
Kinetics of Phenolic Compounds Modification during Maize Flour Fermentation

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. pH, Titratable Acidity (TTA) and Total Soluble Solids (TSS)
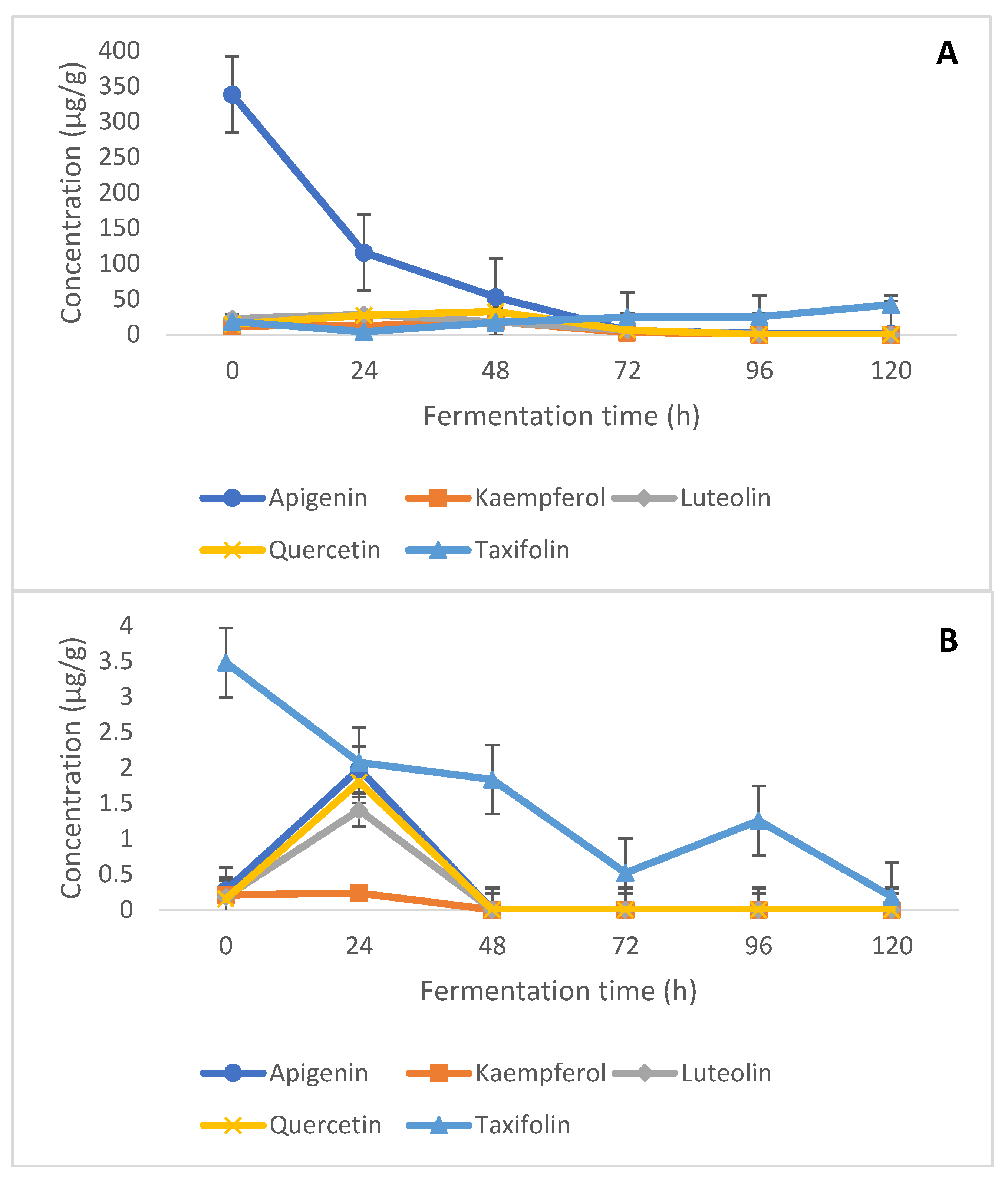
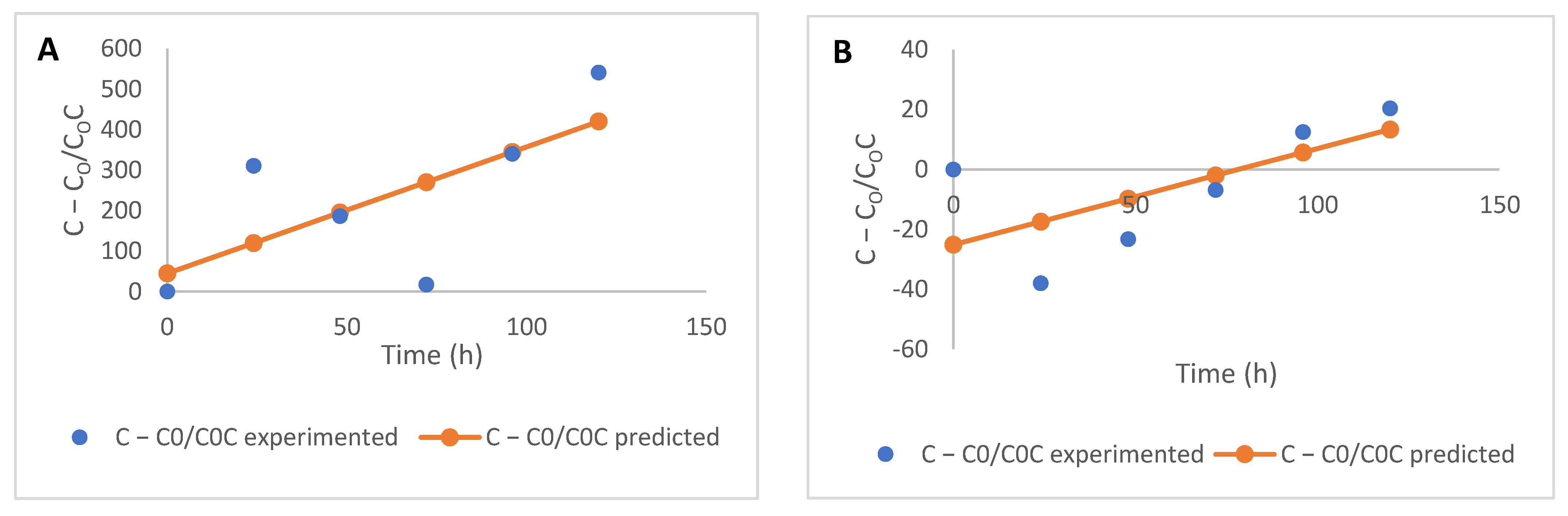
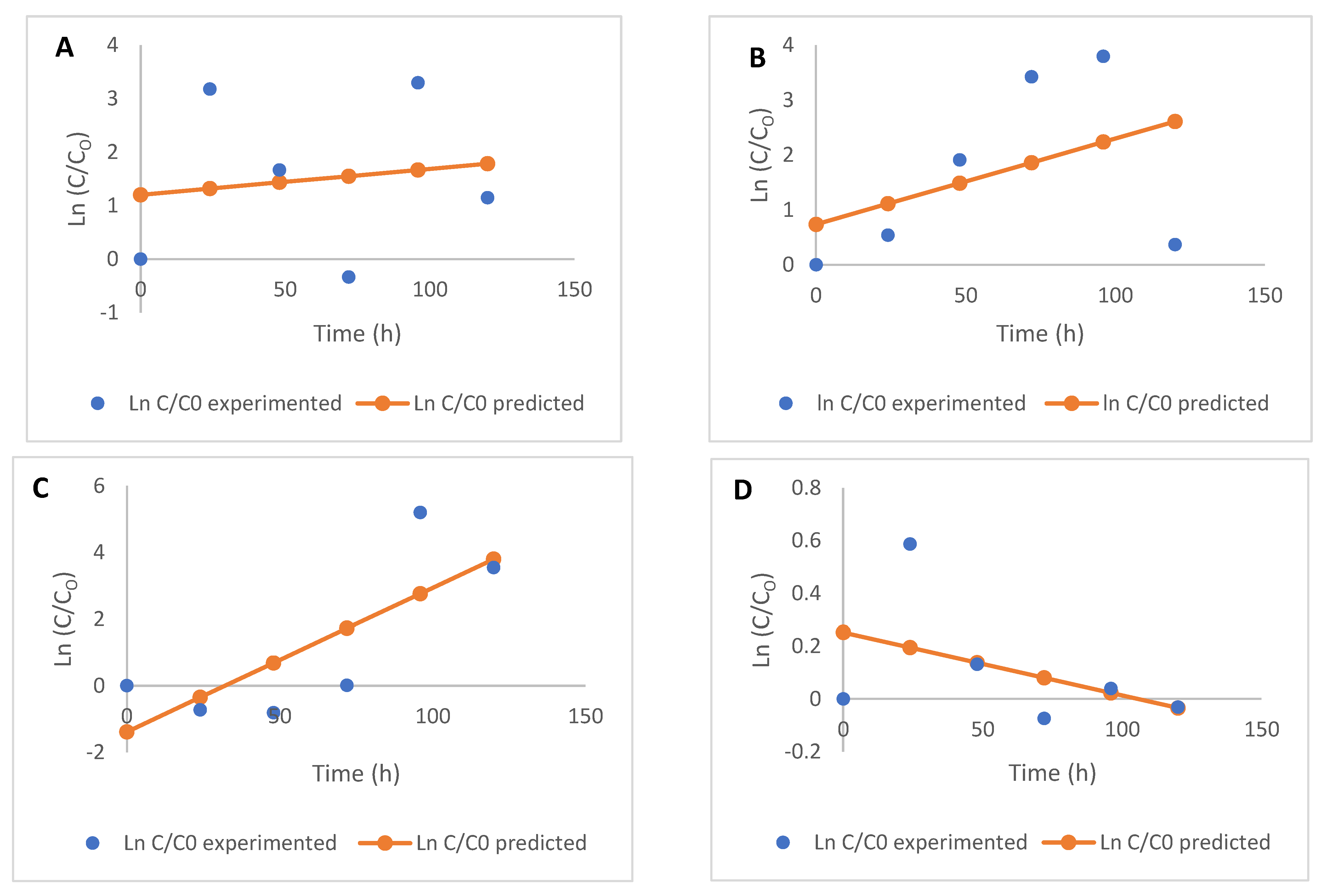
2.2. Free and Bound Flavonoids Contents and Kinetics of Modification
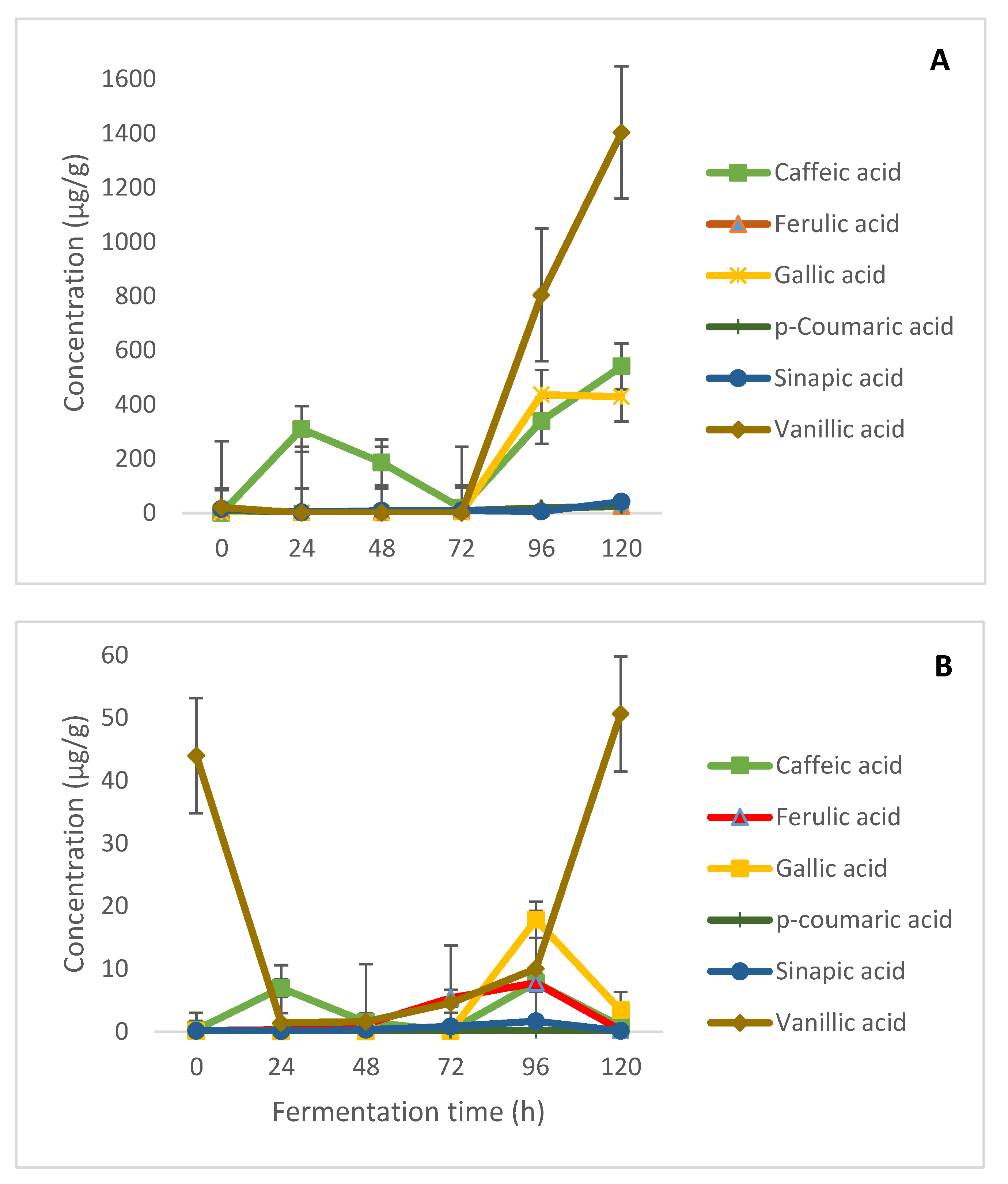
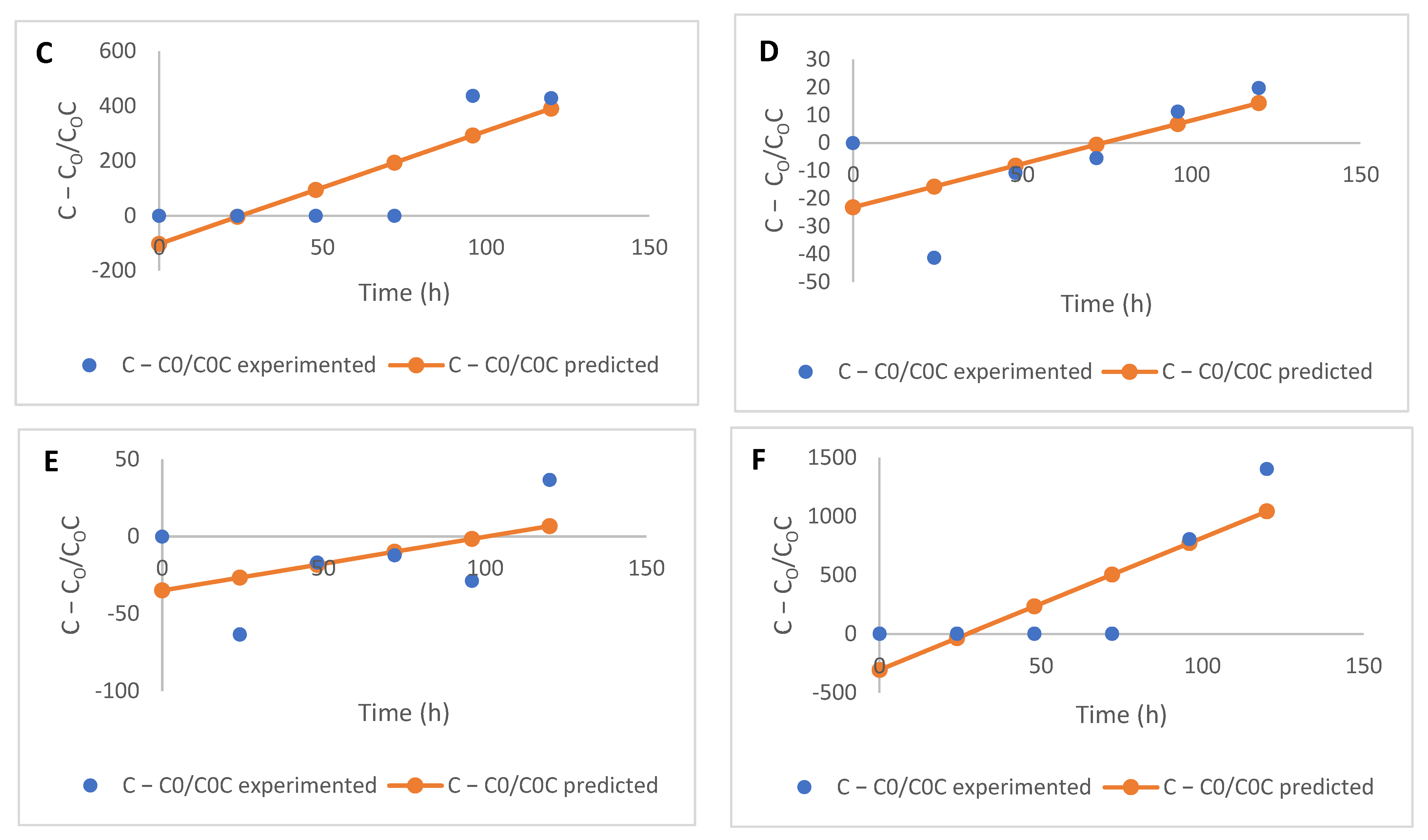
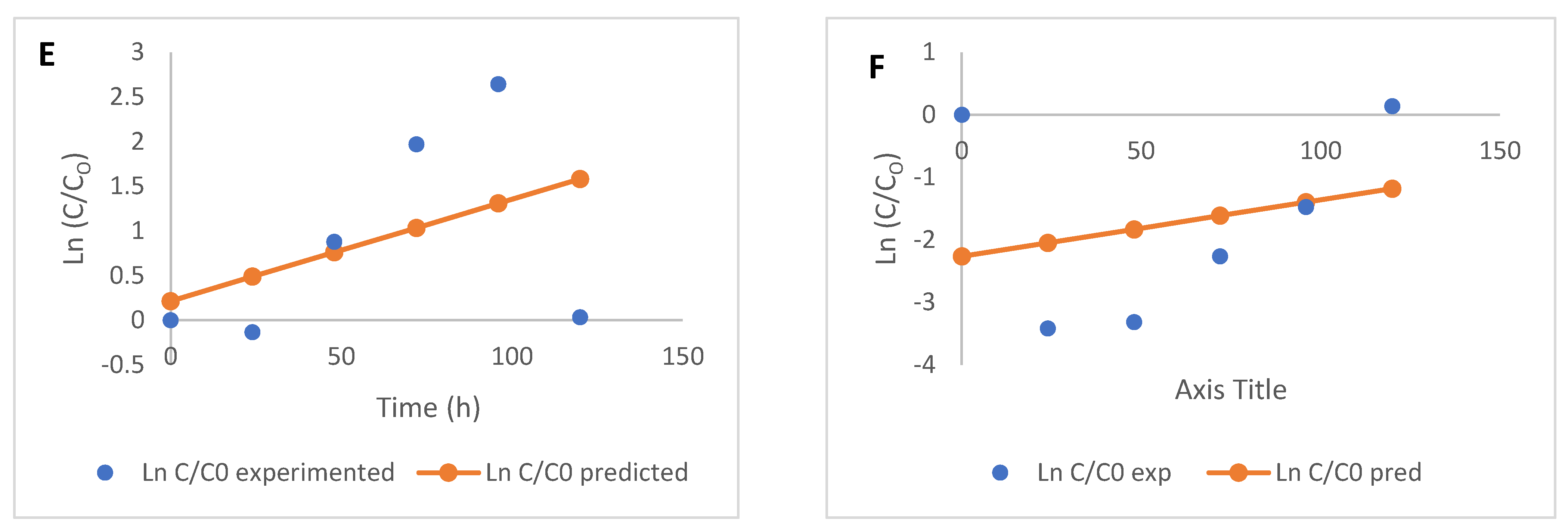
2.3. Free and Bound Phenolic Acids and Kinetics of Modification
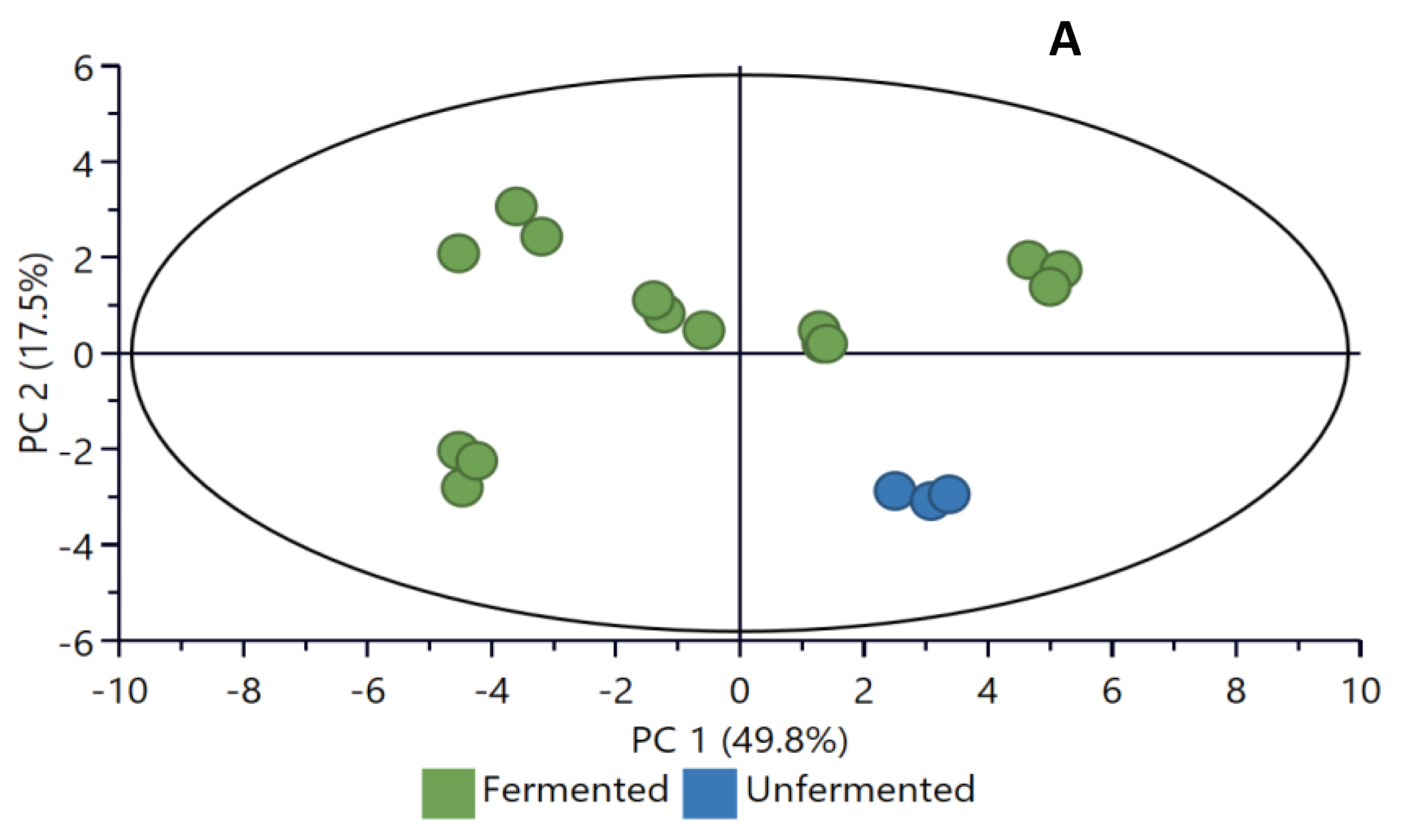
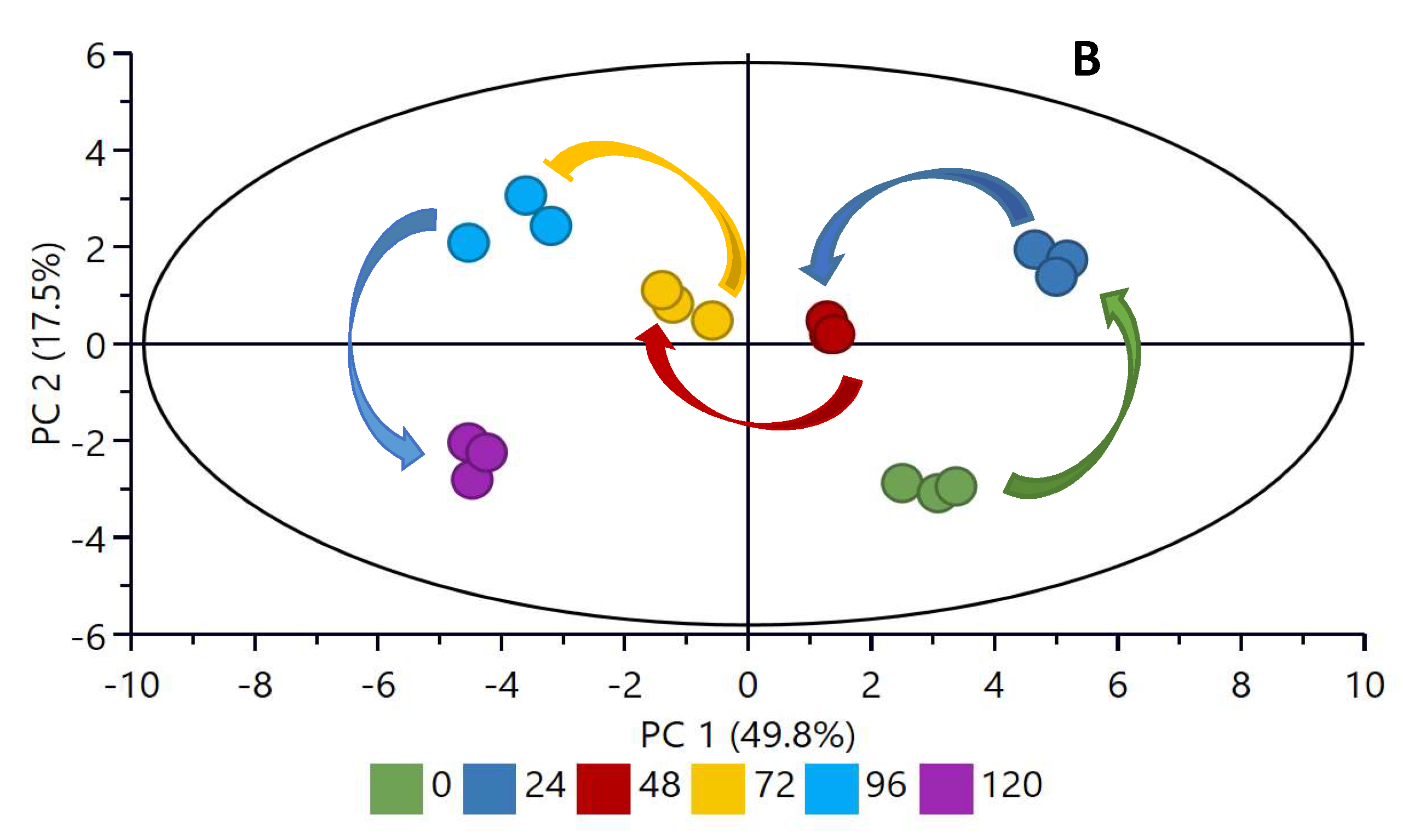
2.4. Principal Component Analysis
3. Materials and Methods
3.1. Fermentation
3.2. Total Soluble Solids (TSS), pH, and Titratable Acidity (TTA)
3.3. Extraction and Quantification of Free and Bound Phenolic Compounds
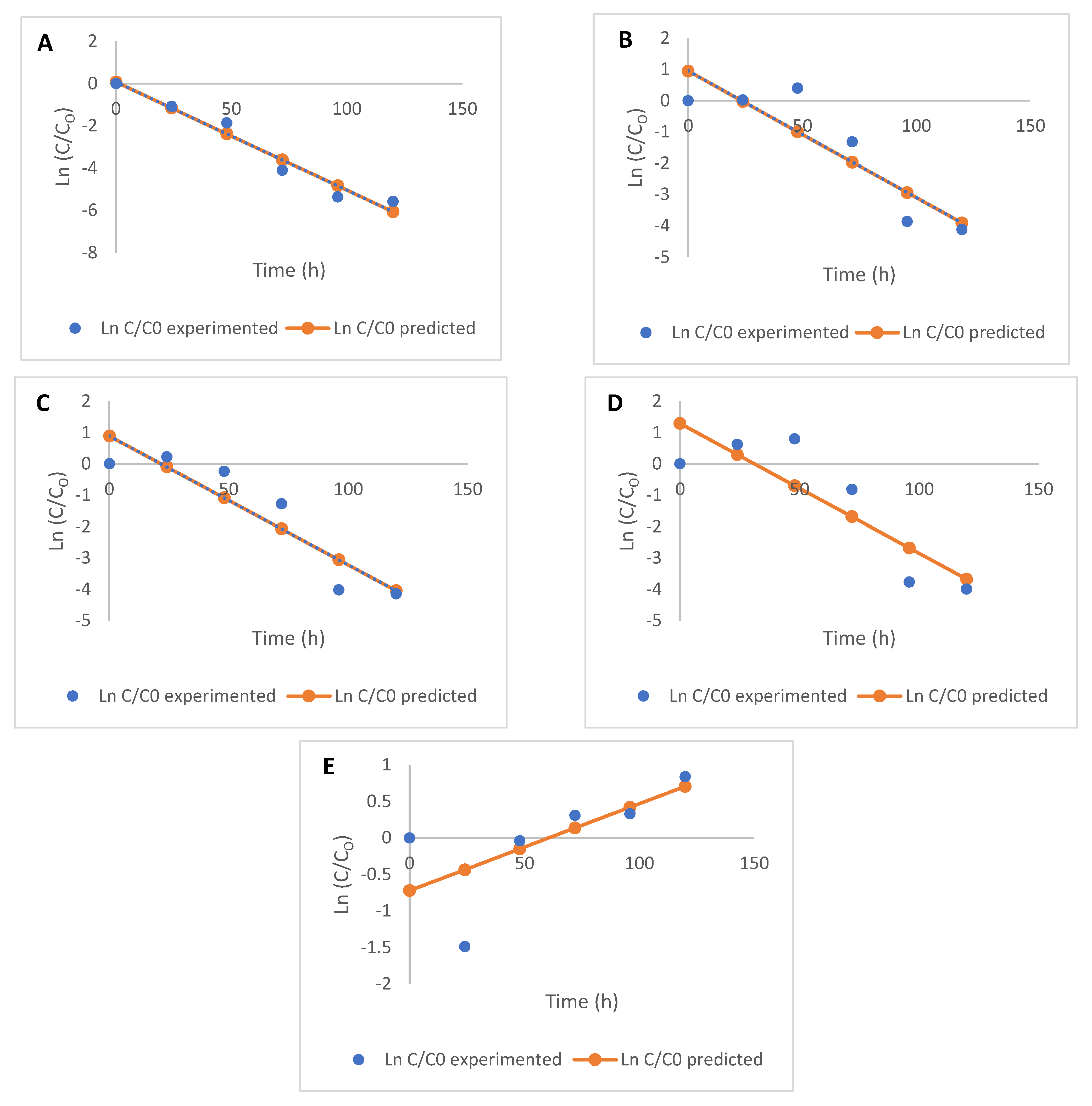
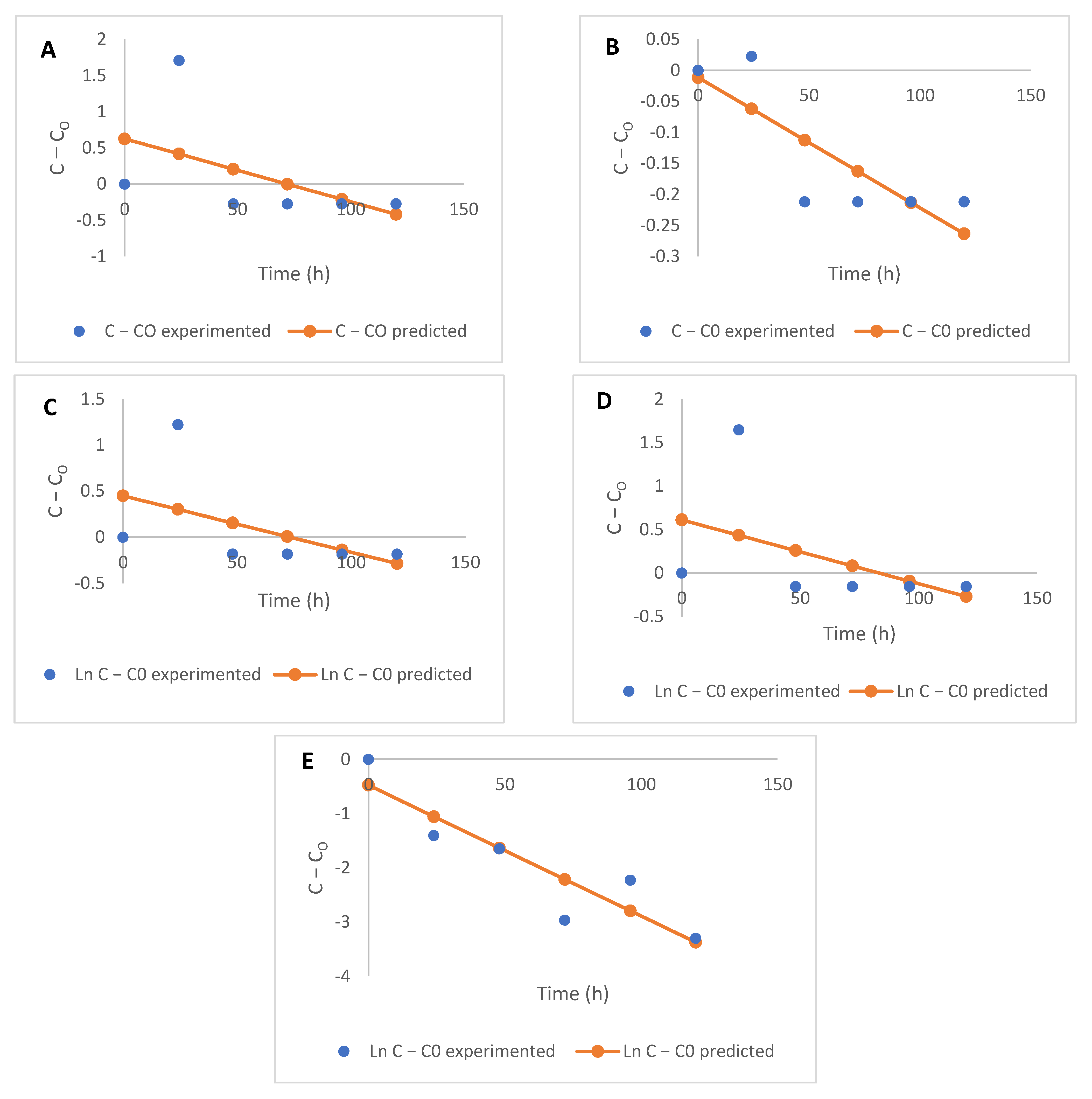
3.4. Kinetic Modelling
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Li, W.; Beta, T. Food sources of phenolics compounds. In Natural Products; Ramawat, K., Mérillon, J.M., Eds.; Springer: Berlin, Germany, 2013; pp. 2527–2558. [Google Scholar]
- Olatunde, O.O.; Benjakul, S.; Vongkamjan, K. Antioxidant and antibacterial properties of guava leaf extracts as affected by solvents used for prior dechlorophyllization. J. Food Biochem. 2018, 42, e12600. [Google Scholar] [CrossRef]
- Bonta, R.K. Dietary phenolic acids and flavonoids as potential anti-cancer agents: Current state of the art and future perspectives. Anti-Cancer Agents Med. Chem 2019, 20, 29–48. [Google Scholar] [CrossRef]
- Adebo, O.A.; Medina-Meza, I. Impact of fermentation on the phenolic compounds and antioxidant activity of whole cereal grains: A mini review. Molecules 2020, 25, 927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, W.; Zhang, P.; Ying, D.; Adhikari, B.; Fang, Z. Fermentation transforms the phenolic profiles and bioactivities of plant-based foods. Biotechnol. Adv. 2021, 49, 107763. [Google Scholar] [CrossRef] [PubMed]
- FAOSTAT. Crop Yields. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 8 June 2021).
- Nyirenda, H.; Mwangomba, W.; Nyirenda, E.M. Delving into possible missing links for attainment of food security in Central Malawi: Farmers’ perceptions and long term dynamics in maize (Zea mays L.) production. Heliyon 2021, 7, e07130. [Google Scholar] [CrossRef]
- Adebo, O.A.; Njobeh, P.B.; Adeboye, A.S.; Adebiyi, J.A.; Sobowale, S.S.; Ogundele, O.M.; Kayitesi, E. Advances in fermentation technology for novel food products. In Innovations in Technologies for Fermented Food and Beverage Industries; Panda, S., Shetty, P., Eds.; Springer: Berlin, Germany, 2018; pp. 71–87. [Google Scholar]
- Adebiyi, J.A.; Obadina, A.O.; Adebo, O.A.; Kayitesi, E. Fermented and malted millet products in Africa: Expedition from traditional/ethnic foods to industrial value added products. Crit Rev. Food Sci. Nutr. 2018, 58, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Adebo, O.A. African sorghum-based fermented foods: Past, current and future prospects. Nutrients 2020, 12, 1111. [Google Scholar] [CrossRef] [PubMed]
- van Boekel, M.A.J.S. On the pros and cons of Bayesian kinetic modeling in food science. Trends Food Sci. Technol. 2020, 99, 181–193. [Google Scholar] [CrossRef]
- Sekwati-Monang, B.; Gänzle, M.G. Microbiological and chemical characterization of ting, a sorghum-based sourdough product from Botswana. Int. J. Food Microbiol. 2011, 150, 115–121. [Google Scholar] [CrossRef]
- Edema, M.O. A modified sourdough procedure for non-wheat bread from maize meal. Food Bioproc. Technol. 2011, 4, 1264–1272. [Google Scholar] [CrossRef]
- Decimo, M.; Quattrini, M.; Ricci, G.; Fortina, M.G.; Brasca, M.; Silvetti, T.; Manini, F.; Erba, D.; Criscuoli, F.; Casiraghi, M.C. Evaluation of microbial consortia and chemical changes in spontaneous maize bran fermentation. AMB Express 2017, 7, 205. [Google Scholar] [CrossRef] [PubMed]
- Adebo, O.A.; Njobeh, P.B.; Adebiyi, J.A.; Kayitesi, E. Co-influence of fermentation time and temperature on physicochemical properties, bioactive components and microstructure of ting (a Southern African food) from whole grain sorghum. Food Biosci. 2018, 25, 118–127. [Google Scholar] [CrossRef]
- Yousif, N.E.; El-Tinay, A.H. Effect of fermentation on sorghum protein fractions and in vitro protein digestibility. Plant. Foods Human Nutr. 2001, 56, 175–182. [Google Scholar] [CrossRef]
- Chai, K.F.; Adzahan, N.M.; Karim, R.; Rukayadi, Y.; Ghazali, H.M. Effects of fermentation time and turning intervals on the physicochemical properties of rambutan (Nephelium lappaceum L.) fruit sweatings. Sains Malays. 2018, 47, 2311–2318. [Google Scholar]
- Padhye, V.W.; Salunkhe, D.K. Biochemical studies on black gram (Phaseolus mungo L.) III. Fermentation of black gram and rice blend and its influence on the in vitro protein digestibilities of proteins. J. Food Biochem. 1979, 2, 327–347. [Google Scholar] [CrossRef]
- El Tinay, A.H.; El Mahdi, Z.M.; El Soubki, A. Supplementation of fermented sorghum kisra bread with legume protein. J. Food Technol. 1985, 20, 679–687. [Google Scholar]
- Yousif, N.E.; El Tinay, A.H. Effect of fermentation on protein fractions and in vitro protein digestibility of maize. Food Chem. 2000, 70, 181–184. [Google Scholar] [CrossRef]
- Hansen, C.E.; del Olmo, M.; Burri, C. Enzyme activities in cocoa beans during fermentation. J. Sci Food Agric. 1998, 77, 273–281. [Google Scholar] [CrossRef]
- Pérez-Jiménez, J.; Torres, J.L. Analysis of nonextractable phenolic compounds in foods: The current state of the art. J. Sci Food Agric. 2011, 59, 12713–12724. [Google Scholar] [CrossRef] [PubMed]
- Righini, S.; Rodriguez, E.J.; Berosich, C.; Grotewold, E.; Casati, P.; Ferreyra, M.L.F. Apigenin produced by maize flavone synthase I and II protects plants against UV-B-induced damage. Plant. Cell Environ. 2019, 42, 495–508. [Google Scholar] [CrossRef]
- Bai, Y.; Findlay, B.; Maldonado, A.F.S.; Schieber, A.; Vederas, J.C.; Ganzle, M.G. Novel pyrano and vinylphenol adducts of deoxyanthocyanidins in sorghum sourdough. J. Agric. Food Chem. 2014, 62, 11536–11546. [Google Scholar] [CrossRef]
- Huynh, N.T.; Van Camp, J.; Smagghe, G.; Raes, K. Improved release and metabolism of flavonoids by steered fermentation processes: A review. Int. J. Mol. Sci. 2014, 15, 19369–19388. [Google Scholar] [CrossRef] [PubMed]
- Ali, F.; Rahul; Naz, F.; Jyoti, S.; Siddique, Y.H. Health functionality of apigenin: A review. Int. J. Food Prop. 2017, 20, 1197–1238. [Google Scholar] [CrossRef]
- Oladeji, B.S.; Akanbi, C.T.; Gbadamosi, S.O. Effects of fermentation on antioxidant properties of flours of a normal endosperm and quality protein maize varieties. J. Food Measur. Character. 2017, 11, 1148–1158. [Google Scholar] [CrossRef]
- Yang, L.; Allred, K.; Dykes, L.; Allred, C.; Awika, J.M. Enhanced action of apigenin and naringenin combination on estrogen receptor activation in non-malignant colonocytes: Implications on sorghum-derived phytoestrogens. Food Funct. 2015, 6, 749–755. [Google Scholar] [CrossRef] [PubMed]
- De Beer, D.; Tobin, J.; Walczak, B.; Van der Rijst, M.; Joubert, E. Phenolic composition of rooibos changes during simulated fermentation: Effect of endogenous enzymes and fermentation temperature on reaction kinetics. Food Res. Int. 2019, 121, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Karaaslan, M.; Yilmaz, F.M.; Cesur, Ö.; Vardin, H.; Ikinci, A.; Dalgiç, A.C. Drying kinetics and thermal degradation of phenolic compounds and anthocyanins in pomegranate arils dried under vacuum conditions. Int. J. Food Sci. Technol. 2014, 49, 595–605. [Google Scholar] [CrossRef]
- Turturică, M.; Stănciuc, N.; Bahrim, G.; Râpeanu, G. Effect of thermal treatment on phenolic compounds from plum (Prunus domestica) extracts–A kinetic study. J. Food Eng. 2016, 171, 200–207. [Google Scholar] [CrossRef]
- Rękas, A.; Ścibisz, I.; Siger, A.; Wroniak, M. The effect of microwave pretreatment of seeds on the stability and degradation kinetics of phenolic compounds in rapeseed oil during long-term storage. Food Chem. 2017, 222, 43–52. [Google Scholar] [CrossRef]
- Ali, A.; Chong, C.H.; Mah, S.H.; Abdullah, L.C.; Choong, T.S.Y.; Chua, B.L. Impact of storage conditions on the stability of predominant phenolic constituents and antioxidant activity of dried piper betle extracts. Molecules 2018, 23, 484. [Google Scholar] [CrossRef] [Green Version]
- Oyedeji, A.B.; Sobukola, O.P.; Henshaw, F.O.; Adegunwa, M.O.; Sanni, L.O.; Tomlins, K.I. Kinetics of mass transfer during deep fat frying of yellow fleshed cassava root slices. Heat Mass Transf. 2016, 52, 1061–1070. [Google Scholar] [CrossRef]
- Oyedeji, A.B.; Sobukola, O.A.; Green, E.; Adebo, O.A. Physical properties and water absorption kinetics of three varieties of Mucuna beans. Sci. Rep. 2021, 11, 5450. [Google Scholar] [CrossRef]
- Moore, J.; Cheng, Z.; Hao, J.; Guo, G.; Liu, J.-G.; Lin, C.; Yu, L. Effects of solid-state yeast treatment on the antioxidant properties and protein and fiber compositions of common hard wheat bran. J. Agric. Food Chem. 2007, 55, 10173–10182. [Google Scholar] [CrossRef]
- Starzyńska-Janiszewska, A.; Stodolaka, B.; Gómez- Caravaca, A.M.; Mickowska, B.; Martin-Garcia, B.; Byczyńskia, L. Mould starter selection for extended solid-state fermentation of quinoa. LWT Food Sci. Technol. 2019, 99, 213–237. [Google Scholar] [CrossRef]
- Svensson, L.; Sekwati-Monang, B.; Lutz, D.L.; Schieber, A.; Gänzle, M.G. Phenolic acids and flavonoids in nonfermented and fermented red sorghum (Sorghum bicolor (L.) Moench). J. Agric. Food Chem. 2010, 58, 9214–9220. [Google Scholar] [CrossRef] [PubMed]
- Hole, A.S.; Rud, I.; Grimmer, S.; Sigl, S.; Narvhus, J.; Sahlstrøm, S. Improved bioavailability of dietary phenolic acids in whole grain barley and oat groat following fermentation with probiotic Lactobacillus acidophilus, Lactobacillus johnsonii, and Lactobacillus reuteri. J. Agric. Food Chem. 2012, 60, 6369–6375. [Google Scholar] [CrossRef] [PubMed]
- Starzyńska-Janiszewska, A.; Stodolak, B.; Socha, R.; Mickowska, B.; Wywrocka-Gurgul, A. Spelt wheat tempe as a value-added whole-grain food product. LWT Food Sci. Technol. 2019, 113, 108250. [Google Scholar] [CrossRef]
- Zhang, L.; Gao, W.; Chen, X.; Wang, H. The effect of bioprocessing on the phenolic acid composition and antioxidant activity of wheat bran. Cereal Chem. 2014, 91, 255–261. [Google Scholar] [CrossRef]
- Kadiri, O. A review on the status of the phenolic compounds and antioxidant capacity of the flour: Effects of cereal processing. Int. J. Food Prop. 2017, 20, S798–S809. [Google Scholar] [CrossRef] [Green Version]
- Skrajda-Brdak, M.; Konopka, I.; Tańska, M.; Czaplicki, S. Changes in the content of free phenolic acids and antioxidative capacity of wholemeal bread in relation to cereal species and fermentation type. Eur. Food Res. Technol. 2019, 245, 2247–2256. [Google Scholar] [CrossRef] [Green Version]
- Vernhet, A.; Carrillo, S.; Rattier, A.; Verbaere, A.; Cheynier, V.; Nguela, J.M. Fate of anthocyanins and proanthocyanidins during the alcoholic fermentation of thermovinified red musts by different Saccharomyces cerevisiae strains. J. Agric. Food Chem. 2020, 68, 3615–3625. [Google Scholar] [CrossRef]
- Aguilar, N.; Albanell, E.; Miñarro, B.; Capellas, M. Chestnut flour sourdough for gluten-free bread making. Eur. Food Res. Technol. 2016, 242, 1795–1802. [Google Scholar] [CrossRef]
- Xiang, J.; Apea-Bah, F.B.; Ndolo, V.U.; Katundu, M.C.; Beta, T. Profile of phenolic compounds and antioxidant activity of finger millet varieties. Food Chem. 2019, 275, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Ravisankar, S.; Abegaz, K.; Awika, J.M. Structural profile of soluble and bound phenolic compounds in teff (Eragrostis tef) reveals abundance of distinctly different flavones in white and brown varieties. Food Chem. 2018, 265–274. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Time (h) | pH | TTA (g/kg) | TSS (°Brix) |
|---|---|---|---|
| 0 | 6.30 c ± 0.14 | 0.27 a ± 0.06 | 0.20 a ± 0 |
| 24 | 5.21 b ± 0.11 | 0.57 b ± 0.03 | 0.20 a ± 0.10 |
| 48 | 4.01 a ± 0.03 | 1.05 c ± 0.18 | 0.53 ab ± 0.25 |
| 72 | 3.88 a ± 0.11 | 1.65 d ± 0.22 | 0.50 ab ± 0.10 |
| 96 | 3.90 a ± 0.04 | 1.45 d ± 0.05 | 0.87 c ± 0.15 |
| 120 | 3.89 a ± 0.08 | 1.53 d ± 0.06 | 0.63 bc ± 0.06 |
| Free Phenolic Compounds | Bound Phenolic Compounds | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Zero Order | First Order | Second Order | Zero Order | First Order | Second Order | |||||||
| (mgh−1) | R2 | (h−1) | R2 | (mgh−1 g−1 h−1) | R2 | (mgh−1) | R2 | (h−1) | R2 | (mgh−1 g−1) | R2 | |
| Flavonoids | ||||||||||||
| Apigenin | −0.015 | 0.71 | −2.470 | 0.96 | −0.007 | 0.82 | −0.009 | 0.24 | −0.007 | 0.15 | −0.007 | 0.15 |
| Kaempferol | −0.040 | 0.62 | −0.134 | 0.79 | −6.655 | 0.74 | −0.002 | 0.66 | −0.0004 | 0.15 | −0.0002 | 0.15 |
| Luteolin | −0.041 | 0.85 | −0.248 | 0.84 | −13.268 | 0.73 | −0.006 | 0.24 | −0.007 | 0.15 | −0.005 | 0.15 |
| Quercetin | −0.214 | 0.48 | −0.041 | 0.74 | −0.214 | 0.48 | −0.007 | 0.20 | −0.009 | 0.15 | −0.006 | 0.15 |
| Taxifolin | 0.012 | 0.66 | 0.227 | 0.45 | 0.538 | 0.40 | −0.024 | 0.82 | −0.021 | 0.76 | −0.432 | 0.60 |
| Phenolic Acids | ||||||||||||
| Caffeic Acid | 0.046 | 0.46 | 3.125 | 0.44 | 3.125 | 0.46 | −0.005 | 0.004 | −0.005 | 0.02 | −0.006 | 0.005 |
| Ferulic Acid | 0.146 | 0.44 | 0.012 | 0.53 | 0.321 | 0.43 | 0.032 | 0.20 | −0.016 | 0.18 | 0.033 | 0.20 |
| Gallic Acid | 0.056 | 0.68 | 4.107 | 0.68 | 4.112 | 0.68 | 0.083 | 0.28 | 0.043 | 0.59 | 0.085 | 0.28 |
| p-Coumaric Acid | 0.012 | 0.60 | 0.139 | 0.47 | 0.312 | 0.44 | −0.0004 | 0.19 | −0.002 | 0.19 | −0.0007 | 0.19 |
| Sinapic Acid | 0.009 | 0.30 | 0.174 | 0.20 | 0.384 | 0.22 | 0.006 | 0.20 | 0.0114 | 0.19 | 0.007 | 0.20 |
| Vanillic Acid | −0.038 | 0.69 | 11.110 | 0.70 | 11.227 | 0.70 | 0.074 | 0.02 | 0.009 | 0.07 | 5.160 | 0.15 |
| Compound | Retention Time (min) | Wavelength (nm) |
|---|---|---|
| Flavonoids | ||
| Apigenin | 11.324 | 336 |
| Kaempferol | 11.347 | 366 |
| Luteolin | 10.670 | 348 |
| Quercetin | 10.597 | 371 |
| Taxifolin | 9.300 | 290 |
| Phenolic acids | ||
| Caffeic acid | 8.597 | 323 |
| Ferulic acid | 9.447 | 322 |
| Gallic acid | 8.528 | 271 |
| p-Coumaric acid | 9.348 | 309 |
| Sinapic acid | 9.016 | 319 |
| Vanillic acid | 8.713 | 261 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adebo, O.A.; Oyedeji, A.B.; Adebiyi, J.A.; Chinma, C.E.; Oyeyinka, S.A.; Olatunde, O.O.; Green, E.; Njobeh, P.B.; Kondiah, K. Kinetics of Phenolic Compounds Modification during Maize Flour Fermentation. Molecules 2021, 26, 6702. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216702
Adebo OA, Oyedeji AB, Adebiyi JA, Chinma CE, Oyeyinka SA, Olatunde OO, Green E, Njobeh PB, Kondiah K. Kinetics of Phenolic Compounds Modification during Maize Flour Fermentation. Molecules. 2021; 26(21):6702. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216702
Chicago/Turabian StyleAdebo, Oluwafemi Ayodeji, Ajibola Bamikole Oyedeji, Janet Adeyinka Adebiyi, Chiemela Enyinnaya Chinma, Samson Adeoye Oyeyinka, Oladipupo Odunayo Olatunde, Ezekiel Green, Patrick Berka Njobeh, and Kulsum Kondiah. 2021. "Kinetics of Phenolic Compounds Modification during Maize Flour Fermentation" Molecules 26, no. 21: 6702. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216702