
Micellar Casein and Whey Powder Hold a TGF-β Activity and Regulate ID Genes In Vitro

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
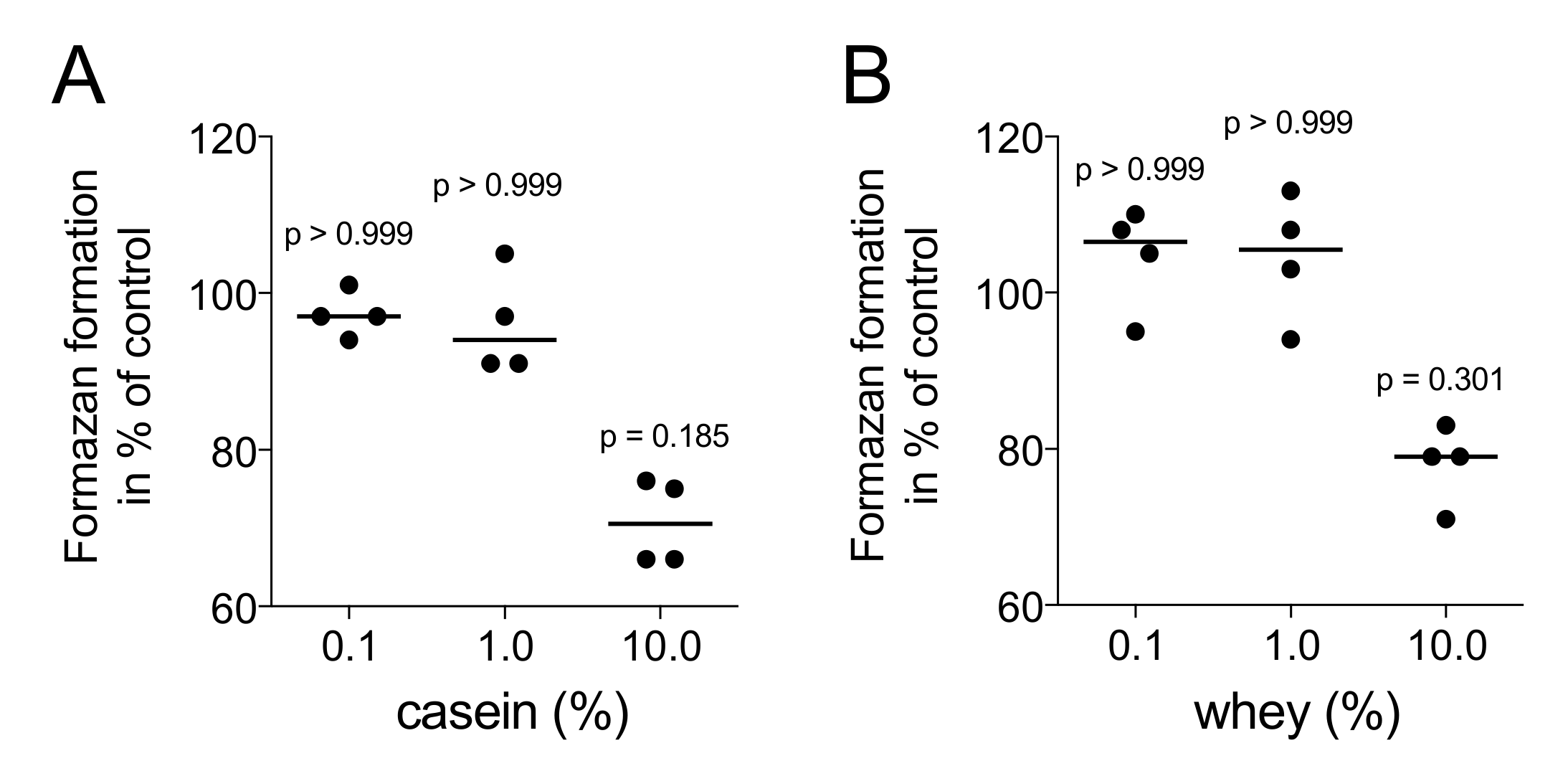
2.1. Casein and Whey Powders Do Not Affect Viability and Proliferation of Gingival Fibroblasts
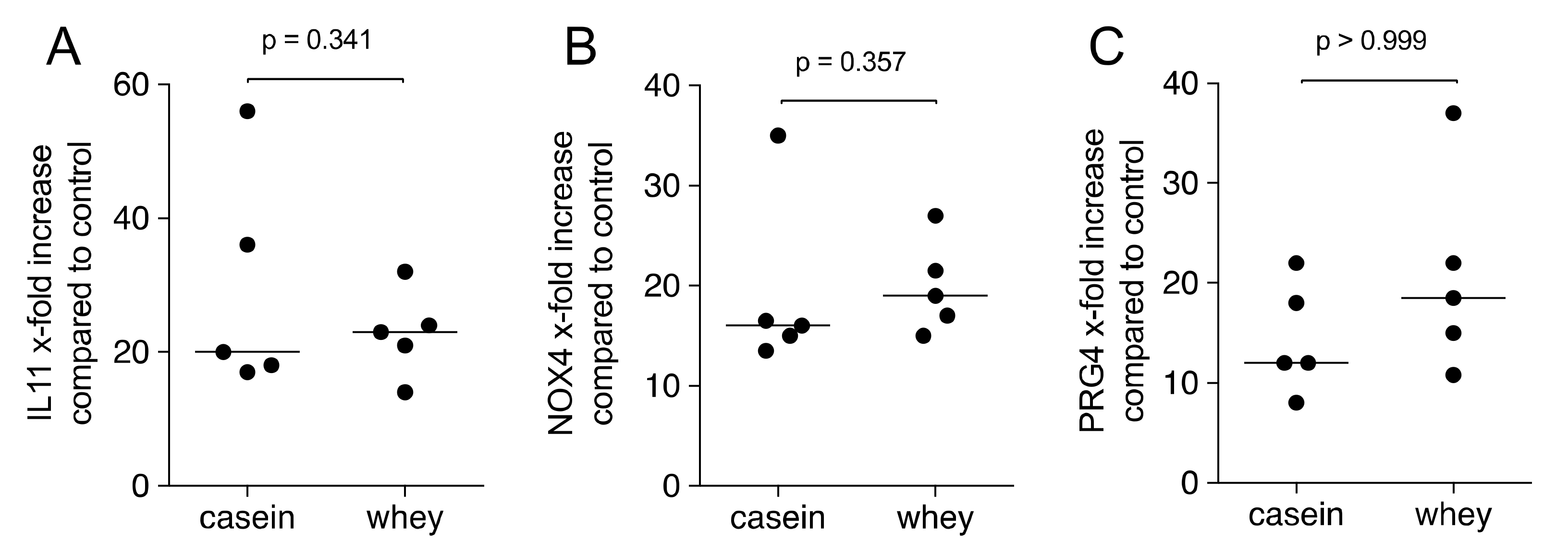
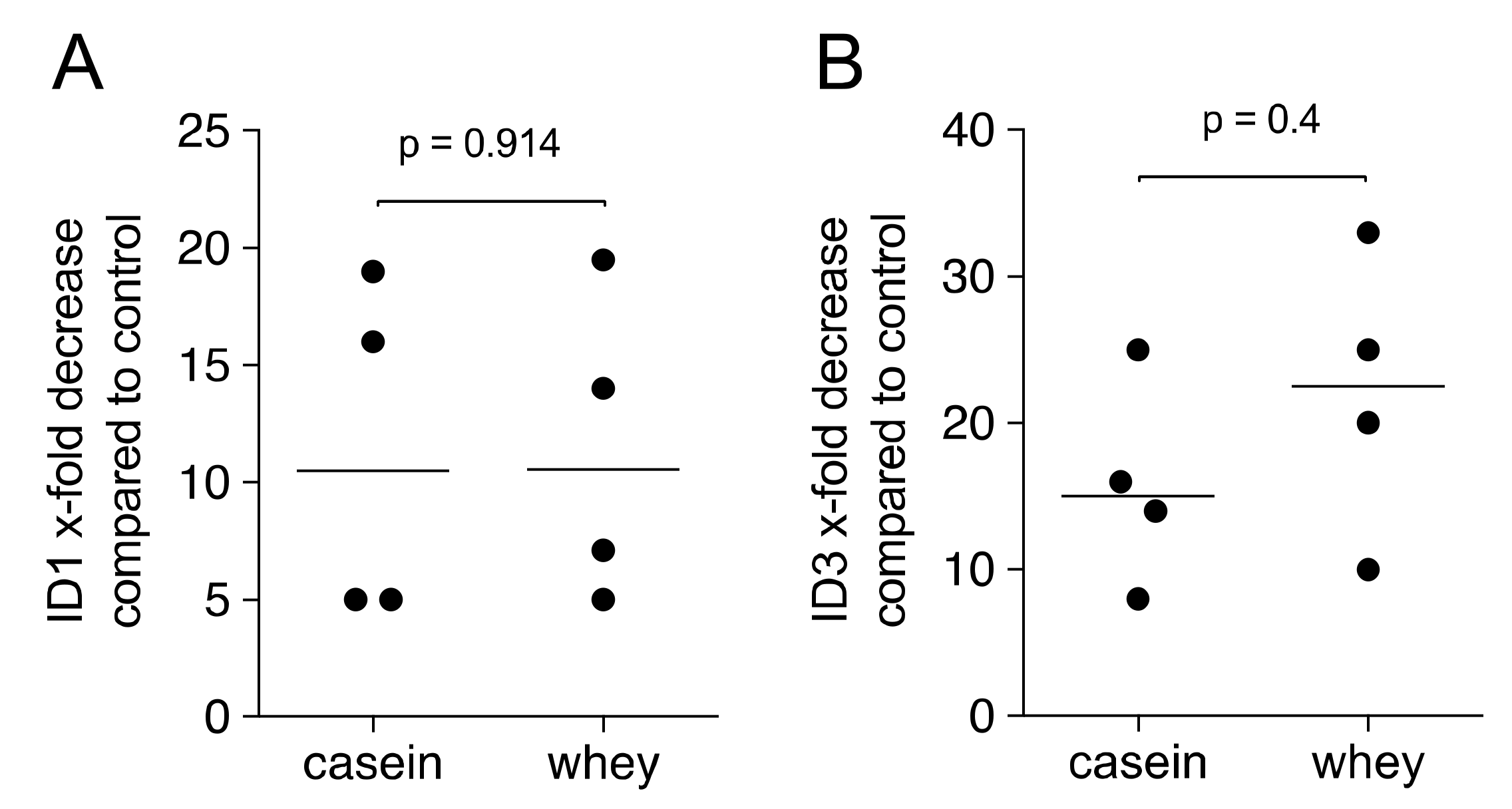
2.2. Casein and Whey Powder Stimulate TGF-β Target Genes Expression in Gingival Fibroblasts
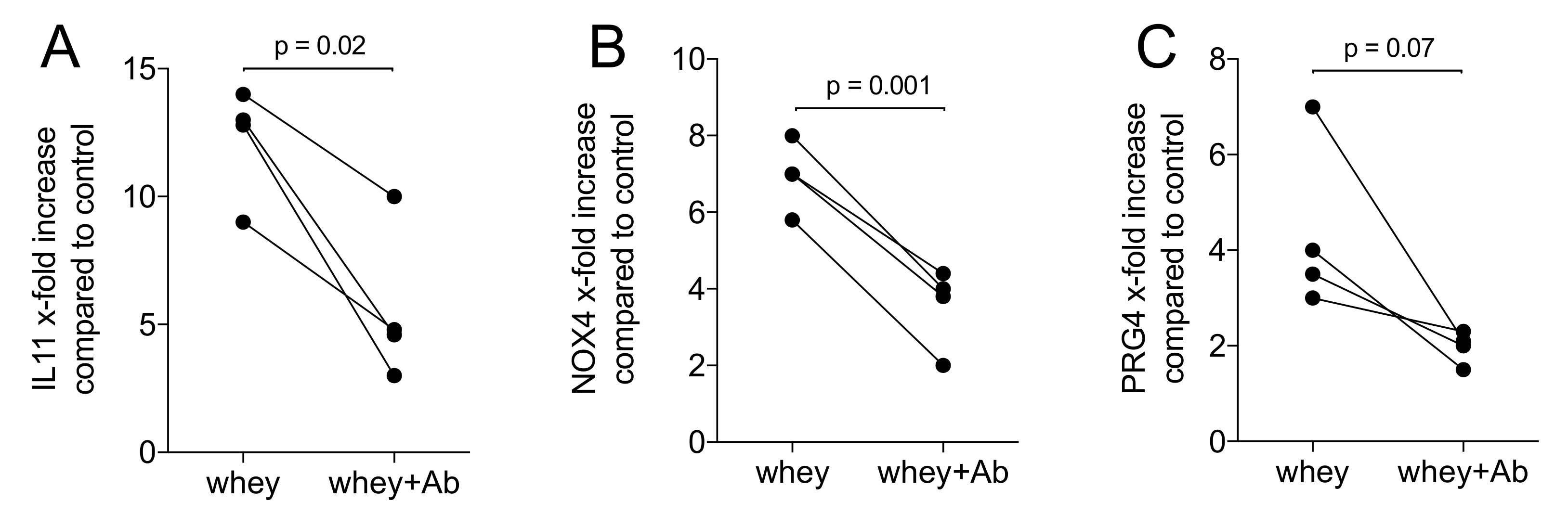
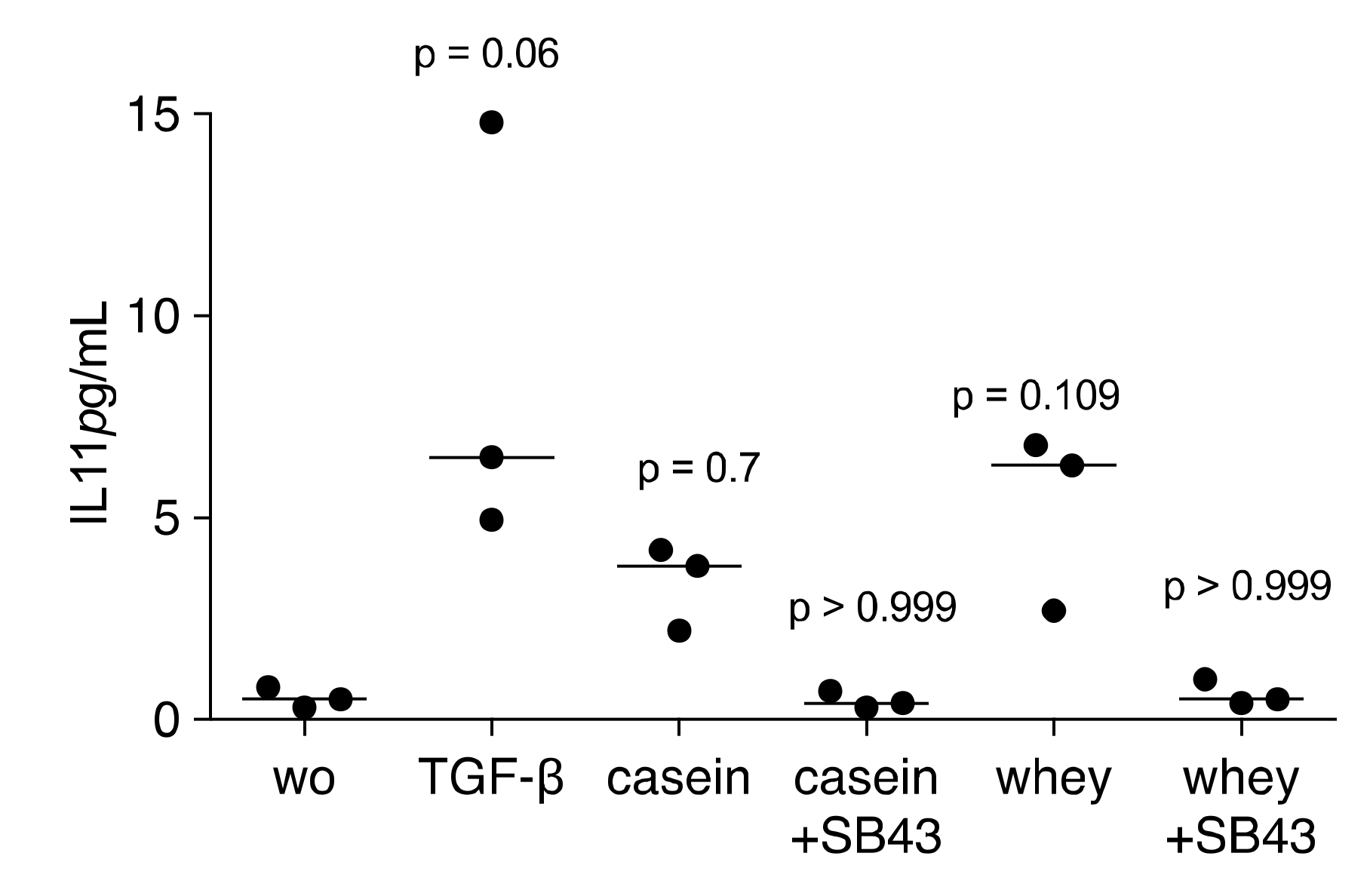
2.3. A TGF-β Neutralizing Antibody Reduce the Whey-Stimulated Gene Expression
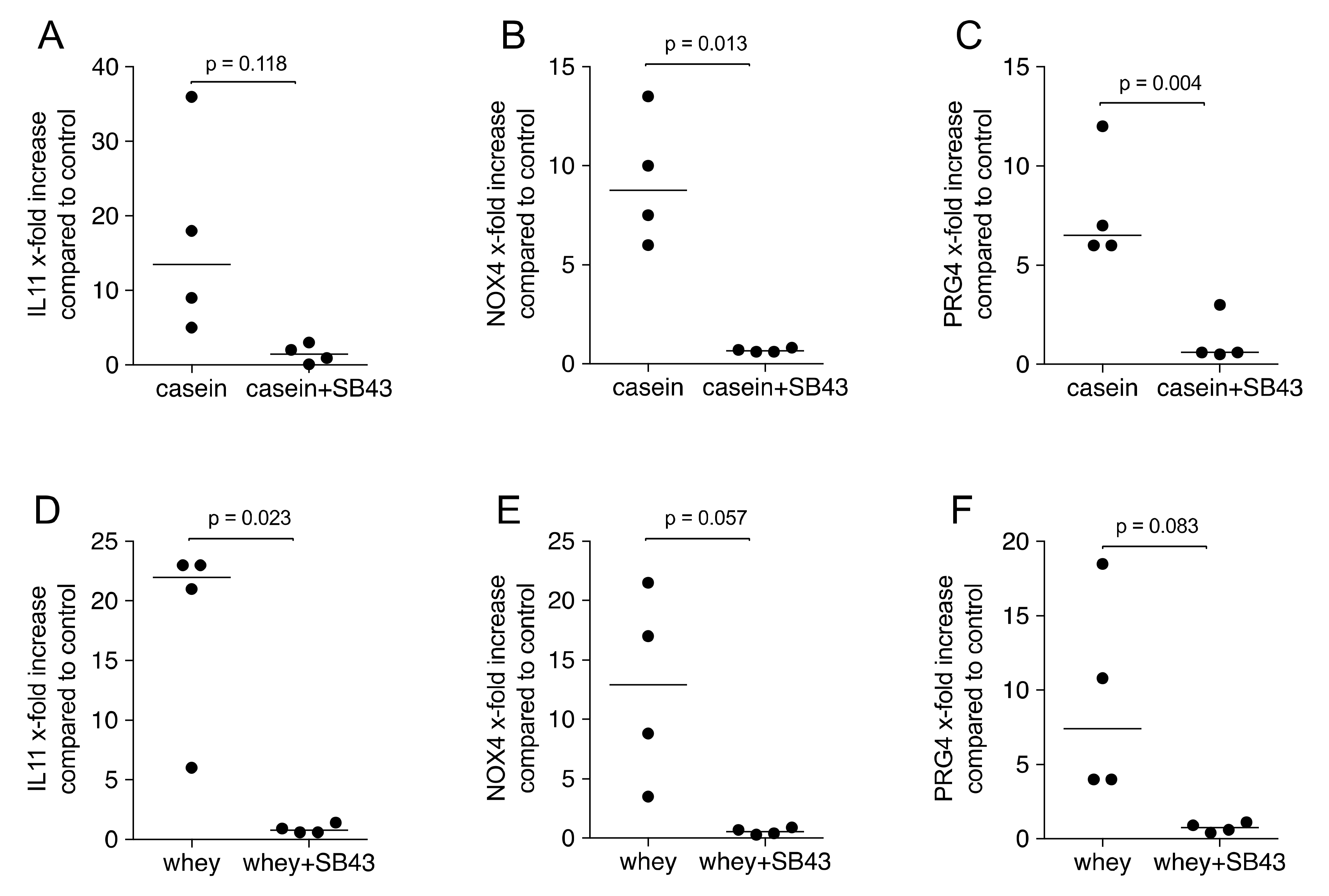
2.4. Gene Expression Is Suppressed by TGF-β Receptor I Kinase Inhibitor SB431542
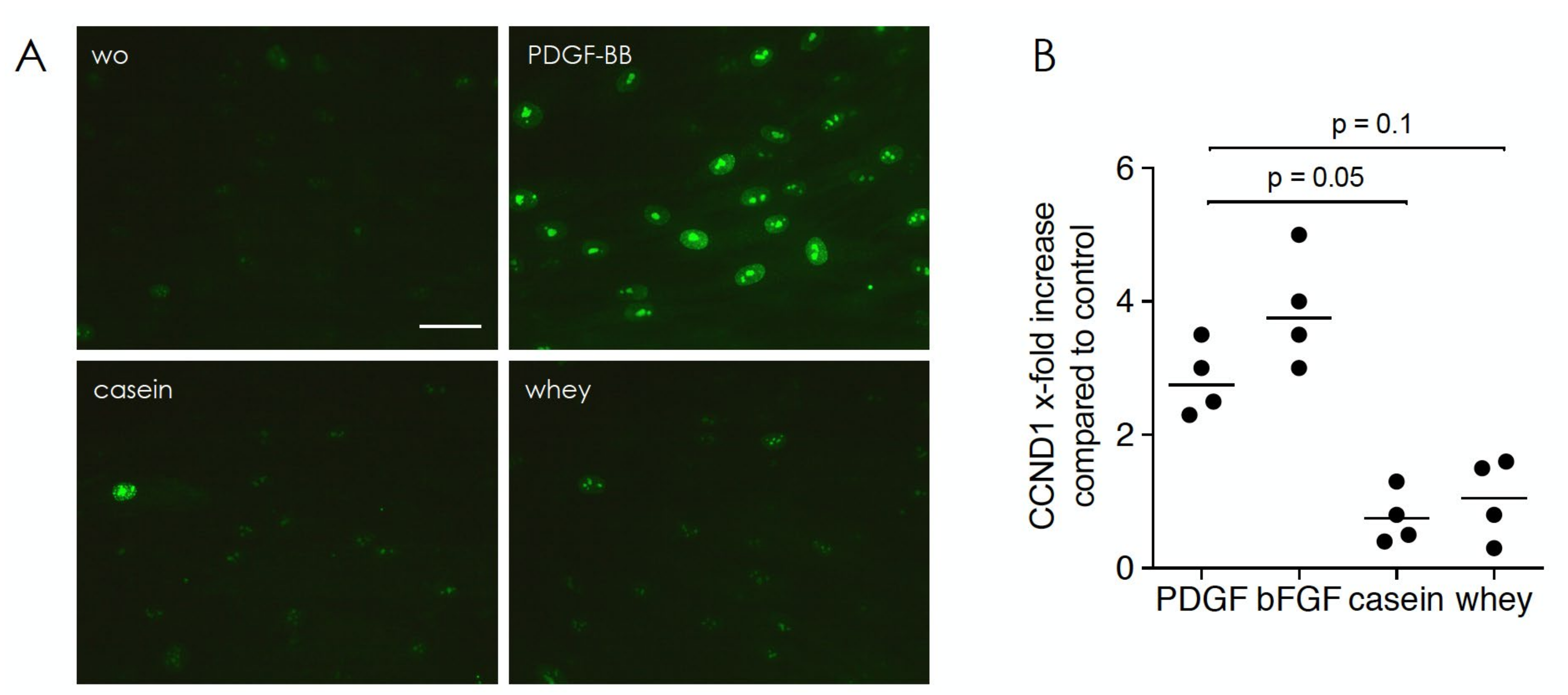
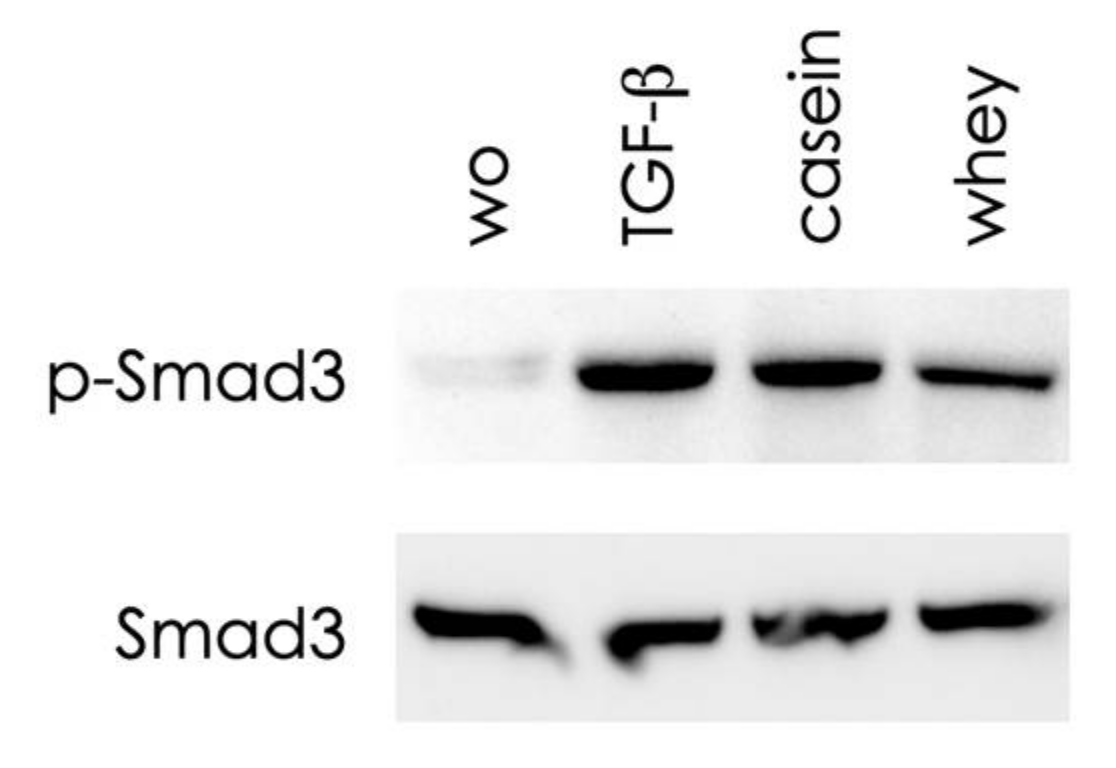
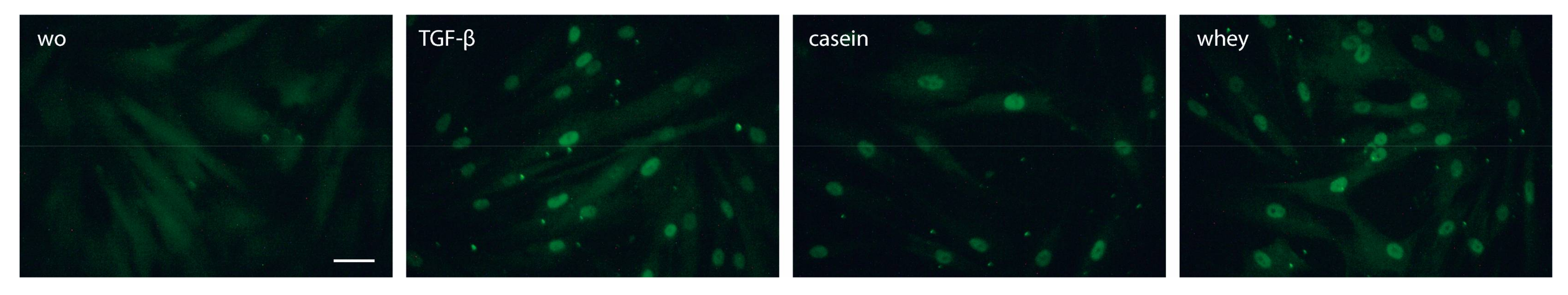
2.5. Casein and Whey Stimulate Phosphorylation and Nuclear Translocation of Smad Proteins
2.6. Gene Expression Analysis of HSC2 Exposed to Casein and Whey Powder
3. Material and Methods
3.1. Aqueous Fractions of Casein and Whey Powder
3.2. Primary Gingival Fibroblasts and Oral Squamous Cells
3.3. Viability Assay
3.4. qRT-PCR Analysis and Immunoassay
3.5. Western Blot Analysis
3.6. Immunofluorescence
3.7. Statistical Analysis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ogra, P.L.; Walker, W.A. Immunology of Human Milk and Lactation: Historical Overview. In Milk, Mucosal Immunity and the Microbiome: Impact on the Neonate; Lönnerdal, B., Ed.; Nestlé Nutr Inst Workshop Ser; Karger: Basel, Switzerland, 2020; Volume 94, pp. 11–26. [Google Scholar] [CrossRef]
- Sitarik, A.R.; Bobbitt, K.R.; Havstad, S.L.; Fujimura, K.E.; Levin, A.M.; Zoratti, E.M.; Kim, H.; Woodcroft, K.J.; Wegienka, G.; Ownby, D.R.; et al. Breast Milk Transforming Growth Factor β Is Associated with Neonatal Gut Microbial Composition. J. Pediatr. Gastroenterol. Nutr. 2017, 65, e60–e67. [Google Scholar] [CrossRef] [PubMed]
- Holvoet, S.; Perrot, M.; de Groot, N.; Prioult, G.; Mikogami, T.; Verhasselt, V.; Nutten, S. Oral Tolerance Induction to Newly Introduced Allergen is Favored by a Transforming Growth Factor-beta-Enriched Formula. Nutrients 2019, 11, 2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozawa, T.; Miyata, M.; Nishimura, M.; Ando, T.; Ouyang, Y.; Ohba, T.; Shimokawa, N.; Ohnuma, Y.; Katoh, R.; Ogawa, H.; et al. Transforming growth factor-beta activity in commercially available pasteurized cow milk provides protection against inflammation in mice. J. Nutr. 2009, 139, 69–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahli, A.; Bosshardt, D.; Sculean, A.; Gruber, R. Emdogain-regulated gene expression in palatal fibroblasts requires TGF-betaRI kinase signaling. PLoS ONE 2014, 9, e105672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, M.; Caballe-Serrano, J.; Bosshardt, D.D.; Ankersmit, H.J.; Buser, D.; Gruber, R. Bone-Conditioned Medium Changes Gene Expression in Bone-Derived Fibroblasts. Int. J. Oral Maxillofac. Implants 2015, 30, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Strauss, F.J.; Stahli, A.; Beer, L.; Mitulovic, G.; Gilmozzi, V.; Haspel, N.; Schwab, G.; Gruber, R. Acid bone lysate activates TGFbeta signalling in human oral fibroblasts. Sci. Rep. 2018, 8, 16065. [Google Scholar] [CrossRef]
- Schafer, S.; Viswanathan, S.; Widjaja, A.A.; Lim, W.W.; Moreno-Moral, A.; DeLaughter, D.M.; Ng, B.; Patone, G.; Chow, K.; Khin, E.; et al. IL-11 is a crucial determinant of cardiovascular fibrosis. Nature 2017, 552, 110–115. [Google Scholar] [CrossRef]
- Widjaja, A.A.; Singh, B.K.; Adami, E.; Viswanathan, S.; Dong, J.; D’Agostino, G.A.; Ng, B.; Lim, W.W.; Tan, J.; Paleja, B.S.; et al. Inhibiting Interleukin 11 Signaling Reduces Hepatocyte Death and Liver Fibrosis, Inflammation, and Steatosis in Mouse Models of Nonalcoholic Steatohepatitis. Gastroenterology 2019, 157, 777–792.e14. [Google Scholar] [CrossRef] [Green Version]
- Jeong, B.Y.; Park, S.R.; Cho, S.; Yu, S.L.; Lee, H.Y.; Park, C.G.; Kang, J.; Jung, D.Y.; Park, M.H.; Hwang, W.M.; et al. TGF-beta-mediated NADPH oxidase 4-dependent oxidative stress promotes colistin-induced acute kidney injury. J. Antimicrob. Chemother. 2018, 73, 962–972. [Google Scholar] [CrossRef]
- Panahipour, L.; Stahli, A.; Haiden, N.; Gruber, R. TGF-beta activity in cow milk and fermented milk products: An in vitro bioassay with oral fibroblasts. Arch. Oral. Biol. 2018, 95, 15–21. [Google Scholar] [CrossRef]
- Panahipour, L.; Tabatabaei, A.A.; Gruber, R. Hypoallergenic infant formula lacks transforming growth factor beta activity and has a lower anti-inflammatory activity than regular infant formula. J. Dairy Sci. 2020, 103, 6771–6781. [Google Scholar] [CrossRef] [PubMed]
- Magacz, M.; Kedziora, K.; Sapa, J.; Krzysciak, W. The Significance of Lactoperoxidase System in Oral Health: Application and Efficacy in Oral Hygiene Products. Int. J. Mol. Sci. 2019, 20, 1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panahipour, L.; Biasi, M.; Bokor, T.S.; Thajer, A.; Haiden, N.; Gruber, R. Milk lactoperoxidase decreases ID1 and ID3 expression in human oral squamous cell carcinoma cell lines. Sci. Rep. 2020, 10, 5836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roschger, C.; Cabrele, C. The Id-protein family in developmental and cancer-associated pathways. Cell Commun. Signal. 2017, 15, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotzer, D.; Krampert, M.; Sulyok, S.; Braun, S.; Stark, H.J.; Boukamp, P.; Werner, S. Id proteins: Novel targets of activin action, which regulate epidermal homeostasis. Oncogene 2006, 25, 2070–2081. [Google Scholar] [CrossRef] [Green Version]
- Caroli, A.M.; Savino, S.; Bulgari, O.; Monti, E. Detecting β-Casein Variation in Bovine Milk. Molecules 2016, 21, 141. [Google Scholar] [CrossRef] [Green Version]
- Abbring, S.; Hols, G.; Garssen, J.; van Esch, B. Raw cow’s milk consumption and allergic diseases–The potential role of bioactive whey proteins. Eur. J. Pharmacol. 2019, 843, 55–65. [Google Scholar] [CrossRef]
- Devries, M.C.; Phillips, S.M. Supplemental protein in support of muscle mass and health: Advantage whey. J. Food. Sci. 2015, 80, A8–A15. [Google Scholar] [CrossRef]
- Gade, J.; Beck, A.M.; Bitz, C.; Christensen, B.; Klausen, T.W.; Vinther, A.; Astrup, A. Protein-enriched, milk-based supplement to counteract sarcopenia in acutely ill geriatric patients offered resistance exercise training during and after hospitalisation: Study protocol for a randomised, double-blind, multicentre trial. BMJ Open 2018, 8, e019210. [Google Scholar] [CrossRef]
- Gryson, C.; Ratel, S.; Rance, M.; Penando, S.; Bonhomme, C.; Le Ruyet, P.; Duclos, M.; Boirie, Y.; Walrand, S. Four-month course of soluble milk proteins interacts with exercise to improve muscle strength and delay fatigue in elderly participants. J. Am. Med. Dir. Assoc. 2014, 15, 958.e1–958.e9. [Google Scholar] [CrossRef]
- Cervo, M.M.C.; Mendoza, D.S.; Barrios, E.B.; Panlasigui, L.N. Effects of Nutrient-Fortified Milk-Based Formula on the Nutritional Status and Psychomotor Skills of Preschool Children. J. Nutr. Metab. 2017, 2017, 6456738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazono, K.; Hellman, U.; Wernstedt, C.; Heldin, C.H. Latent high molecular weight complex of transforming growth factor beta 1. Purification from human platelets and structural characterization. J. Biol. Chem. 1988, 263, 6407–6415. [Google Scholar] [CrossRef]
- Barrett, N.E.; Grandison, A.S.; Lewis, M.J. Contribution of the lactoperoxidase system to the keeping quality of pasteurized milk. J. Dairy. Res. 1999, 66, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Cassiano, L.P.S.; Ventura, T.M.S.; Silva, C.M.S.; Leite, A.L.; Magalhaes, A.C.; Pessan, J.P.; Buzalaf, M.A.R. Protein Profile of the Acquired Enamel Pellicle after Rinsing with Whole Milk, Fat-Free Milk, and Water: An in vivo Study. Caries. Res. 2018, 52, 288–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kensche, A.; Durasch, A.; Konig, B.; Henle, T.; Hannig, C.; Hannig, M. Characterization of the in situ pellicle ultrastructure formed under the influence of bovine milk and milk protein isolates. Arch. Oral Biol. 2019, 104, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Cheaib, Z.; Rakmathulina, E.; Lussi, A.; Eick, S. Impact of Acquired Pellicle Modification on Adhesion of Early Colonizers. Caries. Res. 2015, 49, 626–632. [Google Scholar] [CrossRef] [Green Version]
- Barbour, M.E.; Shellis, R.P.; Parker, D.M.; Allen, G.C.; Addy, M. Inhibition of hydroxyapatite dissolution by whole casein: The effects of pH, protein concentration, calcium, and ionic strength. Eur. J. Oral. Sci. 2008, 116, 473–478. [Google Scholar] [CrossRef]
- Romero, M.J.; Nakashima, S.; Nikaido, T.; Ichinose, S.; Sadr, A.; Tagami, J. Inhibition of hydroxyapatite growth by casein, a potential salivary phosphoprotein homologue. Eur. J. Oral. Sci. 2015, 123, 288–296. [Google Scholar] [CrossRef]
- Ghatak, S.; Hascall, V.C.; Markwald, R.R.; Feghali-Bostwick, C.; Artlett, C.M.; Gooz, M.; Bogatkevich, G.S.; Atanelishvili, I.; Silver, R.M.; Wood, J.; et al. Transforming growth factor beta1 (TGFβ1)-induced CD44V6-NOX4 signaling in pathogenesis of idiopathic pulmonary fibrosis. J. Biol. Chem. 2017, 292, 10490–10519. [Google Scholar] [CrossRef] [Green Version]
- Chavez, R.D.; Sohn, P.; Serra, R. Prg4 prevents osteoarthritis induced by dominant-negative interference of TGF-ss signaling in mice. PLoS ONE 2019, 14, e0210601. [Google Scholar] [CrossRef]
- Oates, T.W.; Rouse, C.A.; Cochran, D.L. Mitogenic effects of growth factors on human periodontal ligament cells in vitro. J. Periodontol. 1993, 64, 142–148. [Google Scholar] [CrossRef]
- Kimura, Y.; Sumiyoshi, M.; Kobayashi, T. Whey peptides prevent chronic ultraviolet B radiation-induced skin aging in melanin-possessing male hairless mice. J. Nutr. 2014, 144, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Gruber, R.; Karreth, F.; Frommlet, F.; Fischer, M.B.; Watzek, G. Platelets are mitogenic for periosteum-derived cells. J. Orthop. Res. 2003, 21, 941–948. [Google Scholar] [CrossRef]
- He, S.; Chen, M.; Lin, X.; Lv, Z.; Liang, R.; Huang, L. Triptolide inhibits PDGF-induced proliferation of ASMCs through G0/G1 cell cycle arrest and suppression of the AKT/NF-kappaB/cyclinD1 signaling pathway. Eur. J. Pharmacol. 2020, 867, 172811. [Google Scholar] [CrossRef] [PubMed]
- Heeboll-Nielsen, A.; Justesen, S.F.; Thomas, O.R. Fractionation of whey proteins with high-capacity superparamagnetic ion-exchangers. J. Biotechnol. 2004, 113, 247–262. [Google Scholar] [CrossRef] [PubMed]
- Wen-Qiong, W.; Yun-Chao, W.; Xiao-Feng, Z.; Rui-Xia, G.; Mao-Lin, L. Whey protein membrane processing methods and membrane fouling mechanism analysis. Food. Chem. 2019, 289, 468–481. [Google Scholar] [CrossRef] [PubMed]
- Baat, C.A.; Tortorello, M.-L. Encyclopedia of Food Microbiology; Academic Press: Cambridge, MA, USA, 2014. [Google Scholar]
- Shiou, S.R.; Yu, Y.; Guo, Y.; Westerhoff, M.; Lu, L.; Petrof, E.O.; Sun, J.; Claud, E.C. Oral administration of transforming growth factor-beta1 (TGF-beta1) protects the immature gut from injury via Smad protein-dependent suppression of epithelial nuclear factor kappaB (NF-kappaB) signaling and proinflammatory cytokine production. J. Biol. Chem. 2013, 288, 34757–34766. [Google Scholar] [CrossRef] [Green Version]
- Oddy, W.H.; McMahon, R.J. Milk-derived or recombinant transforming growth factor-beta has effects on immunological outcomes: A review of evidence from animal experimental studies. Clin. Exp. Allergy 2011, 41, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Nishimine, M.; Nakamura, M.; Mishima, K.; Kishi, M.; Kirita, T.; Sugimura, M.; Konishi, N. Id proteins are overexpressed in human oral squamous cell carcinomas. J. Oral. Pathol. Med. 2003, 32, 350–357. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, F.; Wang, D.; Yang, Z.; Liu, S.; Dong, Z. Prognostic ability of DNA-binding protein inhibitor ID-1 expression in patients with oral squamous cell carcinoma. Oncol. Lett. 2020, 19, 3917–3922. [Google Scholar] [CrossRef] [Green Version]
- Luo, K.J.; Wen, J.; Xie, X.; Fu, J.H.; Luo, R.Z.; Wu, Q.L.; Hu, Y. Prognostic relevance of Id-1 expression in patients with resectable esophageal squamous cell carcinoma. Ann. Thorac. Surg. 2012, 93, 1682–1688. [Google Scholar] [CrossRef] [PubMed]
- Mody, A.A.; Wordinger, R.J.; Clark, A.F. Role of ID Proteins in BMP4 Inhibition of Profibrotic Effects of TGF-beta2 in Human TM Cells. Invest. Ophthalmol. Vis. Sci. 2017, 58, 849–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strong, N.; Millena, A.C.; Walker, L.; Chaudhary, J.; Khan, S.A. Inhibitor of differentiation 1 (Id1) and Id3 proteins play different roles in TGFbeta effects on cell proliferation and migration in prostate cancer cells. Prostate 2013, 73, 624–633. [Google Scholar] [CrossRef] [Green Version]
- Heidebrecht, H.J.; Kulozik, U. Data concerning the fractionation of individual whey proteins and casein micelles by microfiltration with ceramic gradient membranes. Data. Brief. 2019, 25, 104102. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panahipour, L.; Husejnovic, S.; Nasirzade, J.; Semelmayer, S.; Gruber, R. Micellar Casein and Whey Powder Hold a TGF-β Activity and Regulate ID Genes In Vitro. Molecules 2021, 26, 507. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26020507
Panahipour L, Husejnovic S, Nasirzade J, Semelmayer S, Gruber R. Micellar Casein and Whey Powder Hold a TGF-β Activity and Regulate ID Genes In Vitro. Molecules. 2021; 26(2):507. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26020507
Chicago/Turabian StylePanahipour, Layla, Selma Husejnovic, Jila Nasirzade, Stephan Semelmayer, and Reinhard Gruber. 2021. "Micellar Casein and Whey Powder Hold a TGF-β Activity and Regulate ID Genes In Vitro" Molecules 26, no. 2: 507. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26020507