
Eucalyptus gunnii and Eucalyptus pulverulenta ‘Baby Blue’ Essential Oils as Potential Natural Herbicides

 ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
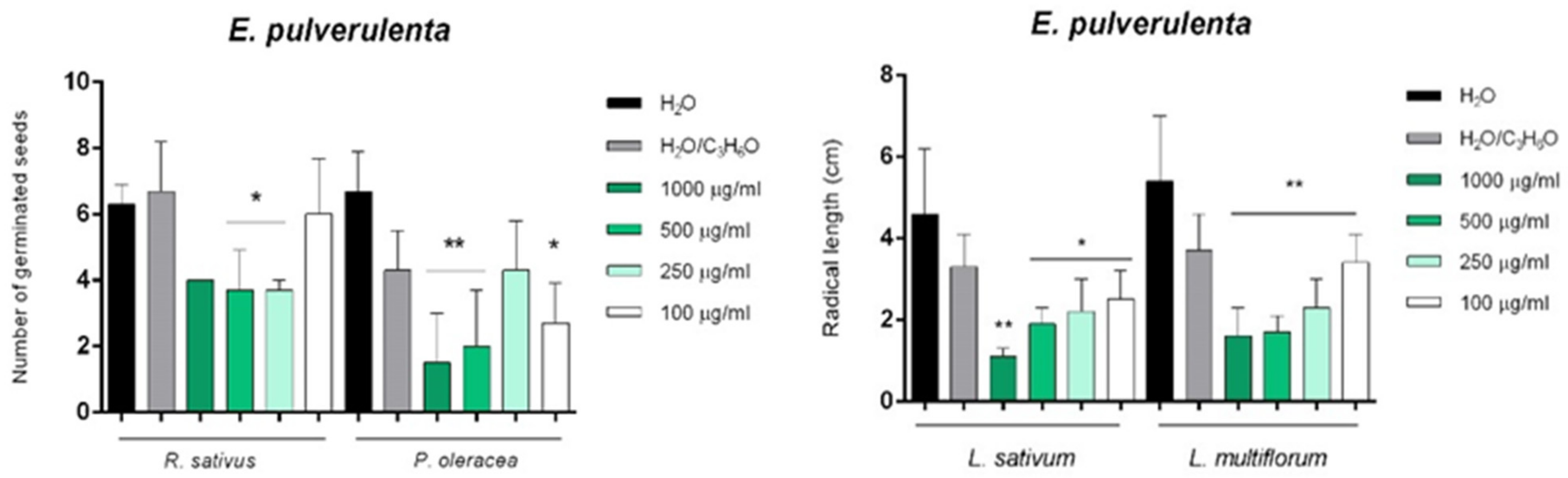
2. Results and Discussion
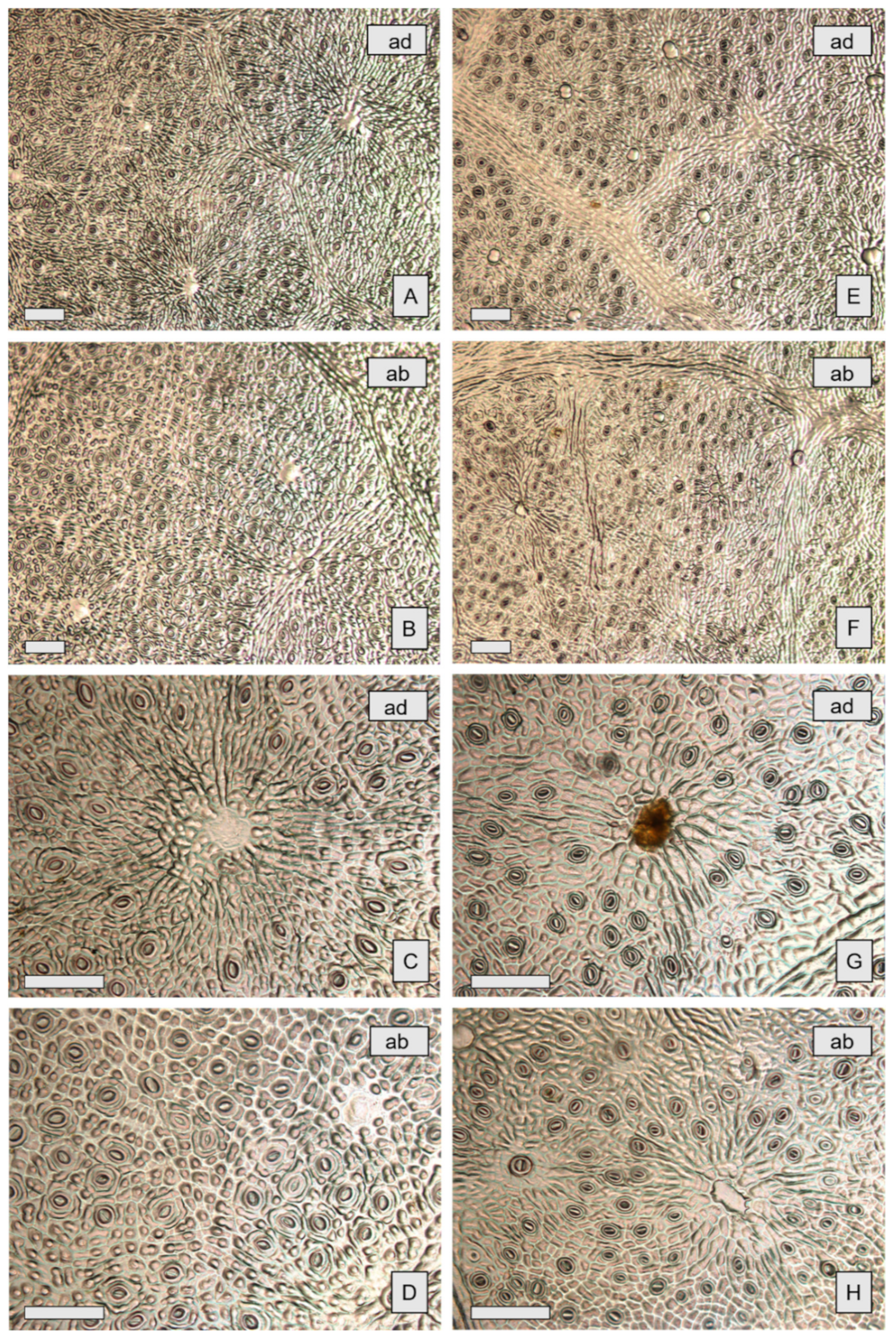
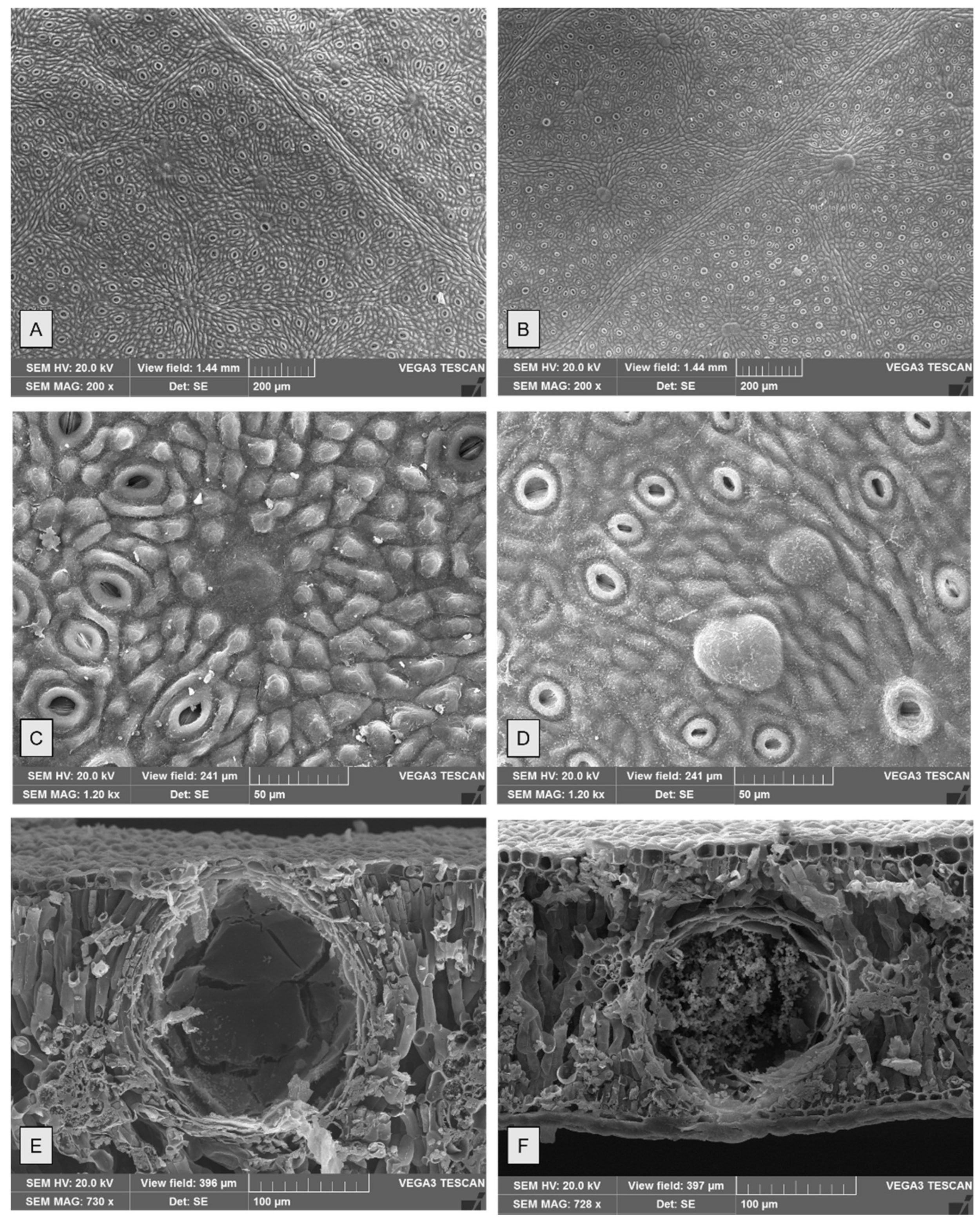
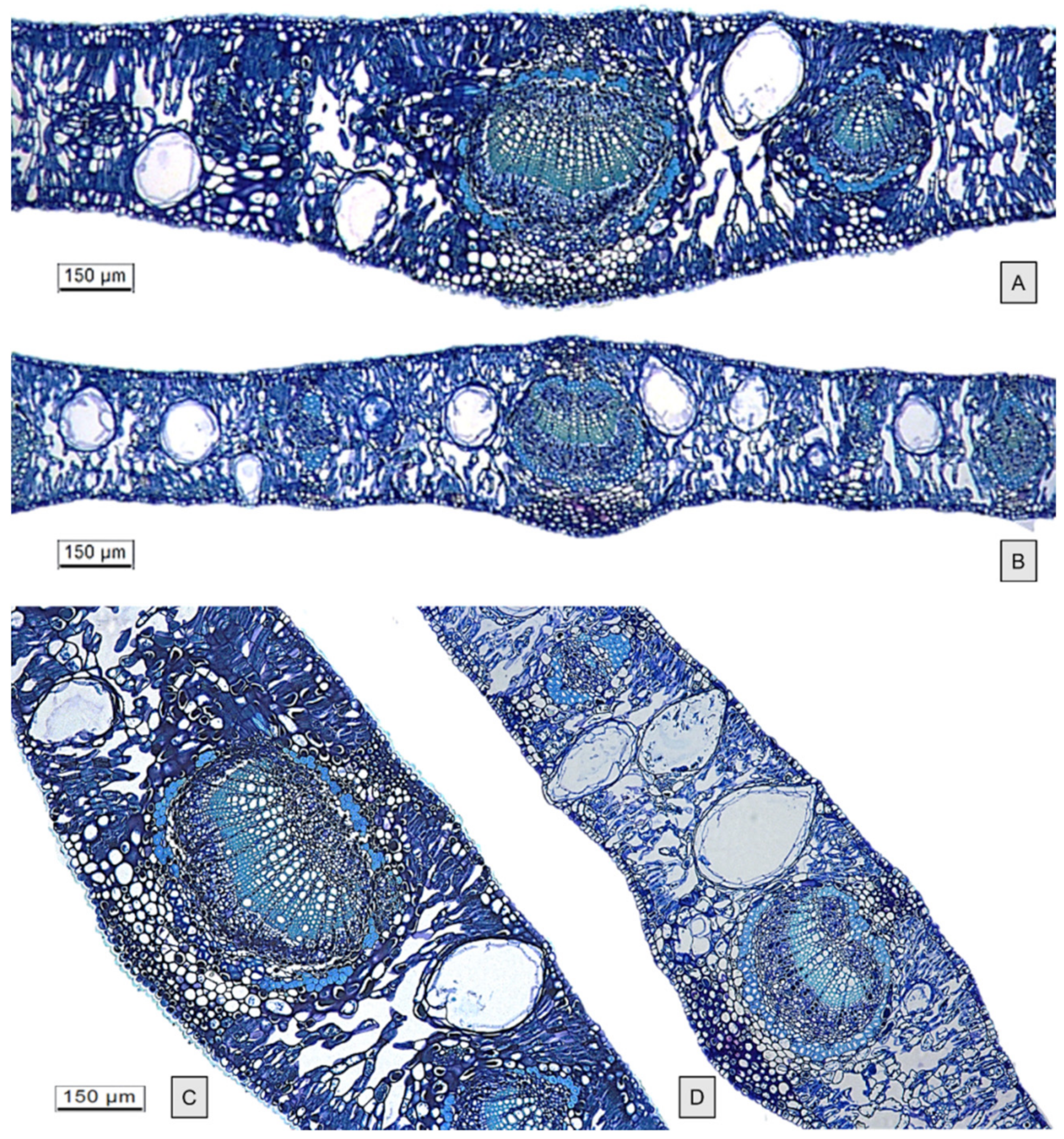
2.1. Micromorphological Studies
2.2. Chemical Composition of Essential Oils
2.3. Phytotoxic and Anti-α-Amylase Activity
2.4. Toxicity
3. Materials and Methods
3.1. Standards and Reagents
3.2. Plant Material
3.3. Extraction of Essential Oils
3.4. Micromorfological Analyses
3.5. GC-FID Analysis
3.6. GC/MS Analysis
3.7. Identification of the Essential Oil Components
3.8. Phytotoxic Activity
3.9. α-Amylase Inhibitory Assay
3.10. Brine Shrimp Lethality Assay
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Dhakad, A.K.; Pandey, V.V.; Beg, S.; Rawat, J.M.; Singh, A. Biological, medicinal and toxicological significance of Eucalyptus leaf essential oil: A review. J. Sci. Food Agric. 2018, 98, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Tomé, M.; Almeida, M.H.; Barreiro, S.; Branco, M.; Deus, E.; Pinto, G.; Silva, J.; Soares, P.; Rodríguez-Soalleir, R. Opportunities and challenges of Eucalyptus plantations in Europe: The Iberian Peninsula experience. Eur. J. For. Res. 2021, 140, 489–510. [Google Scholar] [CrossRef]
- Floris, I.; Cocco, A.; Buffa, F.; Mannu, R.; Satta, A. Insect pests of Eucalyptus plantations in Sardinia (Italy). J. Zool. 2018, 101, 61–71. [Google Scholar] [CrossRef]
- Campisi, E.; Mancianti, F.; Pini, G.; Faggi, E.; Gargani, G. Investigation in central Italy of the possible association between Cryptococcus neoformans var. Gattii and Eucalyptus camaldulensis. Eur. J. Epidemiol. 2003, 18, 357–362. [Google Scholar] [CrossRef]
- Forrest, M. The Performance of a Eucalyptus Gunnii Cut Foliage Plantation over 7 Years. Irish J. Agric. Food Res. 2002, 41, 235–245. Available online: http://0-www-jstor-org.brum.beds.ac.uk/stable/25562467 (accessed on 4 October 2021).
- Pacifici, S.; Ferrante, A.; Mensuali, A.; Serra, G. Postharvest physiology and technology of cut Eucalyptus branches: A review. Agr. Med. 2007, 137, 124–131. [Google Scholar]
- Wirthensohn, M.G.; Collins, G.; Jones, G.P.; Sedgley, M. Variability in waxiness of Eucalyptus gunnii foliage for floriculture. Sci. Hortic. 1999, 82, 279–288. [Google Scholar] [CrossRef]
- Garcia, C.; Montero, G.; Coronado, O.M.; Valdez Salaz, B.; Stoytcheva, M.; Rosas, N.; Ricardo, T.; Sagaste, C. Valorization of Eucalyptus Leaves by Essential Oil Extraction as an Added Value Product in Mexico. Waste Biomass Valor. 2017, 8, 1187–1197. [Google Scholar] [CrossRef]
- Mediavilla, I.; Guillamón, E.; Ruiz, A.; Estepan, L.S. Essential Oils from Residual Foliage of Forest Tree and Shrub Species: Yield and Antioxidant Capacity. Molecules 2021, 26, 3257. [Google Scholar] [CrossRef] [PubMed]
- Smeriglio, A.; Trombetta, D.; Cornara, L.; Valussi, M.; De Feo, V.; Caputo, L. Characterization and Phytotoxicity Assessment of Essential Oils from Plant Byproducts. Molecules 2019, 24, 2941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemke, N.; Murawski, A.; Schmied-Tobies, M.I.H.; Rucic, E.; Hoppe, H.W.; Conrad, A.; Kolossa-Gehring, M. Glyphosate and aminomethylphosphonic acid (AMPA) in urine of children and adolescents in Germany—Human biomonitoring results of the German Environmental Survey 2014–2017 (GerES V). Environ. Int. 2021, 156, 106769. [Google Scholar] [CrossRef]
- Meftaul, I.M.; Venkateswarlu, K.; Dharmarajan, R.; Annamalai, P.; Asaduzzaman, M.; Parven, A.; Megharaj, M. Controversies over human health and ecological impacts of glyphosate: Is it to be banned in modern agriculture? Environ. Pollut. 2020, 263, 114372. Available online: https://0-www-sciencedirect-com.brum.beds.ac.uk/science/article/pii/S0269749119368265 (accessed on 4 October 2021). [CrossRef]
- Setia, N.; Batish, D.R.; Singh, H.P.; Kohli, R.K. Phytotoxicity of volatile oil from Eucalyptus citriodora against some weedy species. J. Environ. Biol. 2007, 28, 63–66. [Google Scholar]
- Kordali, S.; Cakir, A.; Sutay, S. Inhibitory Effects of Monoterpenes on Seed Germination and Seedling Growth. J. Biosci. 2007, 62, 207–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batish, D.; Singh, H.P.; Kohli, R.; Kaur, S. Eucalyptus essential oil as natural pesticide. For. Ecol. Manag. 2008, 256, 2166–2174. [Google Scholar] [CrossRef]
- Rasaeifar, M.; Hosseini, N.; Asl, N.; Zandi, P.; Aghdam, A. Allelopathic effect of Eucalyptus globulus’ essential oil on seed germination and seedling establishment of Amaranthus blitoides and Cyndon dactylon. Trakia J. Sci. 2013, 11, 73–81. [Google Scholar]
- Kanatas, P. Potential role of Eucalyptus spp. and Acacia spp. allelochemicals in weed management. Chil. J. Agric. Res. 2020, 80, 452–458. [Google Scholar] [CrossRef]
- Puig, C.; Revilla, P.; Barreal, M.E.; Reigosa, R.M.; Pedrol, N. On the suitability of Eucalyptus globulus green manure for field weed control. Crop Prot. 2019, 121, 57–65. [Google Scholar] [CrossRef]
- Vaid, S.; Batish, D.; Singh, D.H.; Kohli, R. Phytotoxic effect of eugenol towards two weedy species. Bioscan 2010, 5, 339–341. [Google Scholar]
- Ibáñez, M.D.; Blázquez, M.A. Phytotoxic Effects of Commercial Eucalyptus citriodora, Lavandula angustifolia, and Pinus sylvestris Essential Oils on Weeds, Crops, and Invasive Species. Molecules 2019, 24, 2847. [Google Scholar] [CrossRef] [Green Version]
- Verdeguer, M.; Blázquez, M.; Boira, H. Phytotoxic effects of Lantana camara, Eucalyptus camaldulensis and Eriocephalus africanus essential oils in weeds of Mediterranean summer crops. Biochem. Syst. Ecol. 2009, 37, 362–369. [Google Scholar] [CrossRef]
- Batish, D.R.; Setia, N.; Singh, H.P.; Kohli, R.K. Phytotoxicity of lemon-scented Eucalypt oil and its potential use as a bioherbicide. Crop Prot. 2004, 23, 1209–1214. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Nidhi, S.; Shalinder, K.; Kohli, R.K. Chemical composition and inhibitory activity of essential oil from decaying leaves of Eucalyptus citriodora. Z. Naturforsch. 2006, 61, 52–56. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.; Kaur, S.; Arora, K.; Kohli, R. α-Pinene Inhibits Growth and Induces Oxidative Stress in Roots. Ann. Bot. 2007, 98, 1261–1269. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; An, M.; Wu, H.; Stanton, R.; Lemerle, D. Chemistry and bioactivity of Eucalyptus essential oils. Allelopath. J. 2010, 25, 313–330. [Google Scholar]
- WHO. Quality Control Methods for Herbal Materials; WHO Press: Valletta, Malta, 2011; Available online: https://apps.who.int/iris/handle/10665/44479 (accessed on 4 October 2021).
- Alamgir, A.N.M. Microscopy in Pharmacognosy. In Therapeutic Use of Medicinal Plants and Their Extracts: Volume 1. Progress in Drug Research; Springer: Cham, Swizerland, 2017; Volume 73, Available online: https://0-doi-org.brum.beds.ac.uk/10.1007/978-3-319-63862-1_11 (accessed on 4 October 2021). [CrossRef]
- Santos, L.; Thadeo, M.; Iarema, L.; Meira, R.M.; Ferreira, F. Foliar anatomy and histochemistry in seven species of Eucalyptus. Rev. Arvore 2008, 32, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Migacz, I.; Raman, V.; Nisgoski, S.; Muniz, G.; Manfron, J.; Farago, P.; Khan, I.; Raeski, P. Comparative leaf morpho-anatomy of six species of Eucalyptus cultivated in Brazil. Rev. Bras. Farmacogn. 2018, 28, 273–281. [Google Scholar] [CrossRef]
- Soliman, F.; Fathy, M.; Salama, M.; Saber, F. Botanical study of Eucalyptus cinerea F. Muell. ex Benth. Cultivated in Egypt. World J. Pharm. Pharmaceut. Sci. 2015, 4, 310–328. [Google Scholar]
- King, D.; Gleadow, R.; Woodrow, I. Regulation of oil accumulation in single glands of Eucalyptus polybractea. New Phytol. 2006, 172, 440–451. [Google Scholar] [CrossRef] [PubMed]
- Brooker, M.I.H.; Nicolle, D. Atlas of Leaf Venation and Oil Gland Patterns in the Eucalypts; Csiro Publishing: Collingwood, Australia, 2013. [Google Scholar]
- Lucia, A.; Licastro, S.; Zerba, E.; Masuh, H. Yield, chemical composition, and bioactivity of essential oils from 12 species of Eucalyptus on Aedes aegypti larvae. Entomol. Exp. Appl. 2008, 129, 107–114. [Google Scholar] [CrossRef]
- Bugarin, D.; Grbović, S.; Orčič, D.; Mitić-Ćulafić, D.; Knežević-Vukčević, J.; Mimica-Dukić, N. Essential oil of Eucalyptus gunnii Hook. as a novel source of antioxidant, antimutagenic and antibacterial agents. Molecules 2014, 19, 19007–19020. [Google Scholar] [CrossRef] [PubMed]
- Caputo, L.; Smeriglio, A.; Trombetta, D.; Cornara, L.; Trevena, G.; Valussi, M.; Nazzaro, F. Chemical composition and biological activities of the essential oils of Leptospermum petersonii and Eucalyptus gunnii. Front. Microbiol. 2020, 11, 409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elaissi, A.; Salah, K.H.; Mabrouk, S.; Larbi, K.M.; Chemli, R.; Harzallah-Skhiri, F. Antibacterial activity and chemical composition of 20 Eucalyptus species’ essential oils. Food Chem. 2011, 129, 1427–1434. [Google Scholar] [CrossRef]
- Brophy, J.J.; Lassak, E.V.; Toia, R.F. The steam volatile leaf oil of Eucalyptus pulverulenta. Planta Med. 1985, 51, 170–171. [Google Scholar] [CrossRef]
- Zrira, S.; Bessiere, J.M.; Menut, C.; Elamrani, A.; Benjilali, B. Chemical composition of the essential oil of nine Eucalyptus species growing in Morocco. Flavour Frag. J. 2004, 19, 172–175. [Google Scholar] [CrossRef]
- Ieri, F.; Cecchi, L.; Giannini, E.; Clemente, C.; Romani, A. GC-MS and HS-SPME-GC× GC-TOFMS determination of the volatile composition of essential oils and hydrosols (By-products) from four Eucalyptus species cultivated in Tuscany. Molecules 2019, 24, 226. [Google Scholar] [CrossRef] [Green Version]
- Topiar, M.; Sajfrtova, M.; Pavela, R.; Machalova, Z. Comparison of fractionation techniques of CO2 extracts from Eucalyptus globulus–Composition and insecticidal activity. J. Supercri. Fluids 2015, 97, 202–210. [Google Scholar] [CrossRef]
- Ramezani, S.; Saharkhiz, M.J.; Ramezani, F.; Fotokian, M.H. Use of essential oils as bioherbicides. J. Essent. Oil-Bear. Plants 2008, 11, 319–327. [Google Scholar] [CrossRef]
- Kaur, S.; Pal Singh, H.; Batish, D.; Kumar Kholi, R. Role of monoterpenes in Eucalyptus communities. Curr. Bioact. Compd. 2012, 8, 101–107. [Google Scholar] [CrossRef]
- Kohli, R.K.; Batish, D.R.; Singh, H.P. Eucalypt oils for the control of Parthenium (Parthenium hysterophorus L.). Crop Prot. 1998, 17, 122–199. [Google Scholar] [CrossRef]
- Ibáñez, M.D.; Blázquez, M.A. Phytotoxicity of Essential Oils on Selected Weeds: Potential Hazard on Food Crops. Plants 2018, 7, 79. [Google Scholar] [CrossRef] [Green Version]
- Romagni, J.G.; Allen, S.N.; Dayan, F.E. Allelopathic effects of volatile cineoles on two weedy plant species. J. Chem. Ecol. 2000, 26, 303–313. [Google Scholar] [CrossRef]
- Kaur, S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Chemical characterization and allelopathic potential of volatile oil of Eucalyptus tereticornis against Amaranthus viridis. J. Plant Interact. 2011, 6, 297–302. [Google Scholar] [CrossRef]
- Murtaza, G.; Asghar, R. α-Amylase activities during seed development and germination in pea (Pisum sativum L.) treated with salicylic acid. Pak. J. Bot. 2012, 44, 1823–1829. [Google Scholar]
- Danabas, D.; Ates, M.; Tastan, B.E.; Cimen, I.C.C.; Unal, I.; Aksu, O.; Kutlu, B. Effects of Zn and ZnO nanoparticles on Artemia salina and Daphnia magna organisms: Toxicity, accumulation and elimination. Sci. Total Environ. 2020, 711, 134869. [Google Scholar] [CrossRef]
- Council of Europe. European Pharmacopeia, 5th ed.; Council of Europe: Strasbourg, France, 2004; Volume I, pp. 217–218. [Google Scholar]
- Rogiers, S.; Hardie, W.J.; Smith, J.P. Stomatal density of grapevine leaves (Vitis vinifera L.) responds to soil temperature and atmospheric carbon dioxide. Aust. J. Grape Wine Res. 2011, 17, 147–152. [Google Scholar] [CrossRef]
- O’Brien, T.P.; McCully, M.E. The Study of Plant Structure: Principles and Selected Methods; Termocarphy: Melbourne, Australia, 1981. [Google Scholar]
- Chieco, C.; Rotondi, A.; Morrone, L.; Rapparini, F.; Baraldi, R. An ethanol-based fixation method for anatomical and micromorphological characterization of leaves of various tree species. Biotech. Histochem. 2013, 88, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Pathan, A.K.; Bond, J.; Gaskin, R.E. Sample preparation for scanning electron microscopy of plant surfaces—Horses for courses. Micron 2008, 39, 1049–1061. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Jennings, W.; Shibamoto, T. Qualitative Analysis of Flavour and Fragrance Volatiles by Glass Capillary Gas Chromatography; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicone and Carbowax 20M phases. J. Chromatogr. 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Goodner, K.L. Practical retention index models of OV-101, DB-1, DB-5, and DB-Wax for flavor and fragrance compounds. LWT Food Sci. Technol. 2008, 41, 951–958. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectroscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- McLafferty, F.W. Wiley Registry of Mass Spectral Data, with NIST Spectral Data CD Rom, 7th ed.; John Wiley & Sons: New York, NY, USA, 1998. [Google Scholar]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.; Storms, R.; Tsang, A. A quantitative starch-iodine method for measuring alpha-amylase and glucoamylase activities. Anal. Biochem. 2006, 351, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Sudha, P.; Zinjarde, S.S.; Bhargava, S.Y.; Kumar, A.R. Potent α-amylase inhibitory activity of Indian Ayurvedic medicinal plants. BMC Complement. Altern. Med. 2011, 11, 5. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Oil Gland Mean Density (Glands cm−2) (±S.D.) | Oil Gland Mean Dimensions (μm) (±S.D.) | ||
|---|---|---|---|---|
| Abaxial | Adaxial | Height | Width | |
| E. gunnii | 213 (±66) | 416 (±116) | 199 (±34) | 161 (±31) |
| E. pulverulenta | 750 (±96) | 1321 (±176) | 188 (±32) | 142 (±26) |
| RT | Compound | % | KI a | KI b | Identification c | ||
|---|---|---|---|---|---|---|---|
| EG | EP | ||||||
| 1 | 5.148 | 1-Methyl-1,3-cyclohexadiene | t | 759 | 1183 | 1,2 | |
| 2 | 5.267 | 3-Methyl-2-buten-1-ol | t | 761 | 1328 | 1,2 | |
| 3 | 6.246 | n-Octane | t | 773 | 1,2 | ||
| 4 | 7.299 | (2Z)-Hexenol | t | 787 | 1,2 | ||
| 5 | 12.004 | Santolina triene | t | 848 | 1043 | 1,2 | |
| 6 | 13.849 | α-Pinene | 13.1 | 4.8 | 872 | 1028 | 1,2,3 |
| 7 | 14.036 | Camphene | 0.1 | 0.1 | 874 | 1075 | 1,2,3 |
| 8 | 16.071 | β-Pinene | 0.3 | 0.5 | 901 | 1120 | 1,2,3 |
| 9 | 16.945 | dehydro-1,8 Cineole | t | 912 | 1095 | 1,2 | |
| 10 | 17.952 | Myrcene | 0.3 | 0.4 | 925 | 1173 | 1,2,3 |
| 11 | 18.695 | δ-2-Carene | 0.1 | 0.2 | 934 | 1146 | 1,2,3 |
| 12 | 19.400 | β-Phellandrene | t | 944 | 1206 | 1,2,3 | |
| 13 | 19.500 | p-Cymene | t | 945 | 1279 | 1,2,3 | |
| 14 | 20.574 | 1,8-Cineole | 74.7 | 75.5 | 959 | 1220 | 1,2,3 |
| 15 | 21.870 | (Z)-β-Ocimene | t | 976 | 1240 | 1,2,3 | |
| 16 | 22.438 | (E)-β- Ocimene | 1.9 | 1.0 | 983 | 1260 | 1,2,3 |
| 17 | 25.243 | γ-Terpinene | 0.3 | 0.9 | 1016 | 1254 | 1,2,3 |
| 18 | 27.271 | dehydro-Linalool | 0.5 | 0.5 | 1043 | 1,2 | |
| 19 | 27.644 | 1,3,8-p-Menthatriene | 0.4 | 1048 | 1,2 | ||
| 20 | 28.128 | allo-Ocimene | 0.1 | 1055 | 1382 | 1,2 | |
| 21 | 28.799 | neo-Isopulegol | t | 1064 | 1,2 | ||
| 22 | 29.513 | Borneol | 0.2 | 1074 | 1715 | 1,2,3 | |
| 23 | 29.776 | endo-Fenchol | 0.2 | 1.0 | 1077 | 1,2 | |
| 24 | 31.870 | Terpineol | 4.2 | 1.5 | 1099 | 1710 | 1,2,3 |
| 25 | 32.268 | cis-Verbenol | 0.2 | 0.1 | 1104 | 1663 | 1,2 |
| 26 | 34.118 | Verbenone | 0.2 | 0.2 | 1131 | 1,2 | |
| 27 | 39.679 | Thymol | 0.1 | 0.3 | 1205 | 2172 | 1,2,3 |
| 28 | 40.403 | δ-Elemene | 0.1 | 1216 | 1460 | 1,2,3 | |
| 29 | 41.032 | γ-Elemene | 0.1 | 1226 | 1651 | 1,2,3 | |
| 30 | 41.512 | Myltayl-4(12)-ene | 0.1 | 1233 | 1,2 | ||
| 31 | 42.079 | dihydro-Eugenol | 0.5 | 7.5 | 1242 | 1,2 | |
| 32 | 43.444 | α-Copaene | t | 0.1 | 1262 | 1477 | 1,2,3 |
| 33 | 45.520 | Sibirene | t | 0.7 | 1294 | 1,2 | |
| 34 | 46.066 | Caryophyllene | 0.2 | 1296 | 1607 | 1,2,3 | |
| 35 | 50.831 | α-Guaiene | 0.2 | 0.3 | 1372 | 1583 | 1,2 |
| 36 | 52.204 | Cycloisolongifol-5-ol <trans> | 0.2 | 1395 | 1,2 | ||
| 37 | 56.184 | Globulol | 1.9 | 1457 | 2095 | 1,2 | |
| Total | 98.1 | 97.7 | |||||
| Monoterpene hydrocarbons | 17.1 | 15.4 | |||||
| Oxygenated monoterpenes | 80.3 | 79.1 | |||||
| Sesquiterpenes hydrocarbons | 0.7 | 1.1 | |||||
| Oxygenated sesquiterpenes | 2.1 | ||||||
| Seeds | E. gunnii EO (µg/mL) | E. pulverulenta EO (µg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1000 | 500 | 250 | 100 | 1000 | 500 | 250 | 100 | ||
| L. sativa | Mean ± SD | 7.7 ± 0.6 | 8.3 ± 0.6 | 8.0 ± 0.0 | 9.5 ± 0.7 | 9.0 ± 0.0 | 8.0 ± 0.0 | 8.5 ± 0.7 | 8.5 ± 0.7 |
| % | 9.4 | 2.3 | 0 | 0 | 0 | 5.9 | 0 | 0 | |
| P. oleracea | Mean ± SD | 0.3 ± 0.6 * | 2.3 ± 2.3 | 2.7 ± 2.1 | 3.3 ± 2.1 | 1.7 ± 1.5 ** | 2.0 ± 1.7 ** | 4.3 ± 1.5 | 2.7 ± 1.2 * |
| % | 94.6 | 58.9 | 51.8 | 41.1 | 74.7 | 70.2 | 35.9 | 59.8 | |
| R. sativus | Mean ± SD | 2.7 ± 0.6 ** | 3.7 ± 0.6 * | 5.0 ± 2.6 | 3.7 ± 0.6 * | 4.0 ± 0.0 | 3.7 ± 1.2 * | 3.7 ± 1.5 * | 6.0 ± 1.7 |
| % | 61.5 | 47.2 | 28.6 | 47.8 | 36.6 | 41.3 | 41.3 | 38.4 | |
| S. lycopersicum | Mean ± SD | 2.6 ± 1.5 ** | 4.5 ± 0.7 * | 5.7 ± 0.6 | 5.6 ± 0.6 | 8.3 ± 0.6 | 7.3 ± 0.6 | 7.3 ± 0.6 | 8.2 ± 0.6 |
| % | 67.5 | 43.8 | 27.8 | 30 | 0 | 8.7 | 8.7 | 0 | |
| P. sativum | Mean ± SD | 9.0 ± 1.7 | 9.0 ± 1.7 | 7.3 ± 2.1 | 9.3 ± 1.2 | 9.3 ± 0.6 | 8.7 ± 0.6 | 10 ± 0.0 | 9.7 ± 0.6 |
| % | 7.2 | 7.2 | 24.7 | 4.1 | 4.1 | 10.3 | 3 | 0 | |
| C. sativus | Mean ± SD | 6.0 ± 0.0 | 5.3 ± 0.6 | 5.7 ± 0.6 | 4.3 ± 0.6 | 5.3 ± 0.6 | 5.7 ± 0.6 | 6.0 ± 0.0 | 6.3 ± 0.6 |
| % | 25 | 33.7 | 28.7 | 46.2 | 11.6 | 5 | 0 | 0 | |
| L. sativum | Mean ± SD | 9.0 ± 1.0 | 8.3 ± 0.6 | 7.5 ±0.7 | 9.1 ± 0.3 | 7.0 ± 0.0 | 6.7 ± 1.5 | 6.7 ± 2.1 | 8.0 ± 1.7 |
| % | 5.5 | 2.3 | 11.8 | 0 | 12.5 | 16.2 | 16.2 | 0 | |
| L. multiflorum | Mean ± SD | 7.0 ± 2.0 | 7.5 ± 0.7 | 7.7 ± 0.6 | 7.0 ± 2.6 | 7.3 ± 0.6 | 6.0 ± 1.0 | 7.0 ± 0.0 | 8.3 ± 0.6 |
| % | 12.5 | 6.2 | 3.7 | 12.5 | 0 | 14.2 | 0 | 0 | |
| Seeds | E. gunnii EO (µg/mL) | E. pulverulenta EO (µg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 1000 | 500 | 250 | 100 | 1000 | 500 | 250 | 100 | ||
| L. sativa | Mean ± SD | 1.7 ± 0.6 | 2.5 ± 0.5 | 2.3 ± 0.6 | 2.6 ± 0.7 | 2.2 ± 0.5 | 2.1 ± 0.3 | 2.2 ± 0.5 | 2.1 ± 0.5 |
| % | 37.0 | 7.4 | 17 | 3 | 4.5 | 9.5 | 4.5 | 9.5 | |
| P. oleracea | Mean ± SD | 2.3 ± 0.4 | 2.1 ± 0.2 | 2.2 ± 0.3 | 2.0 ± 0.1 | 1.9 ± 0.4 | 2.1 ± 0.3 | 2.3 ± 0.2 | 2.2 ± 0.2 |
| % | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| R. sativus | Mean ± SD | 1.0 ± 0.2 | 1.2 ± 0.6 | 1.5 ± 0.3 | 1.7 ± 0.4 | 1.8 ± 0.5 | 1.5 ± 0.8 | 1.7 ± 0.3 | 2.4 ± 0.6 |
| % | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| S. lycopersicum | Mean ± SD | 1.3 ± 0.5 ** | 2.2 ± 1.9 * | 2.3 ± 1.7 * | 2.5 ± 1.8 * | 3.2 ± 1 | 4.5 ± 1.7 | 4.6 ± 1.7 | 5.8 ± 1.8 |
| % | 82.6 | 70.7 | 69.3 | 66.7 | 57.3 | 40 | 38.7 | 22.7 | |
| P. sativum | Mean ± SD | 4.1 ± 1.0 | 4.0 ± 1.0 | 4.5 ± 1.1 | 3.9 ± 0.9 | 3.9 ± 0.6 | 3.7 ± 0.8 | 4.5 ± 1.1 | 4.8 ± 0.9 |
| % | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| C. sativus | Mean ± SD | 6.0 ± 1.6 | 7.0 ± 1.0 | 6.9 ± 1.7 | 6.4 ± 0.8 | 5.4 ± 0.7 | 7.5 ± 0.9 | 6.7 ± 1.1 | 7.2 ± 1.9 |
| % | 15.5 | 1.4 | 2.8 | 9.9 | 23.9 | 0 | 5.6 | 0 | |
| L. sativum | Mean ± SD | 2.0 ± 1.4 | 3.0 ± 1.2 | 3.2 ± 1.3 | 2.5 ± 1.8 | 1.1 ± 0.2 ** | 1.9 ± 0.4 * | 2.2 ± 0.8 * | 2.5 ± 0.7 * |
| % | 63.0 | 44.4 | 40.7 | 53.7 | 76.0 | 58.7 | 52.2 | 45.6 | |
| L. multiflorum | Mean ± SD | 2.6 ± 0.6 * | 3.5 ± 1.1 | 4.0 ± 1.0 | 3.5 ± 1.0 | 1.6 ± 0.7 ** | 1.7 ± 0.4 ** | 2.3 ± 0.7 ** | 3.4 ± 0.7 |
| % | 53.5 | 37.5 | 28.6 | 37.5 | 70.4 | 68.5 | 57.4 | 37.0 | |
| Essential Oil | IC50 (µg/mL) |
|---|---|
| E. pulverulenta | 35.9 ± 3.6 |
| E. gunnii | 524.1 ± 10.3 |
| Acarbose (positive control) | 130.2 ± 12.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Danna, C.; Cornara, L.; Smeriglio, A.; Trombetta, D.; Amato, G.; Aicardi, P.; De Martino, L.; De Feo, V.; Caputo, L. Eucalyptus gunnii and Eucalyptus pulverulenta ‘Baby Blue’ Essential Oils as Potential Natural Herbicides. Molecules 2021, 26, 6749. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216749
Danna C, Cornara L, Smeriglio A, Trombetta D, Amato G, Aicardi P, De Martino L, De Feo V, Caputo L. Eucalyptus gunnii and Eucalyptus pulverulenta ‘Baby Blue’ Essential Oils as Potential Natural Herbicides. Molecules. 2021; 26(21):6749. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216749
Chicago/Turabian StyleDanna, Cristina, Laura Cornara, Antonella Smeriglio, Domenico Trombetta, Giuseppe Amato, Pierluca Aicardi, Laura De Martino, Vincenzo De Feo, and Lucia Caputo. 2021. "Eucalyptus gunnii and Eucalyptus pulverulenta ‘Baby Blue’ Essential Oils as Potential Natural Herbicides" Molecules 26, no. 21: 6749. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26216749