. Activated PPAR γ binds to retinoic acid X receptor to regulate transcriptional activation of downstream target genes linked to diabetes. In addition, it is also reported to enhance GLUT-1 and GLUT-4 translocation in the liver and skeletal muscle. It also potentiates insulin sensitivity by reducing TNF-α and elevating adiponectin expression. On the other hand, PPARα is a regulator of fatty acid catabolism and peroxisome proliferation in the liver [15]. Table 1 summarizes the biological effects of insulin in normal and diabetic status [41].
7. Methodology
The key words of: diabetes, plant secondary metabolites, alkaloids, phenols, anthocyanins, flavonoids, stilbenoids, saponins, tannins, polysaccharides, coumarins and terpenes, were searched for in the Pub-Med, SciFinder, EBSCO, ScienceDirect and Google-scholar databases. Relevant papers in the English language from the year 2010 up to 2021 were collected. To qualify for inclusion in this review, publications should have reported biological and chemical data for isolated secondary metabolites with in vitro or in vivo test models of type 2 diabetes.
8. Result and Discussion
8.1. Secondary Metabolites and Antidiabetic Activity
8.1.1. Alkaloids
Alkaloids are a remarkably diverse group of nitrogen-containing compounds with limited allocation in the plant kingdom, especially in angiosperms. They may contain one or more nitrogen atoms within a heterocyclic ring and are classified as pyrrolidine, pyridine, quinoline, isoquinoline, indole, quinazoline, steroidal, diterpenoid, and other alkaloids. They have a wide spectrum of biological activities, including hypoglycemic. The therapeutic impact of alkaloids against diabetes is mediated through various pathways and signaling cascades (Figure 10) [27,42,43,44].
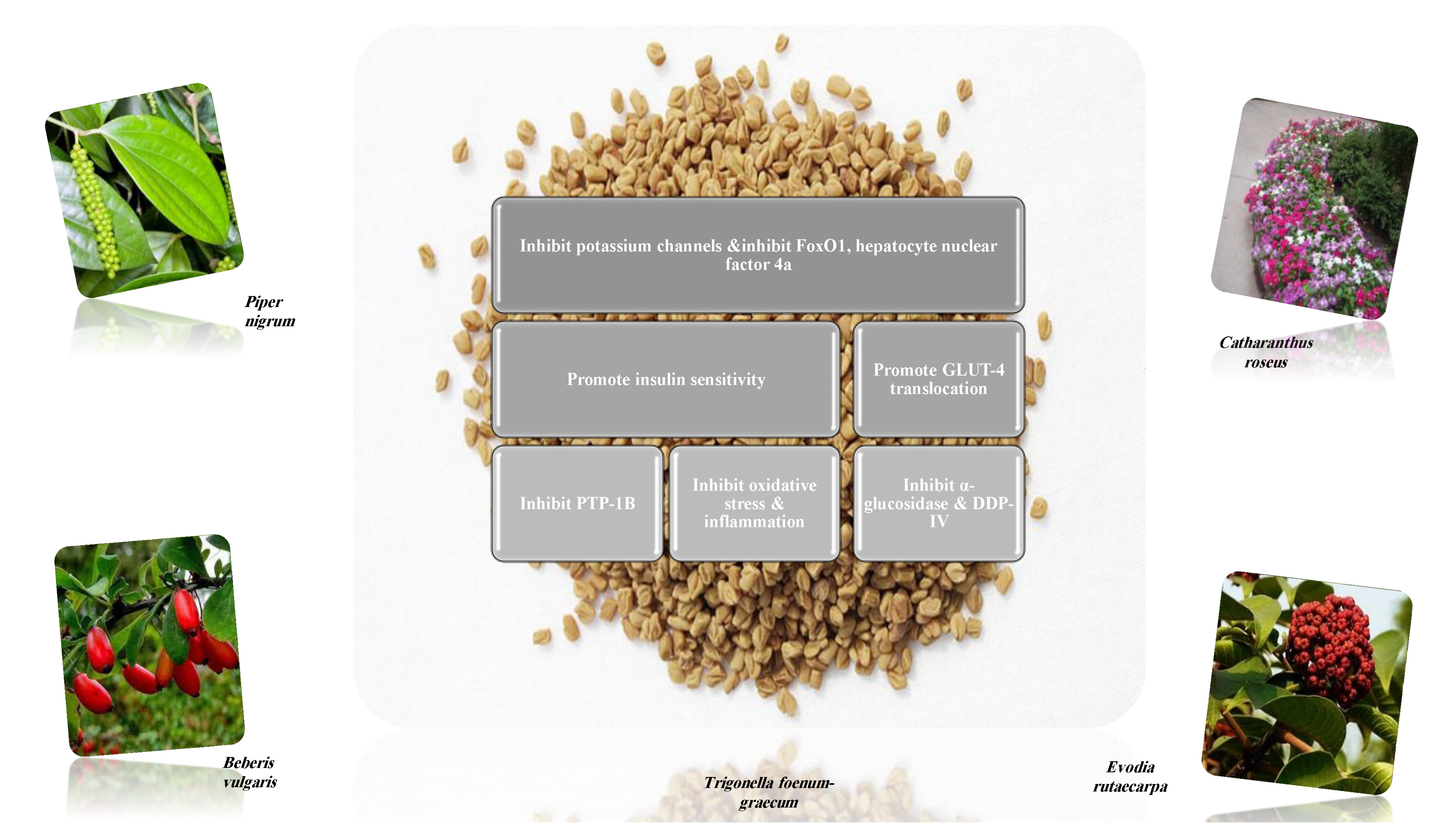
Alkaloids are considered eminent inhibitors of α-glucosidase and α- amylase enzymes, one of the most effective approaches to decrease the level of blood glucose in type 2 diabetes, for example, broussonetine and radicamine [45,46]. Alkaloids inhibit dipeptidyl peptidase-4 (DDP-4), for example, bebeerine that degrades gut incretin hormones, including glucagon-like peptide-1 (GLP-1) and gastric inhibitory polypeptide (GIP). A peptide that induces β-cell differentiation stimulates insulin biosynthesis and release, inhibits gastric emptying, directly reducing food intake and prevents glucagon release from the α-cells of the islets of Langerhans [20,47,48].
Adenosine monophosphate (AMP) activated protein kinase is a cellular fuel sensor and glucose transporter regulator. It stimulates glucose uptake and modulates insulin secretion. Several alkaloids are allosteric activators of AMPK and enhance its enzymatic activity [45,49].
Alkaloids increase glucose-4 transporter translocation, increase insulin secretion via regenerating the pancreatic β-cells and enhanced glucose-stimulated insulin secretion (GSIS) rather than basal insulin secretion [45]. Examples of hypoglycemic alkaloids and their proposed mechanisms of action (Table 2). Several in vitro and in vivo studies were designed to explore the antidiabetic activity of different isolated alkaloids (Table 3).
On the other hand, some were reported to trigger insulin secretion such as bisbenzylisoquinoline, quinolizidine, isoquinoline, indole and pyridine alkaloids [46].
Several plant families: Acanthaceae, Apocynaceae, Bignoniaceae, Campanulaceae, Euphorbiaceae, Fabaceae, Moraceae, Piperaceae, Portulacaceae, Ranunculaceae, and Rutaceae, were investigated for their alkaloids-rich parts. All exhibited various mechanisms of action for their antidiabetic activity. The most frequent mechanism was the digestive enzymes inhibitory activity [42].
Alkaloids have diverse and variable structures. They inhibit enzymes at a lower dose compared to other metabolites. They affect multiple targets to lower hyperglycemia, but α-glucosidase inhibition seems to be the most promising mechanism [46,47,48,50].
Regrettably, the toxicity of some alkaloids, instability under physiologic pH, hence low bioavailability and short half-life, may limit their clinical use. However, new pharmaceutical delivery techniques can overcome the issues. Further investigation is needed to explore their pharmacological mode of action in-vivo and in clinical setting [46,50].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 2.
Proposed antidiabetic effects of different alkaloids.
| Name of Compound | Chemical Structure | Type of Alkaloid | Mechanism | References |
|---|---|---|---|---|
| Berberine |  | Isoquinoline |
| [27,48,51,52] |
| Evodiamine |  | Quinolone |
| [27,46] |
| Glycosin |  | Quinazoline |
| [46] |
| Lupanine |  | Quinolizidine |
| [45,46] |
| Neferine |  | Isoquinoline |
| [27] |
| Piperine |  | Piperidine |
| [27] |
| Oxymatrine |  | Quinolizidine |
| [27,47] |
| Trigonelline |  | Pyridine |
| [27,48] |
| Vidoline |  | Indole |
| [27,48] |
Table 3.
In vitro, in vivo and in silico models for different isolated alkaloids.
| Isolated Compound | Type of Alkaloids | Mechanism | Dose of the Tested Compound (Duration) | Model | References |
|---|---|---|---|---|---|
| Berberine | Isoquinoline | Increased mRNA and protein expressions of GLUT-4 and reduced activation of the hypothalamus-pituitary-adrenal axis. | 200 mg/kg (4 weeks) | High-fat diet and streptozocin-induced diabetic rats | [50] |
| Modulated gut microbiota. | 200 mg/kg (8 weeks) | High-fat diet obese rats | [51] | ||
| Inhibited expression of the gluconeogenic proteins (PEPCK and G-6-Pase) in the liver. | 156 mg/kg (12 weeks) | High-fat diet and streptozocin-induced diabetic rats | [52] | ||
| Increased expression of skeletal muscle GLUT- 4, mRNA had antioxidant activity. | 50, 100 mg/kg (6 weeks) | High- fat and glucose diet hamsters | [53] | ||
| Conophylline | Vinca | Pancreatic β-cells regenerator. | 0.1 µg/mL | In vitro: ICC cell line | [54] |
| Coptisine | Isoquinoline | PTP1B inhibition. | 6.25–50 µM | In vitro: enzymatic assay and in silico | [55] |
| Ephedrine | Phenylalanine derived | DDP-4 inhibition, IC50 was 124 µM | 10−5–10−3 M | In vitro: binding assay and in silico | [56] |
| Evodiamine | Quinolone | Activated AMPK phosphorylation. | 0.1, 1 mg/kg (6 months) | Ageing mice model | [57] |
| Koenidine | Carbazole | GLUT-4 translocation. | 25, 50 µM | In vitro: L6-GLUT4myc myotubes cell line | [58] |
| Lupanine | Quinolizidine | Potentiated insulin release by directly affecting KATP channels. | 0.5 mmol/L 20 mg/kg | In vitro: NS-1E cell line Streptozocin-induced diabetic mice | [59] |
| Magnoflorine | Aporphine | PTP1B inhibition. | 12.5 to 100 μM | In vitro: enzymatic assay and in silico | [55] |
| Neferine | Isoquinoline | Upregulated GLUT-4 expression and plasma membrane fusion. | 150 µM | In vitro: L6 cell line | [60] |
| Nigelladine | Norditerpene | Reduce PTP1B overexpression, promote glycogen synthesis and activated the PI3 K/Akt signaling pathway. | 50 µM | In vitro: L6 cell line | [61] |
| Nuciferine | Aporphine | Insulin secretion stimulator. | 10, 20 mM (24 h) | In vitro: INS1-E cell line | [62] |
| Picrasidine | Cathinone | PTP1B inhibition | IC50: 19.80 ± 0.62 µM | In vitro: Hepatocellular carcinoma (HepG2) cell line | [63] |
| Piperine | Piperidine | Down-regulation of mRNA levels of pro-inflammatory cytokines. | 40 mg/kg (10 weeks) | Monosodium glutamate diabetic mice | [64] |
| Ameliorated dysfunction of β-cell. | 15, 30 mm/kg (8 weeks) | High-fat-induced diet mice | [65] | ||
| Sanguinarine | Benzo-phenanthridine | Activation of AMPK. | 0.2, 1, 10 µM (1 h) | In vitro: cell based assay and in silico | [66] |
| Trigonelline | Pyridine | Reduction of insulin resistance through PAR-γ/GLUT4-leptin/TNF-α signaling pathway. | 40 mg/kg (8 weeks) | High-fat diet and Streptozocin-induced diabetic rats | [67] |
| Protection of β-cells Exhibited antioxidant activity. | 40 mg/kg (8 weeks) | High-fat and high fructose-induced diabetic rat | [68] | ||
| Vindolicine | Indole | PTP1B inhibition and induction of glucose uptake, IC50 was 73.5 ± 11.3 | 12.5, 50 µg/mL (24 h) | In vitro: β-TC6 and C2C12 cell lines | [69] |
| Vindolinine | Indole | PTP1B inhibition and induction of glucose uptake | 12.5, 50 µg/mL IC50: 57.6 ± 10.7 µM (24 h) | In vitro: β-TC6 and C2C12 cell lines | [69] |
| Vindolidine | Indole | PTP1B inhibition and induction of glucose uptake | 12.5, 50 µg/mL IC50: 180.1 ± 19.0 µM (24 h) | In vitro: β-TC6 and C2C12 cell lines | [69] |
| Vindogentianine | Indole alkaloid | PTP1B inhibition and induction of glucose uptake | 12.5, 25, 50, 100 and 200 μg/mL IC50: more than 50 μg/mL (24 h) | In vitro: β-TC6 and C2C12 cell lines | [70] |
8.1.2. Dietary Phenols
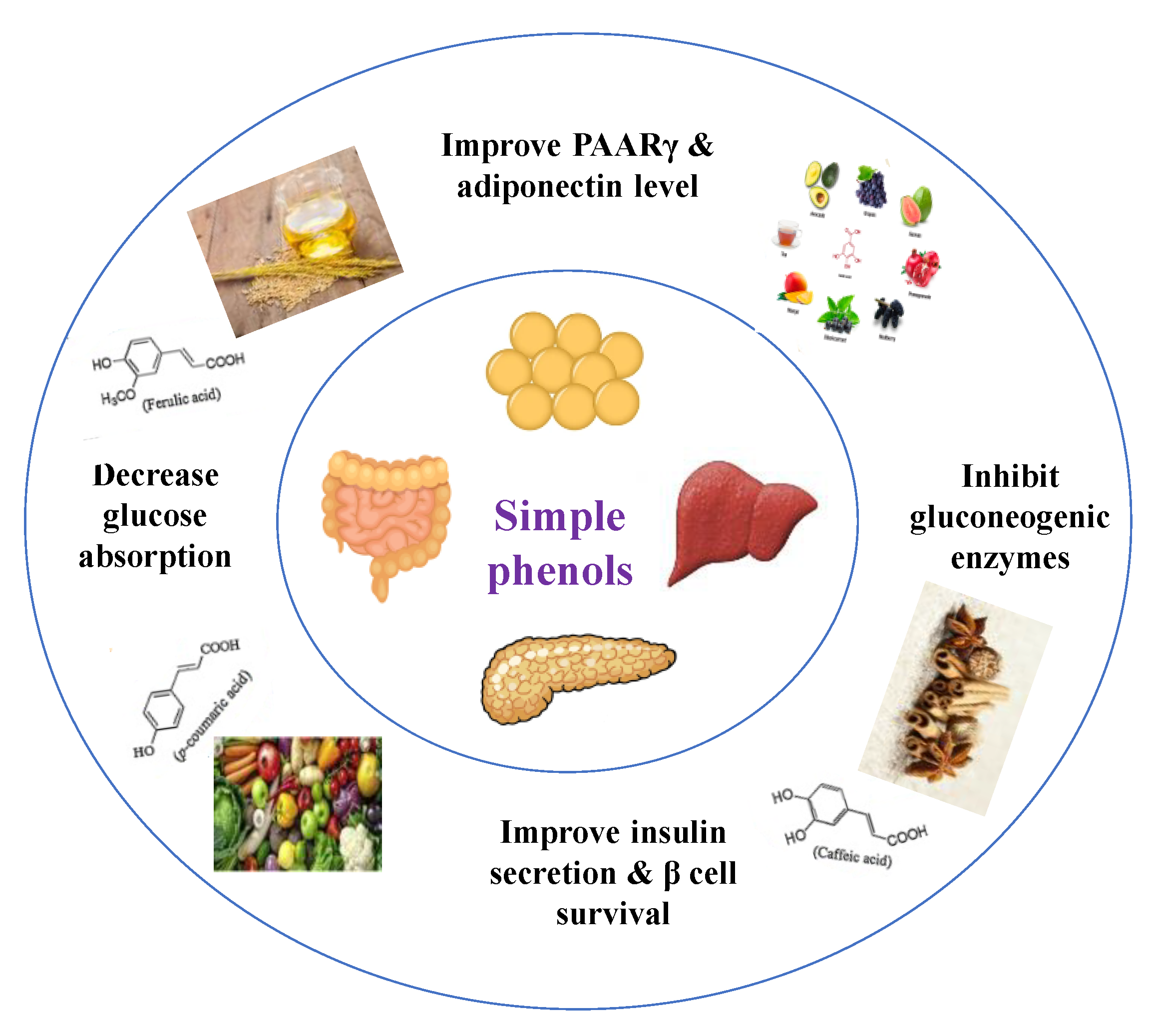
Phenolics are a complex category of bioactive secondary metabolites produced through shikimate and acetate pathways. Phenols are classified into four major groups: simple phenols or phenolic acid, coumarins, lignans, lignins, stilbenes, hydrolyzable and condensed tannins and flavonoids which include flavans, flavonols, flavones, flavanones, isoflavones, and anthocyanins [10,71]. Simple phenols have only one aromatic carboxylic acid with hydroxyl derivatives and two types; hydroxybenzoic acid and hydroxycinnamic acid derivatives. Several simple phenolic acids were assessed clinically and in vivo for their antidiabetic activities. The considerable hypoglycemic activity was shown for several hydroxycinnamic acid derivatives listed in (Table 4) [10].
Dietary phenols have positive health effects in diabetes via protection of pancreatic islet β-cell, reduction of β-cell apoptosis, promotion of β-cell proliferation, attenuation of oxidative stress, activation of insulin signaling, stimulation of pancreas to secrete insulin, inhibition of glucose absorption, regulation of intestinal microbiota, modification of inflammation response, and inhibition of the formation of advanced glycation end-products (Figure 11) [10].
The efficacy of phenols depends primarily on their bioavailability and metabolism after entering the body. Phenolics exhibited a magnificent power to defeat diseases due to their antioxidant potential. They have direct and indirect antioxidant activity by inducing endogenous protective enzymes and positive regulatory effects on signaling pathways. Their antioxidant power depends on the presence of esterified or glycosylated substitution [71,72]. Among all mechanisms reported for phenolic acids to manage diabetes, the best was the inhibition of postprandial hyperglycemia and the antioxidant activity, as diabetes is known as an oxidative stress disorder [71]. Table 5 shows different studied mechanisms of simple phenolics.
In a randomized cross-over trial in healthy overweight men, decaffeinated coffee consumption and chlorogenic acid intake were associated with reducing early glucose and insulin responses during two hours in oral glucose tolerance test (OGTT) [73,74].
Table 4.
Proposed antidiabetic effects of different simple phenols.
| Name of Compound | Chemical Structures | Mechanism | References |
|---|---|---|---|
| Caffeic acid |  |
| [10,75] |
| p-Coumaric acid |  |
| [10,76] |
| Cinnamic acid |  |
| [75,77] |
| Chlorogenic acid |  |
| [75,78] |
| Ellagic acid |  |
| [75,79] |
| Ferulic acid |  |
| [10,77] |
| Gallic acid |  |
| [75,76] |
| Hydroxycinnamic acid |  |
| [10,77] |
| Protocatechuic acid |  |
| [75,80] |
| Syringic acid |  |
| [75,81] |
Table 5.
In vitro, in vivo and in silico models of different isolated simple phenols.
| Compound | Mechanism | Dose of the Tested Compound (Duration) | Model | References |
|---|---|---|---|---|
| Caffeic acid | Exhibited antioxidant effect. | 40 mg/kg (4 weeks) | Nicotinamide-streptozocin-induced diabetic mice | [82] |
| Exhibited antioxidant effect, increased insulin secretion and protected pancreatic cells. | 25, 35 mg/kg (5 weeks) | Streptozocin-induced diabetic rats | [83] | |
| Coumaric acid | Activation of pancreatic GLUT-2, increased level of insulin, decreased gluconeogenic enzymes (glucose-6-phosphatase and fructose-1, 6-bisphosphatase). | 100 mg/kg (4 weeks) | Streptozocin-induced diabetic rats | [84] |
| Decreased the level of TNF-α, increased the levels of PPARγ mRNA and adiponectin. | 40 mg/kg (6 weeks) | Streptozocin-induced diabetic rats | [85] | |
| Exhibited antioxidant effect, increased insulin level, protected pancreatic cells. | 100 mg/kg (4 weeks) | Streptozocin-induced diabetic rats | [86] | |
| Cinnamic acid | Stimulated glucose-induced insulin secretion | 5, 10 mg/kg | Streptozocin-induced diabetic rats | [87] |
| Chlorogenic acid | Increased expression of adiponectin receptors, increased phosphorylation of AMPK in the liver and muscle, increased mRNA and protein levels of PPAR-α in the liver. | 80 mg/kg (12 weeks) | Diabetic mice | [88] |
| Ellagic acid | Activated insulin signaling pathway in the liver by increasing phosphorylated Akt had an antioxidant effect | 50 mg/kg (28 days) | Insulin resistant diabetic rats | [89] |
| Ferulic acid | Improved hepatic glycogenesis by phosphorylating and inhibiting GSK3β, suppressed gluconeogenesis by phosphorylating FoxO1, Reduced IRS1, PKC-ε and PTP1B, which are known to inhibit the insulin signaling. | 50 mg/kg (4 weeks) | High-fat and fructose-induced diet diabetic rat | [90] |
| Reduced GLUT-2 expression. | 50 mg/kg (4 weeks) | High-fat and fructose-induced diet diabetic rat | [91] | |
| Gallic acid | Decreased the level of TNF-α, increased the levels of PPARγ mRNA and adiponectin. | 20 mg/kg (6 weeks) | Streptozocin-induced diabetic rats | [85] |
| Protocatechuic acid | Exhibited antioxidant effect, improved hepatic insulin resistance by modulating IRS1/PI3K/AKT2 pathways. | 100 mg/kg | Streptozocin-induced diabetic rats | [92] |
| Stimulated insulin signaling pathway increasing GLUT4 translocation and glucose uptake. | 1–150 µmol/L | In vitro: human visceral adipocytes | [93] | |
| Attenuated the increase in the expression of gluconeogenic enzymes, restored AKT protein phosphorylation (restores GLUT-4 translocation). | 50, 100 mg/kg (14 days) | Dexamethasone diabetic rats | [94] | |
| Syringic acid | Ameliorated the functional and histological abnormalities and hepatic mitochondria biogenesis (fight insulin resistance). | 25, 50 and 100 mg/kg (6 weeks) | Streptozocin-induced diabetic rats | [95] |
8.1.3. Anthocyanins
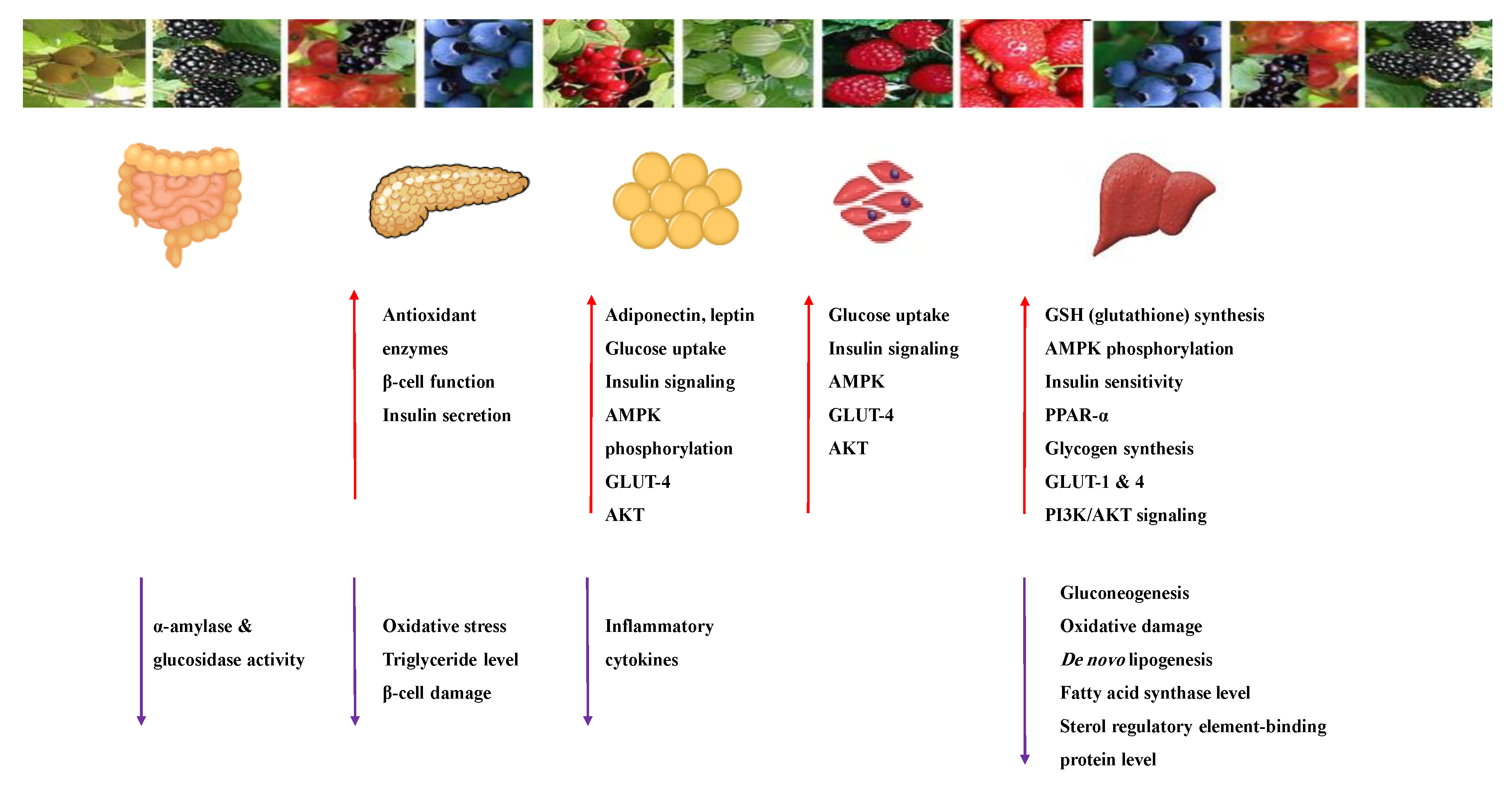
Anthocyanins are widespread natural polyphenolic pigments in the plant kingdom. Six anthocyanidins are commonly synthesized in plants: cyanidin, delphinidin, malvidin, peonidin, pelargonidin and petunidin [96]. Table 6 shows the proposed mechanisms of anthocyanins as antidiabetic agents. They have valuable health effects documented in many in vivo and in vitro studies (Table 7). It appears that they affect several signaling pathways of glucose metabolism (Figure 12) [21,97,98,99].
The antidiabetic effects of anthocyanins are due to their antioxidant and anti-inflammatory properties and their ability to inhibit the α-amylase, α-glucosidase, DPP-IV enzymes and the activity and overexpression of PTP1B protein [41]. The extract and the isolated compounds effectively improved insulin sensitivity and refinement of insulin resistance [100]. On the other hand, anthocyanin-rich foods were tested in type-2 diabetic patients. All studies reported a decrease in insulin resistance and improved lipid profile [100,101].
Anthocyanins keep β-cell viability and induce insulin release and improve insulin sensitivity via downregulation of the adipocytokine retinol-binding protein 4 (RBP4). In addition, they increase GLUT4 in adipose tissue and skeletal muscle through the activation of the AMPK pathway [44,97,100]. They activate AMPK and modulate genes involved in; insulin-glucose signaling pathways, the expression of PPAR γ, adipocyte-specific genes (lipoprotein lipase) and can up-regulate the gene expression of adiponectin and down-regulate plasminogen activator inhibitor-1 and interleukin-6 genes. [96,99,102].
High anthocyanin intake induced a significant reduction in peripheral insulin resistance detected in females aged 18–76 years [103]. The increase in plasma insulin level and the improvement of glycemic control in type 2 diabetic adults were reported after daily consumption of Cornus mas extract for 6 weeks [104,105].
The bioactivities of anthocyanins are different, although they share the same chemical scaffold. The structure–activity relationship influences the bioavailability and bioactivity of anthocyanins. The degree of hydroxylation, methylation, acylation and the sugar moiety attached to make the difference. The in vitro studies showed that the suppression of gut glucose absorption is the major mechanism of anthocyanin anti-diabetic effect through inhibition of α-glucosidase and α-amylase enzymes. The substitution of glucose or galactose enhances the enzyme inhibition activity at the 3-O position or by acylation with ferulic or caffeic acid derivatives [103,105].
The major drawback of anthocyanin use is its poor bioavailability and stability. A new drug design can be used: the encapsulation with protective compounds such as chitosan, cyclodextrin, liposomal micelles or the use of micro or nano-encapsulation. These forms can also offer an extra advantage of controlling the release of the substance. More studies must decide the appropriate consumption and the most effective type as antidiabetic [99,102,103,106].
Table 6.
Proposed antidiabetic effects of different anthocyanins.
| Name of Compound | Chemical Structure | Mechanism | References |
|---|---|---|---|
| Cyanidin |  |
| [41,107] |
| Delphinidin |  |
| [41,106] |
| Pelargonidin |  |
| [41,108] |
Table 7.
In vitro, in vivo and in silico models of different isolated anthocyanins.
| Compound | Mechanism | Dose of the Tested Compound (Duration) | Model | References |
|---|---|---|---|---|
| Cyanidin | Increased intracellular Ca2+ stimulated insulin secretion and the expression of genes involved in this process. | 100 µM | In vitro: β-cells INS-1 | [109] |
| Delphinidin | Inhibited glucose absorption by free fatty acid receptor 1 (FFA1). | 100 µM | In vitro: Caco-2 and HT-29 cells | [110] |
| Inhibited α-amylase Inhibited α-glucosidase. | IC50: 601.56 nM IC50: 268.41 nM | In vitro: enzymatic assay | [111] | |
| Pelargonidin | Inhibited α-amylase Inhibited α-glucosidase. | IC50: 2067.78 nM IC50: 175.04 nM | In vitro: enzymatic assay, in silico | [111,112] |
8.1.4. Flavonoids
Flavonoids are widespread residents in the plant kingdom with approximately 8000 compounds that support plant development, growth and defense. They share 15 carbon skeletons, and according to the modifications on the generic structure, they are classified into six subclasses [16,41]. Flavonoids have been used for their positive health effects, potential biological functions as antioxidants and treating various human diseases, including diabetes [16].
They are considered promising anti-diabetic agents affecting novel targets such as insulin receptors, protein tyrosine phosphatase, peroxisome proliferator-activated receptor-gamma, and adenosine monophosphate-activated protein kinase (Table 8) [113].
They support the regulation of carbohydrate digestion, glucose metabolism in the liver, insulin signaling, insulin secretion, glucose uptake, adipose deposition, β-cell proliferation and apoptosis (Table 9 and Table 10) [41].
Although flavonoids are known for their poor bioavailability, the invention of advanced delivery techniques (microencapsulation, nano-systems, microemulsions, enzymatic methylation) can overcome the problem [41,114]
The optimal human dietary daily consumption level of flavonoids is not confirmed yet, due to the conflicting bioavailability results and diverse molecular structures. However, flavonoids possible genotoxicity and mutagenicity were reported in mammalian experiments. Fortunately, they are not expected to be induced by dietary flavonoid-rich sources [41,114].
Although few clinical trials on humans were conducted to explore the effect of the flavonoids-rich extract on type-2 diabetic patients, data with promising results were noted. In comparison, individual consumption of flavonoids revealed no fruitful impact on diabetes, while the elevated intake of flavonoids in clinical trials showed a decreased risk of developing diabetes. It is necessary to run more clinical trials using isolated flavonoids or defined standardized flavonoid mixtures to explore the efficacy, pharmacokinetics, and safety profiles [16,114].
Recently, a meta-analysis had specified an association between high intake of total flavonoids and the reduced risk of type-2 diabetes in a dose-dependent manner, specifically for anthocyanidins, flavan-3-ols, flavonols, and isoflavones [115].
Alpha-glucosidase inhibition is a promising target for flavonoid antidiabetic activity. Generally, the 3′ position (ring B) favors bulky and minor electron-withdrawing and hydrogen bond donor groups. At position 4′ (ring B), electron-donating and hydrogen bond acceptor groups improve inhibitory activity. In addition, position 7 (ring A) favors bulky and hydrogen bond acceptor groups [113]. More extensive research is needed to relate the flavonoid structure to its activity and efficacy to identify the involved enzymes, receptors, signaling molecules, and transporter or transcription factors.
Table 8.
Antidiabetic effects of flavonoids.
| Glucose Transporter | Hepatic Enzymes | Β-Cells Apoptosis | PPAR | AMPK | Tyrosine Kinase Inhibitor | NF-κB | |
|---|---|---|---|---|---|---|---|
| Target | IRS-1 | G6pase, FD pase PEPCK, G6PD Hexokinase | Bcl-2 family | Gene expression | AMPK | Tyrosine kinase inhibitors | |
 Flavonoid effects  | Activation of IRS-1 | Insulin signaling Liver glycogen | Apoptosis | -------- | -------- | Activity of tyrosine kinase | ------- |
| The synthesis and translocation of GLUT | Hexokinase activity in liver | --------- | Expression of PPARγ | AMPK activation | --------- | Activity of NF-kB |
Table 9.
Proposed antidiabetic effects of different flavonoids.
| Name of Compound | Chemical Structure | Mechanism | References |
|---|---|---|---|
| Apigenin |  |
| [108,116] |
| Baicalein |  |
| [33,117] |
| Chrysin |  |
| [44,118,119] |
| Diosmin |  |
| [108,120] |
| Daidzein |  |
| [108,121] |
| Eriodictyol |  |
| [108,118] |
| Genistein |  |
| [41,121] |
| Fisetin |  |
| [108,119] |
| Kaempferol |  |
| [108,116] |
| Luteolin |  |
| [41,108] |
| Neohesperidin |  |
| [33,122] |
| Naringenin |  |
| [120,123] |
| Naringin |  |
| [110,120,124] |
| Morin |  |
| [108,116] |
| Quercetin |  |
| [35,110,125] |
| Rutin |  |
| [35,44,110] |
| Tangeretin |  |
| [116,126] |
Table 10.
In vitro, in vivo and in silico models of different isolated flavonoids.
| Compound | Mechanism | Dose of the Tested Compound (Duration) | Model | References |
|---|---|---|---|---|
| Apigenin | Exhibited free radical scavenger and a regulator activity to antioxidant defenses of pancreatic cells. | * ND | In silico | [127] |
| Inhibited DPP-4 enzyme. | 1.5 mg/kg for every alternate (4 weeks) | High-fat and high fructose-induced diabetic rats, in silico | [124] | |
| Baicalein | Suppressed hepatic gluconeogenesis via activation of the AMPK and AKT signaling pathways. | 12.5 mM (24 h) | In vitro: HepG-2 cell line | [128] |
| Suppressed expression of PGC-1α (upregulate hepatic gluconeogenic gene expression) and gluconeogenic genes ameliorated hepatic insulin resistance and gluconeogenic activity by inhibiting the p38 MAPK/PGC-1α signal pathway. | 50 mg/kg (21 days) 100 µM (12 h) | High-fat diet-induced insulin-resistant mice Primary hepatocytes | [125] | |
| Promoted glucose uptake through enhancement of GLUT4, PGC-1α, MAPK, AKT and contents. | 100, 200, 400 μM (6,12, 24 h) | In vitro: L6 myoblast cell line, C2C12 cell line, animal model | [129,130] | |
| Chrysin | Modified oxidative stress. | 20, 40, 80 mg/kg | Streptozocin-induced diabetic rats | [131] |
| Activated insulin signal transduction such as IR, IRS-1, Akt | 100 mg/kg (4 weeks) | High fat-induced diet diabetic rats | [132] | |
| Eriodictyol | Exerted glucose-dependent insulinotropic effect through cAMP/PKA pathway. | 200 μM | In vitro: mice islets and MIN6 cell line | [133] |
| Activated glucose utilization, suppressed gluconeogenesis, decreased pro-inflammatory cytokines and increased anti-inflammatory cytokine. | 0.05% (w/w) (16 weeks) | Animal model | [134] | |
| Genistein | Increased insulin level, regenerated β-cells. | 10, 20 mg/kg (4 weeks) | Streptozocin-induced diabetic rats | [135] |
| Inhibited α-amylase Inhibited α-glucosidase. | IC50: 165.51 nM IC50: 1394.36 nM | In vitro: enzymatic assay | [111] | |
| Fisetin | Inhibited high glucose-induced reactive oxygen radical production through the activation of SIRTs and FOXO3a. | 3, 5 and 10 μM (48 h) | In vitro: THP-1 cell line | [136] |
| Hesperidin | Had antioxidant effect, protective effect for β-cells. | 100 mg/kg (15 days) | Streptozocin-induced diabetic rats | [137] |
| Decreased oxidative stress and NF-kB levels and increased while SIRT1 level. | 100 mg/kg (15 days) | Streptozocin-induced diabetic rats | [138] | |
| Improved glycogen content by reinstating the activities of glycogen synthase and glycogen phosphorylase. | 25, 50, 100 mg/kg (4 weeks) | Streptozocin-induced diabetic rats | [139] | |
| Luteolin | Inhibited high glucose-induced reactive oxygen radical production through the activation of SIRTs and FOXO3a. | 3, 5 and 10 μM (48 h) | In vitro: THP-1 cell line | [136] |
| Morin | Improved insulin signaling through inhibition of microRNA-29a (an essential regulator of insulin signaling and gluconeogenesis pathways). | 50 µM (24 h) | In vitro: HepG-2 cell line | [140] |
| Naringenin | Exhibited antioxidant and anti-inflammatory effects. | * ND | In silico | [127] |
| Decreased oxidative stress through promoting nuclear factor E2-related factor 2 (Nrf2), restored insulin expression, promoted glycolysis while inhibiting gluconeogenesis. | 50 mg/kg (45 days) | In vitro: MIN6 cell line, streptozocin-induced diabetic rats | [141] | |
| Improved mRNA expressions of insulin receptor b subunit, GLUT4 and adiponectin. | 100 mg/kg (4 weeks) | Streptozocin-induced diabetic rats | [142] | |
| Naringin | Improved mRNA expressions of insulin receptor b subunit, GLUT4 and adiponectin. | 100 mg/kg (4 weeks) | Streptozocin-induced diabetic rats | [142] |
| Inhibited both intrinsic and extrinsic pathways of β-cell apoptosis, possibly by interfering with DNA damage- and cytokine-induced apoptotic signaling by suppressing pancreatic reactive oxygen species accumulation and leukocyte infiltration. | 50, 100 mg/kg (2 weeks) | Streptozocin-induced diabetic rats | [143] | |
| Quercetin | Activated AMPK- MAPK pathway to induce glucose uptake. | 10, 100 µM (24 h) | In vitro: L6 myoblast cell line | [144] |
| Promoted hepatic glycogen synthesis and reduced blood glucose by increasing Akt phosphorylation, GSK-3 phosphorylation, and GCK protein expression levels. | 10, 50 mg/kg (12 weeks) | Streptozocin-induced diabetic rats | [145] | |
| Exhibited antioxidant effect, protective effect for β-cells. | 100 mg/kg (15 days) | Streptozocin-induced diabetic rats | [137] | |
| Decreased oxidative stress and NF-kB levels and increased while SIRT1 level. | 100 mg/kg (15 days) | Streptozocin-induced diabetic rats | [138] | |
| Tangeretin | Exhibited antiapoptotic property due to its inhibitory effect on oxidative stress. | 0, 10, 20, and 40 μM (12 h) | In vitro: INS-1 cell line | [146] |
| Exhibited a potential insulin action enhancer that functions by inhibiting the MEK-ERK1/2 pathway in hepatocytes. | 10,20 mM (48 h) 25, 50 mg/kg (1 month) | Animal model | [147] | |
| Inhibited α-glucosidase Inhibited α-amylase. | IC50 285.88 nM IC50 682.75 nM | In vitro: enzymatic assay | [111] |
ND * not determined.
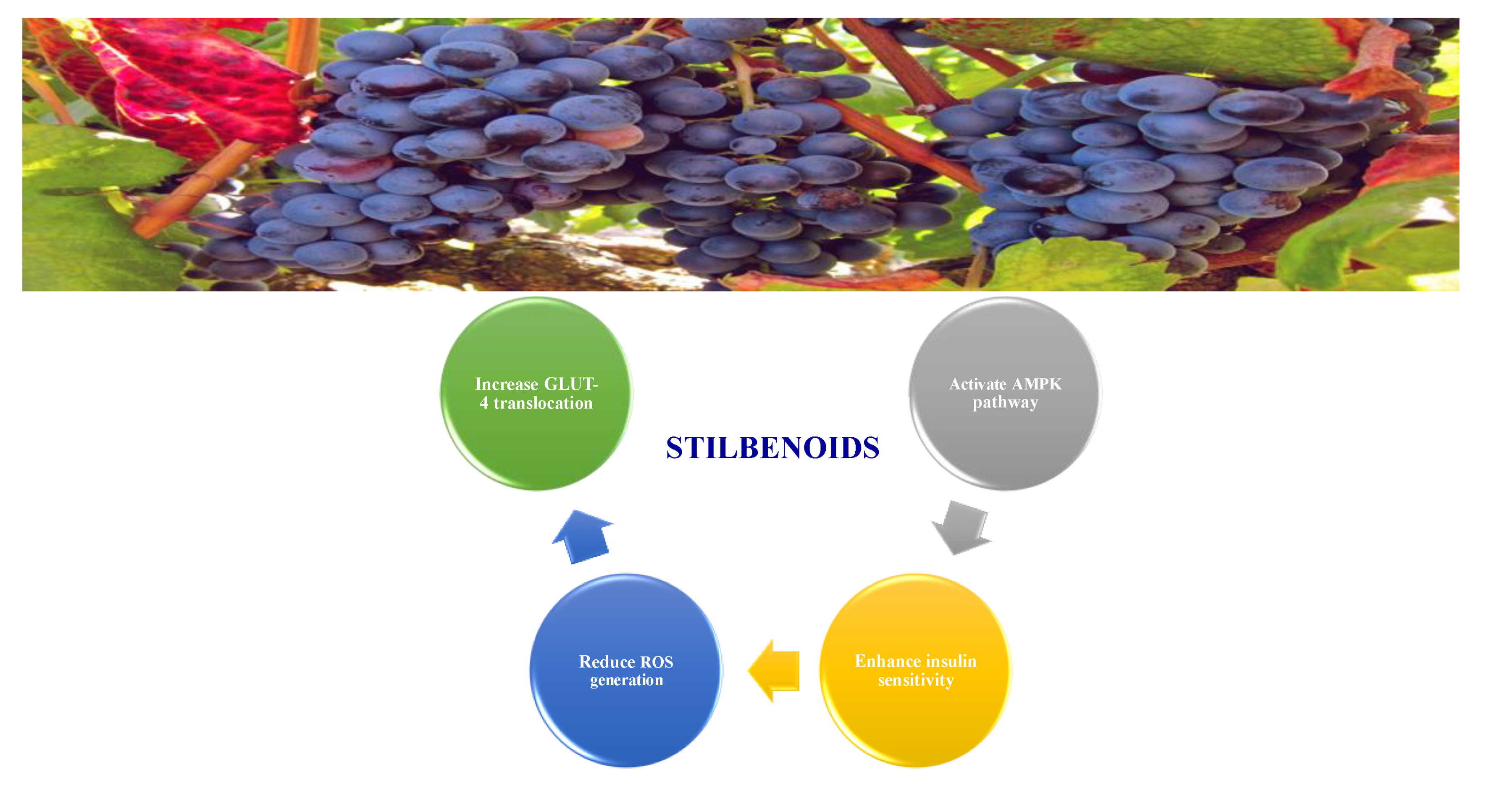
8.1.5. Stilbenoids
Stilbenoids are antimicrobial compounds produced de novo to protect plants from fungal infection and toxins [148]. They share the same backbone (C6-C3-C6) with different substitutions on the aromatic rings. Dietary stilbenoids are rarely reported as antidiabetic agents except for resveratrol, pterostilbene, and polydatin (Figure 13) [10,148]. Table 11 summarizes the hypoglycemic effect of some dietary stilbenoids. Table 12 shows the studied antidiabetic mechanisms of stilbenoids.
8.1.6. Saponins
Saponins are bioactive steroidal or triterpenoid glycosides with numerous biological activities and potent antihyperglycemic activity (Figure 14) [15,33].
The antihyperglycemic activity of saponins from various plants was confirmed in vivo and in vitro models [153]. They reduce hyperglycemia through several proposed mechanisms: restoration of insulin response, improvement in insulin signaling, increase plasma insulin levels, induction of insulin release from the pancreas, activation of glycogen synthesis, inhibition of gluconeogenesis, inhibition of α-glucosidase activity, inhibition of mRNA expression of glycogen phosphorylase and glucose-6-phosphatases, increase the expression of Glut4 (Table 13) [153].
Astragaloside IV significantly decreases blood glucose by attenuating insulin resistance in adipocyte cells and inhibiting glycogen phosphorylase and G6Pase [33].
The tetracyclic triterpene structure is pivotal for the antidiabetic activity of saponins. Ginsenosides, from Ginseng species, were extensively studied for their hypoglycemic activity. Several animal models and cell lines studies have documented the beneficial effect of ginsenoside through affecting several signaling pathways of glucolipid metabolism. On the other hand, several clinical studies were conducted using ginseng extracts. The results were conflicting. To extend the therapeutic effects, SAR, pharmacokinetic and quality control should be investigated. Table 14 shows studied mechanisms of saponins [149,154,155].
Table 13.
Proposed antidiabetic effects of different saponins.
| Name of Compound | Type of Saponin | Chemical Structure | Mechanism | References |
|---|---|---|---|---|
| Arjunolic acid | Triterpene saponins |  |
| [153,156] |
| Astragaloside IV | Steroidal saponins |  |
| [33,153] |
| Diosgenin | Steroidal saponins |  |
| [157,158] |
| Platyconic acid | Triterpene saponin |  |
| [153,157] |
| Ginsenoside K | Steroidal saponin |  |
| [33,159] |
| Ginsenoside Rg3 | Steroidal saponin |  |
| [33,160] |
| Ginsenoside Rg1 | Steroidal saponin |  |
| [33,161] |
8.1.7. Tannins
Tannins are complex polyphenolic compounds of various molecular weights. They are classified as hydrolyzable and condensed tannins. Hydrolyzable tannins are esters of polyol carbohydrate and gallic acid or ellagic acid. Condensed tannins (proanthocyanidins) are oligomers or polymers of polyphenolic bioflavonoids, specifically taking the form of polyhydroxy flavan-3-ol units and flavan-3,4-diols [169,170].
They have several benefits to health, such as free radicals’ scavengers and as activators to antioxidant enzymes. Their benefits encompass the control and management of chronic diseases, including diabetes. They enhance insulin-signaling pathways, such as PI3K (phosphoinositide 3-kinase), p38 MAPK (mitogen-activated protein kinase) activation and GLUT-4 translocation leading to increase glucose uptake [169]. In addition, they reduce the intestinal absorption of glucose and other nutrients, induce β cell regeneration, inhibit α- amylase and α-glucosidase activity and enhance insulin activity on adipose cells (Table 15) [169,170].
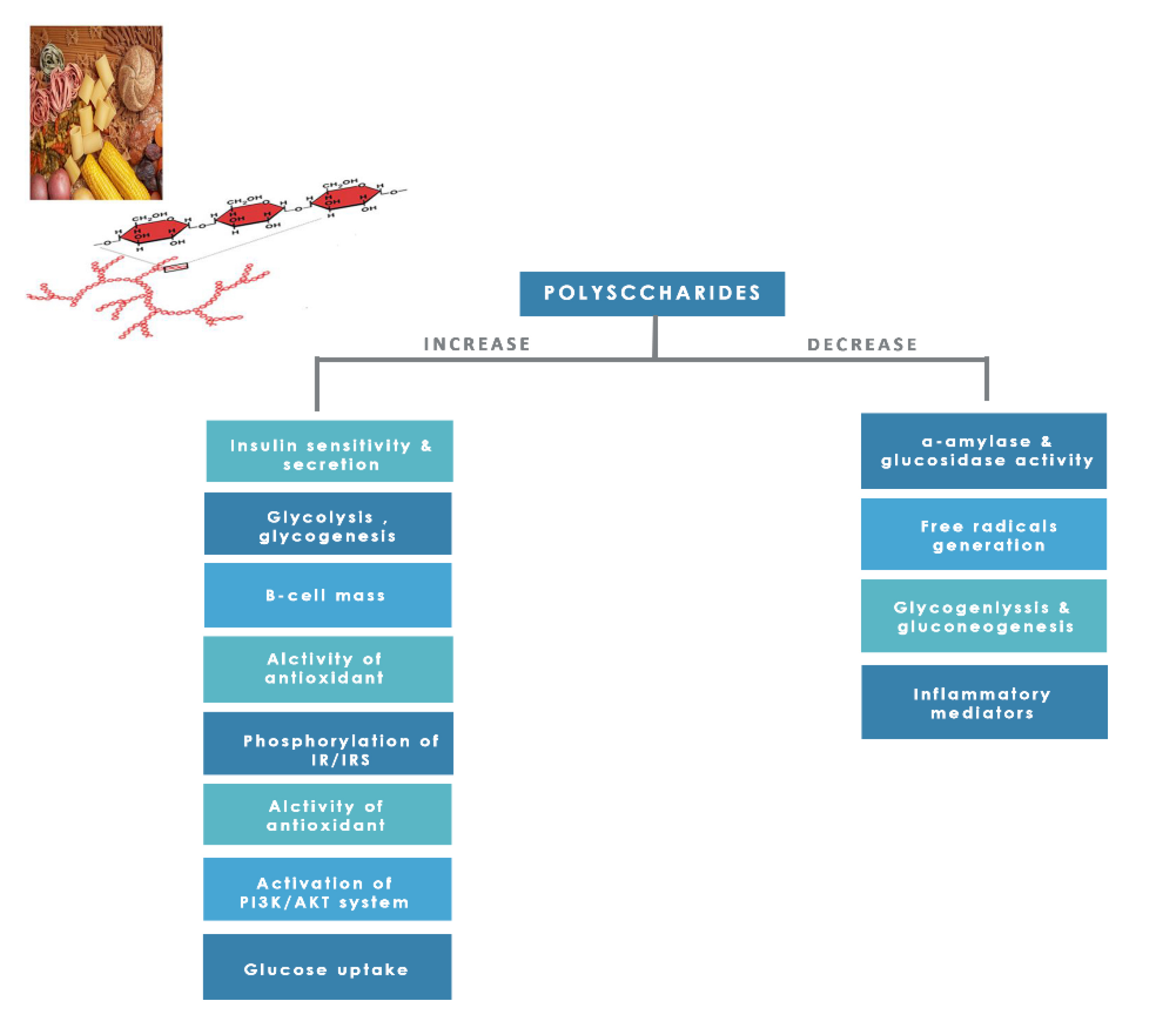
8.1.8. Polysaccharides
Dietary polysaccharides are essential edible compounds for our day-to-day life obtained from plants, grains, fruits, vegetables, and edible mushrooms. Investigators studied them for their numerous pharmacological activities and low toxicity [25]. Polysaccharides showed promising positive effects on human health, such as anticancer, anti-inflammatory action, skin protection, antioxidant, immune modulation, serum cholesterol reduction, and managing diabetes and reducing its complications through several mechanisms (Figure 15) [25,174].
8.1.9. Coumarins
Coumarins are secondary metabolites used widely for their anticoagulation and antithrombotic effects (Figure 16). Abundant in vitro and in vivo studies demonstrated that different coumarin skeletons (simple, furanocoumarin, pyranocoumarin, bicoumarin, and tricoumarin) were good candidates’ against diabetes. Studies have proven that coumarins also exhibit antidiabetic activity through regulating hepatic enzymes, repairing pancreatic β-cells damage, improving insulin signaling, and providing anti-inflammatory and antioxidative properties [157]. Table 16 shows the proposed antidiabetic effects of different coumarins, while Table 17 displays studied antidiabetic mechanisms of coumarins.
Pharmacokinetic profile studies and clinical trials are warranted prior to their use as safe antidiabetics [157]. To enhance their pharmacological potency or pharmacokinetic profile, hybridization between coumarins and other therapeutic pharmacophores opened a new drug method to develop novel hypoglycemic coumarins derivatives. For example, a hybrid molecule of apigenin and coumarin revealed more favorable antidiabetic activity than either compound alone [175].
Table 16.
Proposed antidiabetic effects of different coumarins.
| Name of Compound | Chemical Structure | Mechanism | References |
|---|---|---|---|
| Esculetin |  |
| [157,176] |
| Fraxetin |  |
| [157,176] |
| Umbelliferone |  |
| [177] |
Table 17.
In vitro, in vivo and in silico models of different isolated coumarins.
| Compound | Mechanism | Dose of the Tested Compound (Duration) | Model | References |
|---|---|---|---|---|
| Esculetin | Improved insulin resistance by increasing hepatic GLUT2 and glucokinase mRNA levels and decreased glucose-6-phosphatase mRNA level. | 0.02%, w/w (12 weeks) | C57BL/6J mice high-fat diet diabetic mice, liver histological model | [178] |
| Boosted Akt activation and promoted glucose uptake. | 40 mg/kg (14 days) | Dexamethasone-induced insulin resistance mice, C2C12 cell line | [179] | |
| Fraxetin | Boosted Akt activation and promoted glucose uptake. | 40 mg/kg (14 days) | Dexamethasone-induced insulin resistance mice, C2C12 cell line | [179] |
| Osthole | Increased GLUT4 mRNA expression in skeletal muscle. | 5–10 mg/kg (6 weeks) | High-fat and high-sucrose induced fatty liver with IR rats | [180] |
| Boosted Akt activation and promoted glucose uptake. | 20 mg/kg (14 days) | Dexamethasone-induced insulin resistance mice, C2C12 cell line | [179] | |
| Scopoletin | Stimulated GLUT-4 translocation through activation PI3K and AMPK pathway. | 1, 2.5, 5, 10, 15, 20 (24 h) | 3T3-L1 adipocyte cell lines | [181] |
| Inhibited carbohydrate digestive enzymes. Inhibited α-amylase Inhibited α- glucosidase | IC 50: 37.36 µM IC 50: 85.12 µM | In vitro study | [182] | |
| Umbelliferone | Stimulated muscle glucose uptake and stalled gluconeogenesis and oxidative stress. | 30–240 µg/mL (2 h) | Ex vivo: isolated psoas muscles | [177] |
| Shunted gluconeogenic enzymes, regeneration of the β-cells. | 100 mg/kg (4 weeks) | Alloxan-induced diabetic rat | [183] |
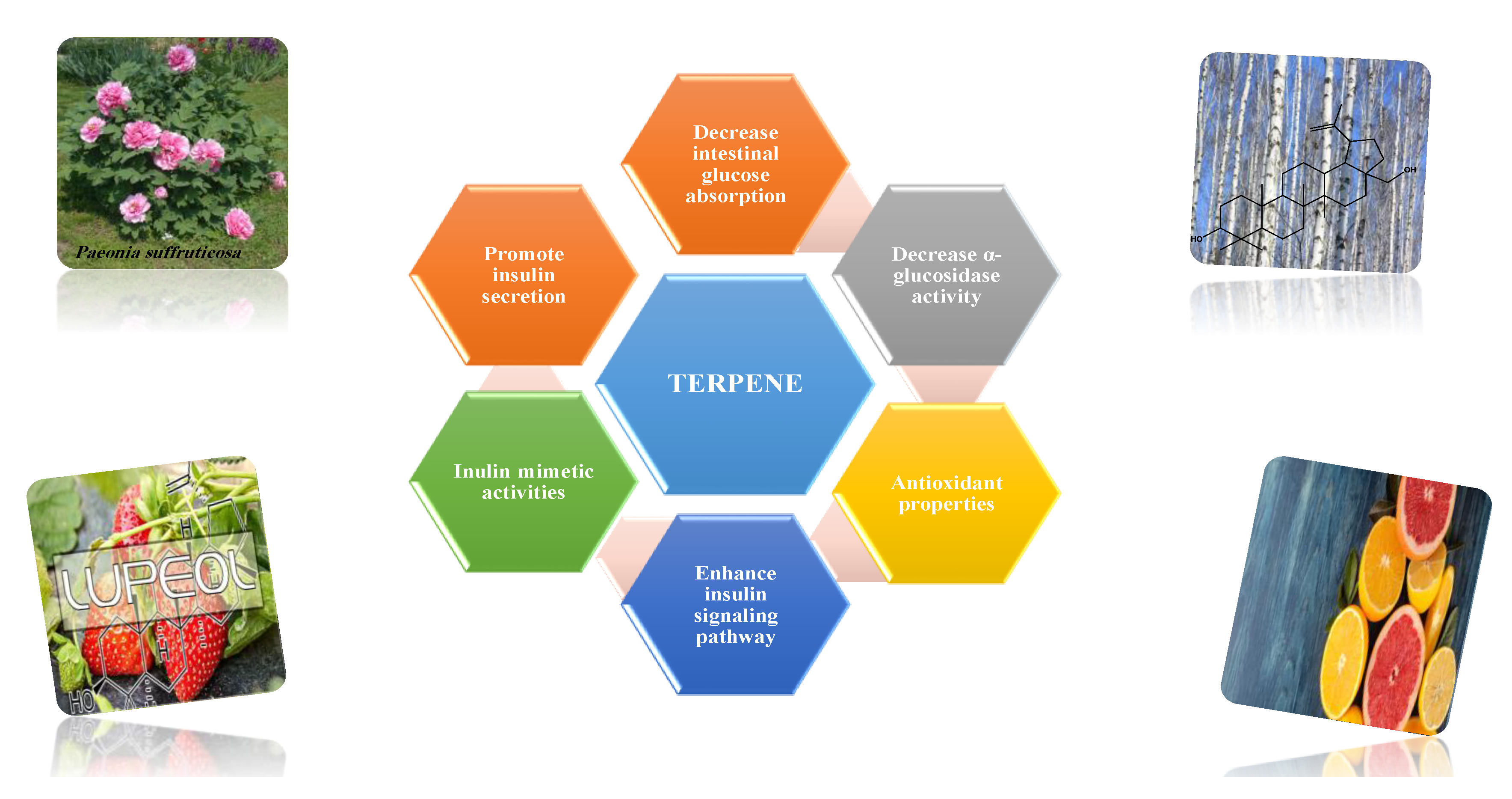
8.1.10. Terpenes
Terpenoids are secondary metabolites constructed from repeating units of isoprene with wide structural diversity [184]. They are categorized into hemiterpenes, monoterpenes, sesquiterpenes, diterpenes, sesterpenes, triterpenes, tetraterpenes, and polyisoprenes (Figure 17). Terpenes and their derivative prevent various diseases and have many valuable properties [185].
Generally, terpenes showed an antidiabetic effect in both in vitro and in vivo studies. They have excellent antioxidant activity and potent α-glucosidase inhibitory activity except for the monoterpenes lacking the phenolic nature in their structures. In addition, they enhanced insulin level and glucose uptake in tissues and can inhibit several signaling pathways in carbohydrate metabolism [184,185]. Several monoterpenes can facilitate glucose uptake via upregulation of the glucose transporter (GLUT4) translocation, enhance insulin signaling pathway, promote insulin secretion, protect pancreatic cells and ameliorate proinflammatory cytokines [184]. In addition, they increase the level of glycogen levels, modulate glucose-6-phosphatase and fructose-1,6-bisphosphatase, reduced glucokinase and glucose-6-phosphate dehydrogenase activities [33,184]. Table 18 shows proposed antidiabetic effects of different terpenes, and Table 19 shows studied antidiabetic mechanisms of terpenes.
Sesquiterpenes are efficacious than other terpenes due to their tightly packed structure. On the contrary, triterpenes with more hydrophilic groups are more preferable to multi-target effects as antidiabetics [185].
Some terpenes were evaluated clinically, but further randomized studies are needed to confirm the declared antidiabetic activity. Additionally, evaluation for pharmacokinetic parameters and safety profiles are needed [185].
Table 18.
Proposed antidiabetic effect of different terpenes.
| Name of Compound | Class of Compound | Chemical Structure | Mechanism | References |
|---|---|---|---|---|
| Bassic acid | Triterpene |  |
| [185] |
| Limonene | Monoterpene |  |
| [185,186] |
| Stevioside | Diterpene |  |
| [185,187] |
| Rebaudioside | Diterpene |  |
| [185,188] |
| Lupeol | Triterpene |  |
| [185,189] |
| Palbinone | Triterpene |  |
| [185,190] |
| Betulin | Triterpene |  |
| [50,190,191] |
Table 19.
In vitro, in vivo and in silico models of different isolated terpenes.
| Compound | Mechanism | Dose of the Tested Compound (Duration) | Model | References |
|---|---|---|---|---|
| Asiatic acid | Exhibited antioxidant activity Increased insulin secretion in rats with sufficient insulin-secreting function Enhanced glucose uptake into skeletal muscle via PI3K-Akt signaling pathway. | 20 mg/kg (45 days) | Streptozocin-induced diabetic rats | [192] |
| Carvacrol | Increased activity of hexokinase, citrate synthase and 6-phosphofructokinase. | 10, 20 mg/kg (4, 6 weeks) | Streptozocin-induced diabetic mice | [193] |
| Limonene | Exhibited antioxidant activity. | 100 mg/kg (8 weeks) | Alloxan-induced diabetic rats | [194] |
| Stevioside | Increased level of GLUT-4. | 1–100 µM (24 h) | Rat L6 myoblast and mouse 3T3-L1 fibroblast cell lines | [195] |
| Increased leptin level Exhibited antioxidant activity, Restored normal pancreatic cell function Increased pyruvate kinase expression and insulin receptor substrate-1. | 300 mg/kg (4 weeks) | Streptozocin-induced diabetic rats | [196] | |
| Lupeol | Shunted gluconeogenic enzymes regenerated o β-cells. | 200 mg/kg (4 weeks) | Alloxan-induced diabetic rat | [183] |
| Inhibited carbohydrate digestive enzyme. | 10 mg/kg | Streptozocin-induced diabetic mice | [191] | |
| Regulated insulin receptor and GLUT-4 protein expression in muscular tissue. | 25 mg/kg (30 days) | High-fat diet and sucrose-induced diabetic mice | [197] | |
| Lupeol | Controlled insulin signaling molecules such as IR and GLUT2 protein expression in hepatocytes. | 25 mg/kg (One month) | High-fat diet diabetic rats | [198] |
| Betulinic acid | Exhibited pancreatic islet regenerative effects. | 10,20, 40 mg/kg (2 weeks) | Streptozocin-nicotinamide-induced diabetic mice | [199] |
| Improved the level of leptin and adiponectin. | 10, 20, 40 mg/kg | Streptozocin-nicotinamide-induced diabetic mice | [200] | |
| Enhanced AMPK phosphorylation, stimulated mRNA expression of glucose transporter 4. | 200 mg/kg | Alloxan- induced diabetic rats | [49] | |
| Inhibited α-glucosidase. | IC50: (1.06 ± 0.02) ×10−5 mol/L | In vitro: enzymatic assay | [201] | |
| Increased basal glucose uptake. | 5, 10 µM (4 days) | HepG2 and 3T3-L1 cell line | [202] | |
| Oleanolic acid | Inhibited α-glucosidase. | IC50: 10.11 ± 0.30 µM | In vitro: enzymatic assay | [203] |
| Ursolic acid | Increased GLUT-4 translocation, increased muscle glycogen content, increased insulin secretion | 0.1, 1 and 10 mg/kg | Hyperglycemic rats | [204] |
| Thymol | Exhibited Antioxidant activity. | 40 mg/kg (28 days) | Streptozocin-induced diabetic rats | [205] |
9. Summary
The management of diabetes is a complicated issue. There is a struggle to lower blood glucose, decrease insulin levels and prohibit insulin resistance. Diabetes is a multifactorial disease, requiring multifactorial strategies to be treated. Although oral antidiabetics and insulin are the keystones for treatment, many obstacles necessitate the investigation for other opportunistic medications. A gastrointestinal burden such as stomach pain and diarrhea are common side-effects for α-glucosidase inhibitors. Thiazolidinediones may cause weight gain, heart failure, and anemia. Other effects, including headache, hypoglycemia, dizziness, nausea and vomiting, fatigue, dyspepsia and genitourinary tract infections, have also been accounted for by conventional anti-diabetic medications. Today, to reverse disease progression by finding a newer drug with novel targets is the main goal to treat diabetes efficiently [42,206,207,208].
Nature is always the savior; animals, microorganisms, marine life and plants all are sources of efficacious and safer drugs. Prospectively, applying natural products in various forms in the healthcare system is extraordinarily bright and urgent. Plant-based medicine is popular, effective, safe, available, and less expensive, serves as a template for drugs and helps detecting new potential targets to treat the disease. In addition, natural products act by affecting multiple targets simultaneously [100,209].
Through various mechanisms of action, plant extract such as Allium cepa, Aloe vera, Alium sativum, Catharanthus roseus, Eucalyptus globules, Trigonella foenum-graecum, Urtica dioica, and isolated molecules such as berberine, ginsenosides, curcumin, stevioside, simple phenolic compounds, anthocyanins, resveratrol, genistein and hesperidin are used for remediation of diabetes [210,211]. Asteraceae, Euphorbiaceae, Fabaceae, Lamiaceae, Liliaceae, and Poaceae are rich in anti-hyperglycemic plant genera [210].
Plant secondary metabolites were reported to inhibit carbohydrate metabolizing enzymes, possess kinase activating capacity, thereby affecting all the metabolic pathways of carbohydrate, lipid and protein, and can intervene in the insulin-signaling pathway, inflammatory response, and oxidative stress and restore molecular aberrations leading to insulin resistance and glucose intolerance [22,27].
The phenolic extracts and pure polyphenols may have different effects on the activity of α-amylase and α-glucosidase. For instance, anthocyanins and flavonols have in vitro α-glucosidase inhibition activity, while flavan-3-ols are responsible for α-amylase inhibitory activity. Simple phenolic acid possesses both in vitro α-glucosidase and α-amylase inhibition. The in vitro antidiabetic activity of saponins is affected by the configuration of the C-23 on the side chain and the type of the molecule attached at the C-3. Moreover, polyphenols can inhibit both enzymes in vivo and influence glucose transporter (SGLT1 and GLUT2), augmenting the postprandial hyperglycemia reduction [212,213].
On the contrary, polyphenol can promote glucose uptake by functional tissues through GLUT4 transporter. Berberine, gallic acid, daidzein, resveratrol and vanillic acid can accelerate glucose transportation by AMPK and PI3K-Akt dependent pathways [213,214].
Enhancement of insulin failure to trigger signal transduction in tissues and the restoring of insulin sensitivity by polyphenols were reported to be mediated by activating the AMPK pathway in many in vivo and in vitro studies. Recently, AMPK was proposed as a major drug target to treat diabetes [20,103]. Its activation prompted upregulation of GLUT-4 and enhancement of glucose uptake, reduction in hepatic glucose production and increase fatty acid oxidation. It had been reported that triterpenes stimulate glucose uptake and glycogen synthesis through AMPK activation [49,97].
Recently, PTP1B was considered a potential target for diabetes. As selective inhibitors of PTP1B, ertiprotafib, trodusquemine and claramine reached clinical trials. Trodusquemine is still in phase 2 clinical trials, while ertiprotafib clinical trials were terminated due to the lack of significant clinical efficacy and undesirable side effects. Alkaloids, flavonoids, phenolic acids, steroids, tannins and terpenes can all inhibit the negative regulation of PTP1B [35,36].
Apigenin, kaempferol, luteolin and quercetin are already known PTP1B inhibitors [215,216]. The screening results strongly revealed the important pattern of substitutions on the main flavonoid skeleton. A hydroxyl group on position 3 (ring C), methoxy groups at 3՝ and 4՝ positions (ring B) and the presence of a less polar group at 7 and 8 positions (ring A), arrangement number and characterization of sugar moiety can affect the activity. Flavonols were reported to possess a potent PTP1B inhibitory effect depending on multiple non-polar substituents. In addition, flavanones were described as significant PTP1B inhibitors. They were associated with the presence of a hydroxyl group at position 5 of ring A and the presence of the isoprenyl group [216,217].
Targeting pancreatic β-cells is a new source for successful antidiabetic agents. For example, berberine and quercetin showed a regeneration power of pancreatic β-cells. Oleanolic acid glycoside, geraniol, kaempferol had an anti-apoptosis effect. Resveratrol increased insulin secretion, and vidoline protects β-cells from cytokines induced apoptosis.
Antioxidant power is a preliminary factor for the prediction of antidiabetic activity [218]. NADPH oxidase inhibitors—such as anthocyanins, xanthine oxidase inhibitors—such as catechin and gallic acid, and SOD mimetics can neutralize free radical generation [219,220,221,222].
In summary, secondary metabolites are excellent research candidates to reduce blood glucose levels as they are digestive enzyme inhibitors, insulin secretagogue, insulin sensitizers, insulin signal transductors and insulin modulators.
10. Conclusions
This review highlighted the scientific evidence of the plant-derived secondary metabolites as potential multi-target antidiabetic agents. They can modulate enzymes, proteins, signaling pathways, cell proliferation, inflammatory mediators, gene transcription and expression. Numerous compounds are promising antidiabetic agents; however, large scale anti-diabetic evaluation is needed. Therefore, it is important to advocate further research to validate and optimize targets, animal models, toxicity, efficacy, pharmacokinetic studies and clinical trials for multi-target hypoglycemic therapy. Computer-aided research may ease the road for the development of more potent and selective plant-based antidiabetic molecules.
Author Contributions
M.B.S. and A.M.A.-O. equally contributed to the writing of the manuscript; G.A.Y.R.S. envisioned the review. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been carried out during sabbatical leave granted to the author Mayadah Shehadeh from the University of Jordan during the academic year 2019–2020.
Institutional Review Board Statement
IRB statement was waived for this review, which does not report new experimental results obtained by authors, but a compilation from published data.
Informed Consent Statement
Not applicable.
Acknowledgments
The authors would like to acknowledge the central library staff at the University of Jordan for their help in acquiring some of the references cited in this manuscript. We are grateful for the three anonymous reviewers who provided useful comments and suggestions on an earlier draft of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, F.-S.; Weng, J.-K. Demystifying traditional herbal medicine with modern approach. Nat. Plants 2017, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dar, R.A.; Shahnawaz, M.; Rasool, S.; Qazi, P.H. Natural product medicines: A literature update. J. Phytopharmacol. 2017, 6, 340–342. [Google Scholar]
- Lahlou, M. The success of natural products in drug discovery. Pharmacol. Pharm. 2013, 4, 17–31. [Google Scholar] [CrossRef] [Green Version]
- Bernardini, S.; Tiezzi, A.; Laghezza Masci, V.; Ovidi, E. Natural products for human health: An historical overview of the drug discovery approaches. Nat. Prod. Res. 2018, 32, 1926–1950. [Google Scholar] [CrossRef]
- Suaifan, G.A.R.Y.; Shehadeh, M.B.; Darwish, R.M.; Al-Ijel, H.; Abbate, V. Design, synthesis and in vivo evaluation of novel glycosylated sulfonylureas as antihyperglycemic agents. Molecules 2015, 20, 20063–20078. [Google Scholar] [CrossRef] [Green Version]
- Verma, S.; Gupta, M.; Popli, H.; Aggarwal, G. Diabetes mellitus treatment using herbal drugs. Int. J. Phytomed. 2018, 10, 1–10. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Verma, R.; Gupta, J. Challenges and future prospects of herbal medicine. Int. Res. Med. Health Sci. 2018, 1, 12–15. [Google Scholar] [CrossRef]
- Rosado-de-Castro, P.H.; Pimentel-Coelho, P.M.; Barbosa da Fonseca, L.M.; de Freitas, G.R.; Mendez-Otero, R. The rise of cell therapy trials for stroke: Review of published and registered studies. Stem Cells Dev. 2013, 22, 2095–2111. [Google Scholar] [CrossRef] [Green Version]
- Chauhan, D.S.; Gupta, P.; Pottoo, F.H.; Amir, M. Secondary metabolites in the treatment of diabetes mellitus: A paradigm shift. Curr. Drug Metab. 2020, 21, 493–511. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhao, C.; Guven, E.C.; Paoli, P.; Simal-Gandara, J.; Ramkumar, K.M.; Wang, S.; Buleu, F.; Pah, A.; Turi, V. Dietary polyphenols as antidiabetic agents: Advances and opportunities. Food Front. 2020, 1, 18–44. [Google Scholar] [CrossRef] [Green Version]
- Ehrenkranz, J.R.; Lewis, N.G.; Ronald Kahn, C.; Roth, J. Phlorizin: A review. Diabetes Metab. Res. Rev. 2005, 21, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Cao, J.; Zhao, T.; Liu, Y.; Khan, A.; Cheng, G. The bioavailability, extraction, biosynthesis and distribution of natural dihydrochalcone: Phloridzin. Int. J. Mol. Sci. 2021, 22, 962. [Google Scholar] [CrossRef]
- Kovacich, N.; Chavez, B. Ertugliflozin (steglatro): A new option for SGLT2 inhibition. Pharm. Ther. 2018, 43, 736. [Google Scholar]
- Triantakonstanti, V.V.; Mountanea, O.G.; Papoulidou, K.-E.C.; Andreou, T.; Koftis, T.V.; Gallos, J.K. Studies towards the synthesis of ertugliflozin from l-Arabinose. Tetrahedron 2018, 74, 5700–5708. [Google Scholar] [CrossRef]
- Bharti, S.K.; Krishnan, S.; Kumar, A.; Kumar, A. Antidiabetic phytoconstituents and their mode of action on metabolic pathways. Ther. Adv. Endocrinol. Metab. 2018, 9, 81–100. [Google Scholar] [CrossRef]
- Alkhalidy, H.; Wang, Y.; Liu, D. Dietary flavonoids in the prevention of T2D: An overview. Nutrients 2018, 10, 438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fred-Jaiyesimi, A.; Kio, A.; Richard, W. α-Amylase inhibitory effect of 3β-olean-12-en-3-yl (9Z)-hexadec-9-enoate isolated from Spondias mombin leaf. Food Chem. 2009, 116, 285–288. [Google Scholar] [CrossRef]
- Dominguez Avila, J.A.; Rodrigo Garcia, J.; Gonzalez Aguilar, G.A.; De la Rosa, L.A. The antidiabetic mechanisms of polyphenols related to increased glucagon-like peptide-1 (GLP1) and insulin signaling. Molecules 2017, 22, 903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turdu, G.; Gao, H.; Jiang, Y.; Kabas, M. Plant dipeptidyl peptidase-IV inhibitors as antidiabetic agents: A brief review. Future Med. Chem. 2018, 10, 1229–1239. [Google Scholar] [CrossRef]
- Les, F.; Cásedas, G.; Gómez, C.; Moliner, C.; Valero, M.S.; López, V. The role of anthocyanins as antidiabetic agents: From molecular mechanisms to in vivo and human studies. J. Physiol. Biochem. 2020, 77, 1–23. [Google Scholar] [CrossRef]
- Gowd, V.; Jia, Z.; Chen, W. Anthocyanins as promising molecules and dietary bioactive components against diabetes–A review of recent advances. Trends Food Sci. Technol. 2017, 68, 1–13. [Google Scholar] [CrossRef]
- Prabhakar, P.K.; Doble, M. A target based therapeutic approach towards diabetes mellitus using medicinal plants. Curr. Diabetes Rev. 2008, 4, 291–308. [Google Scholar] [CrossRef]
- Yaribeygi, H.; Farrokhi, F.R.; Butler, A.E.; Sahebkar, A. Insulin resistance: Review of the underlying molecular mechanisms. J. Cell. Physiol. 2019, 234, 8152–8161. [Google Scholar] [CrossRef]
- Eid, H.M.; Haddad, P.S. Mechanisms of action of indigenous antidiabetic plants from the boreal Forest of northeastern Canada. Adv. Endocrinol. 2014, 2014, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, K.; Xu, B. Anti-diabetic effects and mechanisms of dietary polysaccharides. Molecules 2019, 24, 2556. [Google Scholar] [CrossRef] [Green Version]
- Świderska, E.; Strycharz, J.; Wróblewski, A.; Szemraj, J.; Drzewoski, J.; Śliwińska, A. Role of PI3K/AKT pathway in insulin-mediated glucose uptake. Blood Glucose Lev. 2018, 1, 37–53. [Google Scholar]
- Christodoulou, M.-I.; Tchoumtchoua, J.; Skaltsounis, A.-L.; Scorilas, A.; Halabalaki, M. Natural alkaloids intervening the insulin pathway: New hopes for anti-diabetic agents? Curr. Med. Chem. 2019, 26, 5982–6015. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kim, K.-J.; Koh, E.-J.; Lee, B.-Y. Gelidium elegans extract ameliorates type 2 diabetes via regulation of MAPK and PI3K/Akt signaling. Nutrients 2018, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Han, H.-S.; Kang, G.; Kim, J.S.; Choi, B.H.; Koo, S.-H. Regulation of glucose metabolism from a liver-centric perspective. Exp. Mol. Med. 2016, 48, e218. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, T.; Amrein, H. Gluconeogenesis: An ancient biochemical pathway with a new twist. Fly 2017, 11, 218–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, M.Y.; Zaib, S.; Rahman, M.M.; Jannat, S.; Iqbal, J.; Park, S.K.; Chang, M.S. Didymin, a dietary citrus flavonoid exhibits anti-diabetic complications and promotes glucose uptake through the activation of PI3K/Akt signaling pathway in insulin-resistant HepG2 cells. Chem. Biol. Interact. 2019, 305, 180–194. [Google Scholar] [CrossRef] [PubMed]
- He, J.-H.; Chen, L.-X.; Li, H. Progress in the discovery of naturally occurring anti-diabetic drugs and in the identification of their molecular targets. Fitoterapia 2019, 134, 270–289. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, Y.; Dai, Y.; Peng, J. Natural products for the treatment of type 2 diabetes mellitus: Pharmacology and mechanisms. Pharmacol. Res. 2018, 130, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Schmitz-Peiffer, C.; Biden, T.J. Protein kinase C function in muscle, liver, and β-cells and its therapeutic implications for type 2 diabetes. Diabetes 2008, 57, 1774–1783. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.; Gao, J.; Wei, F.; Zhao, J.; Wang, D.; Wei, J. Therapeutic potential of ginsenosides as an adjuvant treatment for diabetes. Front. Pharmacol. 2018, 9, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ighodaro, O.M. Molecular pathways associated with oxidative stress in diabetes mellitus. Biomed. Pharmacother. 2018, 108, 656–662. [Google Scholar] [CrossRef]
- Kaikini, A.A.; Kanchan, D.M.; Nerurkar, U.N.; Sathaye, S. Targeting mitochondrial dysfunction for the treatment of diabetic complications: Pharmacological interventions through natural products. Pharmacogn. Rev. 2017, 11, 128. [Google Scholar]
- Sivitz, W.I.; Yorek, M.A. Mitochondrial dysfunction in diabetes: From molecular mechanisms to functional significance and therapeutic opportunities. Antioxid. Redox Signal. 2010, 12, 537–577. [Google Scholar] [CrossRef] [Green Version]
- Smith, U. Impaired (‘diabetic’) insulin signaling and action occur in fat cells long before glucose intolerance—Is insulin resistance initiated in the adipose tissue? Int. J. Obes. 2002, 26, 897–904. [Google Scholar] [CrossRef] [Green Version]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Galadari, A.; Thayyullathil, F. Role of ceramide in diabetes mellitus: Evidence and mechanisms. Lipids Health Dis. 2013, 12, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and their anti-diabetic effects: Cellular mechanisms and effects to improve blood sugar levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasouli, H.; Yarani, R.; Pociot, F.; Popović-Djordjević, J. Anti-diabetic potential of plant alkaloids: Revisiting current findings and future perspectives. Pharmacol. Res. 2020, 155, 104723. [Google Scholar] [CrossRef] [PubMed]
- Debnath, B.; Singh, W.S.; Das, M.; Goswami, S.; Singh, M.K.; Maiti, D.; Manna, K. Role of plant alkaloids on human health: A review of biological activities. Mater. Today Chem. 2018, 9, 56–72. [Google Scholar] [CrossRef]
- Bribi, N. Pharmacological activity of alkaloids: A review. Asian J. Bot. 2018, 1, 1–6. [Google Scholar]
- Kumar, A.; Aswal, S.; Semwal, R.B.; Chauhan, A.; Joshi, S.K.; Semwal, D.K. Role of plant-derived alkaloids against diabetes and diabetes-related complications: A mechanism-based approach. Phytochem. Rev. 2019, 18, 1277–1298. [Google Scholar] [CrossRef]
- Ajebli, M.; Khan, H.; Eddouks, M. Natural alkaloids and diabetes mellitus: A Review. Endocr. Metab. Immune Disord. Drug Targets 2020, 21, 111–131. [Google Scholar] [CrossRef]
- Haq, I.U.; Imran, M.; Nadeem, M.; Tufail, T.; Gondal, T.A.; Mubarak, M.S. Piperine: A review of its biological effects. Phytother. Res. 2020, 35, 680–700. [Google Scholar] [CrossRef]
- Dabur, R.; Sharma, B.; Mittal, A. Mechanistic approach of anti-diabetic compounds identified from natural sources. Chem. Biol. Lett. 2018, 5, 63–99. [Google Scholar]
- Song, T.-J.; Park, C.-H.; In, K.-R.; Kim, J.-B.; Kim, J.H.; Kim, M.; Chang, H.J. Antidiabetic effects of betulinic acid mediated by the activation of the AMP-activated protein kinase pathway. PLoS ONE. 2021, 16, e0249109. [Google Scholar] [CrossRef]
- Mi, J.; He, W.; Lv, J.; Zhuang, K.; Huang, H.; Quan, S. Effect of berberine on the HPA-axis pathway and skeletal muscle GLUT4 in type 2 diabetes mellitus rats. Diabetes Metab. Syndr. Obes. Targets Ther. 2019, 12, 1717–1752. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Zhang, Y.; Liu, Y.; Hou, L.; Li, S.; Tian, H.; Zhao, T. Berberine modulates gut microbiota and reduces insulin resistance via the TLR4 signaling pathway. Exp. Clin. Endocrinol. Diabetes 2018, 126, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.-J.; Dong, H.; Li, J.-B.; Xu, L.-J.; Zou, X.; Wang, K.-F.; Lu, F.-E.; Yi, P. Berberine inhibits hepatic gluconeogenesis via the LKB1-AMPK-TORC2 signaling pathway in streptozotocin-induced diabetic rats. World J. Gastroenterol. 2015, 21, 7777–7785. [Google Scholar] [CrossRef]
- Liu, C.; Wang, Z.; Song, Y.; Wu, D.; Zheng, X.; Li, P.; Jin, J.; Xu, N.; Li, L. Effects of berberine on amelioration of hyperglycemia and oxidative stress in high glucose and high fat diet-induced diabetic hamsters in vivo. BioMed Res. Int. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, M.; Hirayama, A.; Tsuchiya, K.; Ohgawara, H.; Nakamura, M.; Umezawa, K. Promotion of β-cell differentiation by the alkaloid conophylline in porcine pancreatic endocrine cells. Biomed. Pharmacother. 2010, 64, 226–231. [Google Scholar] [CrossRef]
- Choi, J.S.; Ali, M.Y.; Jung, H.A.; Oh, S.H.; Choi, R.J.; Kim, E.J. Protein tyrosine phosphatase 1B inhibitory activity of alkaloids from Rhizoma Coptidis and their molecular docking studies. J. Ethnopharmacol. 2015, 171, 28–36. [Google Scholar] [CrossRef]
- Ojeda-Montes, M.J.; Ardid-Ruiz, A.; Tomás-Hernández, S.; Gimeno, A.; Cereto-Massagué, A.; Beltrán-Debón, R.; Mulero, M.; Garcia-Vallvé, S.; Pujadas, G.; Valls, C. Ephedrine as a lead compound for the development of new DPP-IV inhibitors. Future Med. Chem. 2017, 9, 2129–2146. [Google Scholar] [CrossRef]
- Yamashita, H.; Kusudo, T.; Takeuchi, T.; Qiao, S.; Tsutsumiuchi, K.; Wang, T.; Wang, Y. Dietary supplementation with evodiamine prevents obesity and improves insulin resistance in ageing mice. J. Funct. Foods 2015, 19, 320–329. [Google Scholar] [CrossRef]
- Patel, O.P.; Mishra, A.; Maurya, R.; Saini, D.; Pandey, J.; Taneja, I.; Raju, K.S.; Kanojiya, S.; Shukla, S.K.; Srivastava, M.N. Naturally occurring carbazole alkaloids from Murraya koenigii as potential antidiabetic agents. J. Nat. Prod. 2016, 79, 1276–1284. [Google Scholar] [CrossRef]
- Wiedemann, M.; Gurrola-Díaz, C.M.; Vargas-Guerrero, B.; Wink, M.; García-López, P.M.; Düfer, M. Lupanine improves glucose homeostasis by influencing KATP channels and insulin gene expression. Molecules 2015, 20, 19085–19100. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Tian, D.; Song, G.; Ming, Q.; Liu, J.; Shen, J.; Liu, Q.-H.; Yang, X. Neferine promotes GLUT4 expression and fusion with the plasma membrane to induce glucose uptake in L6 cells. Front. Pharmacol. 2019, 10, 999–1015. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Chen, Q.-B.; Xin, X.-L.; Aisa, H.-A. Anti-diabetic effect of three new norditerpenoid alkaloids in vitro and potential mechanism via PI3K/Akt signaling pathway. Biomed. Pharmacother. 2017, 87, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, K.H.; Ta, T.N.; Pham, T.H.M.; Nguyen, Q.T.; Pham, H.D.; Mishra, S.; Nyomba, B.G. Nuciferine stimulates insulin secretion from beta cells—An in vitro comparison with glibenclamide. J. Ethnopharmacol. 2012, 142, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Li, W.; Higai, K.; Koike, K. Canthinone alkaloids are novel protein tyrosine phosphatase 1B inhibitors. Bioorg. Med. Chem. Lett. 2015, 25, 1979–1981. [Google Scholar] [CrossRef]
- Liu, C.; Yuan, Y.; Zhou, J.; Hu, R.; Ji, L.; Jiang, G. Piperine ameliorates insulin resistance via inhibiting metabolic inflammation in monosodium glutamate-treated obese mice. BMC Endocr. Disord. 2020, 20, 1–15. [Google Scholar] [CrossRef]
- Yuan, Y.; Zhou, J.; Hu, R.; Zou, L.; Ji, L.; Jiang, G. Piperine protects against pancreatic β-cell dysfunction by alleviating macrophage inflammation in obese mice. Life Sci. 2021, 274, 119312. [Google Scholar] [CrossRef]
- Choi, J.; He, N.; Sung, M.-K.; Yang, Y.; Yoon, S. Sanguinarine is an allosteric activator of AMP-activated protein kinase. Biochem. Biophys. Res. Commun. 2011, 413, 259–263. [Google Scholar] [CrossRef]
- Li, Y.; Li, Q.; Wang, C.; Lou, Z.; Li, Q. Trigonelline reduced diabetic nephropathy and insulin resistance in type 2 diabetic rats through peroxisome proliferator-activated receptor-γ. Exp. Ther. Med. 2019, 18, 1331–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afifia, N.A.; Ramadana, A.; Erianb, E.Y.; Saleh, D.O.; Sedikb, A.A.; Badawic, M. Trigonelline attenuates hepatic complications and molecular alterations in high fat high fructose-induced insulin resistance in rats. Can. J. Physiol. Pharmacol. 2017, 95, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Tiong, S.H.; Looi, C.Y.; Hazni, H.; Arya, A.; Paydar, M.; Wong, W.F.; Cheah, S.-C.; Mustafa, M.R.; Awang, K. Antidiabetic and antioxidant properties of alkaloids from Catharanthus roseus (L.) G. Don. Molecules 2013, 18, 9770–9784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiong, S.H.; Looi, C.Y.; Arya, A.; Wong, W.F.; Hazni, H.; Mustafa, M.R.; Awang, K. Vindogentianine, a hypoglycemic alkaloid from Catharanthus roseu s (L.) G. Don (Apocynaceae). Fitoterapia 2015, 102, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef]
- Van Dijk, A.E.; Olthof, M.R.; Meeuse, J.C.; Seebus, E.; Heine, R.J.; Van Dam, R.M. Acute effects of decaffeinated coffee and the major coffee components chlorogenic acid and trigonelline on glucose tolerance. Diabetes Care 2009, 32, 1023–1025. [Google Scholar] [CrossRef] [Green Version]
- Olthof, M.R.; van Dijk, A.E.; Deacon, C.F.; Heine, R.J.; van Dam, R.M. Acute effects of decaffeinated coffee and the major coffee components chlorogenic acid and trigonelline on incretin hormones. Nutr. Metab. 2011, 8, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinayagam, R.; Jayachandran, M.; Xu, B. Antidiabetic effects of simple phenolic acids: A comprehensive review. Phytother. Res. 2016, 30, 184–199. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.; Abd El-Twab, S.M.; Yousef, A.I.; Ashour, M.B.; Reheim, E.S.A.; Hamed, M.A.A. New insights into the in vitro, in situ and in vivo antihyperglycemic mechanisms of gallic acid and p-coumaric acid. Arch. Physiol. Biochem. 2020, 1, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Adisakwattana, S. Cinnamic acid and its derivatives: Mechanisms for prevention and management of diabetes and its complications. Nutrients 2017, 9, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajik, N.; Tajik, M.; Mack, I.; Enck, P. The potential effects of chlorogenic acid, the main phenolic components in coffee, on health: A comprehensive review of the literature. Eur. J. Nutr. 2017, 56, 2215–2244. [Google Scholar] [CrossRef] [PubMed]
- Harakeh, S.; Almuhayawi, M.; Al Jaouni, S.; Almasaudi, S.; Hassan, S.; Al Amri, T.; Azhar, N.; Abd-Allah, E.; Ali, S.; El-Shitany, N.; et al. Antidiabetic effects of novel ellagic acid nanoformulation: Insulin-secreting and anti-apoptosis effects. Saudi J. Biol. Sci. 2020, 27, 3474–3480. [Google Scholar] [CrossRef]
- Alegbe, E.O.; Teralı, K.; Olofinsan, K.A.; Surgun, S.; Ogbaga, C.C.; Ajiboye, T.O. Antidiabetic activity-guided isolation of gallic and protocatechuic acids from Hibiscus sabdariffa calyxes. J. Food Biochem. 2019, 43, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Srinivasulu, C.; Ramgopal, M.; Ramanjaneyulu, G.; Anuradha, C.; Kumar, C.S. Syringic acid (SA)‒a review of its occurrence, biosynthesis, pharmacological and industrial importance. Biomed. Pharmacother. 2018, 108, 547–557. [Google Scholar] [CrossRef]
- Mohammed, F.Z.; El-Shehabi, M. Antidiabetic activity of caffeic acid and 18β-glycyrrhetinic acid and its relationship with the antioxidant property. Asian J. Pharm. Clin. Res. 2015, 8, 229–235. [Google Scholar]
- Xu, W.; Luo, Q.; Wen, X.; Xiao, M.; Mei, Q. Antioxidant and anti-diabetic effects of caffeic acid in a rat model of diabetes. Trop. J. Pharm. Res. 2020, 19, 1227–1232. [Google Scholar] [CrossRef]
- Amalan, V.; Vijayakumar, N.; Indumathi, D.; Ramakrishnan, A. Antidiabetic and antihyperlipidemic activity of p-coumaric acid in diabetic rats, role of pancreatic GLUT 2: In vivo approach. Biomed. Pharmacother. 2016, 84, 230–236. [Google Scholar] [CrossRef]
- Abdel-Moneim, A.; Abd El-Twab, S.M.; Yousef, A.I.; Reheim, E.S.A.; Ashour, M.B. Modulation of hyperglycemia and dyslipidemia in experimental type 2 diabetes by gallic acid and p-coumaric acid: The role of adipocytokines and PPARγ. Biomed. Pharmacother. 2018, 105, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Amalan, V.; Vijayakumar, N.; Ramakrishnan, A. p-Coumaric acid regulates blood glucose and antioxidant levels in streptozotocin induced diabetic rats. J. Chem. Pharm. Res. 2015, 7, 831–839. [Google Scholar]
- Hafizur, R.M.; Hameed, A.; Shukrana, M.; Raza, S.A.; Chishti, S.; Kabir, N.; Siddiqui, R.A. Cinnamic acid exerts anti-diabetic activity by improving glucose tolerance in vivo and by stimulating insulin secretion in vitro. Phytomedicine 2015, 22, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Chang, C.; Zhang, L.; Liu, Y.; Huang, X.; Chen, Z. Chlorogenic acid improves late diabetes through adiponectin receptor signaling pathways in db/db mice. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polce, S.A.; Burke, C.; França, L.M.; Kramer, B.; Paes, A.M.d.A.; Carrillo-Sepulveda, M.A. Ellagic acid alleviates hepatic oxidative stress and insulin resistance in diabetic female rats. Nutrients 2018, 10, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narasimhan, A.; Chinnaiyan, M.; Karundevi, B. Ferulic acid exerts its antidiabetic effect by modulating insulin-signalling molecules in the liver of high-fat diet and fructose-induced type-2 diabetic adult male rat. Appl. Physiol. Nutr. Metab. 2015, 40, 769–781. [Google Scholar] [CrossRef]
- Narasimhan, A.; Chinnaiyan, M.; Karundevi, B. Ferulic acid regulates hepatic GLUT2 gene expression in high fat and fructose-induced type-2 diabetic adult male rat. Eur. J. Pharmacol. 2015, 761, 391–397. [Google Scholar] [CrossRef]
- Abdelmageed, M.E.; Shehatou, G.S.; Suddek, G.M.; Salem, H.A. Protocatechuic acid improves hepatic insulin resistance and restores vascular oxidative status in type-2 diabetic rats. Environ. Toxicol. Pharmacol. 2021, 83, 103577. [Google Scholar] [CrossRef]
- Scazzocchio, B.; Varì, R.; Filesi, C.; Del Gaudio, I.; D’Archivio, M.; Santangelo, C.; Iacovelli, A.; Galvano, F.; Pluchinotta, F.R.; Giovannini, C.; et al. Protocatechuic acid activates key components of insulin signaling pathway mimicking insulin activity. Mol. Nutr. Food Res. 2015, 59, 1472–1481. [Google Scholar] [CrossRef]
- El-Sonbaty, Y.A.; Suddek, G.M.; Megahed, N.; Gameil, N.M. Protocatechuic acid exhibits hepatoprotective, vasculoprotective, antioxidant and insulin-like effects in dexamethasone-induced insulin-resistant rats. Biochimie 2019, 167, 119–134. [Google Scholar] [CrossRef]
- Sabahi, Z.; Khoshnoud, M.J.; Khalvati, B.; Hashemi, S.-S.; Farsani, Z.G.; Gerashi, H.M.; Rashedinia, M. Syringic acid improves oxidative stress and mitochondrial biogenesis in the liver of streptozotocin-induced diabetic rats. Asian Pac. J. Trop. Biomed. 2020, 10, 111–119. [Google Scholar]
- Ghosh, D.; Konishi, T. Anthocyanins and anthocyanin-rich extracts: Role in diabetes and eye function. Asia Pac. J. Clin. Nutr. 2007, 16, 200–208. [Google Scholar] [PubMed]
- Li, D.; Wang, P.; Luo, Y.; Zhao, M.; Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. Nutr. 2017, 57, 1729–1741. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, H.; Fernandes, A.; Brás, N.F.; Mateus, N.; de Freitas, V.; Fernandes, I. Anthocyanins as antidiabetic agents—In vitro and in silico approaches of preventive and therapeutic effects. Molecules 2020, 25, 3813. [Google Scholar] [CrossRef]
- Tsuda, T. Dietary anthocyanin-rich plants: Biochemical basis and recent progress in health benefits studies. Mol. Nutr. Food Res. 2012, 56, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Belwal, T.; Nabavi, S.F.; Nabavi, S.M.; Habtemariam, S. Dietary anthocyanins and insulin resistance: When food becomes a medicine. Nutrients 2017, 9, 1111. [Google Scholar] [CrossRef]
- Cao, H.; Ou, J.; Chen, L.; Zhang, Y.; Szkudelski, T.; Delmas, D.; Daglia, M.; Xiao, J. Dietary polyphenols and type 2 diabetes: Human study and clinical trial. Crit. Rev. Food Sci. Nutr. 2019, 59, 3371–3379. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Sharifi-Rad, J.; Cappellini, F.; Reiner, Ž.; Zorzan, D.; Imran, M.; Sener, B.; Kilic, M.; El-Shazly, M.; Fahmy, N.M.; et al. The therapeutic potential of anthocyanins: Current approaches based on their molecular mechanism of action. Front. Pharmacol. 2020, 11, 1300–1320. [Google Scholar] [CrossRef]
- Momtaz, S.; Salek-Maghsoudi, A.; Abdolghaffari, A.H.; Jasemi, E.; Rezazadeh, S.; Hassani, S.; Ziaee, M.; Abdollahi, M.; Behzad, S.; Nabavi, S.M. Polyphenols targeting diabetes via the AMP-activated protein kinase pathway; future approach to drug discovery. Crit. Rev. Clin. Lab. Sci. 2019, 56, 472–492. [Google Scholar] [CrossRef]
- Jennings, A.; Welch, A.A.; Spector, T.; Macgregor, A.; Cassidy, A. Intakes of anthocyanins and flavones are associated with biomarkers of insulin resistance and inflammation in women. J. Nutr. 2014, 144, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Soltani, R.; Gorji, A.; Asgary, S.; Sarrafzadegan, N.; Siavash, M. Evaluation of the effects of Cornus mas L. fruit extract on glycemic control and insulin level in type 2 diabetic adult patients: A randomized double-blind placebo-controlled clinical trial. Evid. Based Complement. Altern. Med. 2015, 2015, 1–5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Zhang, R.; Shi, W.; Li, L.; Liu, H.; Liu, Z.; Wu, L. The Multifunctional Benefits of Naturally Occurring Delphinidin and Its Glycosides. J. Agric. Food Chem. 2019, 67, 11288–11306. [Google Scholar] [CrossRef]
- Rupasinghe, H.V.; Arumuggam, N.; Amararathna, M.; De Silva, A. The potential health benefits of haskap (Lonicera caerulea L.): Role of cyanidin-3-O-glucoside. J. Funct. Foods 2018, 44, 24–39. [Google Scholar] [CrossRef]
- Aba, P.E.; Asuzu, I.U. Mechanisms of actions of some bioactive anti-diabetic principles from phytochemicals of medicinal plants: A review. Indian J. Nat. Prod. Resour. 2018, 9, 85–96. [Google Scholar]
- Suantawee, T.; Elazab, S.T.; Hsu, W.H.; Yao, S.; Cheng, H.; Adisakwattana, S. Cyanidin stimulates insulin secretion and pancreatic β-cell gene expression through activation of l-type voltage-dependent Ca2+ channels. Nutrients 2017, 9, 814. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, J.; Teuber, S.; Morera, F.J.; Ojeda, C.; Flores, C.A.; Hidalgo, M.A.; Núñez, L.; Villalobos, C.; Burgos, R.A. Delphinidin reduces glucose uptake in mice jejunal tissue and human intestinal cells lines through FFA1/GPR40. Int. J. Mol. Sci. 2017, 18, 750. [Google Scholar] [CrossRef] [Green Version]
- Demir, Y.; Durmaz, L.; Taslimi, P.; Gulçin, İ. Antidiabetic properties of dietary phenolic compounds: Inhibition effects on α-amylase, aldose reductase, and α-glycosidase. Biotechnol. Appl. Biochem. 2019, 66, 781–786. [Google Scholar] [CrossRef]
- Rasouli, H.; Hosseini-Ghazvini, S.M.-B.; Adibi, H.; Khodarahmi, R. Differential α-amylase/α-glucosidase inhibitory activities of plant-derived phenolic compounds: A virtual screening perspective for the treatment of obesity and diabetes. Food Funct. 2017, 8, 1942–1954. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Ma, Y.; Cheng, G.; Zhang, Y.; Cai, S. Comparative study of dietary flavonoids with different structures as α-glucosidase inhibitors and insulin sensitizers. J. Agric. Food Chem. 2019, 67, 10521–10533. [Google Scholar] [CrossRef] [PubMed]
- Hussain, T.; Tan, B.; Murtaza, G.; Liu, G.; Rahu, N.; Kalhoro, M.S.; Kalhoro, D.H.; Adebowale, T.O.; Mazhar, M.U.; ur Rehman, Z. Flavonoids and type 2 diabetes: Evidence of efficacy in clinical and animal studies and delivery strategies to enhance their therapeutic efficacy. Pharmacol. Res. 2020, 152, 104629. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Luo, J.; Huang, J.; Wen, Q. Flavonoids intake and risk of type 2 diabetes mellitus: A meta-analysis of prospective cohort studies. Medicine 2018, 97, 1–7. [Google Scholar] [CrossRef]
- Unuofin, J.O.; Lebelo, S.L. Antioxidant effects and mechanisms of medicinal plants and their bioactive compounds for the prevention and treatment of type 2 diabetes: An updated review. Oxid. Med. Cell. Longev. 2020, 2020, 893–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Luo, J.; Jia, Z.; Zhen, W.; Zhou, K.; Gilbert, E.; Liu, D. Baicalein protects against type 2 diabetes via promoting islet β-cell function in obese diabetic mice. Int. J. Endocrinol. 2014, 2014, 846742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Islam, A.; Islam, M.S.; Rahman, M.K.; Uddin, M.N.; Akanda, M.R. The pharmacological and biological roles of eriodictyol. Arch. Pharm. Res. 2020, 43, 1–11. [Google Scholar] [CrossRef]
- Antika, L.D.; Dewi, R.M. Pharmacological aspects of fisetin. Asian Pac. J. Trop. Biomed. 2021, 11, 1–9. [Google Scholar] [CrossRef]
- Marella, S. Flavonoids—The most potent poly-phenols as antidiabetic agents: An overview. Mod. Approaches Drug Des. 2017, 1, 2–5. [Google Scholar] [CrossRef]
- Sun, M.-Y.; Ye, Y.; Xiao, L.; Rahman, K.; Xia, W.; Zhang, H. Daidzein: A review of pharmacological effects. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 117–132. [Google Scholar] [CrossRef] [Green Version]
- Babu, P.V.A.; Liu, D.; Gilbert, E.R. Recent advances in understanding the anti-diabetic actions of dietary flavonoids. J. Nutr. Biochem. 2013, 24, 1777–1789. [Google Scholar] [CrossRef] [Green Version]
- Den Hartogh, D.J.; Tsiani, E. Antidiabetic properties of naringenin: A citrus fruit polyphenol. Biomolecules 2019, 9, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagan, K.; Radika, M.K.; Priyadarshini, E.; Venkatraman, C. A study on the inhibitory potential of DPP-IV enzyme by apigenin through in silico and in vivo approaches. Res. J. Recent Sci. 2015, 2277, 22–29. [Google Scholar]
- Fang, P.; Sun, Y.; Gu, X.; Shi, M.; Bo, P.; Zhang, Z.; Bu, L. Baicalin ameliorates hepatic insulin resistance and gluconeogenic activity through inhibition of p38 MAPK/PGC-1α pathway. Phytomedicine 2019, 64, 153074. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Hur, H.J.; Kwon, D.Y.; Hwang, J.T. Tangeretin stimulates glucose uptake via regulation of AMPK signaling pathways in C2C12 myotubes and improves glucose tolerance in high-fat diet-induced obese mice. Mol. Cell. Endocrinol. 2012, 358, 127–134. [Google Scholar] [CrossRef]
- Wang, N.; Yi, W.J.; Tan, L.; Zhang, J.H.; Xu, J.; Chen, Y.; Qin, M.; Yu, S.; Guan, J.; Zhang, R. Apigenin attenuates streptozotocin-induced pancreatic β cell damage by its protective effects on cellular antioxidant defense. Vitr. Cell. Dev. Biol. Anim. 2017, 53, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jiang, H.; Cao, S.; Chen, Q.; Cui, M.; Wang, Z.; Li, D.; Zhou, J.; Qiu, F.; Kang, N. Baicalin and its metabolites suppresses gluconeogenesis through activation of AMPK or AKT in insulin resistant HepG-2 cells. Eur. J. Med. Chem. 2017, 141, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Yu, M.; Min, W.; Wan, D.; Han, S.; Shan, Y.; Wang, R.; Shi, M.; Zhang, Z.; Bo, P. Effect of baicalin on GLUT4 expression and glucose uptake in myotubes of rats. Life Sci. 2018, 196, 156–161. [Google Scholar] [CrossRef]
- Fang, P.; Yu, M.; Zhang, L.; Wan, D.; Shi, M.; Zhu, Y.; Bo, P.; Zhang, Z. Baicalin against obesity and insulin resistance through activation of AKT/AS160/GLUT4 pathway. Mol. Cell. Endocrinol. 2017, 448, 77–86. [Google Scholar] [CrossRef]
- Samarghandian, S.; Azimi-Nezhad, M.; Samini, F.; Farkhondeh, T. Chrysin treatment improves diabetes and its complications in liver, brain, and pancreas in streptozotocin-induced diabetic rats. Can. J. Physiol. Pharmacol. 2016, 94, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Satyanarayana, K.; Sravanthi, K.; Shaker, I.A.; Ponnulakshmi, R.; Selvaraj, J. Role of chrysin on expression of insulin signaling molecules. J. Ayurveda Integr. Med. 2015, 6, 248. [Google Scholar] [CrossRef] [Green Version]
- Hameed, A.; Hafizur, R.M.; Hussain, N.; Raza, S.A.; Rehman, M.; Ashraf, S.; Ul-Haq, Z.; Khan, F.; Abbas, G.; Choudhary, M.I. Eriodictyol stimulates insulin secretion through cAMP/PKA signaling pathway in mice islets. Eur. J. Pharmacol. 2018, 820, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Kwon, E.-Y.; Do, E.; Lee, D.; Kim, J.; Choi, M.-S. Elucidation of anti-obesity and anti-diabetic function of eriodictyol in diet-induced obese mice. Clin. Nutr. 2018, 37, 146. [Google Scholar] [CrossRef]
- Makena, W.; Hambolu, J.O.; Timbuak, J.A.; Umana, U.E.; Iliya, A.I.; Dibal, N.I. Mormodica charantia L. fruit and genistein ameliorates type 2 diabetes in rats by preventing lipid accumulation, insulin resistance and enhancing beta cell function. J. Diabetes Metab. Disord. 2020, 19, 1303–1310. [Google Scholar] [CrossRef]
- Kim, A.; Lee, W.; Yun, J.-M. Luteolin and fisetin suppress oxidative stress by modulating sirtuins and forkhead box O3a expression under in vitro diabetic conditions. Nutr. Res. Pract. 2017, 11, 430–434. [Google Scholar] [CrossRef] [Green Version]
- Dokumacioglu, E.; Iskender, H.; Sen, T.M.; Ince, I.; Dokumacioglu, A.; Kanbay, Y.; Erbas, E.; Saral, S. The effects of hesperidin and quercetin on serum tumor necrosis factor-alpha and interleukin-6 levels in streptozotocin-induced diabetes model. Pharmacogn. Mag. 2018, 14, 167–173. [Google Scholar]
- Iskender, H.; Dokumacioglu, E.; Sen, T.M.; Ince, I.; Kanbay, Y.; Saral, S. The effect of hesperidin and quercetin on oxidative stress, NF-κB and SIRT1 levels in a STZ-induced experimental diabetes model. Biomed. Pharmacother. 2017, 90, 500–508. [Google Scholar] [CrossRef]
- Sundaram, R.; Nandhakumar, E.; Haseena Banu, H. Hesperidin, a citrus flavonoid ameliorates hyperglycemia by regulating key enzymes of carbohydrate metabolism in streptozotocin-induced diabetic rats. Toxicol. Mech. Methods 2019, 29, 644–653. [Google Scholar] [CrossRef]
- Razavi, T.; Kouhsari, S.M.; Abnous, K. Morin exerts anti-diabetic effects in human HepG2 cells via down-regulation of miR-29a. Exp. Clin. Endocrinol. Diabetes 2019, 127, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Rajappa, R.; Sireesh, D.; Salai, M.B.; Ramkumar, K.M.; Sarvajayakesavulu, S.; Madhunapantula, S.V. Treatment with naringenin elevates the activity of transcription factor Nrf2 to protect pancreatic β-cells from streptozotocin-induced diabetes in vitro and in vivo. Front. Pharmacol. 2019, 9, 1562–1582. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, O.M.; Hassan, M.A.; Abdel-Twab, S.M.; Azeem, M.N.A. Navel orange peel hydroethanolic extract, naringin and naringenin have anti-diabetic potentials in type 2 diabetic rats. Biomed. Pharmacother. 2017, 94, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.J.; Kim, J.H.; Pan, J.H.; Kim, J.K.; Park, T.S.; Kim, Y.J.; Lee, J.H.; Kim, J.H. Naringin protects pancreatic β-cells against oxidative stress-induced apoptosis by inhibiting both intrinsic and extrinsic pathways in insulin-deficient diabetic mice. Mol. Nutr. Food Res. 2018, 62, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Dhanya, R.; Arya, A.; Nisha, P.; Jayamurthy, P. Quercetin, a lead compound against type 2 diabetes ameliorates glucose uptake via AMPK pathway in skeletal muscle cell line. Front. Pharmacol. 2017, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Li, Q.; Li, K.; Zhu, L.; Lin, X.; Lin, X.; Shen, Q.; Li, G.; Xie, X. Quercetin improves glucose and lipid metabolism of diabetic rats: Involvement of Akt signaling and SIRT1. J. Diabetes Res. 2017, 2017, 3417306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Han, J.; Zhou, Z.; Li, D. Tangeretin inhibits streptozotocin-induced cell apoptosis via regulating NF-κB pathway in INS-1 cells. J. Cell. Biochem. 2019, 120, 3286–3293. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Chen, J.; Ren, W.; Zhu, Y.; Zhao, Q.; Zhang, K.; Su, D.; Qiu, C.; Zhang, W.; Li, K. Citrus flavone tangeretin is a potential insulin sensitizer targeting hepatocytes through suppressing MEK-ERK1/2 pathway. Biochem. Biophys. Res. Commun. 2020, 529, 277–282. [Google Scholar] [CrossRef]
- Akinwumi, B.C.; Bordun, K.-A.M.; Anderson, H.D. Biological activities of stilbenoids. Int. J. Mol. Sci. 2018, 19, 792. [Google Scholar] [CrossRef] [Green Version]
- Shao, J.-W.; Jiang, J.-L.; Zou, J.-J.; Yang, M.-Y.; Chen, F.-M.; Zhang, Y.-J.; Jia, L. Therapeutic potential of ginsenosides on diabetes: From hypoglycemic mechanism to clinical trials. J. Funct. Foods 2020, 64, 103630. [Google Scholar] [CrossRef]
- Sun, H.; Liu, X.; Long, S.R.; Ge, H.; Wang, Y.; Yu, S.; Xue, Y.; Zhang, Y.; Li, X.; Li, W. Antidiabetic effects of pterostilbene through PI3K/Akt signal pathway in high fat diet and STZ-induced diabetic rats. Eur. J. Pharmacol. 2019, 859, 172526. [Google Scholar] [CrossRef]
- Gencoglu, H.; Tuzcu, M.; Hayirli, A.; Sahin, K. Protective effects of resveratrol against streptozotocin-induced diabetes in rats by modulation of visfatin/sirtuin-1 pathway and glucose transporters. Int. J. Food Sci. Nutr. 2015, 66, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Elango, B.; Dornadula, S.; Paulmurugan, R.; Ramkumar, K.M. Pterostilbene ameliorates streptozotocin-induced diabetes through enhancing antioxidant signaling pathways mediated by Nrf2. Chem. Res. Toxicol. 2016, 29, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Elekofehinti, O.O. Saponins: Anti-diabetic principles from medicinal plants—A review. Pathophysiology 2015, 22, 95–103. [Google Scholar] [CrossRef]
- Chen, W.; Balan, P.; Popovich, D.G. Review of ginseng anti-diabetic studies. Molecules 2019, 24, 4501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, K.; Takamura, Y.; Iwamoto, T.; Nomura, M.; Iwasaki, H.; Ohdera, M.; Murakoshi, M.; Sugiyama, K.; Matsuyama, K.; Manabe, Y.; et al. Dammarane-type triterpene extracts of Panax notoginseng root ameliorates hyperglycemia and insulin sensitivity by enhancing glucose uptake in skeletal muscle. Biosci. Biotechnol. Biochem. 2017, 81, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Hemalatha, T.; Pulavendran, S.; Balachandran, C.; Manohar, B.M.; Puvanakrishnan, R. Arjunolic acid: A novel phytomedicine with multifunctional therapeutic applications. Indian J. Exp. Boil. 2010, 48, 238–247. [Google Scholar]
- Li, H.; Yao, Y.; Li, L. Coumarins as potential antidiabetic agents. J. Pharm. Pharmacol. 2017, 69, 1253–1264. [Google Scholar] [CrossRef] [Green Version]
- Gan, Q.; Wang, J.; Hu, J.; Lou, G.; Xiong, H.; Peng, C.; Zheng, S.; Huang, Q. The role of diosgenin in diabetes and diabetic complications. J. Steroid Biochem. Mol. Biol. 2020, 198, 105575. [Google Scholar] [CrossRef]
- Sharma, A.; Lee, H.-J. Ginsenoside compound k: Insights into recent studies on pharmacokinetics and health-promoting activities. Biomolecules 2020, 10, 1028. [Google Scholar] [CrossRef]
- Park, M.W.; Ha, J.; Chung, S.H. (S)-ginsenoside Rg3 enhances glucose-stimulated insulin secretion and activates AMPK. Biol. Pharm. Bull. 2008, 31, 748–751. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Yuan, H.D.; Chung, S.H. Ginsenoside Rg1 suppresses hepatic glucose production via AMP-activated protein kinase in HepG2 cells. Biol. Pharm. Bull. 2010, 33, 325–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, S.; Li, W.; Yu, Y.; Yao, F.; Lixiang, A.; Lan, X.; Guan, F.; Zhang, M.; Chen, L. Ginsenoside Compound K suppresses the hepatic gluconeogenesis via activating adenosine-5′ monophosphate kinase: A study in vitro and in vivo. Life Sci. 2015, 139, 8–15. [Google Scholar] [CrossRef]
- Chen, W.; Wang, J.; Luo, Y.; Wang, T.; Li, X.; Li, A.; Li, J.; Liu, K.; Liu, B. Ginsenoside Rb1 and compound K improve insulin signaling and inhibit ER stress-associated NLRP3 inflammasome activation in adipose tissue. J. Ginseng Res. 2016, 40, 351–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabandeh, M.R.; Jafari, H.; Hosseini, S.A.; Hashemitabar, M. Ginsenoside Rb1 stimulates adiponectin signaling in C2C12 muscle cells through up-regulation of AdipoR1 and AdipoR2 proteins. Pharma. Biol. 2015, 53, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Ye, L.; Zhang, H.; Zhao, J.; Wang, G.; Guo, C.; Shang, W. Ginsenoside Rb1 ameliorates liver fat accumulation by upregulating perilipin expression in adipose tissue of db/db obese mice. J. Ginseng Res. 2015, 39, 199–205. [Google Scholar] [CrossRef] [Green Version]
- Song, B.; Ding, L.; Zhang, H.; Chu, Y.; Chang, Z.; Yu, Y.; Guo, D.; Zhang, S.; Liu, X. Ginsenoside Rb1 increases insulin sensitivity through suppressing 11β-hydroxysteroid dehydrogenase type I. Am. J. Transl. Res. 2017, 9, 1049–1057. [Google Scholar]
- Tian, W.; Chen, L.; Zhang, L.; Wang, B.; Li, X.; Fan, K.; Ai, C.; Xia, X.; Li, S.; Li, Y. Effects of ginsenoside Rg1 on glucose metabolism and liver injury in streptozotocin-induced type 2 diabetic rats. Genet. Mol. Res. 2017, 16, 1–13. [Google Scholar] [CrossRef]
- Liu, Y.; Deng, J.; Fan, D. Ginsenoside Rk3 ameliorates high-fat-diet/streptozocin induced type 2 diabetes mellitus in mice via the AMPK/Akt signaling pathway. Food Funct. 2019, 10, 2538–2551. [Google Scholar] [CrossRef]
- Kumari, M.; Jain, S. Tannins: An antinutrient with positive effect to manage diabetes. Res. J. Recent Sci. 2012, 1, 1–8. [Google Scholar]
- Ajebli, M.; Eddouks, M. The promising role of plant tannins as bioactive antidiabetic agents. Curr. Med. Chem. 2019, 26, 4852–4884. [Google Scholar] [CrossRef] [PubMed]
- Türkan, F.; Taslimi, P.; Saltan, F.Z. Tannic acid as a natural antioxidant compound: Discovery of a potent metabolic enzyme inhibitor for a new therapeutic approach in diabetes and Alzheimer’s disease. J. Biochem. Mol. Toxicol. 2019, 33, e22340. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Abu-Izneid, T.; Patel, S.; Pan, X.; Naz, S.; Silva, A.S.; Saeed, F.; Suleria, H.A.R. Proanthocyanidins: A comprehensive review. Biomed. Pharmacother. 2019, 116, 108999. [Google Scholar] [CrossRef]
- Bueno, P.S.A.; Kato-Schwartz, C.G.; de Souza Lima, D.; Bracht, A.; Peralta, R.M.; Seixas, F.A.V. In silico evaluation of condensed and hydrolysable tannins as inhibitors of pancreatic α-amylase. J. Mol. Model. 2019, 25, 1–9. [Google Scholar] [CrossRef]
- Shoukat, M.; Sorrentino, A. Cereal β-glucan: A promising prebiotic polysaccharide and its impact on the gut health. Int. J. Food Sci. Technol. 2021, 56, 2088–2097. [Google Scholar] [CrossRef]
- Pan, G.; Zhao, L.; Xiao, N.; Yang, K.; Ma, Y.; Zhao, X.; Fan, Z.; Zhang, Y.; Yao, Q.; Lu, K.; et al. Total synthesis of 8-(6 ″-umbelliferyl)-apigenin and its analogs as anti-diabetic reagents. Eur. J. Med. Chem. 2016, 122, 674–683. [Google Scholar] [CrossRef]
- Murali, R.; Srinivasan, S.; Ashokkumar, N. Antihyperglycemic effect of fraxetin on hepatic key enzymes of carbohydrate metabolism in streptozotocin-induced diabetic rats. Biochimie 2013, 95, 1848–1854. [Google Scholar] [CrossRef]
- Salau, V.F.; Erukainure, O.L.; Ibeji, C.U.; Koorbanally, N.A.; Islam, M.S. Umbelliferone stimulates glucose uptake; modulates gluconeogenic and nucleotide-hydrolyzing enzymes activities, and dysregulated lipid metabolic pathways in isolated psoas muscle. J. Funct. Foods 2020, 67, 103847. [Google Scholar] [CrossRef]
- Sim, M.-O.; Lee, H.-I.; Ham, J.R.; Seo, K.-I.; Lee, M.-K. Long-term supplementation of esculetin ameliorates hepatosteatosis and insulin resistance partly by activating AdipoR2–AMPK pathway in diet-induced obese mice. J. Funct. Foods 2015, 15, 160–171. [Google Scholar] [CrossRef]
- Mo, Z.; Li, L.; Yu, H.; Wu, Y.; Li, H. Coumarins ameliorate diabetogenic action of dexamethasone via Akt activation and AMPK signaling in skeletal muscle. J. Pharmacol. Sci. 2019, 139, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.-G.; Zhao, X.; Zhong, W.; Xie, M.-L. Osthole improves glucose and lipid metabolism via modulation of PPARα/γ-mediated target gene expression in liver, adipose tissue, and skeletal muscle in fatty liver rats. Pharm. Biol. 2016, 54, 882–888. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.H.; Park, J.E.; Han, J.S. Scopoletin increases glucose uptake through activation of PI3K and AMPK signaling pathway and improves insulin sensitivity in 3T3-L1 cells. Nutr. Res. 2020, 74, 52–61. [Google Scholar] [CrossRef]
- Jang, J.H.; Park, J.E.; Han, J.S. Scopoletin inhibits α-glucosidase in vitro and alleviates postprandial hyperglycemia in mice with diabetes. Eur. J. Pharmacol. 2018, 834, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Ramu, R.; Shirahatti, P.; Zameer, F.; Dhananjaya, B.L. Assessment of in vivo antidiabetic properties of umbelliferone and lupeol constituents of banana (Musa sp. var. Nanjangud Rasa Bale) flower in hyperglycaemic rodent model. PLoS ONE 2016, 11, e0151135. [Google Scholar]
- Kaur, K.K.; Allahbadia, G.; Singh, M. Monoterpenes—A class of terpenoid group of natural products as a source of natural antidiabetic agents in the future—A review. CPQ Nutr. 2019, 3, 1–21. [Google Scholar]
- Panigrahy, S.K.; Bhatt, R.; Kumar, A. Targeting type II diabetes with plant terpenes: The new and promising antidiabetic therapeutics. Biologia 2021, 76, 241–254. [Google Scholar] [CrossRef]
- Anandakumar, P.; Kamaraj, S.; Vanitha, M.K. D-limonene: A multifunctional compound with potent therapeutic effects. J. Food Biochem. 2021, 45, 1–10. [Google Scholar] [CrossRef]
- Ilić, V.; Vukmirović, S.; Stilinović, N.; Čapo, I.; Arsenović, M.; Milijašević, B. Insight into anti-diabetic effect of low dose of stevioside. Biomed. Pharmacother. 2017, 90, 216–221. [Google Scholar] [CrossRef]
- Saravanan, R.; Ramachandran, V. Effect of Rebaudioside A, a diterpenoid on glucose homeostasis in STZ-induced diabetic rats. J. Physiol. Biochem. 2012, 68, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Siddique, H.R.; Saleem, M. Beneficial health effects of lupeol triterpene: A review of preclinical studies. Life Sci. 2011, 88, 285–293. [Google Scholar] [CrossRef]
- Hung, H.-Y.; Qian, K.; Morris-Natschke, S.L.; Hsu, C.-S.; Lee, K.-H. Recent discovery of plant-derived anti-diabetic natural products. Nat. Prod. Rep. 2012, 29, 580–606. [Google Scholar] [CrossRef]
- Lee, H.-A.; Kim, M.-J.; Han, J.-S. Alleviating effects of lupeol on postprandial hyperglycemia in diabetic mice. Toxicol. Res. 2021, 10, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Ramachandran, V.; Saravanan, R. Glucose uptake through translocation and activation of GLUT4 in PI3K/Akt signaling pathway by asiatic acid in diabetic rats. Hum. Exp. Toxicol. 2015, 34, 884–893. [Google Scholar] [CrossRef]
- Li, Y.; Mai, Y.; Qiu, X.; Chen, X.; Li, C.; Yuan, W.; Hou, N. Effect of long-term treatment of Carvacrol on glucose metabolism in Streptozotocin-induced diabetic mice. BMC Complement. Med. Ther. 2020, 20, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, S.; Sarabi, M.M.; Gholami, M.; Assadollahi, V.; Khorramabadi, R.M.; Moradi, F.H.; Ahmadvand, H. D-limonene in diabetic rats. J. Ren. Inj. Prev. 2021, 10, 1–8. [Google Scholar]
- Bhasker, S.; Madhav, H.; Chinnamma, M. Molecular evidence of insulinomimetic property exhibited by steviol and stevioside in diabetes induced L6 and 3T3L1 cells. Phytomedicine 2015, 22, 1037–1044. [Google Scholar] [CrossRef]
- Saleh, O.M.; Awad, N.S.; Soliman, M.M.; Mansour, A.A.; Nassan, M.A. Insulin-mimetic activity of stevioside on diabetic rats: Biochemical, molecular and histopathological study. Afr. J. Tradit. Complement. Altern. Med. 2016, 13, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Shreenithi, S.; Vishnupriya, V.; Ponnulakshmi, R.; Gayathri, R.; Madhan, K.; Shyamaladevi, B.; Selvaraj, J. In silico and in vivo approach to identify the antidiabetic activity of lupeol. Drug Invent. Today 2019, 11, 1113–1116. [Google Scholar]
- Dhivyadharshini, A.; Vishnpriya, V.; Ponnulakshmi, R.; Gayathri, R.; Madhan, K.; Shyamaladevi, B.; Selvaraj, J. Effect of lupeol on insulin receptor substrate-1 and AKt expression in adipose tissue of type-2 diabetic rats. Drug Invent. Today 2019, 12, 801–805. [Google Scholar]
- Birgani, G.A.; Ahangarpour, A.; Khorsandi, L.; Moghaddam, H.F. Anti-diabetic effect of betulinic acid on streptozotocin-nicotinamide induced diabetic male mouse model. Braz. J. Pharm. Sci. 2018, 54, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Ahangarpour, A.; Shabani, R.; Farbood, Y. The effect of betulinic acid on leptin, adiponectin, hepatic enzyme levels and lipid profiles in streptozotocin–nicotinamide-induced diabetic mice. Res. Pharm. Sci. 2018, 13, 142–148. [Google Scholar] [CrossRef]
- Ding, H.; Wu, X.; Pan, J.; Hu, X.; Gong, D.; Zhang, G. New insights into the inhibition mechanism of betulinic acid on α-glucosidase. J. Agric. Food Chem. 2018, 66, 7065–7075. [Google Scholar] [CrossRef] [PubMed]
- Brusotti, G.; Montanari, R.; Capelli, D.; Cattaneo, G.; Laghezza, A.; Tortorella, P.; Loiodice, F.; Peiretti, F.; Bonardo, B.; Paiardini, A.; et al. Betulinic acid is a PPARγ antagonist that improves glucose uptake, promotes osteogenesis and inhibits adipogenesis. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellano, J.; Guinda, A.; Macías, L.; Santos-Lozano, J.; Lapetra, J.; Rada, M. Free radical scavenging and α-glucosidase inhibition, two potential mechanisms involved in the anti-diabetic activity of oleanolic acid. Grasas y Aceites 2016, 67, 142–153. [Google Scholar] [CrossRef] [Green Version]
- Castro, A.J.G.; Frederico, M.J.S.; Cazarolli, L.H.; Mendes, C.P.; Bretanha, L.C.; Schmidt, É.C.; Bouzon, Z.L.; de Medeiros Pinto, V.A.; da Fonte Ramos, C.; Pizzolatti, M.G.; et al. The mechanism of action of ursolic acid as insulin secretagogue and insulinomimetic is mediated by cross-talk between calcium and kinases to regulate glucose balance. Biochim. Biophys. Acta BBA Gen. Subj. 2015, 1850, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, S.; Tripathi, R.; Mohammed, A.; Rizvi, S.I.; Mishra, N. Effects of thymol supplementation against type 2 diabetes in streptozocin- induced rat model. Plant. Arch. 2020, 20, 863–869. [Google Scholar]
- Qi, L.-W.; Liu, E.-H.; Chu, C.; Peng, Y.-B.; Cai, H.-X.; Li, P. Anti-diabetic agents from natural products—an update from 2004 to 2009. Curr. Top. Med. Chem. 2010, 10, 434–457. [Google Scholar] [CrossRef]
- Park, J.; Jang, H.-J. Anti-diabetic effects of natural products an overview of therapeutic strategies. Mol. Cell. Toxicol. 2017, 13, 1–20. [Google Scholar] [CrossRef]
- Belete, T.M. A recent achievement in the discovery and development of novel targets for the treatment of type-2 diabetes mellitus. J. Exp. Pharmacol. 2020, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Osadebe, P.O.; Odoh, E.U.; Uzor, P.F. Natural products as potential sources of antidiabetic drugs. J. Pharm. Res. Int. 2014, 4, 2075–2095. [Google Scholar] [CrossRef]
- Jugran, A.K.; Rawat, S.; Devkota, H.P.; Bhatt, I.D.; Rawal, R.S. Diabetes and plant-derived natural products: From ethnopharmacological approaches to their potential for modern drug discovery and development. Phytother. Res. 2021, 35, 223–245. [Google Scholar] [CrossRef]
- Li, W.; Yuan, G.; Pan, Y.; Wang, C.; Chen, H. Network pharmacology studies on the bioactive compounds and action mechanisms of natural products for the treatment of diabetes mellitus: A review. Front. Pharmacol. 2017, 8, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papoutsis, K.; Zhang, J.; Bowyer, M.C.; Brunton, N.; Gibney, E.R.; Lyng, J. Fruit, vegetables, and mushrooms for the preparation of extracts with α-amylase and α-glucosidase inhibition properties: A review. Food Chem. 2020, 338, 128119. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Miao, M. Dietary polyphenols modulate starch digestion and glycaemic level: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 541–555. [Google Scholar] [CrossRef]
- Sayem, A.S.M.; Arya, A.; Karimian, H.; Krishnasamy, N.; Ashok Hasamnis, A.; Hossain, C.F. Action of phytochemicals on insulin signaling pathways accelerating glucose transporter (GLUT4) protein translocation. Molecules 2018, 23, 258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proença, C.; Freitas, M.; Ribeiro, D.; Sousa, J.L.; Carvalho, F.; Silva, A.M.; Fernandes, P.A.; Fernandes, E. Inhibition of protein tyrosine phosphatase 1B by flavonoids: A structure-activity relationship study. Food Chem. Toxicol. 2018, 111, 474–481. [Google Scholar] [CrossRef]
- Zhao, B.T.; Nguyen, D.H.; Le, D.D.; Choi, J.S.; Min, B.S.; Woo, M.H. Protein tyrosine phosphatase 1B inhibitors from natural sources. Arch. Pharm. Res. 2018, 41, 130–161. [Google Scholar] [CrossRef]
- Jung, H.A.; Paudel, P.; Seong, S.H.; Min, B.-S.; Choi, J.S. Structure-related protein tyrosine phosphatase 1B inhibition by naringenin derivatives. Bioorg. Med. Chem. Lett. 2017, 27, 2274–2280. [Google Scholar] [CrossRef]
- Oh, Y.S. Plant-derived compounds targeting pancreatic beta cells for the treatment of diabetes. Evid. Based Complement. Altern. Med. 2015, 2015, 629863. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Park, S.-Y. Can antioxidants be effective therapeutics for type 2 diabetes? Yeungnam Univ. J. Med. 2021, 38, 83. [Google Scholar] [CrossRef]
- Matos, A.L.; Bruno, D.F.; Ambrósio, A.F.; Santos, P.F. The benefits of flavonoids in diabetic retinopathy. Nutrients 2020, 12, 3169. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Chaturvedi, N.; Singh, A.; Mishra, A. Investigation of hypoglycemic effects, oxidative stress potential and xanthine-oxidase activity of polyphenols (gallic acid, catechin) derived from faba bean on 3T3-L1 cell line: Insights into molecular docking and simulation study. Toxicol. Res. 2020, 9, 308–322. [Google Scholar] [CrossRef] [PubMed]
- Nwabueze, P.O.; Ekeuku, S.O.; Chan, H.K.; Eluri, K.; Fromming, G.R.A. Palmatine inhibits up-regulation of GRP78 and CALR protein in an STZ-induced diabetic rat model. Curr. Pharm. Biotechnol. 2021, 22, 288–298. [Google Scholar]
Figure 1.
Carbohydrate metabolism in the gastrointestinal tract.

Figure 2.
Carbohydrate balance.

Figure 3.
Mechanism of insulin exocytosis.

Figure 4.
Insulin signal transduction, 4E-BP1/2: eukaryotic translation initiation factor 4E binding protein/threonine-protein kinase; AMPK: AMP-activated protein kinase α; AP-1: activator protein 1 transcription factor; c-Raf: RAF proto-oncogene serine/threonine-protein kinase; DEPTOR: DEP domain-containing mTOR-interacting protein; eIF4G: eukaryotic translation initiation factor 4 gamma; eNOS: endothelial nitric oxide synthase; Erk1/2: extracellular signal-regulated kinases 1/2; FoxO: forkhead box O transcription factors; Gab: GRB2-associated-binding protein; GLUT2 or 4: glucose transporter type 2 or 4; GPR120: G-protein coupled receptor 120; GRB2: growth factor receptor-bound protein 2; GS: glycogen synthase; GSK-3: glycogen synthase kinase 3; GβL: G protein beta subunit-like; IKK: IκB kinase; IRS-1: insulin receptor substrate 1; JNK: c-Jun N-terminal kinase; LKB1: liver kinase B1; MEK1/2: mitogen-activated protein kinase 1/2; mTORC: mechanistic target of rapamycin; NF-κB; nuclear factor kappa-light-chain-enhancer of activated B cells; NO: nitrogen oxide; p70S6K: p70S6 kinase PDK1 phosphoinositide-dependent kinase-1; PI3K: phosphoinositide 3-kinase; PKCζ: protein kinase C zeta type; PPAR: peroxisome proliferator-activated receptor; PRR5: proline-rich protein 5; Raptor: regulatory-associated protein of mTOR; Rheb: Ras homolog enriched in brain; Rictor: rapamycin-insensitive companion of mammalian target of rapamycin; RIP: receptor-interacting protein kinases; Sin1: stress-activated map kinase interacting protein 1; SNARE: soluble NSF attachment protein receptor; SOS: son of sevenless protein; SREBP-1: sterol regulatory element-binding protein 1; Synip: syntaxin 4-interacting protein; TAB-1: TGF-beta activated kinase 1.
Figure 4.
Insulin signal transduction, 4E-BP1/2: eukaryotic translation initiation factor 4E binding protein/threonine-protein kinase; AMPK: AMP-activated protein kinase α; AP-1: activator protein 1 transcription factor; c-Raf: RAF proto-oncogene serine/threonine-protein kinase; DEPTOR: DEP domain-containing mTOR-interacting protein; eIF4G: eukaryotic translation initiation factor 4 gamma; eNOS: endothelial nitric oxide synthase; Erk1/2: extracellular signal-regulated kinases 1/2; FoxO: forkhead box O transcription factors; Gab: GRB2-associated-binding protein; GLUT2 or 4: glucose transporter type 2 or 4; GPR120: G-protein coupled receptor 120; GRB2: growth factor receptor-bound protein 2; GS: glycogen synthase; GSK-3: glycogen synthase kinase 3; GβL: G protein beta subunit-like; IKK: IκB kinase; IRS-1: insulin receptor substrate 1; JNK: c-Jun N-terminal kinase; LKB1: liver kinase B1; MEK1/2: mitogen-activated protein kinase 1/2; mTORC: mechanistic target of rapamycin; NF-κB; nuclear factor kappa-light-chain-enhancer of activated B cells; NO: nitrogen oxide; p70S6K: p70S6 kinase PDK1 phosphoinositide-dependent kinase-1; PI3K: phosphoinositide 3-kinase; PKCζ: protein kinase C zeta type; PPAR: peroxisome proliferator-activated receptor; PRR5: proline-rich protein 5; Raptor: regulatory-associated protein of mTOR; Rheb: Ras homolog enriched in brain; Rictor: rapamycin-insensitive companion of mammalian target of rapamycin; RIP: receptor-interacting protein kinases; Sin1: stress-activated map kinase interacting protein 1; SNARE: soluble NSF attachment protein receptor; SOS: son of sevenless protein; SREBP-1: sterol regulatory element-binding protein 1; Synip: syntaxin 4-interacting protein; TAB-1: TGF-beta activated kinase 1.

Figure 5.
Glycolysis.

Figure 6.
Krebs cycle.

Figure 7.
Gluconeogenesis.

Figure 8.
Glycogen metabolism, synthesis and hydrolysis.


Figure 9.
Insulin effect on adipose tissue and muscle tissue.

Figure 10.
Antidiabetic activity of naturally occurring alkaloids.

Figure 11.
Antidiabetic activity of simple phenols.

Figure 12.
Anthocyanins and their protective role against diabetes.

Figure 13.
Antidiabetic activity of stilbenoids.

Figure 14.
Antidiabetic activities of saponins.

Figure 15.
Polysaccharides effects on glucose homeostasis.

Figure 16.
Coumarins effects on glucose homeostasis.

Figure 17.
Antidiabetic activity of terpenes.

Table 1.
Effects of insulin on several organs and proteins.
| Target | Glucose Transporter | Hepatic Enzymes | Β-Cells Apoptosis | PPAR | AMPK | Tyrosine Kinase Inhibitor | NF-κB | |
|---|---|---|---|---|---|---|---|---|
| Normal effects | Enhance glucose uptake by tissues | Enhance glucose metabolism | Control program cell death | Control of lipid metabolism | Energy homeostasis | Control growth factor signaling | Control of β-cells survival | |
| Changes by diabetes |  | GLUT translocation Glucose uptake | Insulin signaling Liver glycogen | Apoptotic regulatory genes Caspases | Lipid metabolism | AMPK activation Glucose homeostasis | Insulin sensitivity Insulin secretion | |
 | Insulin resistance | Insulin resistance Glucose production | Oxidative stress Insulin resistance Mitochondria dysfunction | Hyperglycemia Hyperlipidemia hyperinsulinemia | Insulin resistance | Islet cell function | Proinflammatory cytokines Oxidative stress NF-κB expression | |
Table 11.
Proposed antidiabetic effects of different stilbenoids.
| Name of Compound | Chemical Structure | Mechanism | References |
|---|---|---|---|
| Resveratrol |  |
| [10,102,149] |
| Pterostilbene |  |
| [10,150] |
Table 12.
In vitro, in vivo and in silico models of different isolated stilbenoids.
| Compound | Mechanism | Dose of the Tested Compound (Duration) | Model | References |
|---|---|---|---|---|
| Resveratrol | Suppressed oxidative stress and increased potential to internalize glucose by extrahepatic tissues. | 20 mg/kg (8 weeks) | Streptozocin-induced diabetic rats | [151] |
| Pterostilbene | Ameliorated morphological impairment of the pancreas, increased the protein expression of PPARγ, PI3K, p-Akt, GLUT4 and IRS-1 in adipose tissues. | 20, 40, 80 mg/kg (8 weeks) | Streptozocin-induced diabetic rats | [150] |
| Activated Nrf2, thereby reducing oxidative damage, reverted hexokinase, glucose-6-phosphatase, glucose-6-phosphate dehydrogenase, and fructose-1,6-bisphosphatase, to near-normal levels, improved insulin secretion. | 5, 10 mg/kg (5 weeks) | In vitro: MIN-6 cell line, streptozocin-induced diabetic rats | [152] |
Table 14.
In vitro, in vivo and in silico models of different isolated saponins.
| Compound | Mechanism | Dose of the Tested Compound (duration) | Model | References |
|---|---|---|---|---|
| Ginsenoside K | Inhibited the expression of PEPCK and G6Pase enzymes, increased the activation of AMPK. | 1, 2, 4, 8 μM, (24 h) 30 mg/kg, (4 weeks) | In vitro: HepG2 cell line, streptozocin-induced diabetic mice | [162] |
| Inhibited inflammation and improved insulin signaling in adipose tissue by suppressing ER stress-associated NLRP3 inflammation activation. | 10 µM (24 h) | In vitro: T3-L1 cell line, mice model | [163] | |
| Ginsenoside Rb1 | Increased GLUT-4 translocation through up-regulated adipoR1 and adipoR2 gene. | 0.001–100 mM (1–12 h) | In vitro: C2C12 myotubes cell line | [164] |
| Exhibited insulin-sensitizing effect. | 20 mg/kg (14 days) | Diabetic mice | [165] | |
| Reduced hepatic glucose production, increased glucose uptake in skeletal muscle. | 10 mg/kg every other day (One week) | High-fat-induced diabetic mice | [166] | |
| Inhibited inflammation and improved insulin signaling in adipose tissue by suppressing ER stress-associated NLRP3 inflammation activation. | 10 µM (24 h) | In vitro: T3-L1 cell line, mice model | [163] | |
| Ginsenoside Rg1 | Reduced inflammation by inhibiting JNK activity, reduced caspase-3 and BAX (proapoptotic) proteins, increased BCL-2 (antiapoptotic) protein. | 25, 50 mg/kg (4 weeks) | Streptozocin-induced diabetic rats | [167] |
| Ginsenoside RK3 | Inhibited hepatic gluconeogenesis (inhibited PEPCK and G6pase protein expressions). | 10, 30, 60 mg/kg (4 weeks) | In vitro: HepG2 cell line, high-fat diet and streptozocin-induced mice | [168] |
Table 15.
Antidiabetic effects of different tannins.
| Name of Compound | Mechanism | References |
|---|---|---|
| Tannic acid |
| [170,171] |
| Condensed tannins |
| [172] |
| Hydrolyzable tannins |
| [173] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shehadeh, M.B.; Suaifan, G.A.R.Y.; Abu-Odeh, A.M. Plants Secondary Metabolites as Blood Glucose-Lowering Molecules. Molecules 2021, 26, 4333. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26144333
AMA Style
Shehadeh MB, Suaifan GARY, Abu-Odeh AM. Plants Secondary Metabolites as Blood Glucose-Lowering Molecules. Molecules. 2021; 26(14):4333. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26144333
Chicago/Turabian StyleShehadeh, Mayadah Bashir, Ghadeer A. R. Y. Suaifan, and Ala’ Mustafa Abu-Odeh. 2021. "Plants Secondary Metabolites as Blood Glucose-Lowering Molecules" Molecules 26, no. 14: 4333. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26144333