
Parthenin—A Sesquiterpene Lactone with Multifaceted Biological Activities: Insights and Prospects
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
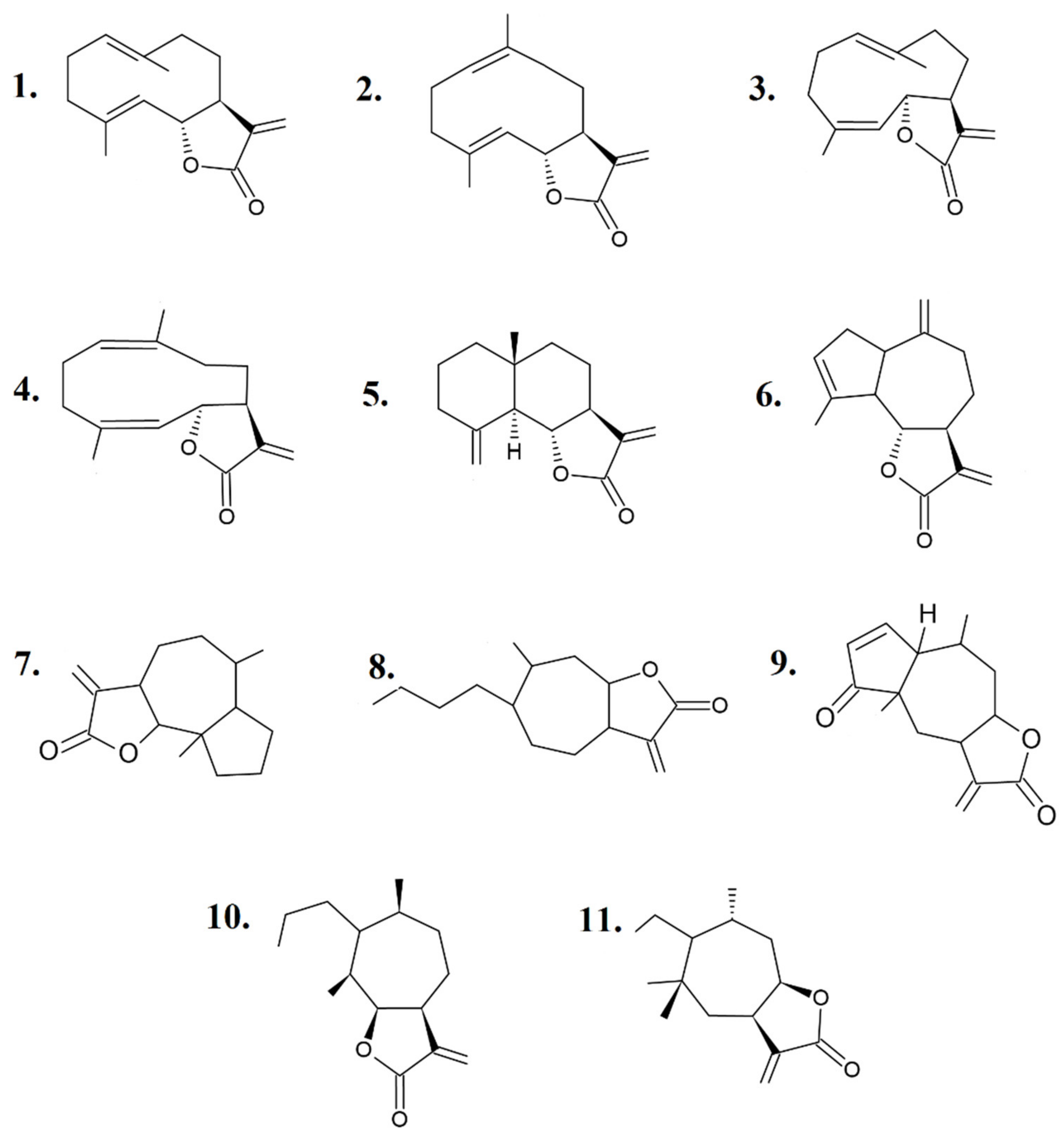
:1. Sesquiterpene Lactones: An Introduction
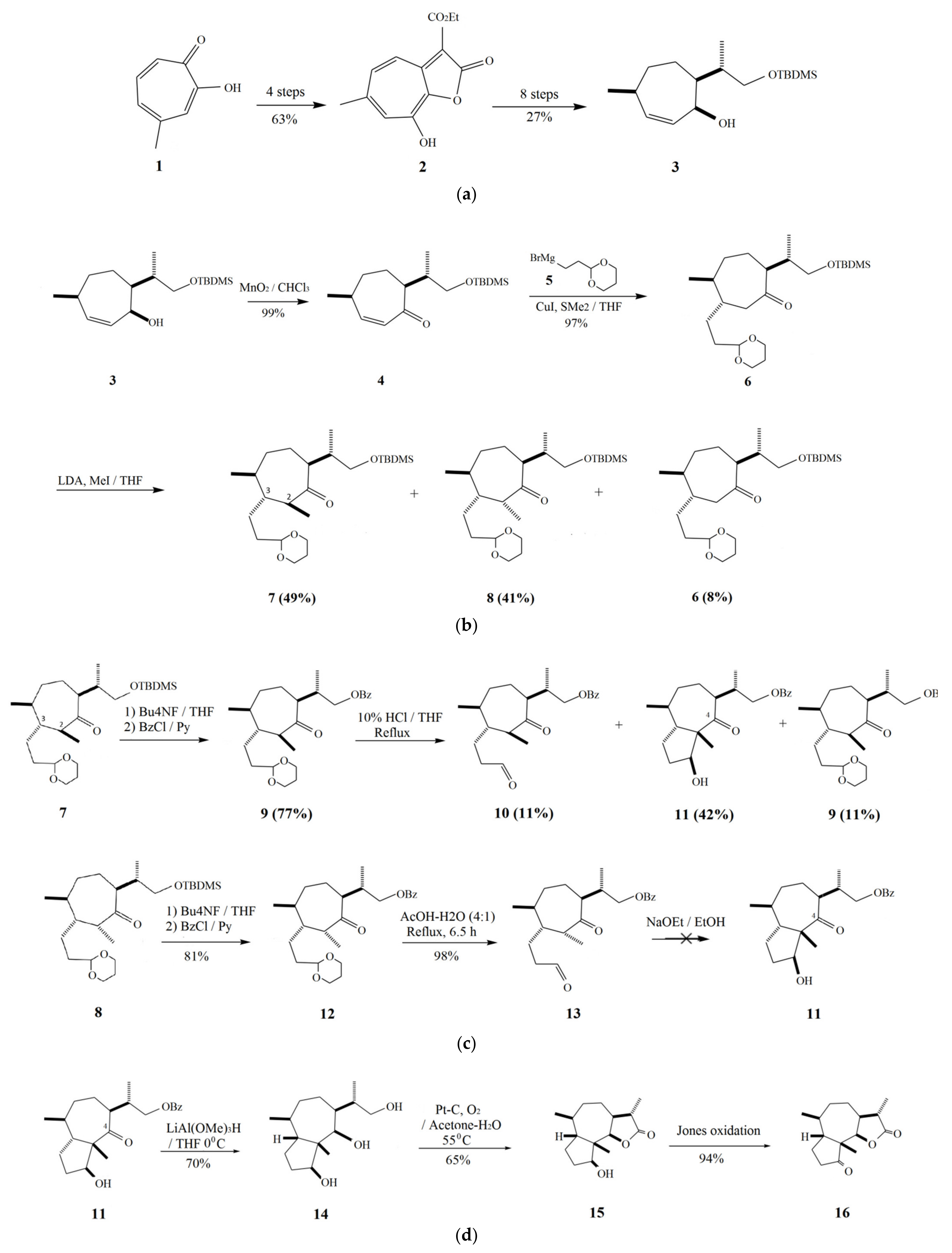
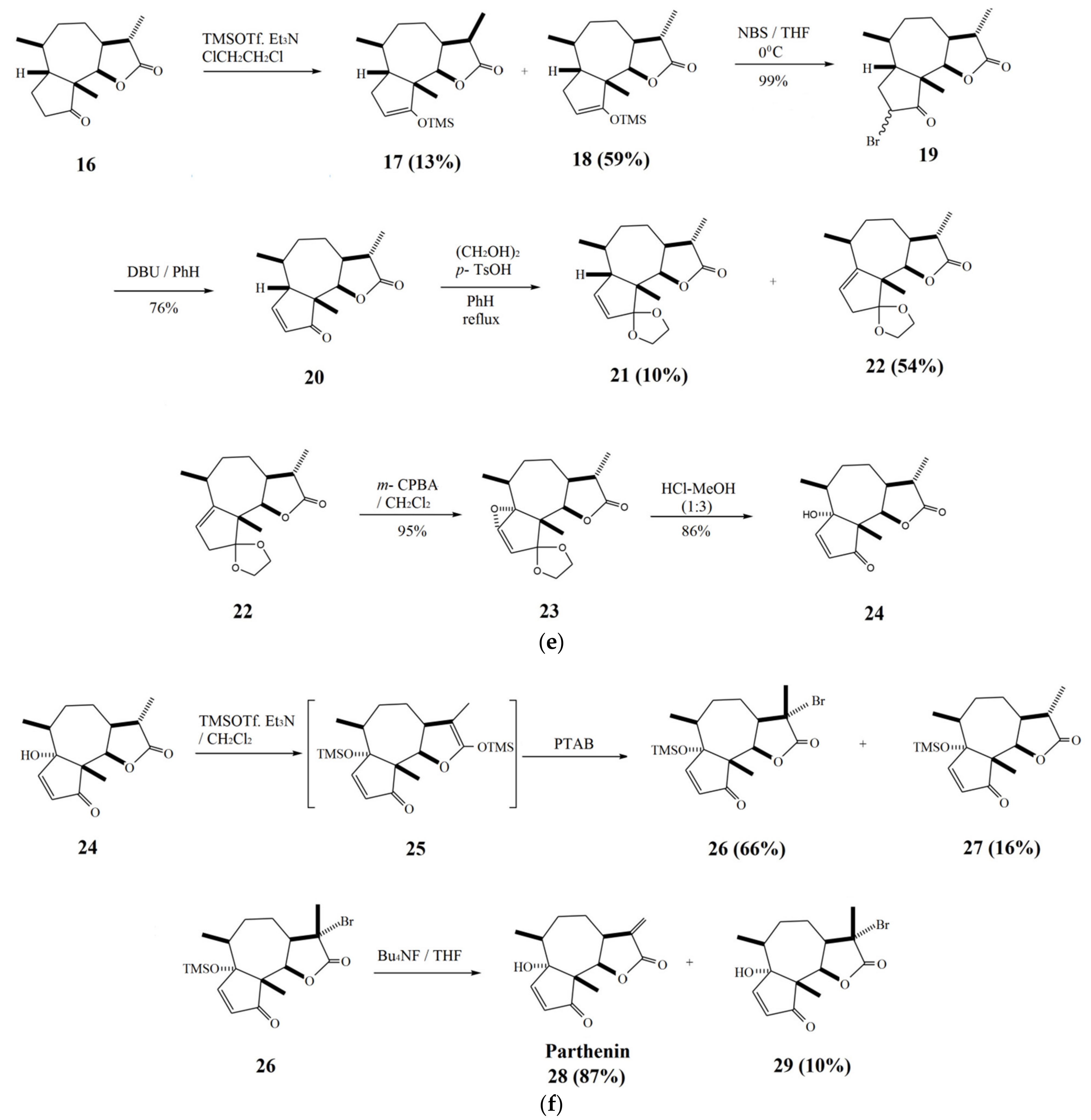
2. Parthenin
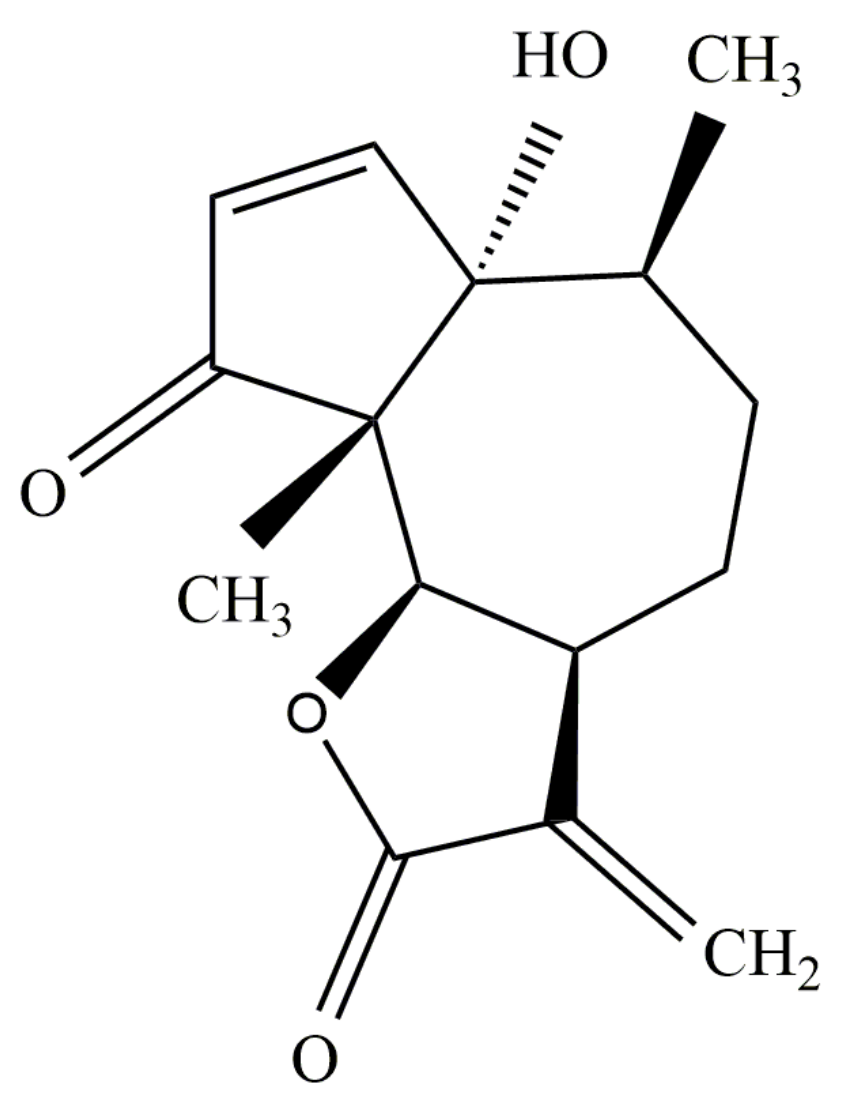
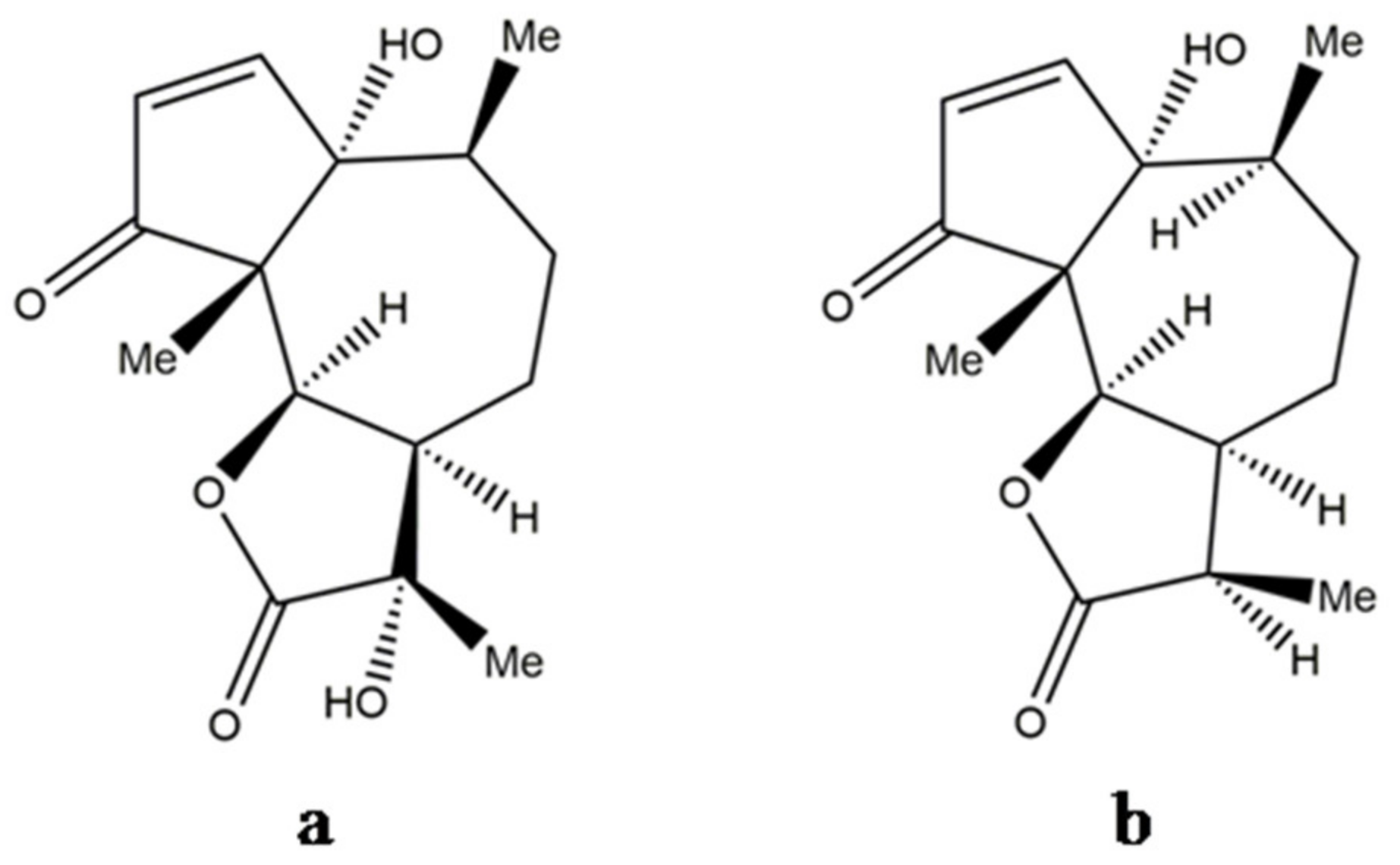
3. Structure of Parthenin
4. Pharmacological Properties of Parthenin
4.1. Anti-Cancerous Activity
4.2. Anti-Malarial Activity
4.3. Others
5. Phytotoxic Property of Parthenin
6. Pesticidal Properties of Parthenin
6.1. Insecticidal Properties
6.2. Fungicidal Properties
7. Toxicological Concerns
8. Prospects and Way Forward
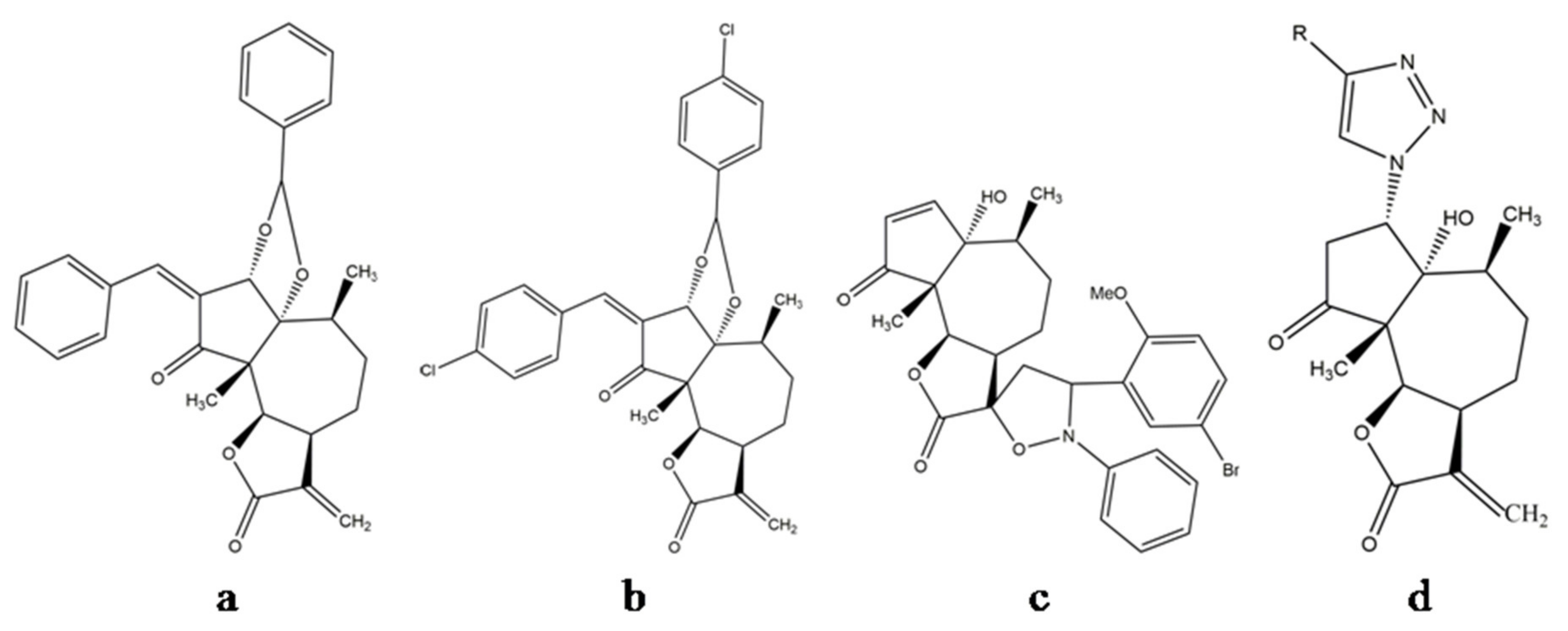
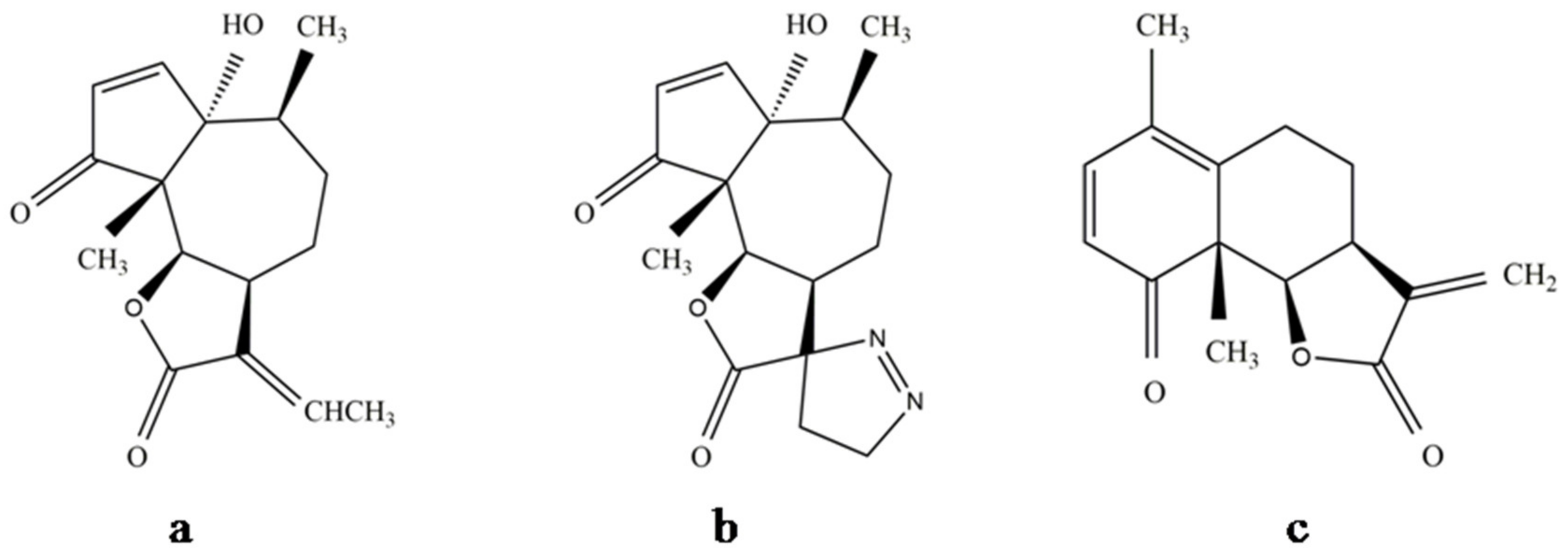
8.1. Development of Structural Analogues
8.2. Selection of Suitable Doses
8.3. Characterisation of Parthenin
8.4. Others
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Sülsen, V.P.; Martino, V.S. Sesquiterpene Lactones- Advances in their Chemistry and Biological Aspects; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Padilla-Gonzalez, G.F.; dos Santos, F.A.; Da Costa, F.B. Sesquiterpene lactones: More than protective plant compounds with high toxicity. Crit. Rev. Plant Sci. 2016, 35, 18–37. [Google Scholar] [CrossRef]
- Chadwick, M.; Trewin, H.; Gawthrop, F.; Wagstaff, C. Sesquiterpenoids lactones: Benefits to plants and people. Int. J. Mol. Sci. 2013, 14, 12780–12895. [Google Scholar] [CrossRef] [Green Version]
- Quintana, J.; Estévez, F. Recent advances on cytotoxic sesquiterpene lactones. Curr. Pharm. Des. 2018, 24, 4355–4361. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.P.; Batish, D.R.; Kohli, R.K.; Saxena, D.B.; Arora, V. Effect of parthenin-a sesquiterpene lactone from Parthenium hysterophorus on early growth and physiology of Ageratum conyzoides. J. Chem. Ecol. 2002, 28, 2169–2179. [Google Scholar] [CrossRef] [PubMed]
- Reddy, D.M.; Qazi, N.A.; Sawant, S.D.; Bandey, A.H.; Srinivas, J.; Shankar, M.; Singh, S.K.; Verma, M.; Chashoo, G.; Saxena, A.; et al. Design and synthesis of spiro derivatives of parthenin as novel anti-cancer agents. Eur. J. Med. Chem. 2011, 46, 3210–3217. [Google Scholar] [CrossRef]
- De Luque, A.P.; Galindo, J.C.G.; Macías, F.A.; Jorrín, J. Sunflower sesquiterpene lactone models induce Orobanche cumana seed germination. Phytochemistry 2000, 53, 45–50. [Google Scholar] [CrossRef]
- Macías, F.A.; Torres, A.; Molinllo, J.M.G.; Varela, R.M.; Castellano, D. Potential allelopathic sesquiterpene lactones from sunflower leaves. Phytochemistry 1996, 43, 1205–1215. [Google Scholar] [CrossRef]
- Rodriguez, E.; Towers, G.H.N.; Mitchell, J.C. Biological activities of sesquiterpene lactones. Phytochemistry 1976, 15, 1573–1580. [Google Scholar] [CrossRef]
- Ivanescu, B.; Miron, A.; Corciova, A. Sesquiterpene lactones from Artemisia genus: Biological activities and methods of analysis. J. Anal. Method. Chem. 2015, 2015, 247685. [Google Scholar] [CrossRef] [Green Version]
- Barbero, M.; Prandi, C. Pseudoguaianolides: Recent advances in synthesis and applications. Nat. Prod. Comm. 2018, 13, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Datta, S.; Saxena, D.B. Pesticidal properties of parthenin (from Parthenium hysterophorus) and related compounds. Pest Manag. Sci. 2001, 57, 95–101. [Google Scholar] [CrossRef]
- Goswami, A.; Ali Shah, B.; Batra, N.; Kumar, A.; Kumar, G.S.; Bhushan, S.; Ahmad, M.F.; Joshi, A.; Singh, J. Multiple pharmacological properties of a novel parthenin analog P16 as evident by its cytostatic and antiangiogenic potential against pancreatic adenocarcinoma PANC-1 cells. Anticancer Agents Med. Chem. 2011, 16, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Milugo, T.K.; Tchouassi, D.P.; Kavishe, R.A.; Dinglasan, R.R.; Torto, B. Derivatization increases mosquito larvicidal activity of the sesquiterpene lactone parthenin isolated from the invasive weed Parthenium hysterophorus. Pest Manag. Sci. 2021, 77, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Shabbir, A.; Adkins, S. Parthenium hysterophorus: A tale of global invasion over two centuries, spread and prevention measures. J. Environ. Manage. 2021, 279, 111751. [Google Scholar] [CrossRef] [PubMed]
- Rice, C.; Wolf, J.; Fleisher, D.H.; Acosta, S.M.; Adkins, S.W.; Bajwa, A.A.; Ziska, L.H. Recent CO2 levels promote increased production of the toxin parthenin in an invasive Parthenium hysterophorus biotype. Nat. Plants 2021, 7, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, C.; Kraus, S.; Walker, F.; Foxcroft, L.C.; Robbertse, P.J.; Hurle, K. The allelochemical parthenin is sequestered at high level in capitate-sessile trichomes on leaf surfaces of Parthenium hysterophorus. J. Plant Dis. Prot. 2004, 19, 253–261. [Google Scholar]
- Kanchan, S.D.; Jayachandra. Allelopathic effects of Parthenium hysterophorus L.: Part IV. Identification of inhibitors. Plant Soil 1980, 55, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Akhtar, N.; Verma, K.K.; Sharma, A. Immunogenetics of cytokine genes in parthenium dermatitis: A review. Eur. Ann. Allergy Clin. Immunol. 2018, 50, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhardt, C.; Van der Laan, M.; Belz, R.G.; Hurle, K.; Foxcroft, L.C. Production dynamics of the allelochemical parthenin in leaves of Parthenium hysterophorus L. J. Plant Dis. Prot. 2006, 20, 427–433. [Google Scholar]
- Belz, R.G. Stimulation versus inhibition—Bioactivity of parthenin, a phytochemical from Parthenium hysterophorus L. Dose Response 2008, 6, 80–96. [Google Scholar] [CrossRef] [Green Version]
- Patel, S. Harmful and beneficial aspects of Parthenium hysterophorus: An update. 3 Biotech 2011, 1, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kok, P.; De Clercq, P.; Vandewalle, M. Pseudoguaianolides. The total synthesis of (±)-neoambrosin (±)-parthenin and (±)-hymenin. Bull. Soc. Chim. Belg. 1978, 87, 615–619. [Google Scholar] [CrossRef]
- Shimoma, F.; Kusaka, H.; Azami, H.; Wada, K.; Suzuki, T.; Hagiwara, H.; Ando, M. Total syntheses of (±)-hymenolin and (±)-parthenin. J. Org. Chem. 1998, 63, 3758–3763. [Google Scholar] [CrossRef]
- Herz, W.; Watanabe, H.; Miyazaki, M.; Kishida, Y. The structures of parthenin and ambrosin. J. Am. Chem. Soc. 1962, 84, 2601–2610. [Google Scholar] [CrossRef]
- Ramos, A.; Rivero, R.; Visozo, A.; Piloto, J.; García, A. Parthenin, a sesquiterpene lactone of Parthenium hysterophorus L. is a high toxicity clastogen. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2002, 514, 19–27. [Google Scholar] [CrossRef]
- Raghu, J.D.; Veerashekar, T.; Kuppast, I.J.; Dharshan, S.; Ravi, M.C. A review on Parthenium hysterophorus Linn. Int. J. Univ. Pharm. Bio Sci. 2014, 3, 110–120. [Google Scholar]
- Lonkar, A.; Nagasampagi, B.A.; Narayanan, C.R.; Landge, A.B.; Sawalkar, D.D. An antigen from Parthenium hysterophorus Linn. Contact Dermatitis 1976, 2, 151–154. [Google Scholar] [CrossRef]
- Heathcock, C.H.; Tice, C.M.; Germroth, T.C. Synthesis of sesquiterpene antitumor lactones. 10. Total synthesis of (±)-parthenin. J. Am. Chem. Soc. 1982, 104, 6081–6091. [Google Scholar] [CrossRef]
- Da Silva, L.P.; Borges, B.A.; Veloso, M.P.; Chagas-Paula, D.A.; Gonçalves, R.V.; Novaes, R.D. Impact of sesquiterpene lactones on the skin and skin-related cells? A systematic review of in vitro and in vivo evidence. Life Sci. 2021, 265, 118815. [Google Scholar] [CrossRef] [PubMed]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Saxena, D.B. Allelopathic effects of parthenin-a sesquiterpene lactone, on germination, and early growth of mung bean (Phaseolus aureus Roxb.). PGRSA Q. 2001, 29, 81–91. [Google Scholar]
- Belz, R.G.; van der Laan, M.; Reinhardt, C.F.; Hurle, K. Soil degradation of parthenin—does it contradict the role of allelopathy in the invasive weed Parthenium hysterophorus L.? J. Chem. Ecol. 2009, 35, 1137–1150. [Google Scholar] [CrossRef] [PubMed]
- Fronczek, F.R.; Vargas, D.; Fischer, N.H.; Chiari, G.; Balza, F.; Towers, G.H.N. Structures of three pseudoguaianolides: Parthenin, hymenolin (11β, 13-dihydroparthenin) and bipinnatin. Acta Cryst. C. 1989, 45, 2006–2010. [Google Scholar] [CrossRef]
- Shah, B.A.; Kaur, R.; Gupta, P.; Kumar, A.; Sethi, V.K.; Andotra, S.S.; Singh, J.; Saxena, A.K.; Taneja, S.C. Structure–activity relationship (SAR) of parthenin analogues with pro-apoptotic activity: Development of novel anti-cancer leads. Bioorganic Med. Chem. Lett. 2009, 19, 4394–4398. [Google Scholar] [CrossRef]
- Picman, A.K.; Rodriguez, E.; Towers, G.H.N. Formation of adducts of parthenin and related sesquiterpene lactones with cysteine and glutathione. Chem. Biol. Interact. 1979, 28, 83–89. [Google Scholar] [CrossRef]
- Isman, M.B. Toxicity and tolerance of sesquiterpene lactones in the migratory grasshopper, Melanoplus sanguinipes (Acrididae). Pestic. Biochem. Physiol. 1985, 24, 348–354. [Google Scholar] [CrossRef]
- Kupchan, S.M.; Fessler, D.C.; Eakin, M.A.; Giacobbe, T.J. Reactions of alpha methylene lactone tumor inhibitors with model biological nucleophiles. Science 1970, 168, 376–378. [Google Scholar] [CrossRef] [PubMed]
- Saxena, D.B.; Dureja, P.; Kumar, B.; Rani, D.; Kohli, R.K. Modification of parthenin. Indian, J. Chem. B. 1991, 30, 849–852. [Google Scholar]
- Belz, R.G.; Reinhardt, C.F.; Foxcroft, L.C.; Hurle, K. Residue allelopathy in Parthenium hysterophorus L.—Does parthenin play a leading role? Crop Prot. 2007, 26, 237–245. [Google Scholar] [CrossRef] [Green Version]
- Hernández, Y.S.; Sánchez, L.B.; Bedia, M.M.G.; Gómez, L.T.; Rodríguez, E.J.; San Miguel, H.M.G.; Mosquera, D.G.; García, L.P.; Dhooghe, L.; Theunis, M.; et al. Determination of parthenin in Parthenium hysterophorus L. by means of HPLC-UV: Method development and validation. Phytochem. Lett. 2011, 4, 134–147. [Google Scholar] [CrossRef]
- Asaoka, M.; Ohkubo, T.; Itahana, H.; Kosaka, T.; Takei, H. Enantioselective synthesis of neoambrosin, parthenin, and dihydroisoparthenin. Tetrahedron 1995, 51, 3115–3128. [Google Scholar] [CrossRef]
- Mukherjee, B.; Chatterjee, M. Antitumor activity of Parthenium hysterophorus and its effect in the modulation of biotransforming enzymes in transplanted murine leukemia. Planta Med. 1993, 59, 513–516. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Malik, F.; Bhushan, S.; Shah, B.A.; Taneja, S.C.; Pal, H.C.; Wani, Z.A.; Mondhe, D.M.; Kaur, J.; Singh, J. A novel parthenin analog exhibits anti-cancer activity: Activation of apoptotic signaling events through robust NO formation in human leukemia HL-60 cells. Chem. Biol. Interact. 2011, 193, 204–215. [Google Scholar] [CrossRef]
- Espinosa-Rivero, J.; Rendón-Huerta, E.; Romero, I. Inhibition of Helicobacter pylori growth and its colonization factors by Parthenium hysterophorus extracts. J. Ethnopharmacol. 2015, 174, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Aggarwal, N.K.; Dhiman, R. Antimicrobial activity of medicinal plant: Parthenium hysterophorus L. Res. J. Med. Plant. 2016, 10, 106–112. [Google Scholar] [CrossRef] [Green Version]
- Muktar, Y.; Mohammadnur, M.; Wondimu, A.; Hiko, A. In vitro antibacterial activity of crude methanol extracts of various parts of Parthenium hysterophorus against pathogenic bacterial strains. Ethiop. Vet. J. 2017, 21, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Maharjan, S.; Shrestha, B.B.; Devkota, A.; Muniappan, R.; Jha, P.K. Temporal and spatial patterns of research on a globally significant invasive weed Parthenium hysterophorus L.: A bibliographic review. Crop Prot. 2020, 135, 104832. [Google Scholar] [CrossRef]
- Mew, D.; Balza, F.; Towers, G.H.N.; Levy, J.G. Anti-tumour effects of the sesquiterpene lactone parthenin. Planta Med. 1982, 45, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Halmuthur, M.S.K.; Saxena, A.K.; Taneja, S.C.; Singh, S.K.; Sethi, V.K.; Qazi, N.A.; Sawant, S.D.; Doma, M.R.; Banday, A.H.; Verma, M.; et al. Spiro Derivatives of Parthenin as Novel Anticancer Agents. U.S. Patent 860985, 23 July 1907. Available online: https://patents.google.com/patent/US860985 (accessed on 3 July 2021).
- Goswami, A.; Shah, B.A.; Kumar, A.; Rizvi, M.A.; Kumar, S.; Bhushan, S.; Malik, F.A.; Batra, N.; Joshi, A.; Singh, J. Antiproliferative potential of a novel parthenin analog P16 as evident by apoptosis accompanied by down-regulation of PI3K/AKT and ERK pathways in human acute lymphoblastic leukemia MOLT-4 cells. Chem. Biol. Interact. 2014, 222, 60–67. [Google Scholar] [CrossRef]
- Saxena, A.; Bhusan, S.; Sachin, B.S.; Kessar, R.R.; Reddy, D.M.; Kumar, H.M.S.; Saxena, A.K. Antineoplastic properties of parthenin derivatives–the other faces of a weed. In Chemistry of Phytopotentials: Health, Energy and Environmental Perspectives; Khemani, L., Srivastava, M., Srivastava, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 13–17. [Google Scholar]
- Khazir, J.; Hyder, I.; Gayatri, J.L.; Yandrati, L.P.; Nalla, N.; Chasoo, G.; Mahajan, A.; Saxena, A.K.; Alam, M.S.; Qazi, G.N.; et al. Design and synthesis of novel 1,2,3-triazole derivatives of coronopilin as anti-cancer compounds. Eur. J. Med. Chem. 2014, 82, 255–262. [Google Scholar] [CrossRef]
- Hooper, M.; Kirby, G.C.; Kulkarni, M.M.; Kulkarni, S.N.; Nagasampagi, B.A.; O’Neill, M.J.; Phillipson, J.D.; Rojatkar, S.R.; Warhurst, D.C. Antimalarial activity of parthenin and its derivatives. Eur. J. Med. Chem. 1990, 25, 717–723. [Google Scholar] [CrossRef]
- Balaich, J.N.; Mathias, D.K.; Torto, B.; Jackson, B.T.; Tao, D.; Ebrahimi, B.; Tarimo, B.B.; Cheseto, X.; Foster, W.A.; Dinglasan, R.R. The non-artemisinin sesquiterpene lactones parthenin and parthenolide block Plasmodium falciparum sexual stage transmission. Antimicrob. Agents Chemother. 2016, 60, 2108–2117. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Kushwaha, P.P.; Kumar, S. Parthenin and its similar structure as potential lead inhibitors of Plasmodium vivax and Plasmodium falciparum Lactate Dehydrogenase. In Phytochemistry: An In-Silico and In-Vitro Update; Kumar, S., Egbuna, C., Eds.; Springer: Singapore, 2019; pp. 565–567. [Google Scholar]
- Sharma, G.L.; Bhutani, K.K. Plant Based Antiamoebic Drugs; Part II. Amoebicidal activity of parthenin isolated from Parthenium hysterophorus. Planta Med. 1988, 54, 120–122. [Google Scholar] [CrossRef]
- Chib, R.; Shah, B.A.; Anand, N.; Pandey, A.; Kapoor, K.; Bani, S.; Gupta, V.K.; Rajnikant; Sethi, V.K.; Taneja, S.C. Psilostachyin, acetylated pseudoguaianolides and their analogues: Preparation and evaluation of their anti-inflammatory potential. Bioorganic Med. Chem. Lett. 2011, 21, 4847–4851. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, C.; Harakishore, K.; Murty, U.S.N.; Das, B. Analogues of parthenin and their antibacterial activity. Arkivoc 2003, 9, 126–132. [Google Scholar]
- Mersie, W.; Singh, M. Allelopathic effect of parthenium (Parthenium hysterophorus L.) extract and residue on some agronomic crops and weeds. J. Chem. Ecol. 1987, 13, 1739–1747. [Google Scholar] [CrossRef]
- Adkins, S.W.; Sowerby, M.S. Allelopathic potential of the weed, Parthenium hysterophorus L., in Australia. Plant Prot. Q. 1996, 11, 20–23. [Google Scholar]
- Singh, H.P.; Batish, D.R.; Pandher, J.K.; Kohli, R.K. Assessment of allelopathic properties of Parthenium hysterophorus residues. Agric. Ecosyst. Environ. 2003, 95, 537–541. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Pandher, J.K.; Kohli, R.K. Phytotoxic effects of Parthenium hysterophorus residues on three Brassica species. Weed Biol. Manag. 2005, 5, 105–109. [Google Scholar] [CrossRef]
- Batish, D.R.; Kohli, R.K.; Singh, H.P.; Saxena, D.B. Studies on herbicidal activity of parthenin, a constituent of Parthenium hysterophorus, towards billgoat weed (Ageratum conyzoides). Curr. Sci. 1997, 73, 369–371. [Google Scholar]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Saxena, D.B.; Kaur, S. Allelopathic effects of parthenin against two weedy species, Avena fatua and Bidens pilosa. Environ. Exp. Bot. 2002, 47, 149–155. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Saxena, D.B.; Kohli, R.K. Weed suppressing ability of parthenin-a sesquiterpene lactone from Parthenium hysterophorus. N.Z. Plant Prot. 2002, 55, 218–221. [Google Scholar] [CrossRef]
- Khosla, S.N.; Kuldeep, S.; Sobti, S.N. Parthenin from Parthenium hysterophorus is phytotoxic too. Indian J. For. 1980, 3, 261–265. [Google Scholar]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Kaur, S.; Saxena, D.B.; Yadav, S. Assessment of phytotoxicity of parthenin. Z. Naturforsch. C. Bio. Sci. 2007, 62, 367–372. [Google Scholar] [CrossRef]
- Pandey, D.K. Phytotoxicity of sesquiterpene lactone parthenin on aquatic weeds. J. Chem. Ecol. 1996, 22, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Kohli, R.K.; Rani, D.; Verma, R.C. A mathematical model to predict the tissue response to parthenin—An allelochemical. Biol. Plant. 1993, 35, 567. [Google Scholar] [CrossRef]
- Batish, D.R.; Kohli, R.K.; Saxena, D.B.; Singh, H.P. Growth regulatory response of parthenin and its derivatives. Plant Growth Regul. 1997, 21, 189–194. [Google Scholar] [CrossRef]
- Belz, R.G. Investigating a potential auxin-related mode of hormetic/inhibitory action of the phytotoxin parthenin. J. Chem. Ecol. 2016, 42, 71–83. [Google Scholar] [CrossRef]
- Belz, R.G.; Cedergreen, N. Parthenin hormesis in plants depends on growth conditions. Environ. Exp. Bot. 2010, 69, 293–301. [Google Scholar] [CrossRef]
- Khosla, S.N.; Sobti, S.N. Parthenin—A promising root inhibitor from Parthenium hysterophorus Linn. Pesticides 1981, 15, 8–11. [Google Scholar]
- Sharma, R.N.; Joshi, V.N. Allomonic principles in Parthenium hysterophorus: Potential as insect control agents and role in the weed’s resistance to serious insect depredation. il. the biological activity of parthenin on insects. Biovigyanam 1977, 3, 225–231. [Google Scholar]
- Datta, S.; Saxena, D.B. Parthenin and azadirachtin-A as antifeedants against Spodoptera litura (Fab). Pestic. Res. J. 1997, 9, 263–266. [Google Scholar]
- Picman, A.K.; Elliott, R.H.; Towers, G.H.N. Cardiac-inhibiting properties of the sesquiterpene lactone, parthenin, in the migratory grasshopper, Melanoplus sanguinipes. Can. J. Zool. 1981, 59, 285–292. [Google Scholar] [CrossRef]
- Arnason, J.T.; Philogène, B.J.R.; Duval, F.; McLachlan, D.; Picman, A.K.; Towers, G.H.N.; Balza, F. Effects of sesquiterpene lactones on development of Aedes atropalpus and relation to partition coefficient. J. Nat. Prod. 1985, 48, 581–584. [Google Scholar] [CrossRef]
- Picman, A.K.; Picman, J. Effect of selected pseudoguaianolides on survival of the flour beetle, Tribolium confusum. Biochem. Syst. Ecol. 1984, 12, 89–93. [Google Scholar] [CrossRef]
- Kaur, R.; Chahal, K.K.; Kang, B.K. Insecticidal potential of parthenin and its transformation products against Tribolium castaneum (Herbst). Pestic. Res. J. 2015, 27, 35–40. [Google Scholar]
- Reddy, S.G.E.; Dolma, S.K.; Verma, P.K.; Singh, B. Insecticidal activities of Parthenium hysterophorus L. extract and parthenin against diamondback moth, Plutella xylostella (L.) and aphid, Aphis craccivora Koch. Toxin Rev. 2018, 37, 161–165. [Google Scholar] [CrossRef]
- Char, M.B.S.; Shankarabhat, S. Parthenin: A growth inhibitor behaviour in different organisms. Experientia 1975, 31, 1164–1165. [Google Scholar] [CrossRef]
- Ganeshan, G.; Jayachandra. Antifungal activity of parthenin. Indian Phytopathol. 1993, 46, 193–194. [Google Scholar]
- Bhusal, D.R.; Ghimire, K.C.; Patel, P.; Bista, M.; Upadhyay, R.; Kumar, B. Temperature and altitude modulate feeding attributes of Mexican beetle, Zygogramma bicolorata Pallister on Parthenium hysterophorus. J Therm. Biol. 2020, 89, 102540. [Google Scholar] [CrossRef]
- Gupta, S.K.; Monika; Gupta, V.; Deepika. An overview of airborne contact dermatitis. Air Water Borne Dis. 2016, 5, 126. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.K.; Sethuraman, G.; Bhat, R. Evolution of clinical pattern of parthenium dermatitis: A study of 74 cases. Contact Dermatitis 2005, 53, 84–88. [Google Scholar] [CrossRef]
- Kaur, M.; Gupta, K.B.; Thakur, S.; Kaur, S.; Dhiman, M. Parthenium hysterophorus mediated inflammation and hyper-responsiveness via NF-κB pathway in human A549 lung cancer cell line. Environ. Toxicol. 2020, 35, 1241–1250. [Google Scholar] [CrossRef]
- Narasimhan, T.R.; Harindranath, N.; Premlata, S.; Murthy, B.S.K.; Rao, P.V.S. Toxicity of the sesquiterpene lactone parthenin to cultured bovine kidney cells. Planta Med. 1985, 51, 194–197. [Google Scholar] [CrossRef]
- Narasimhan, T.R.; Harindranath, N.; Kurup, C.K.; Rao, P.V. Effect of parthenin on mitochondrial oxidative phosphorylation. Biochem. Int. 1985, 11, 239–244. [Google Scholar]
- Božičević, A.; De Mieri, M.; Nassenstein, C.; Wiegand, S.; Hamburger, M. Secondary metabolites in allergic plant pollen samples modulate afferent neurons and murine tracheal rings. J. Nat. Prod. 2017, 80, 2953–2961. [Google Scholar] [CrossRef] [PubMed]
- Bajwa, A.A.; Weston, P.A.; Gurusinghe, S.; Latif, S.; Adkins, S.W.; Weston, L.A. Toxic potential and metabolic profiling of two Australian biotypes of the invasive plant parthenium weed (Parthenium hysterophorus L.). Toxins 2020, 12, 447. [Google Scholar] [CrossRef]
- Raoof, K.M.A.; Siddiqui, M.B. Allelotoxic effect of parthenin on cytomorphology of broad bean (Vicia faba L.). J. Saudi Soc. Agric. Sci. 2013, 12, 143–146. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-León, A.; Torres-Valencia, J.M.; Manríquez-Torres, J.J.; Alvarado-Rodríguez, J.G.; Cerda-García-Rojas, C.M.; Joseph-Nathan, P. The stereochemistry of the 1,3-dipolar cycloadditions of diazomethane to pseudoguaianolides. Tetrahedron: Asymmetry 2017, 28, 367–373. [Google Scholar] [CrossRef]
- Jolly, D.P.; Shikha, A.; Chhabra, B.R.; Dhillon, R.S. Growth regulatory response of parthenin and its derivatives. Pestic. Res. J. 2005, 17, 1–5. [Google Scholar]
- Bhutani, K.K.; Thakur, R.N. The microbiological transformation of parthenin by Beauveria bassiana and Sporotrichum pulverulentum. Phytochemistry 1991, 30, 3599–3600. [Google Scholar] [CrossRef]
- Duke, S.O.; Cedergreen, N.; Velini, E.D.; Belz, R.G. Hormesis: Is it an important factor in herbicide use and allelopathy? Outlook Pest Manage. 2006, 17, 29–33. [Google Scholar] [CrossRef]
- Rajiv, P.; Rajeshwari, S.; Yadav, R.H.; Rajendran, V. Vermiremediation: Detoxification of parthenin toxin from Parthenium weeds. J. Hazard. Mater. 2013, 262, 489–495. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, A.; Kaur, S.; Jandrotia, R.; Singh, H.P.; Batish, D.R.; Kohli, R.K.; Rana, V.S.; Shakil, N.A. Parthenin—A Sesquiterpene Lactone with Multifaceted Biological Activities: Insights and Prospects. Molecules 2021, 26, 5347. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26175347
Kaur A, Kaur S, Jandrotia R, Singh HP, Batish DR, Kohli RK, Rana VS, Shakil NA. Parthenin—A Sesquiterpene Lactone with Multifaceted Biological Activities: Insights and Prospects. Molecules. 2021; 26(17):5347. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26175347
Chicago/Turabian StyleKaur, Amarpreet, Shalinder Kaur, Rupali Jandrotia, Harminder Pal Singh, Daizy Rani Batish, Ravinder Kumar Kohli, Virendra Singh Rana, and Najam A. Shakil. 2021. "Parthenin—A Sesquiterpene Lactone with Multifaceted Biological Activities: Insights and Prospects" Molecules 26, no. 17: 5347. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26175347