
Discovery of a Kojibiose Hydrolase by Analysis of Specificity-Determining Correlated Positions in Glycoside Hydrolase Family 65


Abstract
:
1. Introduction
2. Results
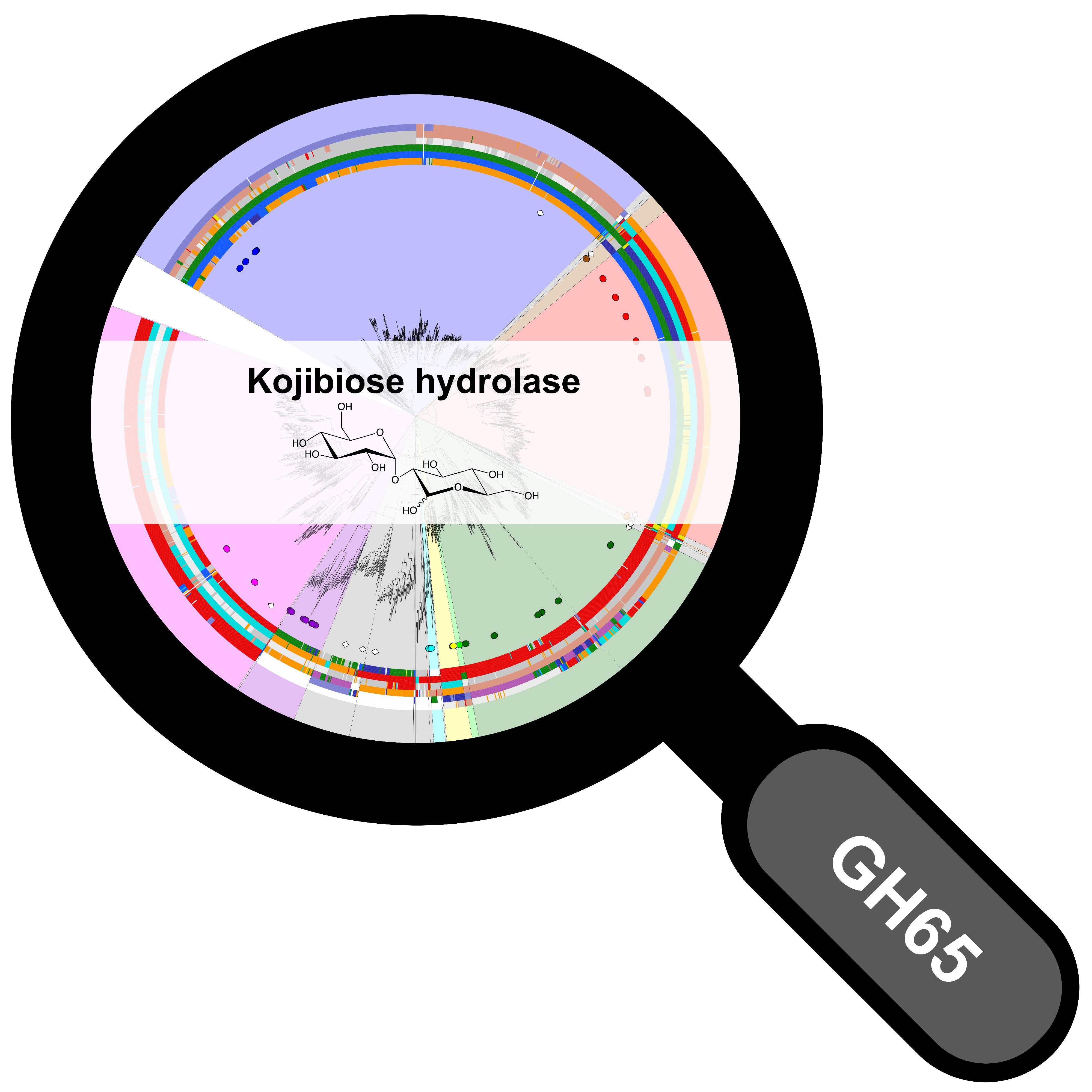
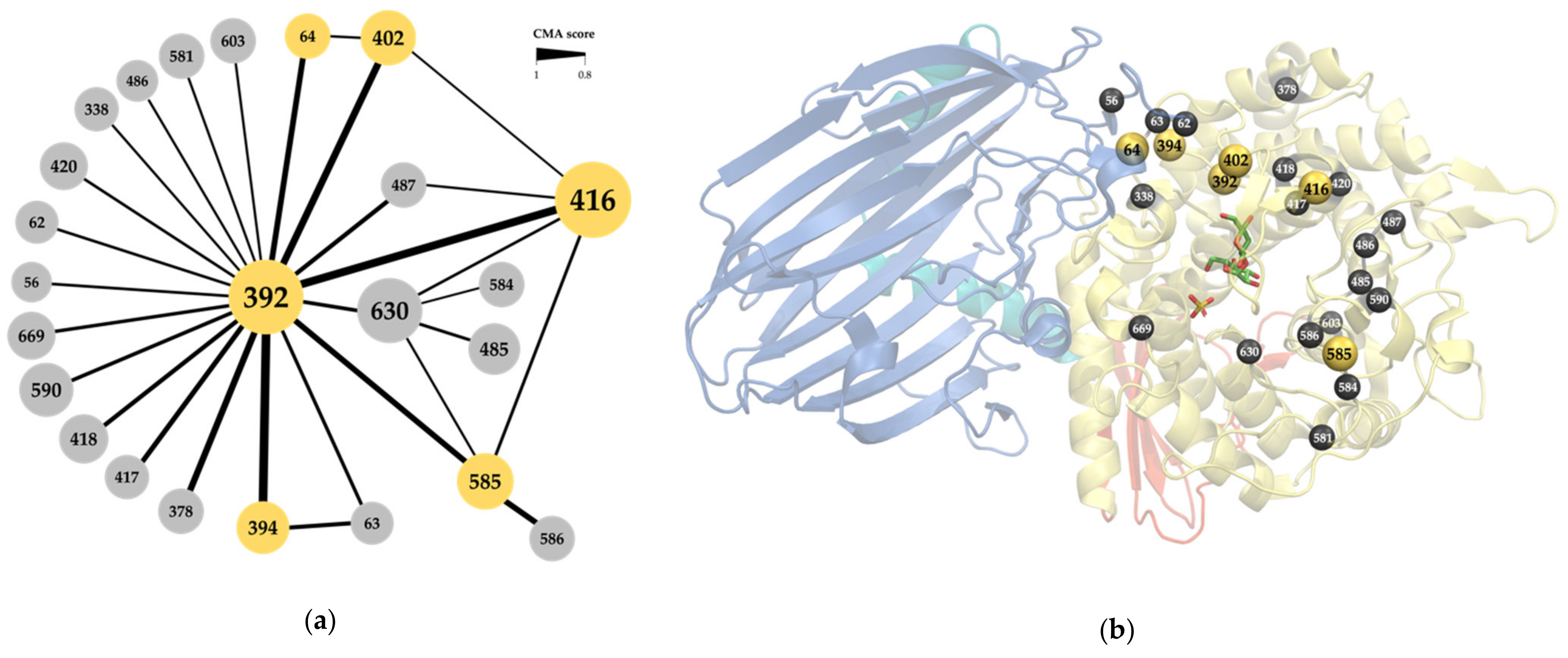
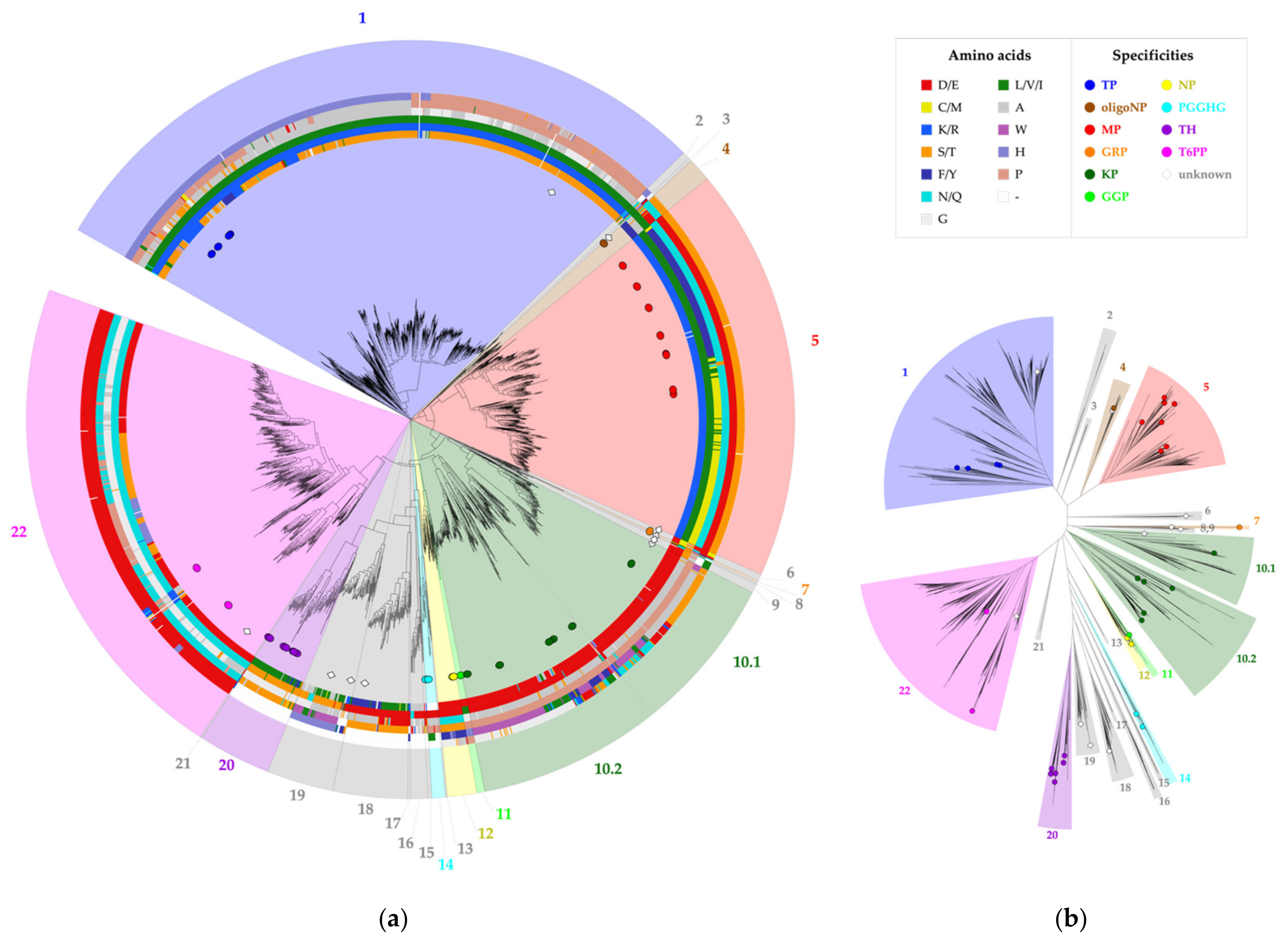
2.1. Correlated Mutations Analysis
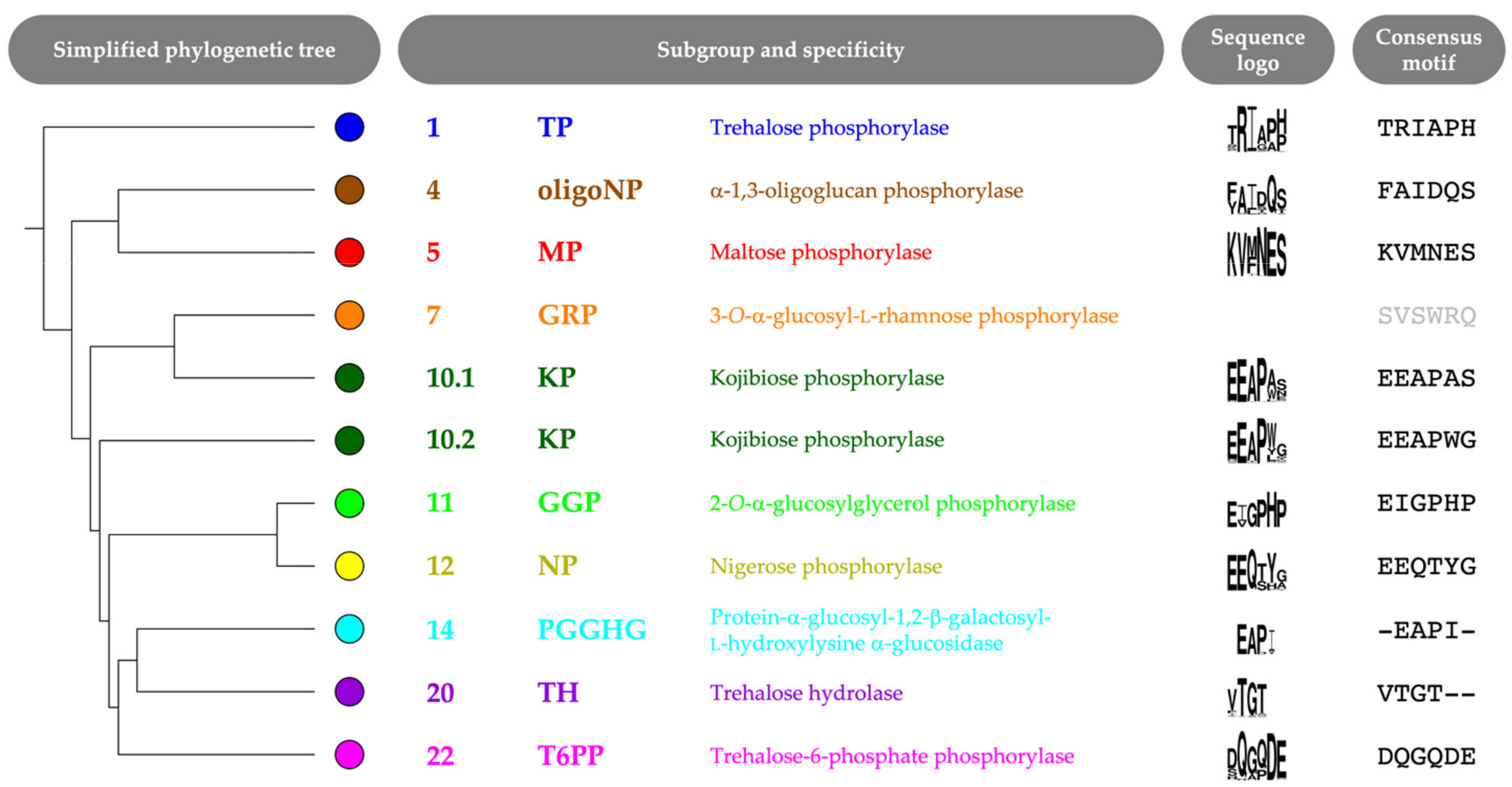
2.2. Correlated Positions as Specificity Determinants
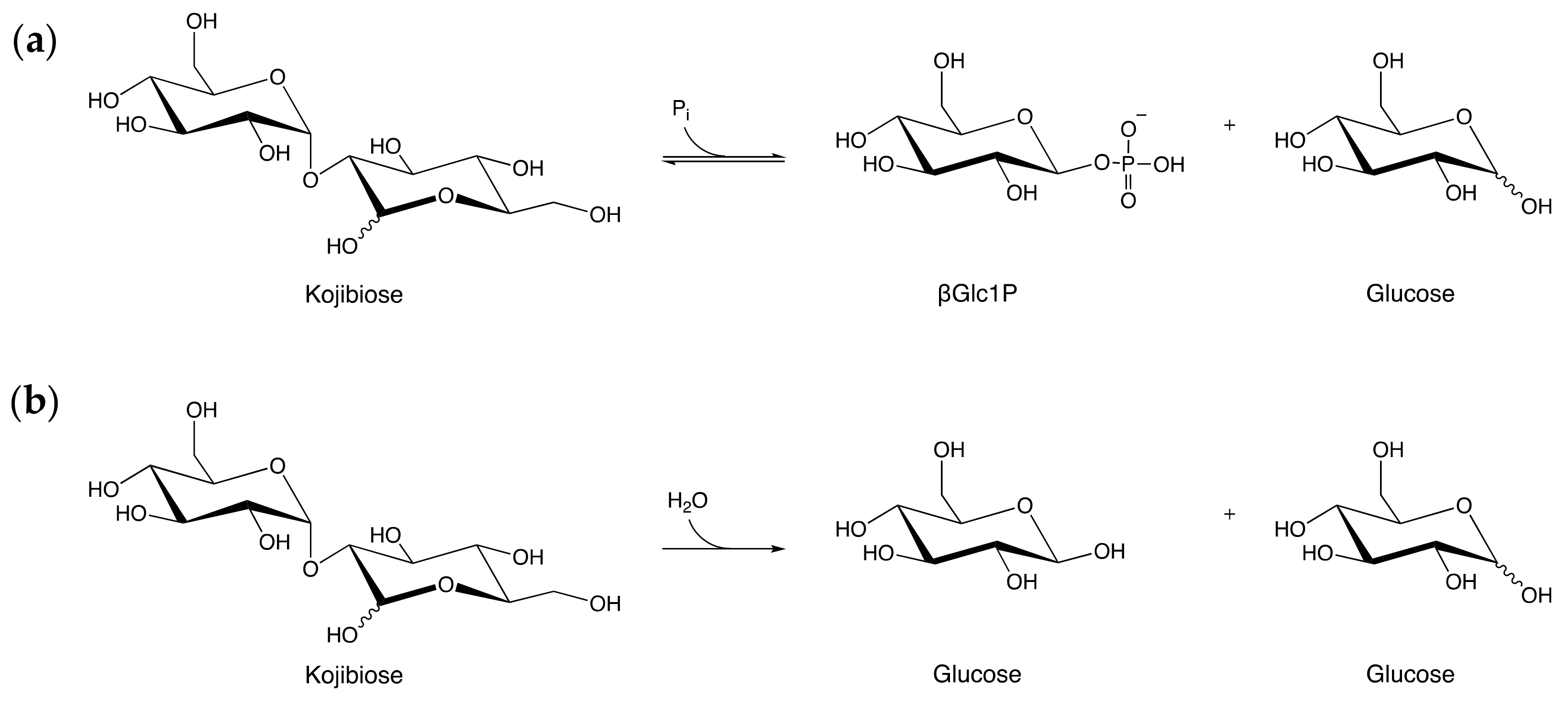
2.3. Specificity-Determining Correlated Positions as Tool for Enzyme Discovery
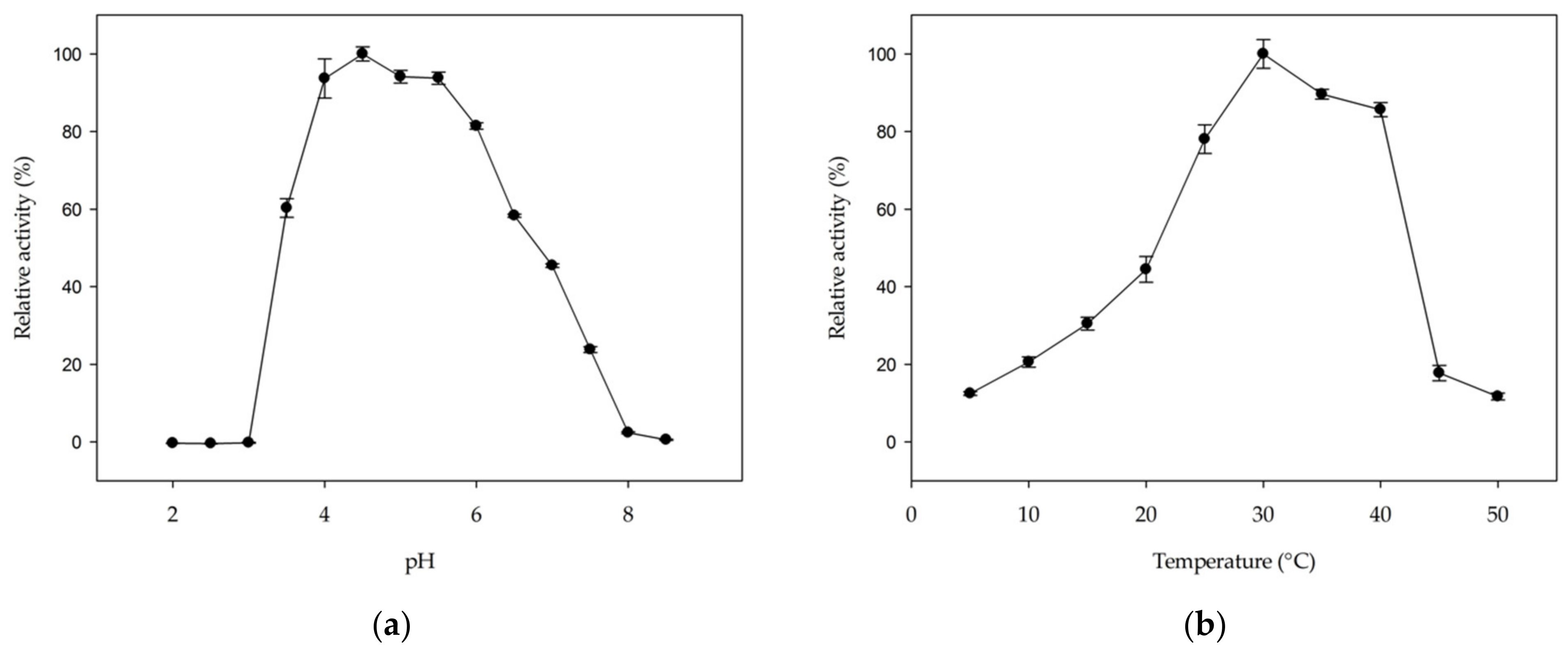
2.4. Optimal pH and Temperature and Kinetic Properties of MmGH
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Sequence Analysis
4.3. Gene Cloning and Transformation
4.4. Enzyme Expression and Purification
4.5. Detection of Reaction Components
4.6. Screening of Potential Substrates
4.7. Optimal pH and Temperature and Kinetic Properties of MmGH
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; You, C. Disaccharide phosphorylases: Structure, catalytic mechanisms and directed evolution. Synth. Syst. Biotechnol. 2021, 6, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Desmet, T.; Soetaert, W. Enzymatic glycosyl transfer: Mechanisms and applications. Biocatal. Biotransform. 2011, 29, 1–18. [Google Scholar] [CrossRef]
- Okada, S.; Yamamoto, T.; Watanabe, H.; Nishimoto, T.; Chaen, H.; Fukuda, S.; Wakagi, T.; Fushinobu, S. Structural and mutational analysis of substrate recognition in kojibiose phosphorylase. FEBS J. 2014, 281, 778–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nihira, T.; Nishimoto, M.; Nakai, H.; Ohtsubo, K.; Kitaoka, M. Characterization of Two α-1,3-Glucoside Phosphorylases from Clostridium phytofermentans. J. Appl. Glycosci. 2014, 61, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Hamazaki, H.; Hamazaki, M.H. Catalytic site of human protein-glucosylgalactosylhydroxylysine glucosidase: Three crucial carboxyl residues were determined by cloning and site-directed mutagenesis. Biochem. Biophys. Res. Commun. 2016, 469, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Hüwel, S.; Haalck, L.; Conrath, N.; Spener, F. Maltose phosphorylase from Lactobacillus brevis: Purification, characterization, and application in a biosensor for ortho-phosphate. Enzyme Microb. Technol. 1997, 21, 413–420. [Google Scholar] [CrossRef]
- Ehrmann, M.A.; Vogel, R.F. Maltose metabolism of Lactobacillus sanfranciscensis: Cloning and heterologous expression of the key enzymes, maltose phosphorylase and phosphoglucomutase. FEMS Microbiol. Lett. 1998, 169, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Inoue, Y.; Yasutake, N.; Oshima, Y.; Yamamoto, Y.; Tomita, T.; Miyoshi, S.; Yatake, T. Cloning of the Maltose Phosphorylase Gene from Bacillus sp. Strain RK-1 and Efficient Production of the Cloned Gene and the Trehalose Phosphorylase Gene from Bacillus stearothermophilus SK-1 in Bacillus subtilis. Biosci. Biotechnol. Biochem. 2002, 66, 2594–2599. [Google Scholar] [CrossRef]
- Hidaka, Y.; Hatada, Y.; Akita, M.; Yoshida, M.; Nakamura, N.; Takada, M.; Nakakuki, T.; Ito, S.; Horikoshi, K. Maltose phosphorylase from a deep-sea Paenibacillus sp.: Enzymatic properties and nucleotide and amino-acid sequences. Enzyme Microb. Technol. 2005, 37, 185–194. [Google Scholar] [CrossRef]
- Nakai, H.; Baumann, M.J.; Petersen, B.O.; Westphal, Y.; Schols, H.; Dilokpimol, A.; Hachem, M.A.; Lahtinen, S.J.; Duus, J.Ø.; Svensson, B. The maltodextrin transport system and metabolism in Lactobacillus acidophilus NCFM and production of novel α-glucosides through reverse phosphorolysis by maltose phosphorylase. FEBS J. 2009, 276, 7353–7365. [Google Scholar] [CrossRef] [PubMed]
- Nihira, T.; Saito, Y.; Kitaoka, M.; Otsubo, K.; Nakai, H. Identification of Bacillus selenitireducens MLS10 maltose phosphorylase possessing synthetic ability for branched α-D-glucosyl trisaccharides. Carbohydr. Res. 2012, 360, 25–30. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, A.; Blancato, V.S.; Repizo, G.D.; Henry, C.; Pikis, A.; Bourand, A.; de Fátima Álvarez, M.; Immel, S.; Mechakra-Maza, A.; Hartke, A.; et al. Enterococcus faecalis utilizes maltose by connecting two incompatible metabolic routes via a novel maltose 6′-phosphate phosphatase (MapP). Mol. Microbiol. 2013, 88, 234–253. [Google Scholar] [CrossRef] [Green Version]
- Chaen, H.; Nakada, T.; Nishimoto, T.; Kuroda, N.; Fukuda, S.; Sugimoto, T.; Kurimoto, M.; Yoshio, T. Purification and characterization of thermostable trehalose phosphorylase from Thermoanaerobium brockii. J. Appl. Glycosci. 1999, 46, 399–405. [Google Scholar] [CrossRef]
- Inoue, Y.; Ishii, K.; Tomita, T.; Yatake, T.; Fukui, F. Characterization of trehalose phosphorylase from Bacillus stearothermophilus SK-1 and nucleotide sequence of the corresponding gene. Biosci. Biotechnol. Biochem. 2002, 66, 1835–1843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Borght, J.; Chen, C.; Hoflack, L.; Van Renterghem, L.; Desmet, T.; Soetaert, W. Enzymatic properties and substrate specificity of the trehalose phosphorylase from Caldanaerobacter subterraneus. Appl. Environ. Microbiol. 2011, 77, 6939–6944. [Google Scholar] [CrossRef] [Green Version]
- Nihira, T.; Saito, Y.; Chiku, K.; Kitaoka, M.; Ohtsubo, K.; Nakai, H. Potassium ion-dependent trehalose phosphorylase from halophilic Bacillus selenitireducens MLS10. FEBS Lett. 2013, 587, 3382–3386. [Google Scholar] [CrossRef] [Green Version]
- Andersson, U.; Levander, F.; Rådström, P. Trehalose-6-phosphate Phosphorylase Is Part of a Novel Metabolic Pathway for Trehalose Utilization in Lactococcus lactis. J. Biol. Chem. 2001, 276, 42707–42713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nihira, T.; Nakai, H.; Chiku, K.; Kitaoka, M. Discovery of nigerose phosphorylase from Clostridium phytofermentans. Appl. Microbiol. Biotechnol. 2012, 93, 1513–1522. [Google Scholar] [CrossRef]
- Chaen, H.; Yamamoto, T.; Nishimoto, T.; Nakada, T.; Fukuda, S.; Sugimoto, T.; Kurimoto, M.; Tsujisaka, Y. Purification and Characterization of a Novel Phosphorylase, Kojibiose Phosphorylase, from Thermoanaerobium brockii. J. Appl. Glycosci. 1999, 46, 423–429. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Maruta, K.; Mukai, K.; Yamashita, H.; Nishimoto, T.; Kubota, M.; Fukuda, S.; Kurimoto, M.; Tsujisaka, Y. Cloning and sequencing of kojibiose phosphorylase gene from Thermoanaerobacter brockii ATCC35047. J. Biosci. Bioeng. 2004, 98, 99–106. [Google Scholar] [CrossRef]
- Yamamoto, T.; Nishio-Kosaka, M.; Izawa, S.; Aga, H.; Nishimoto, T.; Chaen, H.; Fukuda, S. Enzymatic properties of recombinant kojibiose phosphorylase from Caldicellulosiruptor saccharolyticus ATCC43494. Biosci. Biotechnol. Biochem. 2011, 75, 1208–1210. [Google Scholar] [CrossRef]
- Jung, J.H.; Seo, D.H.; Holden, J.F.; Park, C.S. Identification and characterization of an archaeal kojibiose catabolic pathway in the hyperthermophilic Pyrococcus sp. strain ST04. J. Bacteriol. 2014, 196, 1122–1131. [Google Scholar] [CrossRef]
- Mukherjee, K.; Narindoshvili, T.; Raushel, F.M. Discovery of a Kojibiose Phosphorylase in Escherichia coli K-12. Biochemistry 2018, 57, 2857–2867. [Google Scholar] [CrossRef] [PubMed]
- Nihira, T.; Nakai, H.; Kitaoka, M. 3-O-α-D-glucopyranosyl-L-rhamnose phosphorylase from Clostridium phytofermentans. Carbohydr. Res. 2012, 350, 94–97. [Google Scholar] [CrossRef] [PubMed]
- Nihira, T.; Saito, Y.; Ohtsubo, K.; Nakai, H.; Kitaoka, M. 2-O-α-D-Glucosylglycerol Phosphorylase from Bacillus selenitireducens MLS10 Possessing Hydrolytic Activity on β-D-Glucose 1-Phosphate. PLoS ONE 2014, 9, e86548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alizadeh, P.; Klionsky, D.J. Purification and biochemical characterization of the ATH1 gene product, vacuolar acid trehalase, from Saccharomyces cerevisiae. FEBS Lett. 1996, 391, 273–278. [Google Scholar] [CrossRef] [Green Version]
- D’Enfert, C.; Fontaine, T. Molecular characterization of the Aspergillus nidulans treA gene encoding an acid trehalase required for growth on trehalose. Mol. Microbiol. 1997, 24, 203–216. [Google Scholar] [CrossRef]
- Weinstock, K.G.; Bush, D. Nucleic Acid Sequences Relating to Candida albicans for Diagnostics and Therapeutics. U.S. Patent US6747137, 12 February 1999. [Google Scholar]
- Pedreño, Y.; Maicas, S.; Argüelles, J.C.; Sentandreu, R.; Valentin, E. The ATC1 gene encodes a cell wall-linked acid trehalase required for growth on trehalose in Candida albicans. J. Biol. Chem. 2004, 279, 40852–40860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, Z.; Yin, Y.; Cao, Y.; Zhao, H.; Xia, Y. Expression, purification, and characterization of recombinant Metarhizium anisopliae acid trehalase in Pichia pastoris. Protein Expr. Purif. 2007, 54, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Fresneda, R.; Martínez-Esparza, M.; Maicas, S.; Argüelles, J.-C.; Valentín, E. In Candida parapsilosis the ATC1 Gene Encodes for an Acid Trehalase Involved in Trehalose Hydrolysis, Stress Resistance and Virulence. PLoS ONE 2014, 9, e99113. [Google Scholar] [CrossRef]
- Zilli, D.M.W.; Lopes, R.G.; Alves, S.L.; Barros, L.M.; Miletti, L.C.; Stambuk, B.U. Secretion of the acid trehalase encoded by the CgATH1 gene allows trehalose fermentation by Candida glabrata. Microbiol. Res. 2015, 179, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, T.; Yamashita, H.; Mukai, K.; Watanabe, H.; Kubota, M.; Chaen, H.; Fukuda, S. Construction and characterization of chimeric enzymes of kojibiose phosphorylase and trehalose phosphorylase from Thermoanaerobacter brockii. Carbohydr. Res. 2006, 341, 2350–2359. [Google Scholar] [CrossRef] [PubMed]
- Nakai, H.; Petersen, B.O.; Westphal, Y.; Dilokpimol, A.; Abou Hachem, M.; Duus, J.Ø.; Schols, H.A.; Svensson, B. Rational engineering of Lactobacillus acidophilus NCFM maltose phosphorylase into either trehalose or kojibiose dual specificity phosphorylase. Protein Eng. Des. Sel. 2010, 23, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Franceus, J.; Verhaeghe, T.; Desmet, T. Correlated positions in protein evolution and engineering. J. Ind. Microbiol. Biotechnol. 2017, 44, 687–695. [Google Scholar] [CrossRef]
- Kuipers, R.K.P.; Joosten, H.-J.; Verwiel, E.; Paans, S.; Akerboom, J.; van der Oost, J.; Leferink, N.G.H.; van Berkel, W.J.H.; Vriend, G.; Schaap, P.J. Correlated mutation analyses on super-family alignments reveal functionally important residues. Proteins Struct. Funct. Bioinform. 2009, 76, 608–616. [Google Scholar] [CrossRef]
- Forsberg, Z.; Bissaro, B.; Gullesen, J.; Dalhus, B.; Vaaje-Kolstad, G.; Eijsink, V.G.H. Structural determinants of bacterial lytic polysaccharide monooxygenase functionality. J. Biol. Chem. 2018, 293, 1397–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsberg, Z.; Stepnov, A.A.; Nærdal, G.K.; Klinkenberg, G.; Eijsink, V.G.H. Engineering lytic polysaccharide monooxygenases (LPMOs). Methods Enzymol. 2020, 644, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, M.; Gevaert, O.; Van Overtveldt, S.; Lange, J.; Joosten, H.J.; Desmet, T.; Beerens, K. Structure-function relationships in NDP-sugar active SDR enzymes: Fingerprints for functional annotation and enzyme engineering. Biotechnol. Adv. 2021, 48, 107705. [Google Scholar] [CrossRef]
- Egloff, M.P.; Uppenberg, J.; Haalck, L.; van Tilbeurgh, H. Crystal structure of maltose phosphorylase from Lactobacillus brevis: Unexpected evolutionary relationship with glucoamylases. Structure 2001, 9, 689–697. [Google Scholar] [CrossRef] [Green Version]
- Touhara, K.K.; Nihira, T.; Kitaoka, M.; Nakai, H.; Fushinobu, S. Structural basis for reversible phosphorolysis and hydrolysis reactions of 2-O-α-Glucosylglycerol phosphorylase. J. Biol. Chem. 2014, 289, 18067–18075. [Google Scholar] [CrossRef] [Green Version]
- Mittenbuhler, K.; Holzer, H. Purification and characterization of acid trehalase from the yeast suc2 mutant. J. Biol. Chem. 1988, 263, 8537–8543. [Google Scholar] [CrossRef]
- Destruelle, M.; Holzer, H.; Klionsky, D.J. Isolation and characterization of a novel yeast gene, ATH1, that is required for vacuolar acid trehalase activity. Yeast 1995, 11, 1015–1025. [Google Scholar] [CrossRef] [PubMed]
- Sternberg, M.; Spiro, R.G. Studies on the Catabolism of the Hydroxylysine-Linked Disaccharide Units of Basement Membranes and Collagens. Ren. Physiol. 1980, 3, 1–3. [Google Scholar] [CrossRef]
- Männistö, M.K.; Tiirola, M.; McConnell, J.; Häggblom, M.M. Mucilaginibacter frigoritolerans sp. nov., Mucilaginibacter lappiensis sp. nov. and Mucilaginibacter mallensis sp. nov., isolated from soil and lichen samples. Int. J. Syst. Evol. Microbiol. 2010, 60, 2849–2856. [Google Scholar] [CrossRef] [Green Version]
- Kitaoka, M. Diversity of phosphorylases in glycoside hydrolase families. Appl. Microbiol. Biotechnol. 2015, 99, 8377–8390. [Google Scholar] [CrossRef] [PubMed]
- Franceus, J.; Decuyper, L.; D’hooghe, M.; Desmet, T. Exploring the sequence diversity in glycoside hydrolase family 13_18 reveals a novel glucosylglycerol phosphorylase. Appl. Microbiol. Biotechnol. 2018, 102, 3183–3191. [Google Scholar] [CrossRef] [PubMed]
- Franceus, J.; Pinel, D.; Desmet, T. Glucosylglycerate Phosphorylase, an Enzyme with Novel Specificity Involved in Compatible Solute Metabolism. Appl. Environ. Microbiol. 2017, 83, e01434-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceus, J.; Lormans, J.; Cools, L.; D’hooghe, M.; Desmet, T. Evolution of Phosphorylases from N-Acetylglucosaminide Hydrolases in Family GH3. ACS Catal. 2021, 11, 6225–6233. [Google Scholar] [CrossRef]
- Beerens, K.; De Winter, K.; Van de Walle, D.; Grootaert, C.; Kamiloglu, S.; Miclotte, L.; Van de Wiele, T.; Van Camp, J.; Dewettinck, K.; Desmet, T. Biocatalytic synthesis of the rare sugar kojibiose: Process scale-up and application testing. J. Agric. Food Chem. 2017, 65, 6030–6041. [Google Scholar] [CrossRef] [PubMed]
- Franceus, J.; Dhaene, S.; Decadt, H.; Vandepitte, J.; Caroen, J.; Van der Eycken, J.; Beerens, K.; Desmet, T. Rational design of an improved transglucosylase for production of the rare sugar nigerose. Chem. Commun. 2019, 55, 4531–4533. [Google Scholar] [CrossRef] [PubMed]
- Van der Borght, J.; Desmet, T.; Soetaert, W. Enzymatic production of β-D-glucose-1-phosphate from trehalose. Biotechnol. J. 2010, 5, 986–993. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Niu, B.; Gao, Y.; Fu, L.; Li, W. CD-HIT Suite: A web server for clustering and comparing biological sequences. Bioinformatics 2010, 26, 680–682. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. Clustal Omega for making accurate alignments of many protein sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [Green Version]
- Rozewicki, J.; Li, S.; Amada, K.M.; Standley, D.M.; Katoh, K. MAFFT-DASH: Integrated protein sequence and structural alignment. Nucleic Acids Res. 2019, 47, W5–W10. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Brief. Bioinf. 2008, 9, 286–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemoine, F.; Domelevo Entfellner, J.B.; Wilkinson, E.; Correia, D.; Dávila Felipe, M.; De Oliveira, T.; Gascuel, O. Renewing Felsenstein’s phylogenetic bootstrap in the era of big data. Nature 2018, 556, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.-M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrödinger LLC. The PyMOL Molecular Graphics System; Version 2.4; Schrödinger, LLC: New York, NY, USA.
- Gawronski, J.D.; Benson, D.R. Microtiter assay for glutamine synthetase biosynthetic activity using inorganic phosphate detection. Anal. Biochem. 2004, 327, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Blecher, M.; Glassman, A.B. Determination of glucose in the presence of sucrose using glucose oxidase; effect of pH on absorption spectrum of oxidized o-dianisidine. Anal. Biochem. 1962, 3, 343–352. [Google Scholar] [CrossRef]
- Chang, A.; Jeske, L.; Ulbrich, S.; Hofmann, J.; Koblitz, J.; Schomburg, I.; Neumann-Schaal, M.; Jahn, D.; Schomburg, D. BRENDA, the ELIXIR core data resource in 2021: New developments and updates. Nucleic Acids Res. 2021, 49, D498–D508. [Google Scholar] [CrossRef] [PubMed]
- Fierobe, H.-P.; Clarke, A.J.; Tull, D.; Svensson, B. Enzymatic Properties of the Cysteinesulfinic Acid Derivative of the Catalytic-Base Mutant Glu400→Cys of Glucoamylase from Aspergillus awamori. Biochemistry 1998, 37, 3753–3759. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Ueda, Y.; Nakamura, N.; Abe, S. Hydrolysis of low molecular weight isomaltosaccharides by a p-nitrophenyl-α-D-glucopyranoside-hydrolyzing α-glucosidase from a thermophile, Bacillus thermoglucosidius KP 1006. Biochim. Biophys. Acta Enzymol. 1979, 566, 62–66. [Google Scholar] [CrossRef]
- Suzuki, Y.; Uchida, K. Three forms of α-glucosidase from welsh onion. Agric. Biol. Chem. 1984, 48, 1343–1345. [Google Scholar] [CrossRef]
- Wongchawalit, J.; Yamamoto, T.; Nakai, H.; Kim, Y.-M.; Sato, N.; Nishimoto, M.; Okuyama, M.; Mori, H.; Saji, O.; Chanchao, C.; et al. Purification and Characterization of α-Glucosidase I from Japanese Honeybee (Apis cerana japonica) and Molecular Cloning of Its cDNA. Biosci. Biotechnol. Biochem. 2006, 70, 2889–2898. [Google Scholar] [CrossRef] [Green Version]
- Kimura, A.; Takewaki, S.; Matsui, H.; Kubota, M.; Chiba, S. Allosteric properties, substrate specificity, and subsite affinities of honeybee α-glucosidase, I. J. Biochem. 1990, 107, 762–768. [Google Scholar] [CrossRef]
- Kato, N.; Suyama, S.; Shirokane, M.; Kato, M.; Kobayashi, T.; Tsukagoshi, N. Novel α-glucosidase from Aspergillus nidulans with strong transglycosylation activity. Appl. Environ. Microbiol. 2002, 68, 1250–1256. [Google Scholar] [CrossRef] [Green Version]
- Kita, A.; Matsui, H.; Somoto, A.; Kimura, A.; Takata, M.; Chiba, S. Substrate specificity and subsite affinities of crystalline α-glucosidase from Aspergillus niger. Agric. Biol. Chem. 2014, 55, 2327–2335. [Google Scholar] [CrossRef]
- Kim, N.R.; Jeong, D.W.; Ko, D.S.; Shim, J.H. Characterization of novel thermophilic alpha-glucosidase from Bifidobacterium longum. Int. J. Biol. Macromol. 2017, 99, 594–599. [Google Scholar] [CrossRef]
- Bravo-Torres, J.C.; Villagómez-Castro, J.C.; Calvo-Méndez, C.; Flores-Carreón, A.; López-Romero, E. Purification and biochemical characterisation of a membrane-bound α-glucosidase from the parasite Entamoeba histolytica. Int. J. Parasitol. 2004, 34, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Giudicelli, J.; Emiliozzi, R.; Vannier, C.; De Burlet, G.; Sudaka, P. Purification by affinity chromatography and characterization of a neutral α-glucosidase from horse kidney. Biochim. Biophys. Acta Enzymol. 1980, 612, 85–96. [Google Scholar] [CrossRef]
- Tanaka, Y.; Aki, T.; Hidaka, Y.; Furuya, Y.; Kawamoto, S.; Shigeta, S.; Ono, K.; Suzuki, O. Purification and characterization of a novel fungal α-glucosidase from mortierella alliacea with high starch-hydrolytic activity. Biosci. Biotechnol. Biochem. 2002, 66, 2415–2423. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, Y.; Miyake, T.; Suzuki, Y. Properties of crystalline α-glucosidase from mucor javanicus. Agric. Biol. Chem. 1973, 37, 251–259. [Google Scholar] [CrossRef]
- Matsui, H.; Ito, H.; Chiba, S. Low Molecular—Activity α-glucosidase from ungerminated rice seed. Agric. Biol. Chem. 1988, 52, 1859–1860. [Google Scholar] [CrossRef]
- Yamasaki, Y.; Fujimoto, M.; Kariya, J.; Konno, H. Purification and characterization of an α-glucosidase from germinating millet seeds. Phytochemistry 2005, 66, 851–857. [Google Scholar] [CrossRef]
- Kobayashi, I.; Tokuda, M.; Hashimoto, H.; Konda, T.; Nakano, H.; Kitahata, S. Purification and characterization of a new type of α-glucosidase from paecilomyces lilacinus that has transglucosylation activity to produce α-1,3- and α-1,2-linked…. Biosci. Biotechnol. Biochem. 2003, 67, 29–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okuyama, M.; Tanimoto, Y.; Ito, T.; Anzai, A.; Mori, H.; Kimura, A.; Matsui, H.; Chiba, S. Purification and characterization of the hyper-glycosylated extracellular α-glucosidase from Schizosaccharomyces pombe. Enzyme Microb. Technol. 2005, 37, 472–480. [Google Scholar] [CrossRef]
- Choi, K.-H.; Hwang, S.; Cha, J. Identification and characterization of malA in the maltose/maltodextrin operon of Sulfolobus acidocaldarius DSM639. J. Bacteriol. 2013, 195, 1789–1799. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, S.H.; Norén, O.; Sjöström, H.; Danielsen, E.M. Amphiphilic Pig Intestinal Microvillus Maltase/Glucoamylase. Eur. J. Biochem. 1982, 126, 559–568. [Google Scholar] [CrossRef]
- Seo, S.H.; Choi, K.H.; Hwang, S.; Kim, J.; Park, C.S.; Rho, J.R.; Cha, J. Characterization of the catalytic and kinetic properties of a thermostable Thermoplasma acidophilum α-glucosidase and its transglucosylation reaction with arbutin. J. Mol. Catal. B Enzym. 2011, 72, 305–312. [Google Scholar] [CrossRef]
- Alarico, S.; Da Costa, M.S.; Empadinhas, N. Molecular and physiological role of the trehalose-hydrolyzing α-glucosidase from Thermus thermophilus HB27. J. Bacteriol. 2008, 190, 2298–2305. [Google Scholar] [CrossRef] [Green Version]
- Matsui, H.; Yazawa, I.; Chiba, S. Purification and substrate specificity of sweet corn α-glucosidase. Agric. Biol. Chem. 1981, 45, 887–894. [Google Scholar] [CrossRef]
- Saburi, W.; Mori, H.; Saito, S.; Okuyama, M.; Kimura, A. Structural elements in dextran glucosidase responsible for high specificity to long chain substrate. Biochim. Biophys. Acta Proteins Proteom. 2006, 1764, 688–698. [Google Scholar] [CrossRef]
- Kurakata, Y.; Uechi, A.; Yoshida, H.; Kamitori, S.; Sakano, Y.; Nishikawa, A.; Tonozuka, T. Structural Insights into the Substrate Specificity and Function of Escherichia coli K12 YgjK, a Glucosidase Belonging to the Glycoside Hydrolase Family 63. J. Mol. Biol. 2008, 381, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Torres-Rodríguez, B.I.; Flores-Berrout, K.; Villagómez-Castro, J.C.; López-Romero, E. Purification and partial biochemical characterization of a membrane-bound type II-like α-glucosidase from the yeast morphotype of Sporothrix schenckii. Antonie Van Leeuwenhoek 2012, 101, 313–322. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EC | Enzyme | Abbreviation | Substrate | Product | Organisms with Annotated Representatives |
|---|---|---|---|---|---|
| 2.4.1.8 | Maltose phosphorylase | MP | Glcα1→4Glc | βGlc1P | Levilactobacillus brevis [7] Fructilactobacillus sanfranciscensis [8] Bacillus sp. [9] Paenibacillus sp. [10] Lactobacillus acidophilus sp. [11] Salisediminibacterium selenitireducens [12] Enterococcus faecalis [13] |
| 2.4.1.64 | Trehalose phosphorylase | TP | Glcα1→1αGlc | βGlc1P | Thermoanaerobacter brockii [14] Geobacillus stearothermophilus [15] Caldanaerobacter subterraneus [16] Salisediminibacterium selenitireducens [17] |
| 2.4.1.216 | Trehalose-6-phosphate phosphorylase | T6PP | Glcα1→1αGlc6P | βGlc1P | Lactococcus lactis [18] Ca. Kuenenia stuttgartiensis [19] |
| 2.4.1.230 | Kojibiose phosphorylase | KP | Glcα1→2Glc Glcα1→(2Glcα1→)n-22Glc | βGlc1P | Thermoanaerobacter brockii [20,21] Caldicellulosiruptor saccharolyticus [22] Nostoc sp. [19] Pyrococcus sp. [23] Escherichia coli [24] |
| 2.4.1.279 | Nigerose phosphorylase | NP | Glcα1→3Glc | βGlc1P | Lachnoclostridium phytofermentans [5,19] |
| 2.4.1.282 | 3-O-α-glucosyl-l-rhamnose phosphorylase | GRP | Glcα1→3l-Rham | βGlc1P | Lachnoclostridium phytofermentans [25] |
| 2.4.1.332 | 2-O-α-glucosylglycerol phosphorylase | GGP | Glcα1→2Glycerol | βGlc1P | Salisediminibacterium selenitireducens [26] |
| 2.4.1.334 | α-1,3-oligoglucan phosphorylase | oligoNP | Glcα1→(3Glcα1→)n-23Glc | βGlc1P | Lachnoclostridium phytofermentans [5] |
| 3.2.1.28 | Trehalose hydrolase (trehalase) | TH | Glcα1→1αGlc | Glc | Saccharomyces cerevisiae [27] Aspergillus nidulans [28] Candida albicans [29,30] Metarhizium anisopliae [31] Candida parapsilosis [32] Candida glabrata [33] |
| 3.2.1.107 | Protein-α-glucosyl-1,2-β-galactosyl-l-hydroxylysine α-glucosidase | PGGHG | Glcα1→2Galβ1→5Hyl | Glc | Gallus gallus [6] Homo sapiens [6] |
| Substrate | Structure | Specific Activity (U/mg) 1 | Relative Activity (%) |
|---|---|---|---|
| Kojibiose | Glcα1→2Glc | 7.6 ± 0.3 | 100 |
| Nigerose | Glcα1→3Glc | 0.016 ± 0.001 | 0.22 |
| Trehalose | Glcα1→1αGlc | n.d. | - |
| Maltose | Glcα1→4Glc | n.d. | - |
| Isomaltose | Glcα1→6Glc | n.d. | - |
| Sucrose | Glcα1→2βFru | n.d. | - |
| Isomaltulose | Glcα1→6Fru | n.d. | - |
| Turanose | Glcα1→3Fru | n.d. | - |
| Melezitose | Glcα1→3Fruβ2→1Glc | n.d. | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Beul, E.; Jongbloet, A.; Franceus, J.; Desmet, T. Discovery of a Kojibiose Hydrolase by Analysis of Specificity-Determining Correlated Positions in Glycoside Hydrolase Family 65. Molecules 2021, 26, 6321. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26206321
De Beul E, Jongbloet A, Franceus J, Desmet T. Discovery of a Kojibiose Hydrolase by Analysis of Specificity-Determining Correlated Positions in Glycoside Hydrolase Family 65. Molecules. 2021; 26(20):6321. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26206321
Chicago/Turabian StyleDe Beul, Emma, Alana Jongbloet, Jorick Franceus, and Tom Desmet. 2021. "Discovery of a Kojibiose Hydrolase by Analysis of Specificity-Determining Correlated Positions in Glycoside Hydrolase Family 65" Molecules 26, no. 20: 6321. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26206321