
Dendrimers as Non-Viral Vectors in Gene-Directed Enzyme Prodrug Therapy

 , ,
, ,  , , ,
, , ,  , , and
, , and
Abstract
:1. Introduction
- -
- -
- -
- Gene Directed Enzyme Prodrug Therapy (GDEPT) = uses a gene encoding an enzyme that is delivered at the target site through a carrier. The terminology of Genetic Prodrug Activation Therapy (GPAT) [1] is also used in the literature.
- -
- Polymer Directed Enzyme Prodrug Therapy (PDEPT) = passive targeting approach using a prodrug-conjugated polymer, followed by the administration of an enzyme-polymer conjugate, to achieve site-specific activation [8].
2. GDEPT Strategy
- (i)
- Chimeric circular plasmid DNAs (pDNA - plasmid DNA is a small molecule of physically separated DNA and distinct from chromosomal DNA, commonly used for gene duplication, transfer, and manipulation), which are hybrid plasmids containing a specific gene of interest (e.g., a gene encoding an easy-to-follow protein, such as luciferase or a fluorescent protein) [33];
- (ii)
- Messenger RNA (mRNA);
- (iii)
- Short RNA fragments, such as: siRNA, short-interfering RNA or rate-reducing RNA, is a double-stranded RNA that provides interference between gene expression and complementary nucleotide sequences; miRNA, a non-coding RNA molecule that functions by regulating the expression of post-transcription genes; short hairpin RNAs;
- (iv)
- (i)
- Specificity and selectivity: gene expression can be controlled by tumor cell-specific promoters/specific antigens, enzymes and their associated reactions, and can be directed into tumor cells without affecting healthy cells [37,38]. Due to the preferential conversion of the prodrug into the active drug in transfected tumor cells, with a minimal impact on healthy cells, the therapeutic index of the precursor is generally much higher than in conventional antineoplastic chemotherapy [39]. A wide range of tissue-specific promoters have been developed, such as: the human telomerase reverse transcriptase (hTERT) promoter, the carcinoembryonic antigen (CEA) promoter, the osteocalcin (OC) promoter [40]. More recently, other gene promoters have been developed, such as: auxin response factors (ARF), glucose-regulated protein (GRP78), CXC chemokine receptor-4 (CXCR4), osteopontin (OPN) [40,41,42,43,44]. Efficacy has been demonstrated for all of these promoters, but the only one to enter clinical trials was hTERT [45]. The main challenge is the low transcription profile of promoters, which causes the expression of suicide gene at a low level, which is insufficient to convert an optimal fraction of prodrug into cytotoxic molecules. To overcome these obstacles, chimeric and artificial promoters are increasingly studied [46,47].
- (ii)
- Another advantage of this therapeutic strategy compared to conventional therapy is the presence of the bystander effect, which is especially applied in antineoplastic therapy [24].
2.1. Gene Distribution and Delivery Vectors in GDEPT
- (i)
- “Proton sponge theory”: first issued around the 1990s [95,96]. Following the protonation of polymers, their chain elongates due to electrostatic repulsion. Thus, it has been shown that the expansion of the space occupied by the polymer can contribute to the increase in the so-called “umbrella hypothesis” [97,98];
- (ii)
- Membrane destabilization: it has been shown through molecular dynamics that, by elongating the chains of some polymers (such as polyethylenimine), they might be able to interact with the endosomal membrane, leading to the formation of hydrophilic pores in the membrane lipid bilayer. Thus, the lipid bilayer can destabilize, contributing to the release of the polymer from the endosomal space [99,100];
- (iii)
2.2. Gene Delivery Mechanisms through nVV
3. Dendrimers in GDEPT Strategy
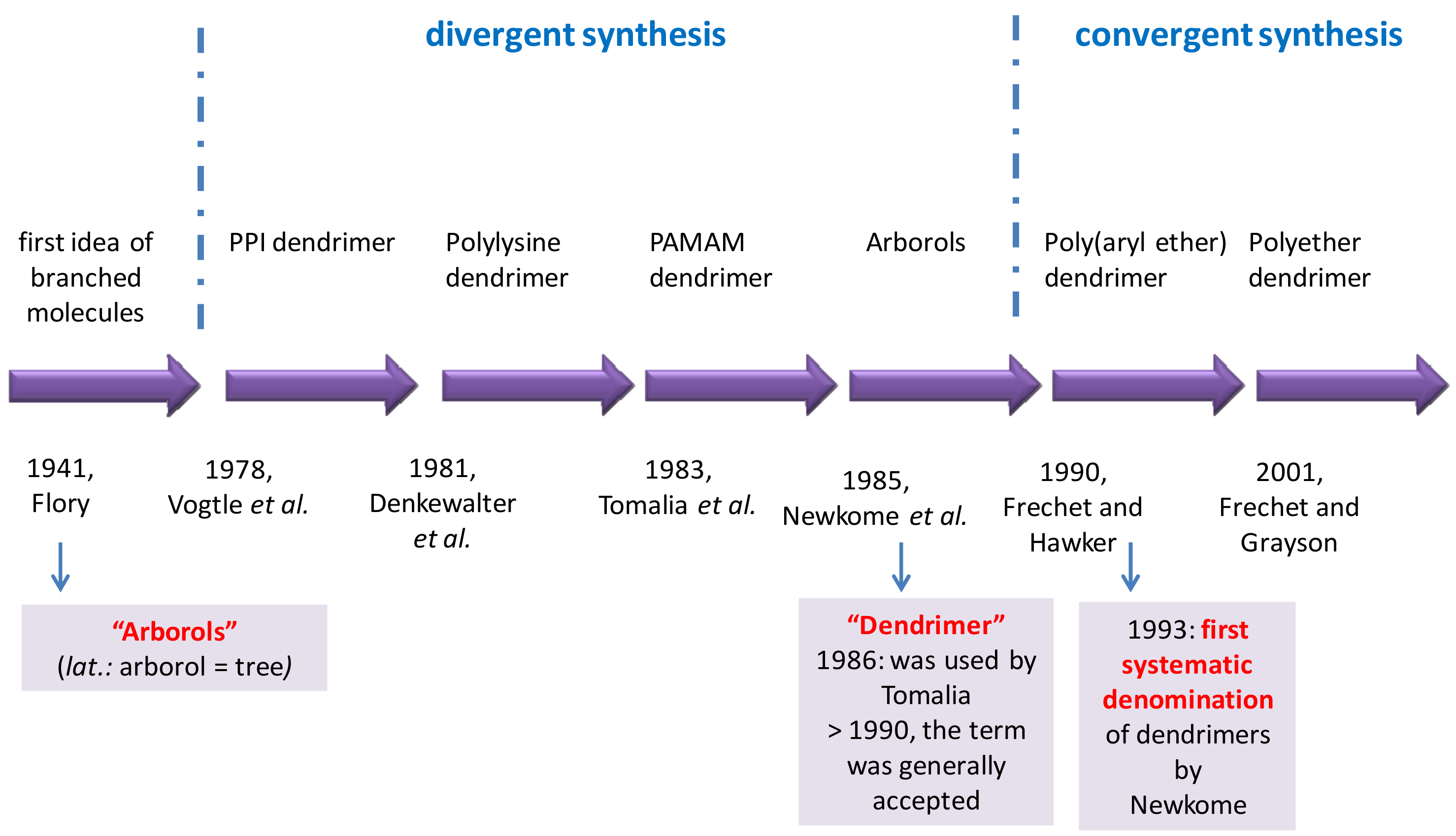
3.1. History of Dendrimers
3.2. Advantages of Dendrimers and Nanoparticles in the Distribution and Release of Drug Molecules at the Site of Action
3.3. The Efficiency of Dendrimers in GDEPT
- (i)
- (ii)
- (iii)
- (iv)
- (v)
- (vi)
- (vii)
- Ovary: siRNA that targets p70S6K in cancer stem cells [175];
- (viii)
- Skin: plasmid DNA encoding p73- inhibition of tumor growth for animals with A431 and B16 tumors [176];
- (ix)
- Blood: positive effects in acute myeloid leukemia using pivotal tumor-suppressor and negative regulator of FLT3 gene for the expression of tyrosine kinases 3 receptor [176].
3.4. In Vitro and In Vivo Studies on the Efficacy of Dendriplexes
3.5. The Stability of Dendrimers and Dendriplexes
3.6. The Toxicity of Dendrimers
3.7. Toxicity Modulation Strategies
- 1)
- PAMAM dendrimers are the most studied dendrimers, representing the dendrimers with the most important potential in gene therapy [216]. Conjugates of a siRNA antibody used in antineoplastic therapy with cationic dendrimers, PAMAM, carbosilane and phosphorus, were studied by quantitative measurements of their fluorescence intensities, zeta potential, light scattering, and dynamic scattering. The complexes were stable and had the potential to protect against nucleolytic separation of siRNA [180,217]. In antineoplastic therapy, the efficacy of transfection in HeLa and HL-60 cells was evaluated by the dendrimer–siRNA complex and the use of siRNA “cocktails”. Phosphoric dendrimers containing siRNA “cocktails” were characterized by the highest transfection rates and cytotoxicity [198,218]. Figure 9 schematically shows how to obtain conjugated phosphorus dendrimers and dendriplexes.
- 2)
- Obtaining dodecylated dendrimers: strategy used in PAMAM dendrimers of generations G2, G3 and G4 for the distribution of siRNA of luciferase-directed BCL-2 genes [219,220]. Remarkable results were obtained by dodecylated G4 dendrimers, with the best delivery efficiency, the 3D spherical shape and the introduction of gold nanoparticles in the central structure of the dendrimers being the most efficient (cell survival rate of over 90%). The ability of dendrimers to compact pDNA was greatly increased, leading to appropriately high delivery of the therapeutic gene [221]. Cyclododecylated dendrimers have been shown to be much more efficient and biocompatible than dodecylated analogs [219,222]. Moreover, the use of several radicals was studied in order to increase biocompatibility (Table 3) [222].
- 3)
- PAMAM G4 dendrimers with Arg terminal groups were developed as efficient nanovectors for the functional delivery of mRNA to capitalize on the unique properties of poly(amidoamine) dendrimers with triethanolamine nucleus (TEA) [223,224]. These dendrimers are structurally flexible and have been shown to be effective in delivering siRNA with numerous advantages in the cell penetration process: the 4th generation dendrimer in this category (G4Arg) has formed stable dendriplexes with siRNA, leading to an improved cellular uptake compared to its dendrimer analogue, which does not carry arginine [224]. In addition, the G4Arg dendrimer has no distinct toxicity. Thus, the addition of Arg radicals to the surface of a dendrimer has been shown to be a favorable option, with the G4Arg dendrimer (Figure 11) being an extremely promising nanovector for the efficient delivery of siRNA with high potential for other therapeutic applications [225].
- 4)
- Fluorinated dendrimers have been studied with the aim of improving transfection and reducing toxicity in gene delivery and improving the release strategy at the target site. Fluorinated PAMAM G5 dendrimers with extremely low N/P ratios (Figure 12) were tested on human embryonic kidney cells (HEK293) and human cervix-carcinoma cell (HeLa) cells. These dendrimers are characterized by serum resistance and elevated effectiveness in the process of gene transfection. G5 dendrimers are the most efficient in providing genes with a minimum toxicity, and the effectiveness of transfection depends on the degree of fluorination [226,227].
- 5)
- In addition to PAMAM dendrimers, different generations of PPI dendrimers have been studied as genetic vectors. Thus, fluorinated PPI dendritic polymers, G3, G4 and G5, were synthesized to improve the transfection efficiency and reduce the cytotoxicity of PPI dendrimers. Fluorinated PPI dendrimers showed a higher transfection efficiency than branched poly(ethyleneimine) and Arg-modified dendrimers in HEK293 and HeLa cells [182].
- 6)
- Other structural modulations applied to dendrimers:
- (i)
- (ii)
- (iii)
- (iv)
- Generation 4 phosphorhydrazone dendrimer with terminal functions modified with piperidine residues (9-G4), used as a dual nVV for both a singular substance, 5-FU, and a cocktail of anti-cancer siRNA, capable of influencing downregulated anti-apoptotic genes (BCL-xL, BCL-2, MCL-1) [165,189,239]. The effect on human cervical carcinoma HeLa cells showed a considerable increase in the cytotoxic effect at low doses of the cytotoxic therapeutic agent 5-FU by complexation with cocktail dendriplexes 9-G4 / siRNA [198,240].
- (v)
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| 5-FU | 5-fluorouracil |
| AAV | adeno-associated virus |
| ADEPT | antibody directed enzyme prodrug therapy |
| ARF | auxin response factors |
| ARF6 | adenosine diphosphate-ribosylation factor 6 |
| ASO | antisense oligonucleotides |
| CCL20 | chemokine (C-C motif) ligand 20 |
| CCR6 | C-C motif chemokine receptor 6 |
| CD | cyclodextrin |
| CEA | carcinoembryonic antigen |
| CRISPR | clustered regularly interspaced short palindromic repeats |
| CXCR4 | CXC chemokine receptor-4 |
| D | diffusion coefficient |
| DEP | dendrimer drug delivery system |
| EDA | ethylenediamine |
| ELAVL1 | embryonic lethal abnormal vision-like 1 |
| FA-3DNA | folic acid derivatized DNA dendrimer nanocarrier |
| G | generation of dendrimer |
| GDEPT | gene-directed enzyme prodrug therapy |
| GPAT | genetic prodrug activation therapy |
| GRAF1 | GTPase regulator associated with focal adhesion kinase-1 |
| GRP78 | glucose-regulated protein |
| HMW | high molecular weight |
| HSV | herpes simplex virus |
| hTERT | human telomerase reverse transcriptase |
| HuR | human antigen R |
| METase | methioninase |
| miRNA | microRNA |
| mRNA | messenger RNA |
| N | ionizable amino groups |
| N/P | ratio between ionizable amino groups and phosphate groups |
| NA | nucleic acid |
| NP | nanoparticle |
| nVGC | non-viral vector/gene complex |
| nVV | non-viral vector |
| OC | osteocalcin |
| OPN | osteopontin |
| P | phosphate group |
| paCD | polycationic amphiphilic CD |
| PAMAM | poly(amidoamine) dendrimer |
| Pc | partition coefficient, |
| PDEPT | polymer directed enzyme prodrug therapy |
| PDMAEMA | poly(2-(N,N-dimethylamino)ethyl methacrylate) dendrimer |
| pDNA | plasmid DNA |
| PEG | polyethlene glycol |
| PEI | polyethylenimine or polyaziridine |
| PLL | lysine polypeptide polymer |
| PPI | poly(propyleneimine) polymer |
| PPI-G4 | poly(propyleneimine) G4 dendrimers |
| RG7388 | selective MDM2 inhibitor that blocks the binding of MDM2 |
| SCID | severe combined immunodeficiency |
| siRNA | short-interfering RNA |
| TEA | triethanolamine |
| TNFα | tumor necrosis factor alpha |
| VDEPT | virus directed enzyme prodrug therapy |
| VV | viral vector |
References
- Sirhan, J.; Karaman, R. Gene directed enzyme prodrug therapy (GDEPT). In Prodrugs Design A New Era; Nova Science Publishers, Inc.: New York, NY, USA, 2014; pp. 210–232. [Google Scholar]
- Singh, Y.; Palombo, M.; Sinko, P. Recent Trends in Targeted Anticancer Prodrug and Conjugate Design. Curr. Med. Chem. 2008, 15, 1802–1826. [Google Scholar] [CrossRef] [Green Version]
- Ginn, S.L.; Amaya, A.K.; Alexander, I.E.; Edelstein, M.; Abedi, M.R. Gene therapy clinical trials worldwide to 2017: An update. J. Gene Med. 2018, 20, e3015. [Google Scholar] [CrossRef] [PubMed]
- Niculescu-Duvaz, I.; Springer, C.J. Antibody-directed enzyme prodrug therapy (ADEPT): A targeting strategy in cancer chemotherapy. Curr. Med. Chem. 1995, 2, 687–706. [Google Scholar]
- Phillips, G.L. Antibody-Drug Conjugates and Immunotoxins: From Pre-Clinical Development to Therapeutic Applications; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-5455-7. [Google Scholar]
- Wold, W.; Toth, K. Adenovirus Vectors for Gene Therapy, Vaccination and Cancer Gene Therapy. Curr. Gene Ther. 2013, 13, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Majhen, D.; Calderon, H.; Chandra, N.; Fajardo, C.A.; Rajan, A.; Alemany, R.; Custers, J. Adenovirus-based vaccines for fighting infectious diseases and cancer: Progress in the field. Hum. Gene Ther. 2014, 25, 301–317. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, G.; Tiwari, R.; Bannerjee, S.; Bhati, L.; Pandey, S.; Pandey, P.; Sriwastawa, B. Drug delivery systems: An updated review. Int. J. Pharm. Investig. 2012, 2, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; McLeod, H.L. Strategies for Enzyme/Prodrug Cancer Therapy. Clin. Cancer Res. 2001, 7, 3314–3324. [Google Scholar]
- Rubanyi, G.M. The future of human gene therapy. Mol. Asp. Med. 2001, 22, 113–142. [Google Scholar] [CrossRef]
- Mulligan, R.C. The basic science of gene therapy. Science 1993, 260, 926–932. [Google Scholar] [CrossRef]
- Yellepeddi, V.K. Vectors for non-viral gene delivery-Clinical and Biomedical applications. Austin Ther. 2015, 2, 1014. [Google Scholar]
- Rai, D.B.; Pooja, D.; Kulhari, H. Dendrimers in gene delivery. In Pharmaceutical Applications of Dendrimers; Elsevier: Amsterdam, The Netherlands; Oxford, UK; Cambridge, UK, 2019; pp. 211–231. ISBN 9780128145272. [Google Scholar]
- Lostalé-Seijo, I.; Montenegro, J. Synthetic materials at the forefront of gene delivery. Nat. Rev. Chem. 2018, 2, 258–277. [Google Scholar] [CrossRef]
- Wirth, T.; Parker, N.; Ylä-Herttuala, S. History of gene therapy. Gene 2013, 525, 162–169. [Google Scholar] [CrossRef]
- Baldi, L.; Hacker, D.L.; Adam, M.; Wurm, F.M. Recombinant protein production by large-scale transient gene expression in mammalian cells: State of the art and future perspectives. Biotechnol. Lett. 2007, 29, 677–684. [Google Scholar] [CrossRef] [Green Version]
- Bandaranayake, A.D.; Almo, S.C. Recent advances in mammalian protein production. FEBS Lett. 2014, 588, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Kale, V.; Chen, M. Gene-Directed Enzyme Prodrug Therapy. AAPS J. 2015, 17, 102–110. [Google Scholar] [CrossRef] [Green Version]
- Staquicini, F.I.; Ozawa, M.G.; Moya, C.A.; Driessen, W.H.; Barbu, E.M.; Nishimori, H.; Soghomonyan, S.; Flores, L.G.; Liang, X.; Paolillo, V.; et al. Systemic combinatorial peptide selection yields a non-canonical iron-mimicry mechanism for targeting tumors in a mouse model of human glioblastoma. J. Clin. Investig. 2011, 121, 161–173. [Google Scholar] [CrossRef]
- Wolff, J.A.; Lederberg, J. An Early History of Gene Transfer and Therapy. Hum. Gene Ther. 1994, 5, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Lederberg, J. What the Double Helix (1953) Has Meant for Basic Biomedical Science: A Personal Commentary. JAMA 1993, 269, 1981–1985. [Google Scholar] [CrossRef] [PubMed]
- Anderson, W.F.; Fletcher, J.C. Gene Therapy in Human Beings: When Is It Ethical to Begin? N. Engl. J. Med. 2009, 303, 1293–1297. [Google Scholar] [CrossRef]
- Both, G.W. Recent progress in gene-directed enzyme prodrug therapy: An emerging cancer treatment. Curr. Opin. Mol. Ther. 2009, 11, 421–432. [Google Scholar] [PubMed]
- Moolten, F.L. Tumor Chemosensitivity Conferred by Inserted Herpes Thymidine Kinase Genes: Paradigm for a Prospective Cancer Control Strategy1. Cancer Res. 1986, 46, 5276–5281. [Google Scholar]
- Dachs, G.U.; Hunt, M.A.; Syddall, S.; Singleton, D.C.; Patterson, A.V. Bystander or no bystander for gene directed enzyme prodrug therapy. Molecules 2009, 14, 4517–4545. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K. Viral Vectors in Gene Therapy. Diseases 2018, 6, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshami, A.A.; Saavedra, A.; Zhang, H.; Kucharczuk, J.C.; Spray, D.C.; Fishman, G.I.; Amin, K.M.; Kaiser, L.R.; Albelda, S.M. Gap junctions play a role in the “bystander effect” of the herpes simplex virus thymidine kinase/ganciclovir system in vitro. Gene Ther. 1996, 3, 85–92. [Google Scholar] [PubMed]
- Xiao, J.; Zhang, G.; Qiu, P.; Liu, X.; Wu, Y.; Du, B.; Li, J.; Zhou, J.; Li, J.; Tan, Y. Tanshinone IIA Increases the Bystander Effect of Herpes Simplex Virus Thymidine Kinase/Ganciclovir Gene Therapy via Enhanced Gap Junctional Intercellular Communication. PLoS ONE 2013, 8, e67662. [Google Scholar] [CrossRef] [PubMed]
- Denny, W.A. Prodrugs for gene-directed enzyme-prodrug therapy (suicide gene therapy). J. Biomed. Biotechnol. 2003, 2003, 48–70. [Google Scholar] [CrossRef] [Green Version]
- Florence, A.T.; Attwood, D. Physicochemical Principles of Pharmacy: In Manufacture, Formulation, and Clinical Use; Pharmaceutical Press: London, UK, 2015; ISBN 9780857112583. [Google Scholar]
- Pandit, N.K.; Soltis, R.P. Introduction to the Pharmaceutical Sciences: An Integrated Approach; Lippincott Williams & Wilkins: Baltimore, MA, USA; Philadelphia, PA, USA, 2012; ISBN 978-1-60-913001-5. [Google Scholar]
- Pan, J.J.; Zhang, S.W.; Chen, C.B.; Xiao, S.W.; Sun, Y.; Liu, C.Q.; Su, X.; Li, D.M.; Xu, G.; Xu, B.; et al. Effect of recombinant adenovirus-p53 combined with radiotherapy on long-term prognosis of advanced nasopharyngeal carcinoma. J. Clin. Oncol. 2009, 27, 799–804. [Google Scholar] [CrossRef] [Green Version]
- Caminade, A.M. Phosphorus dendrimers as nanotools against cancers. Molecules 2020, 25, 3333. [Google Scholar] [CrossRef]
- Wong, J.K.; Mohseni, R.; Hamidieh, A.A.; MacLaren, R.E.; Habib, N.; Seifalian, A.M. Will Nanotechnology Bring New Hope for Gene Delivery? Trends Biotechnol. 2017, 35, 434–451. [Google Scholar] [CrossRef]
- Xu, W.; Jiang, X.; Huang, L. RNA interference technology. In Comprehensive Biotechnology; Elsevier: Amsterdam, The Netherlands; Oxford, UK; Cambridge, UK, 2019; pp. 560–575. ISBN 9780444640475. [Google Scholar]
- Aied, A.; Greiser, U.; Pandit, A.; Wang, W. Polymer gene delivery: Overcoming the obstacles. Drug Discov. Today 2013, 18, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Ribot, E.J.; Miraux, S.; Konsman, J.P.; Bouchaud, V.; Pourtau, L.; Delville, M.H.; Franconi, J.M.; Thiaudière, E.; Voisin, P.J. In vivo MR tracking of therapeutic microglia to a human glioma model. NMR Biomed. 2011, 24, 1361–1368. [Google Scholar] [CrossRef]
- Greco, O.; Dachs, G.U. Gene Directed Enzyme/Prodrug Therapy of Cancer: Historical Appraisal and Future Prospectives. J. Cell. Physiol. 2001, 187, 22–36. [Google Scholar] [CrossRef]
- Karjoo, Z.; Ganapathy, V.; Hatefi, A. Gene-Directed Enzyme Prodrug Cancer Therapy. In Gene Therapy of Cancer, 3rd ed.; Elsevier: San Diego, CA, USA; London, UK; Waltham, MA, USA, 2013; pp. 77–91. [Google Scholar] [CrossRef]
- Kurayoshi, K.; Ozono, E.; Iwanaga, R.; Bradford, A.P.; Komori, H.; Ohtani, K. Cancer cell specific cytotoxic gene expression mediated by ARF tumor suppressor promoter constructs. Biochem. Biophys. Res. Commun. 2014, 450, 240–246. [Google Scholar] [CrossRef]
- Kia, A.; Przystal, J.M.; Nianiaris, N.; Mazarakis, N.D.; Mintz, P.J.; Hajitou, A. Dual systemic tumor targeting with ligand-directed phage and Grp78 promoter induces tumor regression. Mol. Cancer Ther. 2012, 11, 2566–2577. [Google Scholar] [CrossRef] [Green Version]
- Nemunaitis, J.; Tong, A.W.; Nemunaitis, M.; Senzer, N.; Phadke, A.P.; Bedell, C.; Adams, N.; Zhang, Y.A.; Maples, P.B.; Chen, S.; et al. A phase i study of telomerase-specific replication competent oncolytic adenovirus (telomelysin) for various solid tumors. Mol. Ther. 2010, 18, 429–434. [Google Scholar] [CrossRef]
- Zhu, Z.B.; Makhija, S.K.; Lu, B.; Wang, M.; Kaliberova, L.; Liu, B.; Rivera, A.A.; Nettelbeck, D.M.; Mahasreshti, P.J.; Leath, C.A.; et al. Transcriptional targeting of adenoviral vector through the CXCR4 tumor-specific promoter. Gene Ther. 2004, 11, 645–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.G.; Godbey, W.T. The Potential of the Human Osteopontin Promoter and Single-Nucleotide Polymorphisms for Targeted Cancer Gene Therapy. Curr. Gene Ther. 2014, 15, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Kanegae, Y.; Terashima, M.; Kondo, S.; Fukuda, H.; Maekawa, A.; Pei, Z.; Saito, I. High-level expression by tissue/cancer-specific promoter with strict specificity using a single-adenoviral vector. Nucleic Acids Res. 2011, 39, e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuah, M.K.; Petrus, I.; De Bleser, P.; Le Guiner, C.; Gernoux, G.; Adjali, O.; Nair, N.; Willems, J.; Evens, H.; Rincon, M.Y.; et al. Liver-specific transcriptional modules identified by genome-wide in silico analysis enable efficient gene therapy in mice and non-human primates. Mol. Ther. 2014, 22, 1605–1613. [Google Scholar] [CrossRef] [Green Version]
- Fakhiri, J.; Schneider, M.A.; Puschhof, J.; Stanifer, M.; Schildgen, V.; Holderbach, S.; Voss, Y.; El Andari, J.; Schildgen, O.; Boulant, S.; et al. Novel Chimeric Gene Therapy Vectors Based on Adeno-Associated Virus and Four Different Mammalian Bocaviruses. Mol. Ther. Methods Clin. Dev. 2019, 12, 202–222. [Google Scholar] [CrossRef] [Green Version]
- Piotrowski-Daspit, A.S.; Kauffman, A.C.; Bracaglia, L.G.; Saltzman, W.M. Polymeric vehicles for nucleic acid delivery. Adv. Drug Deliv. Rev. 2020, 156, 119–132. [Google Scholar] [CrossRef]
- Pan, X.; Veroniaina, H.; Su, N.; Sha, K.; Jiang, F.; Wu, Z.; Qi, X. Applications and developments of gene therapy drug delivery systems for genetic diseases. Asian J. Pharm. Sci. 2021. [Google Scholar] [CrossRef]
- Belete, T.M. The Current Status of Gene Therapy for the Treatment of Cancer. Biologics 2021, 15, 67–77. [Google Scholar] [CrossRef]
- Nayerossadat, N.; Maedeh, T.; Ali, P.A. Viral and nonviral delivery systems for gene delivery. Adv. Biomed. Res. 2012, 1, 27. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Glass, Z.; Huang, M.; Chen, Z.Y.; Xu, Q. Ex vivo cell-based CRISPR/Cas9 genome editing for therapeutic applications. Biomaterials 2020, 234, 119711. [Google Scholar] [CrossRef] [PubMed]
- Wilbie, D.; Walther, J.; Mastrobattista, E. Delivery Aspects of CRISPR/Cas for in Vivo Genome Editing. Acc. Chem. Res. 2019, 52, 1555–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horodecka, K.; Düchler, M. CRISPR/Cas9: Principle, Applications, and Delivery through Extracellular Vesicles. Int. J. Mol. Sci. 2021, 22, 6072. [Google Scholar] [CrossRef]
- Hromic-Jahjefendic, A.; Lundstrom, K. Viral vector-based melanoma gene therapy. Biomedicines 2020, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Vijayakumar, G.; McCroskery, S.; Palese, P. Engineering Newcastle Disease Virus as an Oncolytic Vector for Intratumoral Delivery of Immune Checkpoint Inhibitors and Immunocytokines. J. Virol. 2020, 94, e01677-19. [Google Scholar] [CrossRef]
- Corti, A.; Pastorino, F.; Curnis, F.; Arap, W.; Ponzoni, M.; Pasqualini, R. Targeted drug delivery and penetration into solid tumors. Med. Res. Rev. 2012, 32, 1078–1091. [Google Scholar] [CrossRef]
- Tamguney, T.M.; Korn, W.M.; McCormick, F. Gene Therapy and Oncolytic Viruses. In The Molecular Basis of Cancer, 4th ed.; Elsevier Inc.: Philadelphia, PA, USA, 2015; pp. 753–768. ISBN 9781455740666. [Google Scholar]
- Naso, M.F.; Tomkowicz, B.; Perry, W.L., III; Strohl, W.R. Adeno-Associated Virus (AAV) as a Vector for Gene Therapy. Biodrugs 2017, 31, 317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milone, M.C.; O’Doherty, U. Clinical use of lentiviral vectors. Leukemia 2018, 32, 1529. [Google Scholar] [CrossRef] [PubMed]
- De Garibay, A.P. Endocytosis in gene therapy with non-viral vectors. Wien. Med. Wochenschr. 2016, 166, 227–235. [Google Scholar] [CrossRef]
- Lundstrom, K. New frontiers in oncolytic viruses: Optimizing and selecting for virus strains with improved efficacy. Biol. Targets Ther. 2018, 12, 43–60. [Google Scholar] [CrossRef] [Green Version]
- Park, H.J.; Yang, F.; Cho, S.W. Nonviral delivery of genetic medicine for therapeutic angiogenesis. Adv. Drug Deliv. Rev. 2012, 64, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.H.; Chang, Y.H.; Lin, S.Y.; Li, K.C.; Hu, Y.C. Recent progresses in gene delivery-based bone tissue engineering. Biotechnol. Adv. 2013, 31, 1695–1706. [Google Scholar] [CrossRef]
- Wu, P.; Chen, H.; Jin, R.; Weng, T.; Ho, J.K.; You, C.; Zhang, L.; Wang, X.; Han, C. Non-viral gene delivery systems for tissue repair and regeneration. J. Transl. Med. 2018, 16, 29. [Google Scholar] [CrossRef] [Green Version]
- Ramamoorth, M.; Narvekar, A. Non viral vectors in gene therapy- an overview. J. Clin. Diagn. Res. 2015, 9, GE01–GE06. [Google Scholar] [CrossRef]
- Liu, N.; Hao, Y.; Yin, Z.; Ma, M.; Wang, L.; Zhang, X. Self-assembled human serum albumin-coated complexes for gene delivery with improved transfection. Pharmazie 2012, 67, 174–181. [Google Scholar] [CrossRef]
- Cooper, R.C.; Yang, H. Duplex of Polyamidoamine Dendrimer/Custom-Designed Nuclear-Localization Sequence Peptide for Enhanced Gene Delivery. Bioelectricity 2020, 2, 150–157. [Google Scholar] [CrossRef]
- Aggarwal, R.; Targhotra, M.; Kumar, B.; Sahoo, P.; Chauhan, M.K. Polyplex: A Promising Gene Delivery System. Int. J. Pharm. Sci. Nanotechnol. 2019, 12, 4681–4686. [Google Scholar] [CrossRef]
- Tarach, P.; Janaszewska, A. Recent Advances in Preclinical Research Using PAMAM Dendrimers for Cancer Gene Therapy. Int. J. Mol. Sci. 2021, 22, 2912. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.R.; Kumar, M.; Murumkar, P.R.; Hazari, P.P.; Mishra, A.K. Gemini Amphiphile-Based Lipoplexes for Efficient Gene Delivery: Synthesis, Formulation Development, Characterization, Gene Transfection, and Biodistribution Studies. Acs Omega 2018, 3, 11802–11816. [Google Scholar] [CrossRef] [PubMed]
- Paris, J.L.; Coelho, F.; Teixeira, A.; Diéguez, L.; Silva, B.F.; Abalde-Cela, S. In Vitro Evaluation of Lipopolyplexes for Gene Transfection: Comparing 2D, 3D and Microdroplet-Enabled Cell Culture. Molecules 2020, 25, 3277. [Google Scholar] [CrossRef]
- Tariq, I.; Ali, M.Y.; Sohail, M.F.; Amin, M.U.; Ali, S.; Bukhari, N.I.; Raza, A.; Pinnapireddy, S.R.; Schäfer, J.; Bakowsky, U. Lipodendriplexes mediated enhanced gene delivery: A cellular to pre-clinical investigation. Sci. Rep. 2020, 10, 21446. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, C.; Sun, Z.; Wu, Y.; You, W.; Mao, Z.; Wang, W. Mesenchymal Stem Cells Engineered by Nonviral Vectors: A Powerful Tool in Cancer Gene Therapy. Pharmaceutics 2021, 13, 913. [Google Scholar] [CrossRef] [PubMed]
- Kesse, S.; Boakye-Yiadom, K.O.; Ochete, B.O.; Opoku-Damoah, Y.; Akhtar, F.; Filli, M.S.; Farooq, M.A.; Aquib, M.; Mily, B.J.; Murtaza, G.; et al. Mesoporous Silica Nanomaterials: Versatile Nanocarriers for Cancer Theranostics and Drug and Gene Delivery. Pharmaceutics 2019, 11, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castillo, R.R.; Lozano, D.; Vallet-Regí, M. Mesoporous Silica Nanoparticles as Carriers for Therapeutic Biomolecules. Pharmaceutics 2020, 12, 432. [Google Scholar] [CrossRef]
- Aslan, C.; Kiaie, S.H.; Zolbanin, N.M.; Lotfinejad, P.; Ramezani, R.; Kashanchi, F.; Jafari, R. Exosomes for mRNA delivery: A novel biotherapeutic strategy with hurdles and hope. BMC Biotechnol. 2021, 21, 20. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, Z.; Wu, G.; Yang, Z.; Cui, Y.; Li, H.; Zhang, Y. Fluorinated Carbon Nanotube Superamphiphobic Coating for High-Efficiency and Long-Lasting Underwater Antibiofouling Surfaces. Acs Appl. Bio Mater. 2021, 4, 6351–6360. [Google Scholar] [CrossRef]
- Lin, L.; Fan, Y.; Gao, F.; Jin, L.; Li, D.; Sun, W.; Li, F.; Qin, P.; Shi, Q.; Shi, X.; et al. UTMD-Promoted Co-Delivery of Gemcitabine and miR-21 Inhibitor by Dendrimer-Entrapped Gold Nanoparticles for Pancreatic Cancer Therapy. Theranostics 2018, 8, 1923. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, D.M.; Correa, D.S.; Medeiros, E.S.; Oliveira, J.E.; Mattoso, L.H.C. Advances in Functional Polymer Nanofibers: From Spinning Fabrication Techniques to Recent Biomedical Applications. Acs Appl. Mater. Interfaces 2020, 12, 45673–45701. [Google Scholar] [CrossRef]
- Sofi, H.S.; Ashraf, R.; Khan, A.H.; Beigh, M.A.; Majeed, S.; Sheikh, F.A. Reconstructing nanofibers from natural polymers using surface functionalization approaches for applications in tissue engineering, drug delivery and biosensing devices. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 94, 1102–1124. [Google Scholar] [CrossRef] [PubMed]
- Dufay, M.; Jimenez, M.; Degoutin, S. Effect of Cold Plasma Treatment on Electrospun Nanofibers Properties: A Review. Acs Appl. Bio Mater. 2020, 3, 4696–4716. [Google Scholar] [CrossRef]
- Al Qtaish, N.; Gallego, I.; Villate-Beitia, I.; Sainz-Ramos, M.; López-Méndez, T.B.; Grijalvo, S.; Eritja, R.; Soto-Sánchez, C.; Martínez-Navarrete, G.; Fernández, E.; et al. Niosome-Based Approach for In Situ Gene Delivery to Retina and Brain Cortex as Immune-Privileged Tissues. Pharmaceutics 2020, 12, 198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carballo-Pedrares, N.; Kattar, A.; Concheiro, A.; Alvarez-Lorenzo, C.; Rey-Rico, A. Niosomes-based gene delivery systems for effective transfection of human mesenchymal stem cells. Mater. Sci. Eng. C 2021, 128, 112307. [Google Scholar] [CrossRef]
- Iqbal, S.; Blenner, M.; Alexander-Bryant, A.; Larsen, J. Polymersomes for Therapeutic Delivery of Protein and Nucleic Acid Macromolecules: From Design to Therapeutic Applications. Biomacromolecules 2020, 21, 1327–1350. [Google Scholar] [CrossRef]
- Sharma, A.K.; Prasher, P.; Aljabali, A.A.; Mishra, V.; Gandhi, H.; Kumar, S.; Mutalik, S.; Chellappan, D.K.; Tambuwala, M.M.; Dua, K.; et al. Emerging era of “somes”: Polymersomes as versatile drug delivery carrier for cancer diagnostics and therapy. Drug Deliv. Transl. Res. 2020, 10, 1171–1190. [Google Scholar] [CrossRef]
- Pack, D.W.; Hoffman, A.S.; Pun, S.; Stayton, P.S. Design and development of polymers for gene delivery. Nat. Rev. Drug Discov. 2005, 4, 581–593. [Google Scholar] [CrossRef]
- Majidi, A.; Nikkhah, M.; Sadeghian, F.; Hosseinkhani, S. Development of novel recombinant biomimetic chimeric MPG-based peptide as nanocarriers for gene delivery: Imitation of a real cargo. Eur. J. Pharm. Biopharm. 2016, 107, 191–204. [Google Scholar] [CrossRef]
- Kulkarni, M.; Greiser, U.; O’Brien, T.; Pandit, A. Liposomal gene delivery mediated by tissue-engineered scaffolds. Trends Biotechnol. 2010, 28, 28–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonetta, L. The inside scoop—Evaluating gene delivery methods. Nat. Methods 2005, 2, 875–882. [Google Scholar] [CrossRef]
- Bono, N.; Ponti, F.; Mantovani, D.; Candiani, G. Non-viral in vitro gene delivery: It is now time to set the bar! Pharmaceutics 2020, 12, 183. [Google Scholar] [CrossRef] [Green Version]
- Smedt, D.S.; Martens, T.F.; Remaut, K.; Demeester, J.; De Smedt, S.C.; Braeckmans, K. This item is the archived peer-reviewed author-version of: Intracellular delivery of nanomaterials: How to catch endosomal escape in the act intracellular delivery of nanomaterials: How to catch endosomal escape in the act. Nano Today 2014, 9, 344–364. [Google Scholar] [CrossRef] [Green Version]
- Pangarkar, C.; Dinh, A.T.; Mitragotri, S. Endocytic pathway rapidly delivers internalized molecules to lysosomes: An analysis of vesicle trafficking, clustering and mass transfer. J. Control. Release 2012, 162, 76–83. [Google Scholar] [CrossRef]
- Selby, L.I.; Cortez-Jugo, C.M.; Such, G.K.; Johnston, A.P. Nanoescapology: Progress toward understanding the endosomal escape of polymeric nanoparticles. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2017, 9, e1452. [Google Scholar] [CrossRef] [PubMed]
- Behr, J.P.; Demeneix, B.; Loeffler, J.P.; Perez-Mutul, J. Efficient gene transfer into mammalian primary endocrine cells with lipopolyamine-coated DNA. Proc. Natl. Acad. Sci. USA 1989, 86, 6982–6986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haensler, J.; Francis, C.; Szoka, J. Polyamidoamine cascade polymers mediate efficient transfection of cells in culture. Bioconjug. Chem. 2002, 4, 372–379. [Google Scholar] [CrossRef]
- Ziebarth, J.D.; Wang, Y. Understanding the Protonation Behavior of Linear Polyethylenimine in Solutions through Monte Carlo Simulations. Biomacromolecules 2009, 11, 29–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermeulen, L.M.; Brans, T.; Samal, S.K.; Dubruel, P.; Demeester, J.; De Smedt, S.C.; Remaut, K.; Braeckmans, K. Endosomal Size and Membrane Leakiness Influence Proton Sponge-Based Rupture of Endosomal Vesicles. ACS Nano 2018, 12, 2332–2345. [Google Scholar] [CrossRef] [Green Version]
- Choudhury, C.K.; Kumar, A.; Roy, S. Characterization of Conformation and Interaction of Gene Delivery Vector Polyethylenimine with Phospholipid Bilayer at Different Protonation State. Biomacromolecules 2013, 14, 3759–3768. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Yeo, Y. Controlled drug release from pharmaceutical nanocarriers. Chem. Eng. Sci. 2015, 125, 75–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behr, J.P. The Proton Sponge: A Trick to Enter Cells the Viruses Did Not Exploit. CHIMIA Int. J. Chem. 1997, 51, 34–36. [Google Scholar]
- Benjaminsen, R.V.; Mattebjerg, M.A.; Henriksen, J.R.; Moghimi, S.M.; Andresen, T.L. The Possible “Proton Sponge” Effect of Polyethylenimine (PEI) Does Not Include Change in Lysosomal pH. Mol. Ther. 2013, 21, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.; Kim, J.; Herrera, M.; Mukherjee, A.; Kabanov, A.V.; Sahay, G. Brief update on endocytosis of nanomedicines. Adv. Drug Deliv. Rev. 2019, 144, 90–111. [Google Scholar] [CrossRef]
- Wang, Y.; Zheng, M.; Meng, F.; Zhang, J.; Peng, R.; Zhong, Z. Branched Polyethylenimine Derivatives with Reductively Cleavable Periphery for Safe and Efficient In Vitro Gene Transfer. Biomacromolecules 2011, 12, 1032–1040. [Google Scholar] [CrossRef] [PubMed]
- Kano, A.; Moriyama, K.; Yamano, T.; Nakamura, I.; Shimada, N.; Maruyama, A. Grafting of poly(ethylene glycol) to poly-lysine augments its lifetime in blood circulation and accumulation in tumors without loss of the ability to associate with siRNA. J. Control. Release 2011, 149, 2–7. [Google Scholar] [CrossRef]
- Zakeri, A.; Kouhbanani, M.A.; Beheshtkhoo, N.; Beigi, V.; Mousavi, S.M.; Hashemi, S.A.; Karimi Zade, A.; Amani, A.M.; Savardashtaki, A.; Mirzaei, E.; et al. Polyethylenimine-based nanocarriers in co-delivery of drug and gene: A developing horizon. Nano Rev. Exp. 2018, 9, 1488497. [Google Scholar] [CrossRef] [Green Version]
- Tack, F.; Bakker, A.; Maes, S.; Dekeyser, N.; Bruining, M.; Elissen-Roman, C.; Janicot, M.; Brewster, M.; Janssen, H.M.; De Waal, B.F.; et al. Modified poly(propylene imine) dendrimers as effective transfection agents for catalytic DNA enzymes (DNAzymes). J. Drug Target. 2008, 14, 69–86. [Google Scholar] [CrossRef]
- Moghimi, S.M.; Symonds, P.; Murray, J.C.; Hunter, A.C.; Debska, G.; Szewczyk, A. A two-stage poly(ethylenimine)-mediated cytotoxicity: Implications for gene transfer/therapy. Mol. Ther. 2005, 11, 990–995. [Google Scholar] [CrossRef]
- Bugno, J.; Hsu, H.J.; Pearson, R.M.; Noh, H.; Hong, S. Size and surface charge of engineered poly(amidoamine) dendrimers modulate tumor accumulation and penetration: A model study using multicellular tumor spheroids. Mol. Pharm. 2016, 13, 2155–2163. [Google Scholar] [CrossRef] [PubMed]
- Moura, L.I.; Malfanti, A.; Peres, C.; Matos, A.I.; Guegain, E.; Sainz, V.; Zloh, M.; Vicent, M.J.; Florindo, H.F. Functionalized branched polymers: Promising immunomodulatory tools for the treatment of cancer and immune disorders. Mater. Horiz. 2019, 6, 1956–1973. [Google Scholar] [CrossRef]
- Fox, L.J.; Richardson, R.M.; Briscoe, W.H. PAMAM dendrimer-cell membrane interactions. Adv. Colloid Interface Sci. 2018, 257, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kesharwani, P.; Banerjee, S.; Gupta, U.; Mohd Amin, M.C.; Padhye, S.; Sarkar, F.H.; Iyer, A.K. PAMAM dendrimers as promising nanocarriers for RNAi therapeutics. Mater. Today 2015, 18, 565–572. [Google Scholar] [CrossRef]
- Kadlecova, Z.; Rajendra, Y.; Matasci, M.; Baldi, L.; Hacker, D.L.; Wurm, F.M.; Klok, H.A. DNA delivery with hyperbranched polylysine: A comparative study with linear and dendritic polylysine. J. Control. Release 2013, 169, 276–288. [Google Scholar] [CrossRef]
- Jiang, Y.; Arounleut, P.; Rheiner, S.; Bae, Y.; Kabanov, A.V.; Milligan, C.; Manickam, D.S. SOD1 nanozyme with reduced toxicity and MPS accumulation. J. Control. Release 2016, 231, 38–49. [Google Scholar] [CrossRef] [PubMed]
- Misselwitz, B.; Schmitt-Willich, H.; Ebert, W.; Frenzel, T.; Weinmann, H.J. Pharmacokinetics of Gadomer-17, a new dendritic magnetic resonance contrast agent. Magn. Reson. Mater. Phys. Biol. Med. 2001, 12, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kakkar, A. Designing Dendrimer and Miktoarm Polymer Based Multi-Tasking Nanocarriers for Efficient Medical Therapy. Molecules 2015, 20, 16987–17015. [Google Scholar] [CrossRef]
- Agarwal, S.; Zhang, Y.; Maji, S.; Greiner, A. PDMAEMA based gene delivery materials. Mater. Today 2012, 15, 388–393. [Google Scholar] [CrossRef]
- Santos, A.; Veiga, F.; Figueiras, A. Dendrimers as Pharmaceutical Excipients: Synthesis, Properties, Toxicity and Biomedical Applications. Materials 2020, 13, 65. [Google Scholar] [CrossRef] [Green Version]
- Trombino, S.; Curcio, F.; Poerio, T.; Pellegrino, M.; Russo, R.; Cassano, R. Chitosan Membranes Filled with Cyclosporine A as Possible Devices for Local Administration of Drugs in the Treatment of Breast Cancer. Molecules 2021, 26, 1889. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Howard, K.A.; Dong, M.; Andersen, M.; Rahbek, U.L.; Johnsen, M.G.; Hansen, O.C.; Besenbacher, F.; Kjems, J. The influence of polymeric properties on chitosan/siRNA nanoparticle formulation and gene silencing. Biomaterials 2007, 28, 1280–1288. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Tan, Y.F.; Wong, Y.S.; Liew, M.W.; Venkatraman, S. Recent Advances in Chitosan-Based Carriers for Gene Delivery. Mar. Drugs 2019, 17, 381. [Google Scholar] [CrossRef] [Green Version]
- Ortega-Caballero, F.; Mellet, C.O.; Le Gourriérec, L.; Guilloteau, N.; Di Giorgio, C.; Vierling, P.; Defaye, J.; García Fernández, J.M. Tailoring beta-cyclodextrin for DNA complexation and delivery by homogeneous functionalization at the secondary face. Org. Lett. 2008, 10, 5143–5146. [Google Scholar] [CrossRef]
- Lai, W.F. Cyclodextrins in non-viral gene delivery. Biomaterials 2014, 35, 401. [Google Scholar] [CrossRef]
- Pichon, C.; Billiet, L.; Midoux, P. Chemical vectors for gene delivery: Uptake and intracellular trafficking. Curr. Opin. Biotechnol. 2010, 21, 640–645. [Google Scholar] [CrossRef]
- Raftery, R.M.; Walsh, D.P.; Castaño, I.M.; Heise, A.; Duffy, G.P.; Cryan, S.A.; O’Brien, F.J. Delivering Nucleic-Acid Based Nanomedicines on Biomaterial Scaffolds for Orthopedic Tissue Repair: Challenges, Progress and Future Perspectives. Adv. Mater. 2016, 28, 5447–5469. [Google Scholar] [CrossRef] [PubMed]
- Braun, S. Non-viral Vector for Muscle-Mediated Gene Therapy. In Muscle Gene Therapy; Springer International Publishing: Cham, Switzerland, 2019; pp. 157–178. [Google Scholar]
- Pouton, C.W.; Seymour, L.W. Key issues in non-viral gene delivery. Adv. Drug Deliv. Rev. 2001, 46, 187–203. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.; Shao, N.; Hu, Z.; Chen, H.; Xu, L.; Wang, C.; Cheng, Y.; Xiao, J. Triazine-modified dendrimer for efficient TRAIL gene therapy in osteosarcoma. Acta Biomater. 2015, 17, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Palm, W. Metabolic functions of macropinocytosis. Philos. Trans. R. Soc. B 2019, 374, 20180285. [Google Scholar] [CrossRef] [PubMed]
- Xiang, S.; Zhang, X. Cellular Uptake Mechanism of Non-Viral Gene Delivery and Means for Improving Transfection Efficiency. In Gene Therapy—Tools and Potential Applications; IntechOpen: Rijeka, Croatia, 2013. [Google Scholar] [CrossRef] [Green Version]
- Do Carmo, D.R. Synthesis and a Preliminary Characterization of Poly(Propylene)Imine Hexadecylamine Dendrimer (DAB-Am-16) Modified with Methyl Acrylate. Am. Chem. Sci. J. 2013, 3, 314–324. [Google Scholar] [CrossRef]
- Augustus, E.N.; Allen, E.T.; Nimibofa, A.; Donbebe, W. A review of synthesis, characterization and applications of functionalized dendrimers. Am. J. Polym. Sci. 2017, 7, 8–14. [Google Scholar] [CrossRef]
- Gupta, A.; Dubey, S.; Mishra, M. Unique Structures, Properties and Applications of Dendrimers. J. Drug Deliv. Ther. 2018, 8, 328–339. [Google Scholar] [CrossRef]
- Abbasi, E.; Aval, S.F.; Akbarzadeh, A.; Milani, M.; Nasrabadi, H.T.; Joo, S.W.; Hanifehpour, Y.; Nejati-Koshki, K.; Pashaei-Asl, R. Dendrimers: Synthesis, applications, and properties. Nanoscale Res. Lett. 2014, 9, 247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faheem, A.M.; Abdelkader, D.H. Novel drug delivery systems. In Engineering Drug Delivery Systems; Woodhead Publishing: Cambridge, UK, 2019; pp. 1–16. [Google Scholar] [CrossRef]
- Wang, H.; Chang, H.; Zhang, Q.; Cheng, Y. Fabrication of Low-Generation Dendrimers into Nanostructures for Efficient and Nontoxic Gene Delivery. Top. Curr. Chem. 2017, 375, 62. [Google Scholar] [CrossRef]
- Mariyam, M.; Ghosal, K.; Thomas, S.; Kalarikkal, N.; Latha, M.S. Dendrimers: General Aspects, Applications and Structural Exploitations as Prodrug/Drug-delivery Vehicles in Current Medicine. Mini Rev. Med. Chem. 2018, 18, 439–457. [Google Scholar] [CrossRef]
- Cheng, Y.; Xu, Z.; Ma, M.; Xu, T. Dendrimers as drug carriers: Applications in different routes of drug administration. J. Pharm. Sci. 2008, 97, 123–143. [Google Scholar] [CrossRef]
- Bharali, D.J.; Khalil, M.; Gurbuz, M.; Simone, T.M.; Mousa, S.A. Nanoparticles and cancer therapy: A concise review with emphasis on dendrimers. Int. J. Nanomed. 2009, 4, 1–7. [Google Scholar]
- Chis, A.A.; Dobrea, C.; Morgovan, C.; Arseniu, A.M.; Rus, L.L.; Butuca, A.; Juncan, A.M.; Totan, M.; Vonica-Tincu, A.L.; Cormos, G.; et al. Applications and Limitations of Dendrimers in Biomedicine. Molecules 2020, 25, 3982. [Google Scholar] [CrossRef]
- Tomalia, D.A.; Naylor, A.M.; Goddard, W.A. Starburst Dendrimers: Molecular-Level Control of Size, Shape, Surface Chemistry, Topology, and Flexibility from Atoms to Macroscopic Matter. Angew. Chem. Int. Ed. Engl. 1990, 29, 138–175. [Google Scholar] [CrossRef]
- Fradet, A.; Chen, J.; Hellwich, K.H.; Horie, K.; Kahovec, J.; Mormann, W.; Stepto, R.F.; Vohlídal, J.; Wilks, E.S. Nomenclature and terminology for dendrimers with regular dendrons and for hyperbranched polymers (IUPAC Recommendations 2017). Pure Appl. Chem. 2019, 91, 523–561. [Google Scholar] [CrossRef] [Green Version]
- De Araújo, R.V.; da Silva Santos, S.; Ferreira, E.I.; Giarolla, J. New advances in general biomedical applications of PAMAM dendrimers. Molecules 2018, 23, 2849. [Google Scholar] [CrossRef] [Green Version]
- Sapra, R.; Verma, R.P.; Maurya, G.P.; Dhawan, S.; Babu, J.; Haridas, V. Designer Peptide and Protein Dendrimers: A Cross-Sectional Analysis. Chem. Rev. 2019, 119, 11391–11441. [Google Scholar] [CrossRef]
- Siafaka, P.I.; Okur, N.Ü.; Karavas, E.; Bikiaris, D.N. Surface Modified Multifunctional and Stimuli Responsive Nanoparticles for Drug Targeting: Current Status and Uses. Int. J. Mol. Sci. 2016, 17, 1440. [Google Scholar] [CrossRef]
- De Jong, W.H.; Borm, P.J. Drug delivery and nanoparticles: Applications and hazards. Int. J. Nanomed. 2008, 3, 133. [Google Scholar] [CrossRef] [Green Version]
- Golshan, M.; Rostami-Tapeh-Esmail, E.; Salami-Kalajahi, M.; Roghani-Mamaqani, H. A review on synthesis, photophysical properties, and applications of dendrimers with perylene core. Eur. Polym. J. 2020, 137, 109933. [Google Scholar] [CrossRef]
- Svenson, S.; Chauhan, A.S. Dendrimers for enhanced drug solubilization. Nanomedicine 2008, 3, 679–702. [Google Scholar] [CrossRef]
- Winalski, C.S.; Shortkroff, S.; Mulkern, R.V.; Schneider, E.; Rosen, G.M. Magnetic resonance relaxivity of dendrimer-linked nitroxides. Magn. Reson. Med. 2002, 48, 965–972. [Google Scholar] [CrossRef] [PubMed]
- Maliakal, A.J.; Turro, N.J.; Bosman, A.W.; Cornel, J.; Meijer, E.W. Relaxivity Studies on Dinitroxide and Polynitroxyl Functionalized Dendrimers: Effect of Electron Exchange and Structure on Paramagnetic Relaxation Enhancement. J. Phys. Chem. A 2003, 107, 8467–8475. [Google Scholar] [CrossRef]
- Zhang, S.; Lloveras, V.; Pulido, D.; Liko, F.; Pinto, L.F.; Albericio, F.; Royo, M.; Vidal-Gancedo, J. Radical Dendrimers Based on Biocompatible Oligoethylene Glycol Dendrimers as Contrast Agents for MRI. Pharmaceutics 2020, 12, 772. [Google Scholar] [CrossRef]
- Francese, G.; Dunand, F.A.; Loosli, C.; Merbach, A.E.; Decurtins, S. Functionalization of PAMAM dendrimers with nitronyl nitroxide radicals as models for the outer-sphere relaxation in dentritic potential MRI contrast agents. Magn. Reson. Chem. 2003, 41, 81–83. [Google Scholar] [CrossRef]
- Vanerio, N.; Stijnen, M.; de Mol, B.A.; Kock, L.M. Biomedical Applications of Photo- and Sono-Activated Rose Bengal: A Review. Photobiomodul. Photomed. Laser Surg. 2019, 37, 383–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.J.; Gorman, C.; Menegatti, S. Synthesis, structure, and function of internally functionalized dendrimers. J. Polym. Sci. 2021, 59, 10–28. [Google Scholar] [CrossRef]
- Esfand, R.; Tomalia, D.A. Poly(amidoamine) (PAMAM) dendrimers: From biomimicry to drug delivery and biomedical applications. Drug Discov. Today 2001, 6, 427–436. [Google Scholar] [CrossRef]
- Tomalia, D.A.; Fréchet, J.M. Discovery of dendrimers and dendritic polymers: A brief historical perspective. J. Polym. Sci. Part A Polym. Chem. 2002, 40, 2719–2728. [Google Scholar] [CrossRef]
- Tomalia, D.A.; Hall, M.; Hedstrand, D.M. Starburst Dendrimers. 3. The Importance of Branch Junction Symmetry in the Development of Topological Shell Molecules. J. Am. Chem. Soc. 1987, 109, 1601–1603. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhao, L.; Li, Y.; Xu, T. Design of biocompatible dendrimers for cancer diagnosis and therapy:Current status and future perspectives. Chem. Soc. Rev. 2011, 40, 2673–2703. [Google Scholar] [CrossRef]
- Xiong, Z.; Shen, M.; Shi, X. Dendrimer-based strategies for cancer therapy: Recent advances and future perspectives. Sci. China Mater. 2018, 61, 1387–1403. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Q.; Chang, H.; Cheng, Y. Surface-Engineered Dendrimers in Gene Delivery. Chem. Rev. 2015, 115, 5274–5300. [Google Scholar] [CrossRef]
- Chaplot, S.P.; Rupenthal, I.D. Dendrimers for gene delivery—A potential approach for ocular therapy? J. Pharm. Pharmacol. 2014, 66, 542–556. [Google Scholar] [CrossRef] [PubMed]
- Somani, S.; Blatchford, D.R.; Millington, O.; Stevenson, M.L.; Dufès, C. Transferrin-bearing polypropylenimine dendrimer for targeted gene delivery to the brain. J. Control. Release 2014, 188, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Somani, S.; Robb, G.; Pickard, B.S.; Dufès, C. Enhanced gene expression in the brain following intravenous administration of lactoferrin-bearing polypropylenimine dendriplex. J. Control. Release 2015, 217, 235–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, W.; Shao, K.; Huang, R.; Han, L.; Liu, Y.; Li, J.; Kuang, Y.; Ye, L.; Lou, J.; Jiang, C. Gene delivery targeted to the brain using an Angiopep-conjugated polyethyleneglycol-modified polyamidoamine dendrimer. Biomaterials 2009, 30, 6976–6985. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Zhang, A.; Wang, H.; Pu, P.; Jiang, X.; Kang, C.; Chang, J. Tat-BMPs-PAMAM conjugates enhance therapeutic effect of small interference RNA on U251 glioma cells in vitro and in vivo. Hum. Gene Ther. 2010, 21, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Mayilsamy, K.; Markoutsa, E.; Das, M.; Chopade, P.; Puro, D.; Kumar, A.; Gulick, D.; Willing, A.E.; Mohapatra, S.S.; Mohapatra, S. Treatment with shCCL20-CCR6 nanodendriplexes and human mesenchymal stem cell therapy improves pathology in mice with repeated traumatic brain injury. Nanomed. Nanotechnol. Biol. Med. 2020, 29, 102247. [Google Scholar] [CrossRef]
- Kukowska-Latallo, J.F.; Raczka, E.; Quintana, A.; Chen, C.; Rymaszewski, M.; Baker, J.R. Intravascular and Endobronchial DNA Delivery to Murine Lung Tissue Using a Novel, Nonviral Vector. Hum. Gene Ther. 2000, 11, 1385–1395. [Google Scholar] [CrossRef]
- Kang, W.Y.; Wang, H.L.; Wang, H.; Wang, X.F.; Wang, C.Z.; Fu, Q.H.; Ding, Q.L.; Wu, W.M.; Fang, Y.; Wang, Z.Y. In vivo transfection and expression of human coagulant factor VIII cDNA in mice. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2004, 12, 188–193. [Google Scholar]
- Shcharbin, D.; Janaszewska, A.; Klajnert-Maculewicz, B.; Ziemba, B.; Dzmitruk, V.; Halets, I.; Loznikova, S.; Shcharbina, N.; Milowska, K.; Ionov, M. How to study dendrimers and dendriplexes III. Biodistribution, pharmacokinetics and toxicity in vivo. J. Control. Release 2014, 181, 40–52. [Google Scholar] [CrossRef]
- Ding, J.J.; Guo, C.Y.; Cai, Q.L.; Lin, Y.H.; Wang, H. In vivo expression of green fluorescent protein gene and immunogenicity of ES312 vaccine both mediated by starburst polyamidoamine dendrimers. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2005, 27, 499–503. [Google Scholar]
- Mamede, M.; Saga, T.; Ishimori, T.; Higashi, T.; Sato, N.; Kobayashi, H.; Brechbiel, M.W.; Konishi, J. Hepatocyte targeting of 111In-labeled oligo-DNA with avidin or avidin-dendrimer complex. J. Control. Release 2004, 95, 133–141. [Google Scholar] [CrossRef]
- Maruyama-Tabata, H.; Harada, Y.; Matsumura, T.; Satoh, E.; Cui, F.; Iwai, M.; Kita, M.; Hibi, S.; Imanishi, J.; Sawada, T.; et al. Effective suicide gene therapy in vivo by EBV-based plasmid vector coupled with polyamidoamine dendrimer. Gene Ther. 2000, 7, 53–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Robaian, M.; Chiam, K.Y.; Blatchford, D.R.; Dufès, C. Therapeutic efficacy of intravenously administered transferrin-conjugated dendriplexes on prostate carcinomas. Nanomedicine 2014, 9, 421–434. [Google Scholar] [CrossRef] [Green Version]
- Altwaijry, N.; Somani, S.; Parkinson, J.A.; Tate, R.J.; Keating, P.; Warzecha, M.; Mackenzie, G.R.; Leung, H.Y.; Dufès, C. Regression of prostate tumors after intravenous administration of lactoferrin-bearing polypropylenimine dendriplexes encoding TNF-α, TRAIL, and interleukin-12. Drug Deliv. 2018, 25, 679–689. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Kala, S.; Yung, S.; Chan, T.M.; Cao, Y.; Jiang, Y.; Liu, X.; Giorgio, S.; Peng, L.; Wong, A.S. Blocking Stemness and Metastatic Properties of Ovarian Cancer Cells by Targeting p70S6K with Dendrimer Nanovector-Based siRNA Delivery. Mol. Ther. 2018, 26, 70. [Google Scholar] [CrossRef] [PubMed]
- Lemarié, F.; Croft, D.R.; Tate, R.J.; Ryan, K.M.; Dufès, C. Tumor regression following intravenous administration of a tumor-targeted p73 gene delivery system. Biomaterials 2012, 33, 2701–2709. [Google Scholar] [CrossRef]
- Zhang, C.; Pan, D.; Li, J.; Hu, J.; Bains, A.; Guys, N.; Zhu, H.; Li, X.; Luo, K.; Gong, Q.; et al. Enzyme-responsive peptide dendrimer-gemcitabine conjugate as a controlled-release drug delivery vehicle with enhanced antitumor efficacy. Acta Biomater. 2017, 55, 153–162. [Google Scholar] [CrossRef]
- Wang, S.; Wang, F.; Zhang, Q.; Cheng, Y. A core-shell structured polyplex for efficient and non-toxic gene delivery. J. Mater. Chem. B 2017, 5, 5101–5108. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.; Huang, S.; Chen, Z.; Li, Y.; Liu, K.; Liu, Y.; Du, L. Low cytotoxicity fluorescent PAMAM dendrimer as gene carriers for monitoring the delivery of siRNA. J. Nanopart. Res. 2015, 17, 385. [Google Scholar] [CrossRef]
- Ionov, M.; Lazniewska, J.; Dzmitruk, V.; Halets, I.; Loznikova, S.; Novopashina, D.; Apartsin, E.; Krasheninina, O.; Venyaminova, A.; Milowska, K.; et al. Anticancer siRNA cocktails as a novel tool to treat cancer cells. Part (A). Mechanisms of interaction. Int. J. Pharm. 2015, 485, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Li, J.; Fan, Y.; Qiu, J.; Cao, L.; Laurent, R.; Mignani, S.; Caminade, A.M.; Majoral, J.P.; Shi, X. Revisiting Cationic Phosphorus Dendrimers as a Nonviral Vector for Optimized Gene Delivery Toward Cancer Therapy Applications. Biomacromolecules 2020, 21, 2502–2511. [Google Scholar] [CrossRef]
- Liu, H.; Wang, Y.; Wang, M.; Xiao, J.; Cheng, Y. Fluorinated poly(propylenimine) dendrimers as gene vectors. Biomaterials 2014, 35, 5407–5413. [Google Scholar] [CrossRef] [PubMed]
- Franiak-Pietryga, I.; Ostrowska, K.; Maciejewski, H.; Appelhans, D.; Misiewicz, M.; Ziemba, B.; Bednarek, M.; Bryszewska, M.; Borowiec, M. PPI-G4 Glycodendrimers Upregulate TRAIL-Induced Apoptosis in Chronic Lymphocytic Leukemia Cells. Macromol. Biosci. 2017, 17, 1600169. [Google Scholar] [CrossRef]
- Dzmitruk, V.; Apartsin, E.; Ihnatsyeu-Kachan, A.; Abashkin, V.; Shcharbin, D.; Bryszewska, M. Dendrimers show promise for siRNA and microrna therapeutics. Pharmaceutics 2018, 10, 126. [Google Scholar] [CrossRef] [Green Version]
- Dias, A.P.; da Silva Santos, S.; da Silva, J.V.; Parise-Filho, R.; Ferreira, E.I.; El Seoud, O.; Giarolla, J. Dendrimers in the context of nanomedicine. Int. J. Pharm. 2020, 573, 118814. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Kang, C.S.; Yuan, X.B.; Zhou, X.; Xu, P.; Han, L.; Wang, G.X.; Jia, Z.; Zhong, Y.; Yu, S.; et al. Co-delivery of as-miR-21 and 5-FU by poly(amidoamine) dendrimer attenuates human glioma cell growth in vitro. J. Biomater. Sci. Polym. Ed. 2010, 21, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.; Ren, Y.; Shi, Z.; Long, L.; Pu, P.; Sheng, J.; Yuan, X.; Kang, C. Sequence-Dependent Synergistic Inhibition of Human Glioma Cell Lines by Combined Temozolomide and miR-21 Inhibitor Gene Therapy. Mol. Pharm. 2012, 9, 2636–2645. [Google Scholar] [CrossRef]
- Candela-Noguera, V.; Vivo-Llorca, G.; de Greñu, B.D.; Alfonso, M.; Aznar, E.; Orzáez, M.; Marcos, M.D.; Sancenón, F.; Martínez-Máñez, R. Gene-Directed Enzyme Prodrug Therapy by Dendrimer-Like Mesoporous Silica Nanoparticles against Tumor Cells. Nanomaterials 2021, 11, 1298. [Google Scholar] [CrossRef]
- Huang, Y.H.; Peng, W.; Furuuchi, N.; Gerhart, J.; Rhodes, K.; Mukherjee, N.; Jimbo, M.; Gonye, G.E.; Brody, J.R.; Getts, R.C.; et al. Delivery of Therapeutics Targeting the mRNA-Binding Protein HuR Using 3DNA Nanocarriers Suppresses Ovarian Tumor Growth. Cancer Res. 2016, 76, 1549–1559. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.F.; Zhang, H.T.; Xin, L. Hyaluronic acid-modified polyamidoamine dendrimer G5-entrapped gold nanoparticles delivering METase gene inhibits gastric tumor growth via targeting CD44+ gastric cancer cells. J. Cancer Res. Clin. Oncol. 2018, 144, 1463–1473. [Google Scholar] [CrossRef]
- Chen, K.; Xin, X.; Qiu, L.; Li, W.; Guan, G.; Li, G.; Qiao, M.; Zhao, X.; Hu, H.; Chen, D. Co-delivery of p53 and MDM2 inhibitor RG7388 using a hydroxyl terminal PAMAM dendrimer derivative for synergistic cancer therapy. Acta Biomater. 2019, 100, 118–131. [Google Scholar] [CrossRef]
- Bai, C.Z.; Choi, S.; Nam, K.; An, S.; Park, J.S. Arginine modified PAMAM dendrimer for interferon beta gene delivery to malignant glioma. Int. J. Pharm. 2013, 445, 79–87. [Google Scholar] [CrossRef]
- Wang, G.; Hu, W.; Chen, H.; Shou, X.; Ye, T.; Xu, Y. Cocktail Strategy Based on NK Cell-Derived Exosomes and Their Biomimetic Nanoparticles for Dual Tumor Therapy. Cancers 2019, 11, 1560. [Google Scholar] [CrossRef] [Green Version]
- Zhupanyn, P.; Ewe, A.; Büch, T.; Malek, A.; Rademacher, P.; Müller, C.; Reinert, A.; Jaimes, Y.; Aigner, A. Extracellular vesicle (ECV)-modified polyethylenimine (PEI) complexes for enhanced siRNA delivery in vitro and in vivo. J. Control. Release 2020, 319, 63–76. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, M.; Drzewinska, J.; Dzmitruk, V.; Shcharbin, D.; Klajnert, B.; Appelhans, D.; Bryszewska, M. Stability of Dendriplexes Formed by Anti-HIV Genetic Material and Poly(propylene imine) Dendrimers in the Presence of Glucosaminoglycans. J. Phys. Chem. B 2012, 116, 14525–14532. [Google Scholar] [CrossRef]
- Monteagudo, S.; Pérez-Martínez, F.C.; Pérez-Carrión, M.D.; Guerra, J.; Merino, S.; Sánchez-Verdú, M.P.; Ceña, V. Inhibition of p42 MAPK using a nonviral vector-delivered siRNA potentiates the anti-tumor effect of metformin in prostate cancer cells. Nanomedicine 2012, 7, 493–506. [Google Scholar] [CrossRef]
- Liu, C.; Shao, N.; Wang, Y.; Cheng, Y. Clustering Small Dendrimers into Nanoaggregates for Efficient DNA and siRNA Delivery with Minimal Toxicity. Adv. Healthc. Mater. 2016, 5, 584–592. [Google Scholar] [CrossRef]
- Ihnatsyeu-Kachan, A.; Dzmitruk, V.; Apartsin, E.; Krasheninina, O.; Ionov, M.; Loznikova, S.; Venyaminova, A.; Miłowska, K.; Shcharbin, D.; Mignani, S.; et al. Multi-Target Inhibition of Cancer Cell Growth by SiRNA Cocktails and 5-Fluorouracil Using Effective Piperidine-Terminated Phosphorus Dendrimers. Colloids Interfaces 2017, 1, 6. [Google Scholar] [CrossRef] [Green Version]
- Taratula, O.; Garbuzenko, O.B.; Kirkpatrick, P.; Pandya, I.; Savla, R.; Pozharov, V.P.; He, H.; Minko, T. Surface-engineered targeted PPI dendrimer for efficient intracellular and intratumoral siRNA delivery. J. Control. Release 2009, 140, 284–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janaszewska, A.; Lazniewska, J.; Trzepiński, P.; Marcinkowska, M.; Klajnert-Maculewicz, B. Cytotoxicity of dendrimers. Biomolecules 2019, 9, 330. [Google Scholar] [CrossRef] [Green Version]
- Albertazzi, L.; Gherardini, L.; Brondi, M.; Sato, S.S.; Bifone, A.; Pizzorusso, T.; Ratto, G.M.; Bardi, G. In Vivo Distribution and Toxicity of PAMAM Dendrimers in the Central Nervous System Depend on Their Surface Chemistry. Mol. Pharm. 2012, 10, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Ziemba, B.; Janaszewska, A.; Ciepluch, K.; Krotewicz, M.; Fogel, W.A.; Appelhans, D.; Voit, B.; Bryszewska, M.; Klajnert, B. In vivo toxicity of poly(propyleneimine) dendrimers. J. Biomed. Mater. Res. Part A 2011, 99, 261–268. [Google Scholar] [CrossRef]
- Bullen, H.A.; Hemmer, R.; Haskamp, A.; Cason, C.; Wall, S.; Spaulding, R.; Rossow, B.; Hester, M.; Caroway, M.; Haik, K.L.; et al. Evaluation of Biotinylated PAMAM Dendrimer Toxicity in Models of the Blood Brain Barrier: A Biophysical and Cellular Approach. J. Biomater. Nanobiotechnol. 2011, 2, 485–493. [Google Scholar] [CrossRef] [Green Version]
- Gothwal, A.; Malik, S.; Gupta, U.; Jain, N.K. Toxicity and biocompatibility aspects of dendrimers. In Pharmaceutical Applications of Dendrimers; Elsevier: Amsterdam, The Netherlands, 2020; pp. 251–274. [Google Scholar] [CrossRef]
- Thiagarajan, G.; Greish, K.; Ghandehari, H. Charge affects the oral toxicity of poly(amidoamine) dendrimers. Eur. J. Pharm. Biopharm. 2013, 84, 330–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agashe, H.B.; Dutta, T.; Garg, M.; Jain, N.K. Investigations on the toxicological profile of functionalized fifth-generation poly(propylene imine) dendrimer. J. Pharm. Pharm. 2010, 58, 1491–1498. [Google Scholar] [CrossRef]
- Kesharwani, P.; Gajbhiye, V.; Tekade, R.K.; Jain, N.K. Evaluation of dendrimer safety and efficacy through cell line studies. Curr. Drug Targets 2011, 12, 1478–1497. [Google Scholar] [CrossRef] [PubMed]
- Yellepeddi, V.K.; Kumar, A.; Palakurthi, S. Surface modified poly(amido)amine dendrimers as diverse nanomolecules for biomedical applications. Expert Opin. Drug Deliv. 2009, 6, 835–850. [Google Scholar] [CrossRef]
- Jain, K.; Kesharwani, P.; Gupta, U.; Jain, N.K. Dendrimer toxicity: Let’s meet the challenge. Int. J. Pharm. 2010, 394, 122–142. [Google Scholar] [CrossRef]
- Duncan, R.; Izzo, L. Dendrimer biocompatibility and toxicity. Adv. Drug Deliv. Rev. 2005, 57, 2215–2237. [Google Scholar] [CrossRef]
- Treatment of Non-Responding to Conventional Therapy Inoperable Liver Cancers by In Situ Introduction of ImDendrim—Full Text View—ClinicalTrials.gov. Available online: https://clinicaltrials.gov/ct2/show/NCT03255343 (accessed on 19 September 2021).
- DEP® Drug Delivery—Starpharma. Available online: https://starpharma.com/drug_delivery (accessed on 19 September 2021).
- Ashrafizadeh, M.; Hushmandi, K.; Moghadam, E.R.; Zarrin, V.; Kashani, S.H.; Bokaie, S.; Najafi, M.; Tavakol, S.; Mohammadinejad, R.; Nabavi, N.; et al. Progress in delivery of siRNA-based therapeutics employing nano-vehicles for treatment of prostate cancer. Bioengineering 2020, 7, 91. [Google Scholar] [CrossRef] [PubMed]
- Starpharma to Commence DEP® Cabazitaxel Phase 1/2 Trial—Starpharma. Available online: https://starpharma.com/news/356 (accessed on 19 September 2021).
- Starpharma Commences Phase 1/2 DEP® Irinotecan Trial—Starpharma. Available online: https://starpharma.com/news/427 (accessed on 19 September 2021).
- Abedi-Gaballu, F.; Dehghan, G.; Ghaffari, M.; Yekta, R.; Abbaspour-Ravasjani, S.; Baradaran, B.; Ezzati Nazhad Dolatabadi, J.; Hamblin, M.R. PAMAM dendrimers as efficient drug and gene delivery nanosystems for cancer therapy. Appl. Mater. Today 2018, 12, 177–190. [Google Scholar] [CrossRef]
- Dzmitruk, V.; Szulc, A.; Shcharbin, D.; Janaszewska, A.; Shcharbina, N.; Lazniewska, J.; Novopashina, D.; Buyanova, M.; Ionov, M.; Klajnert-Maculewicz, B.; et al. Anticancer siRNA cocktails as a novel tool to treat cancer cells. Part (B). Efficiency of pharmacological action. Int. J. Pharm. 2015, 485, 288–294. [Google Scholar] [CrossRef]
- Contin, M.; Garcia, C.; Dobrecky, C.; Lucangioli, S.; D’Accorso, N. Advances in drug delivery, gene delivery and therapeutic agents based on dendritic materials. Future Med. Chem. 2019, 11, 1791–1810. [Google Scholar] [CrossRef]
- Shen, W.; Liu, H.; Ling-Hu, Y.; Wang, H.; Cheng, Y. Enhanced siRNA delivery of a cyclododecylated dendrimer compared to its linear derivative. J. Mater. Chem. B 2016, 4, 5654–5658. [Google Scholar] [CrossRef]
- Chang, H.; Zhang, Y.; Li, L.; Cheng, Y. Efficient delivery of small interfering RNA into cancer cells using dodecylated dendrimers. J. Mater. Chem. B 2015, 3, 8197–8202. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.; Wen, S.; Guo, R.; Wang, S.; Shi, X. Partially acetylated dendrimer-entrapped gold nanoparticles with reduced cytotoxicity for gene delivery applications. J. Nanosci. Nanotechnol. 2015, 15, 4094–4105. [Google Scholar] [CrossRef]
- Sadekar, S.; Ghandehari, H. Transepithelial transport and toxicity of PAMAM dendrimers: Implications for oral drug delivery. Adv. Drug Deliv. Rev. 2012, 64, 571–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Hu, K.; Cheng, Y. Tailoring the dendrimer core for efficient gene delivery. Acta Biomater. 2016, 35, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.; Wang, H.; Shao, N.; Wang, M.; Wang, X.; Cheng, Y. Surface-engineered dendrimers with a diaminododecane core achieve efficient gene transfection and low cytotoxicity. Bioconjug. Chem. 2014, 25, 342–350. [Google Scholar] [CrossRef]
- Liu, C.; Liu, X.; Rocchi, P.; Qu, F.; Iovanna, J.L.; Peng, L. Arginine-terminated generation 4 PAMAM dendrimer as an effective nanovector for functional siRNA delivery in vitro and in vivo. Bioconjug. Chem. 2014, 25, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cheng, Y. The effect of fluorination on the transfection efficacy of surface-engineered dendrimers. Biomaterials 2014, 35, 6603–6613. [Google Scholar] [CrossRef]
- Wang, M.; Liu, H.; Li, L.; Cheng, Y. A fluorinated dendrimer achieves excellent gene transfection efficacy at extremely low nitrogen to phosphorus ratios. Nat. Commun. 2014, 5, 3053. [Google Scholar] [CrossRef] [Green Version]
- Klein, E.; Ciobanu, M.; Klein, J.; MacHi, V.; Leborgne, C.; Vandamme, T.; Frisch, B.; Pons, F.; Kichler, A.; Zuber, G.; et al. “HFP” fluorinated cationic lipids for enhanced lipoplex stability and gene delivery. Bioconjug. Chem. 2010, 21, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cheng, Y. Structure-activity relationships of fluorinated dendrimers in DNA and siRNA delivery. Acta Biomater. 2016, 46, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Müller, K.; Faeh, C.; Diederich, F. Fluorine in pharmaceuticals: Looking beyond intuition. Science 2007, 317, 1881–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, Y.; Wang, Y.; Hu, J.; Li, T.; Liu, H.; Zhang, Q.; Cheng, Y. Self-Assembled Fluorodendrimers Combine the Features of Lipid and Polymeric Vectors in Gene Delivery. Angew. Chem. Int. Ed. 2015, 54, 11647–11651. [Google Scholar] [CrossRef]
- Kannan, R.M.; Nance, E.; Kannan, S.; Tomalia, D.A. Emerging concepts in dendrimer-based nanomedicine: From design principles to clinical applications. J. Intern. Med. 2014, 276, 579–617. [Google Scholar] [CrossRef] [PubMed]
- Arima, H.; Arizono, M.; Higashi, T.; Yoshimatsu, A.; Ikeda, H.; Motoyama, K.; Hattori, K.; Takeuchi, T.; Hirayama, F.; Uekama, K. Potential use of folate-polyethylene glycol (PEG)-appended dendrimer (G3) conjugate with α-cyclodextrin as DNA carriers to tumor cells. Cancer Gene Ther. 2012, 19, 358–366. [Google Scholar] [CrossRef] [Green Version]
- Arima, H.; Yoshimatsu, A.; Ikeda, H.; Ohyama, A.; Motoyama, K.; Higashi, T.; Tsuchiya, A.; Niidome, T.; Katayama, Y.; Hattori, K.; et al. Folate-PEG-appended dendrimer conjugate with α-cyclodextrin as a novel cancer cell-selective siRNA delivery carrier. Mol. Pharm. 2012, 9, 2591–2604. [Google Scholar] [CrossRef] [PubMed]
- Li, J.L.; Tang, B.; Yuan, B.; Sun, L.; Wang, X.G. A review of optical imaging and therapy using nanosized graphene and graphene oxide. Biomaterials 2013, 34, 9519–9534. [Google Scholar] [CrossRef]
- Xue, L.; Yan, Y.; Kos, P.; Chen, X.; Siegwart, D.J. PEI fluorination reduces toxicity and promotes liver-targeted siRNA delivery. Drug Deliv. Transl. Res. 2021, 11, 255–260. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, J.; Chen, C. Gold Nanorods Based Platforms for Light-Mediated Theranostics. Theranostics 2013, 3, 223. [Google Scholar] [CrossRef] [PubMed]
- Wong, B.S.; Yoong, S.L.; Jagusiak, A.; Panczyk, T.; Ho, H.K.; Ang, W.H.; Pastorin, G. Carbon nanotubes for delivery of small molecule drugs. Adv. Drug Deliv. Rev. 2013, 65, 1964–2015. [Google Scholar] [CrossRef]
- Taratula, O.; Garbuzenko, O.; Savla, R.; Andrew Wang, Y.; He, H.; Minko, T. Multifunctional Nanomedicine Platform for Cancer Specific Delivery of siRNA by Superparamagnetic Iron Oxide Nanoparticles-Dendrimer Complexes. Curr. Drug Deliv. 2010, 8, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-Fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Soršak, E.; Valh, J.V.; Urek, S.K.; Lobnik, A. Application of PAMAM dendrimers in optical sensing. Analyst 2015, 140, 976–989. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polymer | Structure | Efficiency |
|---|---|---|
| PEI = Polyethylenimine or polyaziridine. Polymer with an amine group repeating unit and two aliphatic groups as spacer (CH2-CH2). They contain secondary amine groups. |  | PEI is known as one of the most efficient nonviral agent currently used in GDEPT strategy [104]. The efficiency of transfection using PEI is largely due to the intense proton buffering effect and the "proton sponge effect" in the endosomal stage. At physiological pH, about 15–20% of PEI amines are protonated, which gives PEI the very intense capacity of a proton sponge [105]. |
| Branched PEI Ethylenediamine-ethylenimine copolymer; Aziridine-1,2-diaminoethane copolymer; Ethylenediamine - ethylenimine polymer |  | Every third atom in the PEI structure is a nitrogen atom with a very large distribution of primary, secondary and tertiary amine groups [104]. The superiority of PEI over other polymers such as PLL (lysine polypeptide) is due to the high loading density and the flexibility of the chain. PEI electrostatically condenses high molecular weight DNA in the form of polypeptic nanoparticles (10–100 nm), which are able to achieve endocytosis [106]. |
| PPI – G4 poly(propyleneimine) G4 dendrimers with fully branched amine groups |  | Changes in the terminal groups of the PPI dendrimer lead to a decreased toxicity, regardless of its internal structure. These structural modulations contribute to the aqueous solubility and increased stability of hydrolysis. Changes in the internal structure are also favorable by quaternization of internal tertiary amines, with the formation of cationic ammonium groups [107]. High-molecular-weight (HMW) modulations have led to a decreased cytotoxicity, while maintaining the potential for DNA binding and condensation [108]. |
| (a) Linear poly(amidoamine) dendrimers (PAMAM); (b) Branched PAMAM dendrimers: spheroidal, cascade polymers, the size and surface charge of which determine the generation number; ethylenediamine (EDA), ammonia (NH3) or cystamine are used as initiating nuclei, obtaining numerous ramifications. |  | They contain a multitude of secondary and tertiary amines, which cause an intense "proton sponge" effect. PAMAM forms spherical polymers with good water solubility due to the presence of surface amine groups [109,110,111,112]. |
| Linear PLL |  | At physiological pH, amino groups are positively charged, so they have a reduced proton-buffering capacity to facilitate endosomal escape. In general, PLL has a relatively weak gene transfection activity in the absence of a lysosomal disruptive agent. In addition, unmodified in vitro PLL has increased cytotoxicity [113,114]. |
| Branched PLL |  | They have a large surface and branched structure, which, together with the numerous functional groups, facilitates the binding of different biological entities or molecules [115,116]. |
| PDMAEMA poly(2-(N,N-dimethylamino)ethyl methacrylate) |  | The synthesis of PDMAEMA derivatives with a molecular weight of approximately 60 kDa by controlled radical polymerization is optimal for the efficacy and toxicity of controlled transfection in GDEPT [117,118]. |
| Chitosan |  | Chitosan, a natural polymer derived from chitin [119], is used as a targeting and release vector in GDEPT therapy due to its unique characteristics (cationic structure, biocompatibility, relatively low production cost and the possibility of easy functional modulation), at an optimal molecular mass between 65 and 170 kDa [120,121]. |
| Cyclodextrin (CD) |  | CDs generally have a limited transfection efficiency, which can be increased by various modulations/structural derivatizations: polycationic amphiphilic CDs (paCD), modification of functional groups, hydrophilic–hydrophobic balance and linker length. Another favourable modulation is represented by the modification of β-CD with a pyridylamino, alkylimidazole, methoxyethylamino or primary amine group in position 6 of the glucose units [122,123]. |
| Advantages. | Disadvantages |
|---|---|
| Chemical: controllable synthesis and degradation, homogenous structure, good solubility or miscibility, numerous functional groups | Chemical: low solubility of higher generation dendrimers |
| Pharmaceutical: lower viscosity, multiple possibilities to bond the active substances, high bioavailability | Pharmaceutical: poor quality control, difficult technological transfer from research to clinical practice, tendency to aggregate in aqueous solutions |
| Pharmacological: high capacity to penetrate membranes or barriers, a good ratio between their therapeutic efficacy and toxicity, increased bioavailability and efficiency, prolonged half-life, specific activity | Pharmacological: superficial penetration into tissues of dendrimers with higher tendency to aggregate in aqueous solutions |
| Economic disadvantages: higher cost of production |
| Radicals | Permeability | Cytotoxicity | Cellular Uptake |
|---|---|---|---|
| Acetyl | increase | decrease | |
| Lauroyl | increase | decrease | Increase |
| Amino acid | increase | increase | |
| PEG | decrease | decrease | Decrease |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chis, A.A.; Dobrea, C.M.; Rus, L.-L.; Frum, A.; Morgovan, C.; Butuca, A.; Totan, M.; Juncan, A.M.; Gligor, F.G.; Arseniu, A.M. Dendrimers as Non-Viral Vectors in Gene-Directed Enzyme Prodrug Therapy. Molecules 2021, 26, 5976. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195976
Chis AA, Dobrea CM, Rus L-L, Frum A, Morgovan C, Butuca A, Totan M, Juncan AM, Gligor FG, Arseniu AM. Dendrimers as Non-Viral Vectors in Gene-Directed Enzyme Prodrug Therapy. Molecules. 2021; 26(19):5976. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195976
Chicago/Turabian StyleChis, Adriana Aurelia, Carmen Maximiliana Dobrea, Luca-Liviu Rus, Adina Frum, Claudiu Morgovan, Anca Butuca, Maria Totan, Anca Maria Juncan, Felicia Gabriela Gligor, and Anca Maria Arseniu. 2021. "Dendrimers as Non-Viral Vectors in Gene-Directed Enzyme Prodrug Therapy" Molecules 26, no. 19: 5976. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26195976