
Sensitivity of Botrytis cinerea Isolates Complex to Plant Extracts


Laboratory of Plant Protection, Institute of Horticulture, Lithuanian Research Centre for Agriculture and Forestry, Kauno st. 30, LT-54333 Babtai, Lithuania
*
Author to whom correspondence should be addressed.
Molecules 2021, 26(15), 4595; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26154595
Submission received: 7 June 2021
/
Revised: 10 July 2021
/
Accepted: 26 July 2021
/
Published: 29 July 2021
(This article belongs to the Special Issue Chitosan and Natural Compounds for the Control of Preharvest and Postharvest Diseases of Horticultural Commodities)
Abstract
:New agricultural strategies aim to reduce the use of pesticides due to their damage to the environment and humans, and the caused resistance to pathogens. Therefore, alternative sources of antifungal compounds from plants are under investigation lately. Extracts from plants have a wide composition of chemical compounds which may complicate the development of pathogen resistance. Botrytis cinerea, causing grey mould, is an important horticultural and ornamental pathogen, responsible for the relevant yield and quality losses. B. cinerea isolated from a different plant host may differ in the sensitivity to antifungal substances from plants. Assessing the importance of research covering a wide range of pathogens for the rapid development of biopesticides, this study aims to determine the sensitivity of the B. cinerea isolate complex (10 strains) to plant extracts, describe morphological changes caused by the extract treatment, and detect differences between the sensitivity of different plant host isolates. The results showed the highest sensitivity of the B. cinerea isolates complex to cinnamon extract, and the lowest to laurel extract. In contrast, laurel extract caused the most changes of morphological attributes in the isolates. Five B. cinerea isolates from plant hosts of raspberry, cabbage, apple, bell pepper, and rose were grouped statistically according to their sensitivity to laurel extract. Meanwhile, the bell pepper isolate separated from the isolate complex based on its sensitivity to clove extract, and the strawberry and apple isolates based on their sensitivity to cinnamon extract.
1. Introduction
New strategies for reducing pesticide use are an inevitable, however, scientifically and practically demanding challenge. In the case of fungicides, it is already known that they have adverse effects on the environment and humans [1,2]. In the long run, they also caused pathogen resistance [3,4]. If not adequately managed, widespread disease control methods can become ineffective in the long run. With all the knowledge about pesticide residues in food, consumers tend to choose higher quality and more environmentally and human-safe products [5]. Therefore, alternative sources of antifungal substances are under investigation lately [6,7,8,9,10,11]. One alternative source is active compounds obtained from plants [12], such as botanical pesticides.
One of the advantages of plant-based pesticides is their biological activity from many accumulated chemical compounds, which result in a less expected pathogen resistance [13]. The sensitivity of pathogens to active compounds from plants has been described to some extent in the literature [2,7,14,15]. Morphological changes in fungal structures were observed after treatment with alternative antifungal compounds: salt solutions [16], zinc oxide nanoparticles [6], specific light spectrum [17], essential oils [1] or extracts [18], and their active components [19]. As demand for the replacement of synthetic fungicides is extremely high [14], in vitro antifungal studies should be expanded/continued to group plant materials according to their antifungal effect on the pathogen complex. This would increase the research and development of high-impact bioactive substances from plants against fungal pathogens.
Botrytis cinerea is known to develop fungicide resistance [4,20,21,22]. This pathogen infects many plant species and causes grey mould [23,24,25] during the growth phase and after harvest [18]. It is responsible for one of the most significant economic losses of horticultural crop production [1,4,10]. Genetic differences between B. cinerea cryptic species group II isolates from different plant hosts were indicated in previous studies [23]. Therefore, we would like to emphasize the importance of evaluating the sensitivity of isolates from different plant hosts or even different plant parts in the search for new active substances, as this pathogen is not host-specific, and is virulent independently from plant species [26]. Natural substances with broad antifungal properties, affecting a wide range of pathogens, harmful to various plants, should be considered for further studies as this would fasten the development of new products that could equate and supplement the fungicides used for B. cinerea control nowadays.
Therefore, the study of pathogens from different plant hosts should become an integral part of plant bioactive substances studies. As B. cinerea is harmful to various horticultural and ornamental crops [25], our study focused on isolates from several plant host species. Furthermore, we hypothesise that plant extracts cause morphological changes to pathogen colonies, which could be noticeable visually. An initial visual assessment of the pathogen is important for further, in-depth studies to determine the mechanism of action of the active substances. Our study aimed to determine the sensitivity of the B. cinerea isolate complex to plant extracts, evaluate the morphological changes caused by the extract treatment, and detect the differences between the sensitivity of different plant host isolates. The collected data can be further used to develop plant-based antifungal products with significant effects on B. cinerea.
2. Results
2.1. Sensitivity of B. cinerea Isolates Complex to Plant Extracts
This study investigated the sensitivity of the B. cinerea isolates complex to plant extracts under different concentrations. The isolates complex showed the highest sensitivity to cinnamon extract. All isolates showed no visual growth at an 800 µL/L concentration, 60% at 600 µL/L concentration, and 20% at 500 µL/L concentration (Table 1). B. cinerea isolates BC1 (strawberry) and BC6 (apple) were sensitive to the lowest concentration of cinnamon extract. After treatment with 800 µL/L of clove extract, the growth of 70% of the isolates was suppressed (radial colony growth was equal zero, Table 1). The isolate from the strawberry was the most sensitive to clove extract as the growth B. cinerea from this plant host was not visible from 500 µL/L concentration. Meanwhile, 30% of the isolates (plant hosts—apple, leek, and rose) still expressed growth at the highest investigated concentration of clove extract (800 µL/L). Opposite to clove and cinnamon extracts, the B. cinerea complex of isolates was not sensitive to laurel extract, 4200 µL/L concentration was not efficient to inhibit none of the isolates from ten plant hosts. However, B. cinerea from strawberry and apple showed sensitivity to laurel extract at earlier days of investigation. Overall, these two isolates had the highest sensitivity to extracts, examined in this study. Statistical differences in each isolate’s sensitivity to different concentrations of the extract are presented in Table 1.
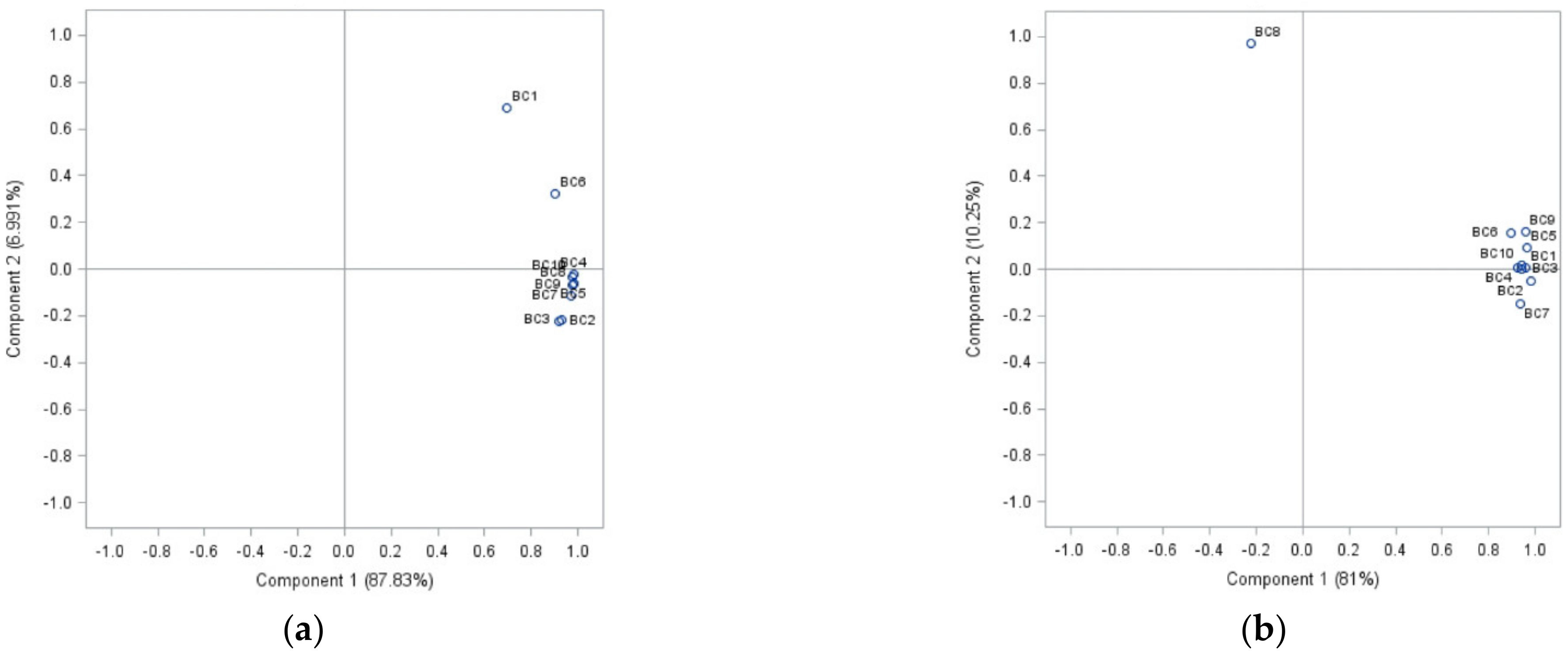
An examination of the Botrytis cinerea isolate complex with PCA showed a visualization of differences between the sensitivity of isolates belonging to the complex used in this study after treatment with cinnamon, clove, and laurel extracts (Figure 1).
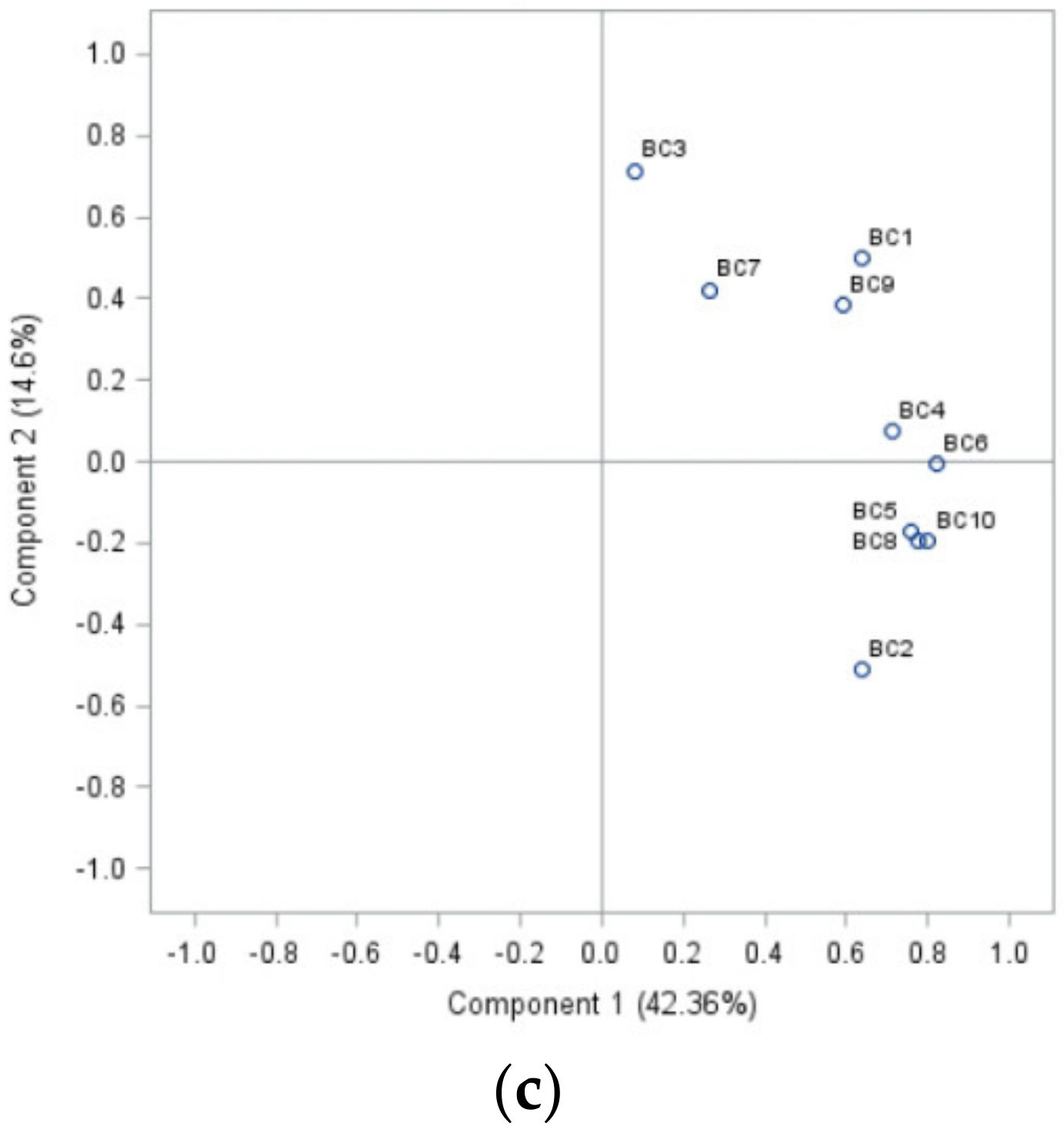
Analysing the sensitivity of isolates to cinnamon extract, the first principal component described 87.83% of the variance (Figure 1a). Isolates BC1 (strawberry) and BC6 (apple) separated from the other isolates group according to their sensitivity to different concentrations of cinnamon extract. An analysis of treatment with clove extract, PCA1, represented 81% of the data, and PCA2 10.25% (91.25% overall) (Figure 1b). B. cinerea from bell pepper (BC8) differed from other isolates’ sensitivity compared to the sensitivity to clove extract. Half of the isolates formed a group based on their similarities and differences in laurel extract sensitivity. Strawberry and leek isolates (BC1 and BC9) formed a separate group. Additionally, individual differences were indicated in onion (BC2), tomato (BC3) and grape (BC7) isolates sensitivity to laurel extract. In this case, 56.96% of total data variance was explained (PCA1 and PCA2—42.36% and 14.6%, respectively) (Figure 1c).
2.2. Morphological Attributes of the Isolates
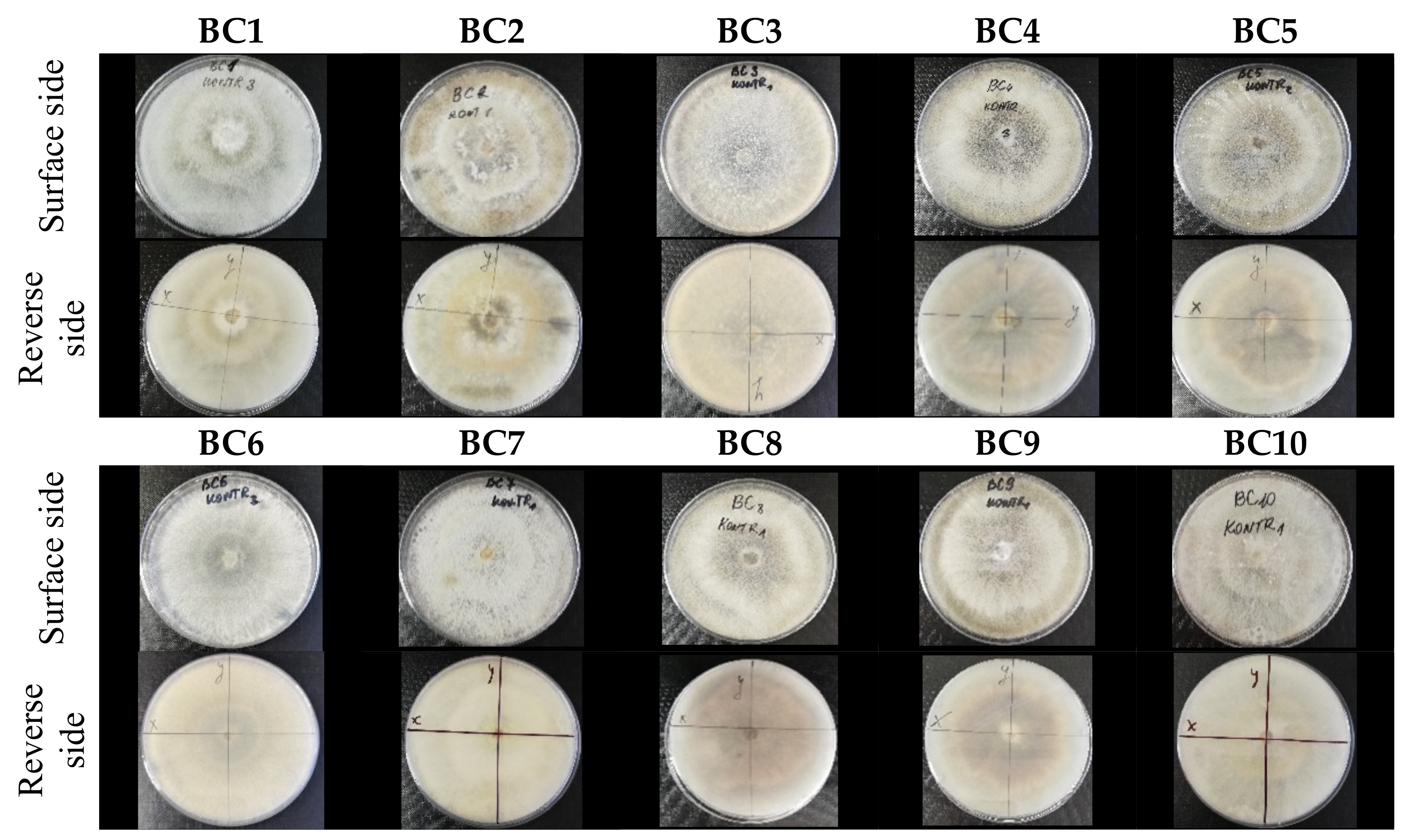
Botrytis cinerea isolates with visible colony growth after treatment with extracts were compared to the control plates (Figure 2), where no extract was added to the medium. Observed changes of type of mycelium and colony colour are presented in Table 2. Clove and laurel extracts caused the most mycelium changes of the B. cinerea complex. With laurel extract, mycelium changes in isolate BC3 were observed from 3600 µL/L, and from 3200 µL/L in BC7. All investigated extracts similarly affected colour changes in isolate colonies. Colour changes were visible from treatment with 300 µL/L concentration of cinnamon extract, and from 3200 µL treatment with laurel extract. Furthermore, clove extract induced pigmentation of the medium in the most of B. cinerea plates, and treatment with cinnamon and laurel extracts had no significant effect.
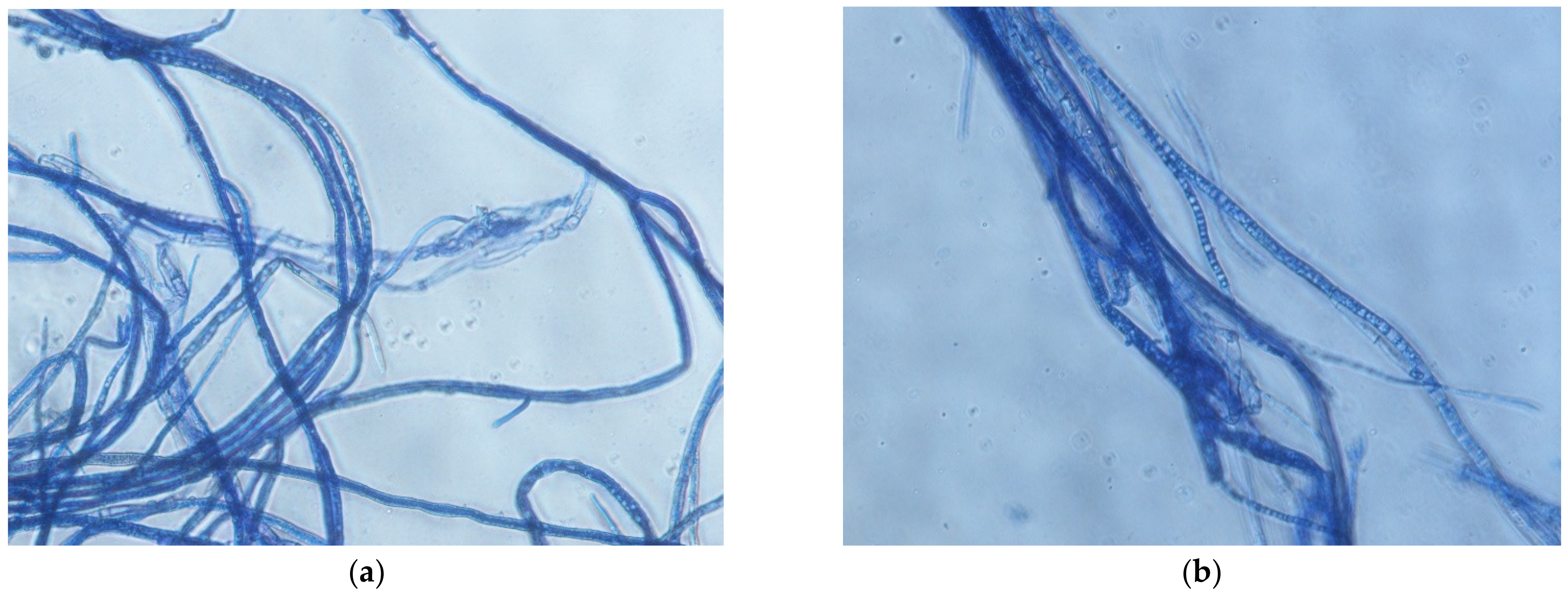
B. cinerea isolates complex colonies were examined for the presence of conidia. Conidia were detected in most of the isolates, except for BC1 (strawberry) and BC6 (apple) (Table 3). The conidia length and width in control colonies of B. cinerea isolates varied—17.36–20.91 µm and 13.55–15.82 µm, respectively. Overall, no significant differences were observed between the control colonies conidia size and cinnamon extract-treated colonies. An amount of 400 and 600 µL/L clove extract treatment resulted in smaller conidia in examined samples of BC2 isolate (plant host—onion). Meanwhile, other isolates produced conidia in a similar size as the control. Laurel extract-treated isolates varied in conidia size the most compared to all extract treatments. Significantly smaller conidia were found in BC2, BC4 and BC8 isolates (onion, raspberry, bell pepper) colonies, and the BC7 (grape) isolate had a higher length and width of conidia compared to the control. Shrunk, twisted hyphae with coagulated inner structures were observed under the microscope after treatment with different concentrations of the examined extracts. An example of the B. cinerea hyphae after treatment with laurel extract is shown in Figure 3.
3. Discussion
In previously reported studies, the sensitivity of B. cinerea from various plant hosts was investigated and antifungal concentrations of extracts and essential oils were determined [1,10,11,14,15]. Mycelial growth of B. cinerea from tomato was inhibited by origanum essential oil at 12.8 µg/mL and lavender and rosemary essential oils at 25.6 µg/mL [1]. In another tomato B. cinerea study, the pathogen expressed high sensitivity to 400 and 500 mg/mL concentration of the Vernonia amygdalina dichloromethane extract [18]. Strawberry B. cinerea showed sensitivity to Cymbopogon martinii and Mentha spicata essential oils between 250 and 500 µL/L [10], and to Aloysia citriodora, C. winterianus, Lippia alba and Ocimum americanum oils at 0.8–1.4 mL/L [5].
In our study, the B. cinerea pathogen complex, consisting of ten strains from different plant hosts, was strongly sensitive to clove and cinnamon extract in vitro. Isolates from strawberry and apple (BC1 and BC6) significantly differed from other isolates in the complex as they expressed a sensitivity to the lowest concentration of cinnamon extract. The strawberry isolate (BC1) was also more sensitive to clove extract; however, other isolates were inhibited only with the highest tested concentration or still expressed growth. These varied sensitivity results illustrate the need for the investigation of more than one strain of the pathogen as the range of the antifungal concentration of the extract was observed individual to every plant host isolate and may be too low to inhibit all of the complex. Similarly, to our results, four B. cinerea isolates (plant hosts lettuce, strawberry, cucumber, and pepper) had a high sensitivity to cinnamon extract in Wahab et al.’s [8] study. The full inhibition of B. cinerea (kiwi fruit) growth was reached at 1000 µg/mL of laurel essential oil, extracted with supercritical carbon dioxide [14], and the growth of Aspergillus carbonarius was stopped with 0.3% of laurel essential oil [2]. In contrast, neither of the isolates from our B. cinerea complex were sensitive to laurel extract. Despite relatively high tested concentrations, it should be increased in further in vitro studies.
Isolates BC4, BC5, BC6, BC8, and BC10 (plant hosts: raspberry, cabbage, apple, bell pepper, and rose) were grouped statistically according to their growth feature after laurel extract treatment with different concentrations. Three of the isolates in the formed group had the transposa genotype, and two Vacuma. Meanwhile, only the bell pepper isolate (BC8) separated from the isolate complex after treatment with different clove extract concentrations—and strawberry (BC1) and apple (BC6) isolates—after treatment with different cinnamon extract concentrations. Our isolate complex had a higher sensitivity to cinnamon and clove extract compared to the one described in da Silva et al.’s [11] study; there, B. cinerea isolated from strawberries was inhibited by essential oils of Eucalyptus staigeriana and. E. urograndis using concentrations of 2000 and 4000 µL/L, respectively. Completely inhibited growth of B. cinerea from grapevine fruit was also observed at a lower—1500 µL/L—concentration of Eucalyptus sp. and Zhumeria majdae oils in another study [15]. Meanwhile, no sensitivity was observed at the highest (4200 µL/L) concentration of laurel extract, considering it less effective against B. cinerea than oils of the Eucalyptus genus. Additionally, B. cinerea was sensitive to lower or equal concentrations of cinnamon and clove extracts, compared to the antifungal concentrations of various essential oils described by Fontana et al. [5].
Considering that B. cinerea demonstrated morphological differences while grown on a different medium [24], changes of morphological attributes could be expected for isolates grown on a medium with an additional factor—plant extract. Although the B. cinerea isolates complex was highly sensitive to cinnamon extract, only a few isolates showed changed mycelium—even fewer changed the colour of the colonies. On the contrary, most complex isolates reacted to laurel extract treatment by changing the mycelium structure and colour despite the low sensitivity. We can assume that the possible antifungal mechanism of laurel extract was active as morphological attributes were affected, however, not strong enough to inhibit the growth of the pathogen. A changed surface and other features of B. cinerea hyphae were detected before [6,19]. In other investigations, structural alterations of B. cinerea hyphae were detected after exposure with Eucalyptus essential oils, causing the development of wrinkles, torsion, peeling and dehydration, and authors of the study suggested that hyphae changes are caused regarding essential oils action on the cell wall of the pathogen [11]. Our primary visual assessment of B. cinerea hyphae after treatment with extracts agree with other studies, as several changes were observed under microscope.
The conidia size in our control colonies of B. cinerea isolates varied and were smaller than described of the isolates in Kumari et al.’s [26] study. However, this feature may depend on each isolate’s individuality and the environmental conditions, including geographical region. Examples of how treatment with essential oils results in degraded hyphae with changed or damaged cell structures could be found in the literature [1]. Shrunk conidia, wrinkled, and deformed hyphae were observed on B. cinerea after V. amygdalina dichloromethane extract treatment [18]. Overall, cinnamon and clove extracts did not reduce the conidia size of the isolates complex in our study, except in the clove extract and isolate from the onion case. However, it was observed that there was a tendency after laurel extract treatment that half of the isolates germinated significantly smaller conidia, mostly at 2600 µL/L concentration. Yusoff et al. [18] suggest that the shrinkage of conidia could prevent the dispersion of the grey mould disease. To confirm the opposing results of the laurel extract impact on the morphology of B. cinerea, deeper morphological studies should be continued. In contrast to our study, no conidiation was observed in oil-treated samples of tomato B. cinerea [1].
To determine materials that may supplement chemical fungicides in the future, the effect on a wide range of plant host pathogens is essential. That is the reason why in this work we evaluated the sensitivity of the fungal pathogen B. cinerea complex, consisting of ten strains from different plant hosts to plant extracts, aiming to determine the possible plant-based materials with a significant impact on the control of horticultural and ornamental pathogens.
4. Materials and Methods
4.1. Botrytis Cinerea Isolates Complex
As Botrytis cinerea is a harmful pest of horticultural and ornamental crops, isolates from various plant hosts were selected as an investigation object. Ten single-spore B. cinerea isolates (Table 4) belonging to the cryptic species group I (resistant to fenhexamid) and group II (sensitive to fenhexamid) were selected for the study with the purpose to investigate their sensitivity to potential biopesticides—plant extracts.
Single-spore isolates were obtained from infected parts of plants with visual symptoms of grey mould. Species and phylogenetic identification were performed under conditions specified in the study by Rasiukevičiūtė et al. [27]. The transposable element type of each isolate is presented in Table 1. Single spore isolates were stored in the Lithuanian Research Centre for Agriculture and Forestry, Institute of Horticulture, Plant protection laboratory pathogen collection, and renewed on the PDA two weeks before the start of the experiments.
4.2. Plant Extracts
Dried cinnamon bark (Cinnamomum zeylanicum), clove buds (Syzygium aromaticum) and laurel leaves (Laurus nobilis) were extracted using the CO2 extraction method as described in the previous study [28]. Composition of the volatile compounds responsible for antifungal activities of each extract was determined previously using gas chromatography/mass spectrometry [29]. The main component of the cinnamon extract was trans-cinnamaldehyde, clove—eugenol and eugenol acetate—, and laurel—eucalyptol and alpha-terpinyl acetate [29,30].
4.3. Determination of the Sensitivity to the Extracts
The sensitivity of B. cinerea isolates complex was tested for the following concentrations: cinnamon extract—100–800 µL/L—, clove extract—100–800 µL/L—, and laurel extract—2600–4200 µL/L. The experiment was designed with four replicates per each treatment concentration. Sterile PDA medium was prepared as specified in the manufacturer’s instruction and the appropriate amounts of the extracts were separately poured into the medium and mixed on the rotary shaker for 10 min. Prepared extract-enriched medium was poured into Petri dishes. Previously cut 7 mm diameter mycelium plug of B. cinerea (each isolate separately) was put in the centre of the Petri dish. No extracts were added to the control plates of the isolates. Plates with mycelium plugs were placed for incubation at 22 °C in the dark for seven days. After seven days, radial colony growth (cm) of the B. cinerea isolates was measured. If radial colony growth was significantly reduced compared to control treatment, isolate expressed sensitivity to the tested concentration of the extract. If radial colony growth was fully reduced (equal to zero), isolate was considered as highly sensitive to the treatment.
4.4. Examination of Morphological Attributes of the Isolates Complex
Only concentrations with visual pathogen growth were included in the morphological attributes’ evaluation. Seven days after inoculation, fungal colonies treated with different extract concentrations visually compared to the ones in the control plates. Examined morphological attributes were mycelium and colour, evaluating the front and reverse sides of the colonies. Differences from the control plate were marked as described:
Mycelium:
(+)—extract treatment caused mycelium changes compared to the control plate.
(–)—fungal colony in the extract-treated plate matches the type of mycelium with the control plate.
Colour:
(+)—extract treatment caused different pigmentation compared to the control plate.
(–)—fungal colony in the extract-treated plate matches the colour with the control plate.
Furthermore, all colonies with visual fungal growth were examined for the presence of conidia. An amount of 1 mm of fungal mycelium was taken from the margin of the colony with a sterile needle and placed in 50% lactophenol blue solution on a microscope glass slide, mixed and covered with a glass coverslip and examined at 40× magnification using a phase-contrast light microscope Nikon Eclipse 80i (Nikon Instrument Inc., Melville, NY, USA). In case conidia were formed, the length and width (µm) of 30 conidia were measured using NIS-Elements D 3.2 programme (Nikon Instrument Inc., Melville, NY, USA). Four measurements per treatment were performed. The same program was used to capture observed morphological changes of hyphae.
4.5. Statistical Analysis
Statistical analysis was performed with the SAS Enterprise Guide programme (SAS Institute Inc., Cary, NC, USA)). Differences between means were analysed using Duncan Multiple Range Test (p < 0.05). Principal component analysis was also performed to group the analysed isolates of Botrytis cinerea complex according to the sensitivity to each of the extracts.
5. Conclusions
In conclusion, the study revealed varied sensitivity of the complex of different plant hosts B. cinerea isolates from cryptic species group I and II to clove, cinnamon, and laurel extracts. Isolates had the highest sensitivity to cinnamon extract as their growth was completely reduced at the lowest concentrations from all tested extracts. Different plant hosts B. cinerea isolates were grouped statistically according to their sensitivity to plant extracts. Morphological attribute changes (colour, mycelium, and conidia size) after treatment with extracts were described. Contrasting results were obtained in laurel extract, as this extract had the most impact on the changes of morphological attributes. However, the isolates complex was not susceptible to the investigated concentrations. This information gives possible directions for further studies evaluating the antifungal mechanism of oils. It is emphasized that the advantage of natural products from plants is their multicomponent composition providing a lower possibility for the pathogen to develop resistance quickly. Based on the results in our study, the cinnamon extract has promising antifungal properties, causing sensitivity to B. cinerea, infecting various plant hosts.
Author Contributions
Conceptualization, L.D. and A.V.; methodology, L.D.; software, L.D.; validation, L.D.; formal analysis, L.D.; investigation, L.D.; resources, A.V.; data curation, A.V.; writing—original draft preparation, L.D.; writing—review and editing, L.D. and A.V.; visualization, L.D.; supervision, A.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors express a gratitude to Neringa Rasiukevičiūtė from the Lithuanian Research Centre for Agriculture and Forestry for consultations regarding the experiments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Soylu, E.M.; Kurt, Ş.; Soylu, S. In vitro and in vivo antifungal activities of the essential oils of various plants against tomato grey mould disease agent Botrytis cinerea. Int. J. Food Microbiol. 2010, 143, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Dammak, I.; Hamdi, Z.; El Euch, S.K.; Zemni, H.; Mliki, A.; Hassouna, M.; Lasram, S. Evaluation of antifungal and anti-ochratoxigenic activities of Salvia officinalis, Lavandula dentata and Laurus nobilis essential oils and a major monoterpene constituent 1,8-cineole against Aspergillus carbonarius. Ind. Crops Prod. 2019, 128, 85–93. [Google Scholar] [CrossRef]
- Saito, S.; Xiao, C.L. Fungicide resistance in Botrytis cinerea populations in California and its influence on control of gray mold on stored mandarin fruit. Plant. Dis. 2018, 102, 2545–2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbey, J.A.; Percival, D.; Abbey, L.; Asiedu, S.K.; Prithiviraj, B.; Schilder, A. Biofungicides as alternative to synthetic fungicide control of grey mould (Botrytis cinerea)–prospects and challenges. Biocontrol Sci. Technol. 2018, 29, 207–228. [Google Scholar] [CrossRef]
- Fontana, D.C.; Neto, D.D.; Pretto, M.M.; Mariotto, A.B.; Caron, B.O.; Kulczynski, S.M.; Schmidt, D. Using essential oils to control diseases in strawberries and peaches. Int. J. Food Microbiol. 2021, 338, 108980. [Google Scholar] [CrossRef]
- He, L.; Liu, Y.; Mustapha, A.; Lin, M. Antifungal activity of zinc oxide nanoparticles against Botrytis cinerea and Penicillium expansum. Microbiol. Res. 2011, 166, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Morcia, C.; Malnati, M.; Terzi, V. In vitro antifungal activity of terpinen-4-ol, eugenol, carvone, 1,8-cineole (eucalyptol) and thymol against mycotoxigenic plant pathogens. Food Addit. Contam. Part A 2012, 29, 415–422. [Google Scholar] [CrossRef] [Green Version]
- Wahab, H.A.; Malek, A.; Ghobara, M. Effects of some plant extracts, bioagents, and organic compounds on Botrytis and Sclerotinia molds. Acta Agrobot. 2020, 73, 7321. [Google Scholar] [CrossRef]
- Oliveira, M.S.; Costa, W.A.; Pereira, D.S.; Botelho, J.R.S.; Menezes, T.O.A.; Andrade, E.H.A.; Silva, S.H.M.; Filho, A.P.S.S.; Carvalho Junior, R.N. Chemical composition and phytotoxic activity of clove (Syzygium aromaticum) essential oil obtained with supercritical CO2. J. Supercrit. Fluids 2016, 118, 185–193. [Google Scholar] [CrossRef]
- De Oliveira Filho, J.G.; da Cruz Silva, G.; de Aguiar, A.C.; Cipriano, L.; de Azeredo, H.M.C.; Bogusz Junior, S.; Ferreira, M.D. Chemical composition and antifungal activity of essential oils and their combinations against Botrytis cinerea in strawberries. J. Food Meas. Charact. 2021, 15, 1815–1825. [Google Scholar] [CrossRef]
- Da Silva, P.P.M.; de Oliveira, J.; dos Biazotto, A.M.; Parisi, M.M.; da Glória, E.M.; Spoto, M.H.F. Essential oils from Eucalyptus staigeriana F. Muell. ex Bailey and Eucalyptus urograndis W. Hill ex Maiden associated to carboxymethylcellulose coating for the control of Botrytis cinerea Pers. Fr. and Rhizopus stolonifer (Ehrenb.:Fr.) Vuill. in strawberr. Ind. Crops Prod. 2020, 156, 112884. [Google Scholar] [CrossRef]
- Pinto, L.; Bonifacio, M.A.; De Giglio, E.; Cometa, S.; Logrieco, A.F.; Baruzzi, F. Unravelling the antifungal effect of red thyme oil (Thymus vulgaris L.) compounds in vapor phase. Molecules 2020, 25, 4761. [Google Scholar] [CrossRef] [PubMed]
- Packiam, S.M. Green Pesticides: Eco-friendly Technology for Integrated Pest Management. Acta Sci. Agric. 2018, 2, 1. [Google Scholar]
- De Corato, U.; Maccioni, O.; Trupo, M.; Di Sanzo, G. Use of essential oil of Laurus nobilis obtained by means of a supercritical carbon dioxide technique against post harvest spoilage fungi. Crop. Prot. 2010, 29, 142–147. [Google Scholar] [CrossRef]
- Davari, M.; Ezazi, R. Chemical composition and antifungal activity of the essential oil of Zhumeria majdae, Heracleum persicum and Eucalyptus sp. against some important phytopathogenic fungi. J. Mycol. Med. 2017, 27, 463–468. [Google Scholar] [CrossRef]
- Youssef, K.; Roberto, S.R.; de Oliveira, A.G. Ultra-structural alterations in Botrytis cinerea—the causal agent of gray mold—treated with salt solutions. Biomolecules 2019, 9, 582. [Google Scholar] [CrossRef] [Green Version]
- Meng, L.; Mestdagh, H.; Ameye, M.; Audenaert, K.; Höfte, M.; Van Labeke, M.C. Phenotypic Variation of Botrytis cinerea Isolates Is Influenced by Spectral Light Quality. Front. Plant. Sci. 2020, 11, 1233. [Google Scholar] [CrossRef]
- Yusoff, S.F.; Haron, F.F.; Mohamed, M.T.M.; Asib, N.; Sakimin, S.Z.; Kassim, F.A.; Ismail, S.I. Antifungal activity and phytochemical screening of Vernonia amygdalina extract against Botrytis cinerea causing gray mold disease on tomato fruits. Biology 2020, 9, 286. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, J.; Chen, H.; Fan, Y.; Shi, Z. Antifungal activity of eugenol against Botrytis cinerea. Trop. Plant. Pathol. 2010, 35, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Veloukas, T.; Leroch, M.; Hahn, M.; Karaoglanidis, G.S. Detection and molecular characterization of boscalid-resistant Botrytis cinerea isolates from strawberry. Plant. Dis. 2011, 95, 1302–1307. [Google Scholar] [CrossRef] [Green Version]
- Cosseboom, S.D.; Schnabel, G.; Hu, M. Competitive ability of multi-fungicide resistant Botrytis cinerea in a blackberry planting over three years. Pestic. Biochem. Physiol. 2020, 163, 1–7. [Google Scholar] [CrossRef]
- Nielsen, B.J.; Jensen, N.L.; Hartvig, P.; Hjelmroth, L.; Weber, R.W.S. Fungicide Resistance in Botrytis in Danish Strawberry Production. Erwerbs-Obstbau 2021, 63, 1–6. [Google Scholar] [CrossRef]
- Fournier, E.; Giraud, T. Sympatric genetic differentiation of a generalist pathogenic fungus, Botrytis cinerea, on two different host plants, grapevine and bramble. J. Evol. Biol. 2008, 21, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Rasiukevičiutė, N.; Moročko-Bičevska, I.; Sasnauskas, A. Characterisation of growth variability and mycelial compatibility of Botrytis cinerea isolates originated from apple and strawberry in Lithuania. Proc. Latv. Acad. Sci. Sect. B 2017, 71, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, D.M.Q.; Martins, S.D.S.; Guterres, D.C.; Martins, M.D.; Araújo, L.; Guimarães, L.M.S.; Alfenas, A.C.; Furtado, G.Q. Diversity, prevalence and phylogenetic positioning of Botrytis species in Brazil. Fungal Biol. 2020, 124, 940–957. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Tayal, P.; Sharma, E.; Kapoor, R. Analyses of genetic and pathogenic variability among Botrytis cinerea isolates. Microbiol. Res. 2014, 169, 862–872. [Google Scholar] [CrossRef]
- Rasiukevičiūtė, N.; Rugienius, R.; Šikšnianienė, J.B. Genetic diversity of Botrytis cinerea from strawberry in Lithuania. Zemdirbyste-Agriculture 2018, 105, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Šernaitė, L.; Rasiukevičiūtė, N.; Dambrauskienė, E.; Viškelis, P.; Valiuškaitė, A. Biocontrol of strawberry pathogen Botrytis cinerea using plant extracts and essential oils. Zemdirbyste-Agriculture 2020, 107, 147–152. [Google Scholar] [CrossRef]
- Šernaitė, L.; Rasiukevičiūtė, N.; Valiuškaitė, A. The extracts of cinnamon and clove as potential biofungicides against strawberry grey mould. Plants 2020, 9, 613. [Google Scholar] [CrossRef]
- Šernaitė, L.; Rasiukevičiūtė, N.; Valiuškaitė, A. Application of plants extracts to control postharvest gray mold and susceptibility of apple fruits to B. cinerea from different plant hosts. Foods 2020, 9, 1430. [Google Scholar] [CrossRef]
Figure 1.
Principal components (p < 0.05), showing differences between the sensitivity of separate Botrytis cinerea isolates from ten different plant hosts complex (BC1-BC10) to cinnamon extract (a), clove extract (b), and laurel extract (c).
Figure 1.
Principal components (p < 0.05), showing differences between the sensitivity of separate Botrytis cinerea isolates from ten different plant hosts complex (BC1-BC10) to cinnamon extract (a), clove extract (b), and laurel extract (c).


Figure 2.
Colony morphology of B. cinerea control isolates grown for 7 days on PDA, at 22 °C in the dark.
Figure 2.
Colony morphology of B. cinerea control isolates grown for 7 days on PDA, at 22 °C in the dark.

Figure 3.
Hyphae morphology of B. cinerea isolate from apple, grown on PDA with 3600 µL/L (a) and 4200 µL/L (b) observed under 40× magnification.
Figure 3.
Hyphae morphology of B. cinerea isolate from apple, grown on PDA with 3600 µL/L (a) and 4200 µL/L (b) observed under 40× magnification.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison of the sensitivity of Botrytis cinerea isolate complex (BC1-BC10) to different concentrations of the extracts (µL/L) 7 days after inoculation. Data presented as radial colony growth of pathogens (cm).
Table 1.
Comparison of the sensitivity of Botrytis cinerea isolate complex (BC1-BC10) to different concentrations of the extracts (µL/L) 7 days after inoculation. Data presented as radial colony growth of pathogens (cm).
| Extract | µL/L | BC1 | BC2 | BC3 | BC4 | BC5 | BC6 | BC7 | BC8 | BC9 | BC10 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cinnamon | 100 | 7.80a | 7.80a | 7.80a | 7.80a | 7.80a | 7.80a | 7.80a | 7.80a | 7.80a | 7.80a |
| 200 | 7.80a | 7.80a | 7.71a | 7.80a | 7.80a | 7.80a | 7.80a | 7.80a | 7.80a | 7.54ba | |
| 300 | 0.58g | 7.80a | 7.80a | 7.80a | 7.80a | dgfe | 7.80a | 7.80a | 7.80a | 7.45ba | |
| 400 | 0.75g | 7.80a | 7.73a | 6.61ba | 7.76a | 6.63ba | 7.80a | 7.80a | 7.62a | 6.55b | |
| 500 | 0.00g | 4.80dc | 5.19c | 0.86g | 0.80i | 0.00i | 2.26f | 0.70i | 1.06fe | 1.76jhi | |
| 600 | 0.00g | 0.00g | 0.00e | 0.00g | 0.00i | 0.00i | 0.00g | 0.00i | 0.00f | 0.00k | |
| 800 | 0.00g | 0.00g | 0.00e | 0.00g | 0.00i | 0.00i | 0.00g | 0.00i | 0.00f | 0.00k | |
| Clove | 100 | 7.80a | 7.80a | 7.80a | 7.08ba | 7.80a | 7.80a | 7.80a | 7.80a | 7.80a | 6.69b |
| 200 | 7.14a | 6.30b | 6.96ba | 6.91ba | 6.01b | 7.08ba | 7.13a | 6.91b | 5.48cb | 4.41dc | |
| 300 | 5.29b | 4.56dce | 5.35c | 6.90ba | 4.79ed | 3.50dfe | 5.61cb | 6.19cb | 4.29cd | 3.56dfe | |
| 400 | 5.25b | 4.08dce | 3.05d | 4.38dce | 3.68fg | 5.25bc | 3.70e | 4.35fg | 3.63d | 3.87dce | |
| 500 | 0.00g | 2.58f | 0.00e | 2.83f | 3.28g | 0.00i | 4.55ced | 3.71g | 2.14e | 2.45ghf | |
| 600 | 0.00g | 0.02g | 1.22e | 1.29g | 2.24h | 1.61hgi | 1.44f | 2.01h | 1.77e | 1.38ji | |
| 800 | 0.00g | 0.00g | 0.00e | 0.00g | 0.00i | 0.99hi | 0.00g | 0.00i | 0.13f | 0.91jk | |
| Laurel | 2600 | 4.68cbd | 4.56dce | 6.93ba | 6.63ba | 5.30cbd | 6.65ba | 4.75cbd | 5.93cd | 5.61b | 4.78c |
| 2800 | 2.91efd | 4.38dce | 6.42bac | 4.95dc | 5.80cb | 6.18ba | 5.08cbd | 5.77cde | 5.27cb | 4.43dc | |
| 3000 | 3.08cefd | 3.90dfce | 5.30c | 4.48dce | 4.74ed | 5.54bc | 4.61ced | 4.98fe | 4.30cd | 3.84dce | |
| 3200 | 4.89cb | 3.29dfe | 6.58bac | 3.49fe | 5.10cd | 2.81dgfe | 4.66ced | 4.93fe | 4.79cbd | 2.95gfe | |
| 3400 | 1.93gef | 4.80dc | 5.25c | 4.86dce | 5.51cbd | 5.59bc | 5.29cb | 5.08fde | 5.65b | 2.99gfe | |
| 3600 | 1.44gf | 3.08fe | 5.98bc | 4.75dce | 4.24fe | 2.11hgfe | 5.77b | 4.45fg | 4.81cbd | 2.24ghi | |
| 3800 | 1.46gf | 3.25fe | 5.19c | 4.80dce | 4.06feg | 3.86dce | 5.01cbd | 5.07fde | 4.85cbd | 3.11gfe | |
| 4000 | 0.79g | 4.08dce | 5.10c | 3.99dfe | 3.83fg | 1.84hgf | 4.14ed | 4.59f | 4.51cbd | 2.73ghfe | |
| 4200 | 3.41cebd | 4.84c | 3.51d | 5.74bc | 5.45cbd | 4.35dc | 5.58cb | 5.76cde | 4.89cbd | 3.86dce |
Different letters show statistical differences of each isolate sensitivity to extract treatment according to the Duncan Multiple range test at the 5% probability level (p < 0.05).
Table 2.
Mycelium and colour changes in B. cinerea isolate complex after treatment with cinnamon, clove, and laurel extracts. Colony morphology illustrations represent a surface side of B. cinerea grown for 7 days on PDA with investigated extract, at 22 °C in the dark.
Table 2.
Mycelium and colour changes in B. cinerea isolate complex after treatment with cinnamon, clove, and laurel extracts. Colony morphology illustrations represent a surface side of B. cinerea grown for 7 days on PDA with investigated extract, at 22 °C in the dark.
| Isolate | Cinnamon Extract | Clove Extract | Laurel Extract | ||||||
|---|---|---|---|---|---|---|---|---|---|
| M | C | CM | M | C | CM | M | C | CM | |
| BC1 | − | − |  | + | + |  | + | − |  |
| BC2 | − | − |  | + | − |  | − | − |  |
| BC3 | + | − |  | + | − |  | + | − |  |
| BC4 | − | − |  | + | − |  | + | − |  |
| BC5 | − | + |  | + | + |  | + | + |  |
| BC6 | − | − |  | − | − |  | − | − |  |
| BC7 | + | + |  | + | + |  | + | + |  |
| BC8 | + | − |  | − | − |  | + | + |  |
| BC9 | − | − |  | + | − |  | + | + |  |
| BC10 | − | + |  | − | − |  | + | − |  |
M—mycelium; C—colour; CM—colony morphology. “+” Visible changes of the morphological attribute. “−“ No visible changes of the morphological attribute.
Table 3.
Conidia size (µm) of B. cinerea isolates after treatment with cinnamon, clove, and laurel extracts.
Table 3.
Conidia size (µm) of B. cinerea isolates after treatment with cinnamon, clove, and laurel extracts.
| Extract Isolate | Control | Cinnamon | Clove | Laurel | |||||
|---|---|---|---|---|---|---|---|---|---|
| 0 µL/L | 400 µL/L | 600 µL/L | 400 µL/L | 600 µL/L | 2600 µL/L | 2800 µL/L | 3000 µL/L | ||
| BC2 | Length | 19.38a | 18.93a | – | 13.56b | – | 14.82b | n.d. | 19.60a |
| Width | 14.54ba | 16.05a | 11.87b | 12.56b | 15.72a | ||||
| BC3 | Length | n.d. | 19.91a | n.d. | 19.07a | n.d. | |||
| Width | 16.05a | 12.34b | |||||||
| BC4 | Length | 19.92dc | 22.13a | 19.00ed | 20.30bdc | 21.02bac | 17.66e | 21.76ba | |
| Width | 15.43b | 13.03c | 15.66b | 16.39ba | 15.45b | 16.15ba | 17.31a | ||
| BC5 | Length | 19.54ba | 21.39a | 19.75ba | 18.24ba | 20.31ba | 16.94b | 21.07a | |
| Width | 15.24ba | 15.87a | 15.22ba | 15.51ba | 16.62a | 13.15b | 15.56ba | ||
| BC7 | Length | 20.91b | 19.87b | n.d. | – | 19.26b | 24.70a | n.d. | |
| Width | 14.60b | 13.81b | 16.46b | 21.08a | |||||
| BC8 | Length | 19.13a | 20.35a | 20.53a | 20.58a | 20.92a | 19.09a | 16.75b | |
| Width | 15.82ba | 15.55ba | 16.06a | 15.30ba | 16.62a | 16.15a | 14.08b | ||
| BC9 | Length | 18.43bac | 17.85bc | 20.35a | 17.31c | 17.83bc | 19.92ba | 17.52bc | 18.09bac |
| Width | 14.34bc | 14.59bc | 15.09bac | 13.82c | 14.21bc | 16.21a | 13.77c | 15.42ba | |
| BC10 | Length | 17.36a | 17.45a | – | n.d. | – | 17.22a | 17.74a | n.d. |
| Width | 13.55a | 14.87a | 13.78a | 14.70a | |||||
“n.d.” Means not detected. “–“ Sign means that no colony growth of the pathogen was observed. Different letters show statistical differences of each isolate conidia length/width after treatment with extracts according to the Duncan Multiple range test at the 5% probability level (p < 0.05).
Table 4.
Origin, cryptic species groups, and genotypes of the Botrytis cinerea isolates complex used in the study.
Table 4.
Origin, cryptic species groups, and genotypes of the Botrytis cinerea isolates complex used in the study.
| Isolate Code | Plant Host | Latin Name | Group | Transposable Element Type | ||||
|---|---|---|---|---|---|---|---|---|
| I | II | F | B | V | T | |||
| BC1 | Strawberry | Fragaria magna f. ananassa | + | + | ||||
| BC2 | Onion | Allium cepa | + | + | ||||
| BC3 | Tomato | Solanum lycopersicum | + | + | ||||
| BC4 | Raspberry | Rubus idaeus | + | + | ||||
| BC5 | Cabbage/leaf | Brassica oleracea | + | + | ||||
| BC6 | Apple | Malus domestica | + | + | ||||
| BC7 | Grapevine | Vitis spp. | + | + | ||||
| BC8 | Bell pepper | Capsicum annuum | + | + | ||||
| BC9 | Leek | Allium porrum | + | + | ||||
| BC10 | Rose | Rosa spp. | + | + | ||||
| Total | 1 | 9 | 1 | 1 | 4 | 4 | ||
F—Flipper, B—Boty, V—Vacuma, T—Transposa, I—group—resistant to fenhexamid, II—sensitive. “+” indicates cryptic species group/transposable element type of the isolate.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Dėnė, L.; Valiuškaitė, A. Sensitivity of Botrytis cinerea Isolates Complex to Plant Extracts. Molecules 2021, 26, 4595. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26154595
AMA Style
Dėnė L, Valiuškaitė A. Sensitivity of Botrytis cinerea Isolates Complex to Plant Extracts. Molecules. 2021; 26(15):4595. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26154595
Chicago/Turabian StyleDėnė, Lina, and Alma Valiuškaitė. 2021. "Sensitivity of Botrytis cinerea Isolates Complex to Plant Extracts" Molecules 26, no. 15: 4595. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26154595