
An Insight into the Anti-Angiogenic and Anti-Metastatic Effects of Oridonin: Current Knowledge and Future Potential

 , and
, and
Abstract
:1. Introduction
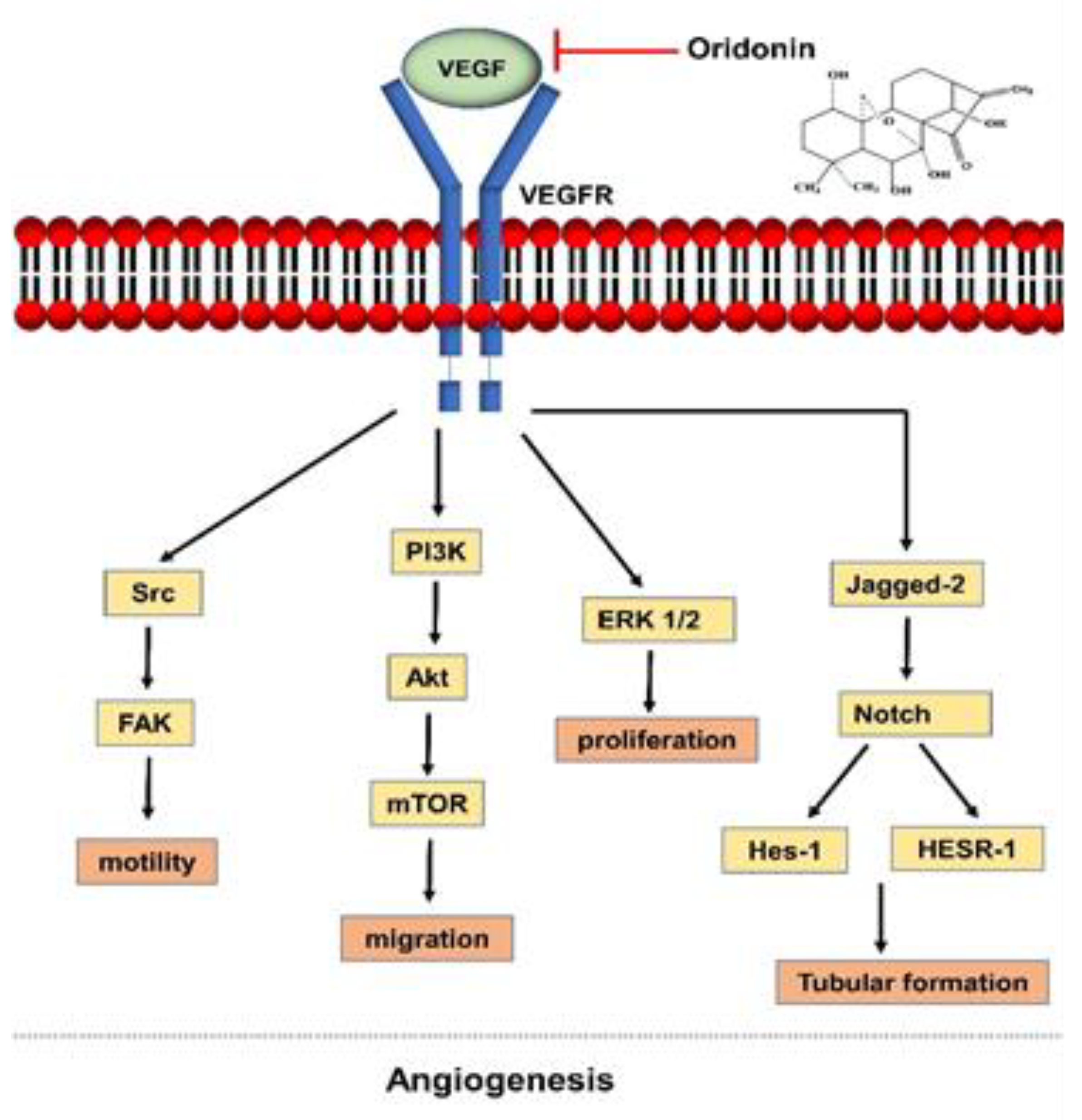
2. The Process of Angiogenesis as a Target of Oridonin
3. Oridonin in EMT
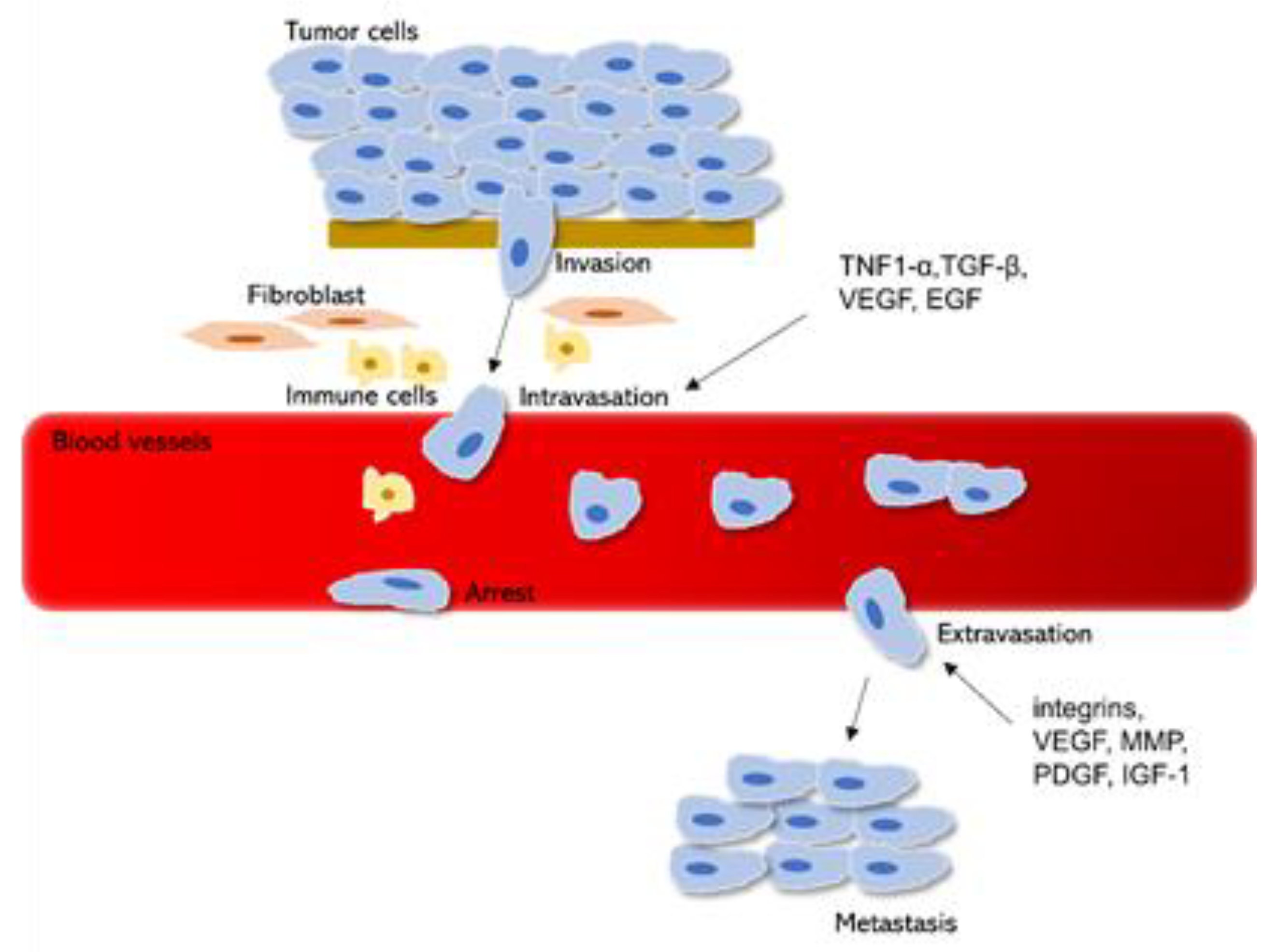
4. Cancer Invasion and Metastasis
4.1. Oridonin Inhibits TGF-β/Smad Pathway
4.2. Oridonin Blocks the Activation of EGF/EGFR/ERK Signaling Pathways
4.3. Oridonin Inhibits the Phosphorylation of mTOR Signaling Pathway
4.4. Oridonin Downregulates the lncRNA AFAP1-AS1 Expression
4.5. Oridonin Increases the Expression of p53
5. Oridonin in Hypoxia
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| EMT | epithelial-to-mesenchymal transition |
| ECM | extracellular matrix |
| VEGF | vascular endothelial growth factor |
| EGF | epidermal growth factor |
| TGF | transforming growth factor |
| VEGFR | vascular endothelial growth factor receptor |
| MMP | matrix metalloproteinases |
| ZEB1 | zinc finger E-box binding homeobox 1 |
| ADAM | metalloproteinase |
| CTC | circulating tumor cells |
| TNF1-α | tumor necrosis factor 1-α |
| PDGF | platelet-derived growth factor |
| IGF-1 | insulin-like growth factor-1 |
| lncRNAs | long non-coding RNAs |
| EGFR | endothelial growth factor receptor |
| mTOR | mammalian target of rapamycin |
| NSCLC | human non-small cell lung cancer |
| ERK | extracellular-signal-regulated kinase |
| PP2A | protein phosphatase 2A |
| FAK | focal adhesion kinase |
| FOXP3 | forkhead box P3 |
| HIF-1α | hypoxia-inducible factor 1 |
References
- Dillekås, H.; Rogers, M.S.; Straume, O. Are 90% of deaths from cancer caused by metastases? Cancer Med. 2019, 8, 5574–5576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Cancer Society. Cancer Facts and Figures; American Cancer Society (ACS): Atlanta, GA, USA, 2020. [Google Scholar]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging biological principles of metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera, E.; Gomez, H. Chemotherapy resistance in metastatic breast cancer: The evolving role of ixabepilone. In Proceedings of the Breast Cancer Research; Springer: Berlin/Heidelberg, Germany, 2010; Volume 12, p. S2. [Google Scholar]
- Luo, H.; Vong, C.T.; Chen, H.; Gao, Y.; Lyu, P.; Qiu, L.; Zhao, M.; Liu, Q.; Cheng, Z.; Zou, J.; et al. Naturally occurring anti-cancer compounds: Shining from Chinese herbal medicine. Chin. Med. 2019, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Liu, J.; Zhang, H. Efficacy of rabdosia rubescens in the treatment of gingivitis. J. Huazhong Univ. Sci. Technol. Med. Sci. 2009, 29, 659. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Liu, B.; Mao, M. Study on the Application of Chinese Patent Drug and Chinese Formula of Rabdosia Rubescens. IOP Conf. Ser. Mater. Sci. Eng. 2018, 301, 12060. [Google Scholar] [CrossRef]
- Fujita, E.; Fujita, T.; Shibuya, M. Diterpenoid constituents of Isodon trichocarpus and Isodon japonicus (terpenoids IV). Tetrahedron Lett. 1966, 7, 3153–3162. [Google Scholar] [CrossRef]
- Wang, S.; Yu, L.; Yang, H.; Li, C.; Hui, Z.; Xu, Y.; Zhu, X. Oridonin attenuates synaptic loss and cognitive deficits in an Aβ1–42-induced mouse model of Alzheimer’s disease. PLoS ONE 2016, 11, e0151397. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Jiang, H.; Chen, Y.; Ye, J.; Wang, A.; Wang, C.; Liu, Q.; Liang, G.; Deng, X.; Jiang, W. Oridonin is a covalent NLRP3 inhibitor with strong anti-inflammasome activity. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Ouyang, P.; Gu, K.; Rehman, T.; Zhang, T.; Yin, Z.; Fu, H.; Lin, J.; He, C.; Shu, G. The antibacterial mechanism of oridonin against methicillin-resistant Staphylococcus aureus (MRSA). Pharm. Biol. 2019, 57, 710–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhong, Z.; Wan, J.; Tan, W.; Wu, G.; Chen, M.; Wang, Y. Oridonin induces apoptosis, inhibits migration and invasion on highly-metastatic human breast cancer cells. Am. J. Chin. Med. 2013, 41, 177–196. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Xie, F.; Li, M.; Liang, Z.; Xu, W.; Yang, J.; Liu, C.; Li, H.; Zhou, H.; Qu, L.-H. Oridonin induces autophagy via inhibition of glucose metabolism in p53-mutated colorectal cancer cells. Cell Death Dis. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Shen, C.; Li, Y.; Zhang, T.; Huang, H.; Ren, J.; Hu, Z.; Xu, J.; Xu, B. Oridonin overcomes the gemcitabine resistant PANC-1/Gem cells by regulating GST pi and LRP/1 ERK/JNK signalling. OncoTargets Ther. 2019, 12, 5751–5765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Bi, Y.; Xu, Y.; Zhang, Z.; Xu, W.; Zhang, S.; Chen, J. Oridonin inhibits the migration and epithelial-to-mesenchymal transition of small cell lung cancer cells by suppressing FAK-ERK1/2 signalling pathway. J. Cell. Mol. Med. 2020, 24, 4480–4493. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.-L.; Ghoorun, R.A.; Wu, X.-H.; Chen, H.-L.; Zhou, Q.; Wu, X.-B. Oridonin induces apoptosis in HGC-27 cells by activating the JNK signaling pathway. Oncol. Lett. 2020, 19, 255–260. [Google Scholar] [CrossRef]
- Lu, J.; Chen, X.; Qu, S.; Yao, B.; Xu, Y.; Wu, J.; Jin, Y.; Ma, C. Oridonin induces G2/M cell cycle arrest and apoptosis via the PI3K/Akt signaling pathway in hormone-independent prostate cancer cells. Oncol. Lett. 2017, 13, 2838–2846. [Google Scholar] [CrossRef] [Green Version]
- Ren, K.-K.; Wang, H.-Z.; Xie, L.-P.; Chen, D.-W.; Liu, X.; Sun, J.; Nie, Y.-C.; Zhang, R.-Q. The effects of oridonin on cell growth, cell cycle, cell migration and differentiation in melanoma cells. J. Ethnopharmacol. 2006, 103, 176–180. [Google Scholar] [CrossRef]
- Nagy, J.A.; Dvorak, H.F. Heterogeneity of the tumor vasculature: The need for new tumor blood vessel type-specific targets. Clin. Exp. Metastasis 2012, 29, 657–662. [Google Scholar] [CrossRef] [Green Version]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: Causes, consequences, challenges and opportunities. Cell. Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [Green Version]
- Nasir, A. Angiogenic signaling pathways and anti-angiogenic therapies in human cancer. In Predictive Biomarkers in Oncology; Springer: Cham, Switzerland, 2019; pp. 243–262. [Google Scholar]
- Bergers, G.; Benjamin, L.E. Tumorigenesis and the angiogenic switch. Nat. Rev. Cancer 2003, 3, 401–410. [Google Scholar] [CrossRef]
- Almog, N.; Ma, L.; Raychowdhury, R.; Schwager, C.; Erber, R.; Short, S.; Hlatky, L.; Vajkoczy, P.; Huber, P.E.; Folkman, J. Transcriptional switch of dormant tumors to fast-growing angiogenic phenotype. Cancer Res. 2009, 69, 836–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuazo-Gaztelu, I.; Casanovas, O. Unraveling the role of angiogenesis in cancer ecosystems. Front. Oncol. 2018, 8, 248. [Google Scholar] [CrossRef] [PubMed]
- Isogai, S.; Lawson, N.D.; Torrealday, S.; Horiguchi, M.; Weinstein, B.M. Angiogenic network formation in the developing vertebrate trunk. Development 2003, 130, 5281–5290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zecchin, A.; Kalucka, J.; Dubois, C.; Carmeliet, P. How endothelial cells adapt their metabolism to form vessels in tumors. Front. Immunol. 2017, 8, 1750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shawber, C.J.; Kitajewski, J. Notch function in the vasculature: Insights from zebrafish, mouse and man. Bioessays 2004, 26, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Liang, N.; Zhang, J.; Xie, J.; Liu, F.; Xu, D.; Yu, X.; Tian, Y. Advanced research on vasculogenic mimicry in cancer. J. Cell. Mol. Med. 2015, 19, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Tewari, K.S.; Sill, M.W.; Long, H.J., III; Penson, R.T.; Huang, H.; Ramondetta, L.M.; Landrum, L.M.; Oaknin, A.; Reid, T.J.; Leitao, M.M. Improved survival with bevacizumab in advanced cervical cancer. N. Engl. J. Med. 2014, 370, 734–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finke, J.; Ko, J.; Rini, B.; Rayman, P.; Ireland, J.; Cohen, P. MDSC as a mechanism of tumor escape from sunitinib mediated anti-angiogenic therapy. Int. Immunopharmacol. 2011, 11, 856–861. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, M. Vascular endothelial growth factor (VEGF) and its receptor (VEGFR) signaling in angiogenesis: A crucial target for anti-and pro-angiogenic therapies. Genes Cancer 2011, 2, 1097–1105. [Google Scholar] [CrossRef]
- Sa‑Nguanraksa, D.; Chuangsuwanich, T.; Pongpruttipan, T. High vascular endothelial growth factor gene expression predicts poor outcome in patients with non-luminal A breast cancer. Mol. Clin. Oncol. 2015, 3, 1103–1108. [Google Scholar] [CrossRef] [Green Version]
- George, D.J.; Halabi, S.; Shepard, T.F.; Vogelzang, N.J.; Hayes, D.F.; Small, E.J.; Kantoff, P.W. Prognostic significance of plasma vascular endothelial growth factor levels in patients with hormone-refractory prostate cancer treated on Cancer and Leukemia Group B 9480. Clin. Cancer Res. 2001, 7, 1932–1936. [Google Scholar] [PubMed]
- Horikawa, N.; Abiko, K.; Matsumura, N.; Hamanishi, J.; Baba, T.; Yamaguchi, K.; Yoshioka, Y.; Koshiyama, M.; Konishi, I. Expression of vascular endothelial growth factor in ovarian cancer inhibits tumor immunity through the accumulation of myeloid-derived suppressor cells. Clin. Cancer Res. 2017, 23, 587–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanotelli, M.R.; Reinhart-King, C.A. Mechanical forces in tumor angiogenesis. In Biomechanics in Oncology; Springer: New York, NY, USA, 2018; pp. 91–112. [Google Scholar]
- Dong, Y.; Zhang, T.; Li, J.; Deng, H.; Song, Y.; Zhai, D.; Peng, Y.; Lu, X.; Liu, M.; Zhao, Y.; et al. Oridonin inhibits tumor growth and metastasis through anti-angiogenesis by blocking the notch signaling. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.-H.; Pi, J.; Cai, J.-Y. Oridonin exhibits anti-angiogenic activity in human umbilical vein endothelial cells by inhibiting VEGF-induced VEGFR-2 signaling pathway. Pathol. Res. Pract. 2020, 216, 153031. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Xie, K.; Sheng, D.; Wan, X.; Zhu, G. Antiangiogenic effects of oridonin. BMC Complement. Altern. Med. 2017, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Wang, Q.; Shen, S.; Wei, X.; Li, G. Oridonin inhibits vegf-a-associated angiogenesis and epithelial-mesenchymal transition of breast cancer in vitro and in vivo. Oncol. Lett. 2018, 16, 2289–2298. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Zhang, X.; Li, C.; Guan, H. Oridonin inhibits breast cancer growth and metastasis through blocking the Notch signaling. Saudi Pharm. J. 2017, 25, 638–643. [Google Scholar] [CrossRef]
- Karar, J.; Maity, A. PI3K/AKT/mTOR pathway in angiogenesis. Front. Mol. Neurosci. 2011, 4, 51. [Google Scholar] [CrossRef] [Green Version]
- Pujade-Lauraine, E.; Hilpert, F.; Weber, B.; Reuss, A.; Poveda, A.; Kristensen, G.; Sorio, R.; Vergote, I.; Witteveen, P.; Bamias, A. Bevacizumab combined with chemotherapy for platinum-resistant recurrent ovarian cancer: The AURELIA open-label randomized phase III trial. Obstet. Gynecol. Surv. 2014, 69, 402–404. [Google Scholar] [CrossRef]
- Li, J.; Wu, Y.; Wang, D.; Zou, L.; Fu, C.; Zhang, J.; Leung, G.P.-H. Oridonin synergistically enhances the anti-tumor efficacy of doxorubicin against aggressive breast cancer via pro-apoptotic and anti-angiogenic effects. Pharmacol. Res. 2019, 146. [Google Scholar] [CrossRef]
- Steeg, P.S. Targeting metastasis. Nat. Rev. Cancer 2016, 16, 201. [Google Scholar] [CrossRef] [PubMed]
- Thiery, J.P. Epithelial–mesenchymal transitions in tumour progression. Nat. Rev. Cancer 2002, 2, 442–454. [Google Scholar] [CrossRef] [PubMed]
- De Craene, B.; Berx, G. Regulatory networks defining EMT during cancer initiation and progression. Nat. Rev. Cancer 2013, 13, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Du, Y.; Beckford, J.; Alachkar, H. Upregulation of the EMT marker vimentin is associated with poor clinical outcome in acute myeloid leukemia. J. Transl. Med. 2018, 16, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Mendez, M.G.; Kojima, S.; Goldman, R.D. Vimentin induces changes in cell shape, motility, and adhesion during the epithelial to mesenchymal transition. FASEB J. 2010, 24, 1838–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, C.-T.; Cheng, H.-W.; Huang, C.-M.; Li, H.-R.; Ou, M.-H.; Huang, J.-R.; Khoo, K.-H.; Yu, H.W.; Chen, Y.-Q.; Wang, Y.-K. Fibronectin in cell adhesion and migration via N-glycosylation. Oncotarget 2017, 8, 70653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamaria, P.G.; Moreno-Bueno, G.; Portillo, F.; Cano, A. EMT: Present and future in clinical oncology. Mol. Oncol. 2017, 11, 718–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gui, Z.; Luo, F.; Yang, Y.; Shen, C.; Li, S.; Xu, J. Oridonin inhibition and miR-200b-3p/ZEB1 axis in human pancreatic cancer. Int. J. Oncol. 2017, 50, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Lou, S.; Xu, J.; Wang, B.; Li, S.; Ren, J.; Hu, Z.; Xu, B.; Luo, F. Downregulation of lncRNA AFAP1-AS1 by oridonin inhibits the epithelial-to-mesenchymal transition and proliferation of pancreatic cancer cells. Acta Biochim. Biophys. Sin. 2018, 51, 814–825. [Google Scholar] [CrossRef]
- Sun, Y.; Jiang, X.; Lu, Y.; Zhu, J.; Yu, L.; Ma, B.; Zhang, Q. Oridonin prevents epithelial-mesenchymal transition and TGF-β1-induced epithelial-mesenchymal transition by inhibiting TGF-β1/Smad2/3 in osteosarcoma. Chem. Biol. Interact. 2018, 296, 57–64. [Google Scholar] [CrossRef]
- Wieczorek, E.; Jablonska, E.; Wasowicz, W.; Reszka, E. Matrix metalloproteinases and genetic mouse models in cancer research: A mini-review. Tumor Biol. 2015, 36, 163–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, B.N.; Bhowmick, N.A. Role of EMT in metastasis and therapy resistance. J. Clin. Med. 2016, 5, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Tan, W.; Du, B.; Liu, W.; Li, W.; Che, D.; Zhang, G. Oridonin effectively reverses cisplatin drug resistance in human ovarian cancer cells via induction of cell apoptosis and inhibition of matrix metalloproteinase expression. Mol. Med. Rep. 2016, 13, 3342–3348. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yan, S.; Tao, Z.; Wang, C.; Huang, M.; Zhang, X.; Du, Y. Oridonin inhibits the proliferation, migration and invasion of human osteosarcoma cells via suppression of matrix metalloproteinase expression and STAT3 signalling pathway. J. BU ON 2019, 24, 1175–1180. [Google Scholar]
- Zhang, Y.; Wang, L.; Zi, Y.; Zhang, L.; Guo, Y.; Huang, Y. Oridonin effectively reverses the drug resistance of cisplatin involving induction of cell apoptosis and inhibition of MMP expression in human acute myeloid leukemia cells. Saudi J. Biol. Sci. 2017, 24, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; He, Z.; Cao, W.; Cai, F.; Zhang, L.; Huang, Q.; Fan, C.; Duan, C.; Wang, X.; Wang, J.; et al. Oridonin inhibits gefitinib-resistant lung cancer cells by suppressing EGFR/ERK/MMP-12 and CIP2A/Akt signaling pathways. Int. J. Oncol. 2016, 48, 2608–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.-Q.; Chen, K.; Ye, Q.; Jiang, X.-H.; Sun, Y.-W. Oridonin inhibits pancreatic cancer cell migration and epithelial-mesenchymal transition by suppressing Wnt/β-catenin signaling pathway. Cancer Cell Int. 2016, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, X.; Ye, M.; Jing, P.; Xiong, J.; Han, Z.; Kong, J.; Li, M.; Lai, X.; Chang, N.; et al. FBW7 loss promotes epithelial-to-mesenchymal transition in non-small cell lung cancer through the stabilization of Snail protein. Cancer Lett. 2018, 419, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of extracellular matrix remodelling in tumour progression and metastasis. Nat. Commun. 2020, 11, 1–19. [Google Scholar]
- Chiang, S.P.H.; Cabrera, R.M.; Segall, J.E. Tumor cell intravasation. Am. J. Physiol. Physiol. 2016, 311, C1–C14. [Google Scholar] [CrossRef] [Green Version]
- Fontebasso, Y.; Dubinett, S.M. Drug development for metastasis prevention. Crit. Rev. Oncog. 2015, 20, 449–473. [Google Scholar] [CrossRef] [PubMed]
- Fares, J.; Fares, M.Y.; Khachfe, H.H.; Salhab, H.A.; Fares, Y. Molecular principles of metastasis: A hallmark of cancer revisited. Signal Transduct. Target. Ther. 2020, 5, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Valastyan, S.; Weinberg, R.A. Tumor metastasis: Molecular insights and evolving paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenbroucke, R.E.; Libert, C. Is there new hope for therapeutic matrix metalloproteinase inhibition? Nat. Rev. Drug Discov. 2014, 13, 904–927. [Google Scholar] [CrossRef]
- Teixeira, A.F.; Dijke, P.T.; Zhu, H.-J. On-Target Anti-TGF-β Therapies Are Not Succeeding in Clinical Cancer Treatments: What Are Remaining Challenges? Front. Cell Dev. Biol. 2020, 8, 605. [Google Scholar] [CrossRef]
- Bu, H.-Q.; Shen, F.; Cui, J. The inhibitory effect of oridonin on colon cancer was mediated by deactivation of TGF-β1/smads-pai-1 signaling pathway in vitro and vivo. OncoTargets Ther. 2019, 12, 7467–7476. [Google Scholar] [CrossRef] [Green Version]
- Xia, R.; Chen, S.-X.; Qin, Q.; Chen, Y.; Zhang, W.-W.; Zhu, R.R.; Deng, A.-M. Oridonin suppresses proliferation of human ovarian cancer cells via blockage of mTOR signaling. Asian Pac. J. Cancer Prev. 2016, 17, 667–671. [Google Scholar] [CrossRef] [Green Version]
- Bi, E.; Liu, D.; Li, Y.; Mao, X.; Wang, A.; Wang, J. Oridonin induces growth inhibition and apoptosis in human gastric carcinoma cells by enhancement of p53 expression and function. Braz. J. Med. Biol. Res. 2018, 51. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Yang, S.; Sun, G.; Huang, W.; Zhang, Y. Transforming growth factor-beta polymorphisms and serum level in the development of osteosarcoma. DNA Cell Biol. 2014, 33, 802–806. [Google Scholar] [CrossRef]
- Ahel, J.; Hudorović, N.; Vičić-Hudorović, V.; Nikles, H. Tgf-Beta in the Natural History of Prostate Cancer. Acta Clin. Croat. 2019, 58, 128–138. [Google Scholar] [CrossRef] [Green Version]
- Yang, L. TGFβ and cancer metastasis: An inflammation link. Cancer Metastasis Rev. 2010, 29, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Margadant, C.; Sonnenberg, A. Integrin–TGF-β crosstalk in fibrosis, cancer and wound healing. EMBO Rep. 2010, 11, 97–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarwar, M.D.; Xia, Y.-X.; Liang, Z.-M.; Tsang, S.W.; Zhang, H.-J. Mechanistic pathways and molecular targets of plant-derived anticancer ent-kaurane diterpenes. Biomolecules 2020, 10, 144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.-Y.; Wang, Q.; Shen, S.; Wei, X.-L.; Li, G.-X. Oridonin inhibits migration, invasion, adhesion and TGF-ß1-induced epithelial-mesenchymal transition of melanoma cells by inhibiting the activity of PI3K/AKT/GSK-3ß signaling pathway. Oncol. Lett. 2018, 15, 1362–1372. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Chen, T.; Ma, Z.; Qiao, C.; Yuan, F.; Guo, X.; Liu, J.; Shen, Y.; Yu, L.; Xiang, A. Oridonin inhibits 4T1 tumor growth by suppressing Treg differentiation via TGF-β receptor. Int. Immunopharmacol. 2020, 88, 106831. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Jang, M.H.; Kim, E.J.; Kim, H.J.; Lee, H.J.; Kim, Y.J.; Kim, J.H.; Kang, E.; Kim, S.-W.; Kim, I.A. High EGFR gene copy number predicts poor outcome in triple-negative breast cancer. Mod. Pathol. 2014, 27, 1212–1222. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Li, D.; Sun, L. High levels of EGFR expression in tumor stroma are associated with aggressive clinical features in epithelial ovarian cancer. OncoTargets Ther. 2016, 9, 377. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.-Y.; Tsai, P.-C.; Tseng, C.-H.; Chen, Y.; Chang, L.-S.; Lin, S.-R. Inhibition of EGF/EGFR activation with naphtho [1, 2-b] furan-4, 5-dione blocks migration and invasion of MDA-MB-231 cells. Toxicol. In Vitro 2013, 27, 1–10. [Google Scholar] [CrossRef]
- Guo, Y.; Pan, W.; Liu, S.; Shen, Z.; Xu, Y.; Hu, L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007. [Google Scholar] [CrossRef] [Green Version]
- Jackson, J.L.; Young, M.R.I. Protein phosphatase-2A regulates protein tyrosine phosphatase activity in Lewis lung carcinoma tumor variants. Clin. Exp. Metastasis 2003, 20, 357–364. [Google Scholar] [CrossRef]
- Benefield, J.; Meisinger, J.; Petruzzelli, G.J.; Young, M.R. Endothelial cell response to human head and neck squamous cell carcinomas involves downregulation of protein phosphatases-1/2A, cytoskeletal depolymerization and increased motility. Invasion Metastasis 1997, 17, 210–220. [Google Scholar] [PubMed]
- Zhang, J.; Hochwald, S.N. The role of FAK in tumor metabolism and therapy. Pharmacol. Ther. 2014, 142, 154–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, A.; Kunz, J.; Hall, M.N. TOR2 is required for organization of the actin cytoskeleton in yeast. Proc. Natl. Acad. Sci. USA 1996, 93, 13780–13785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dada, S.; Demartines, N.; Dormond, O. mTORC2 regulates PGE2-mediated endothelial cell survival and migration. Biochem. Biophys. Res. Commun. 2008, 372, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, Z. Oridonin inhibits metastasis of human ovarian cancer cells by suppressing the mTOR pathway. Arch. Med. Sci. 2019, 15, 1017–1027. [Google Scholar] [CrossRef]
- Lu, L.; Barbi, J.; Pan, F. The regulation of immune tolerance by FOXP3. Nat. Rev. Immunol. 2017, 17, 703. [Google Scholar] [CrossRef]
- Zhang, C.; Xu, Y.; Hao, Q.; Wang, S.; Li, H.; Li, J.; Gao, Y.; Li, M.; Li, W.; Xue, X. FOXP 3 suppresses breast cancer metastasis through downregulation of CD 44. Int. J. Cancer 2015, 137, 1279–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, H.; Gao, Y.; Fan, X.; Liu, X.; Peng, L.; Ci, X. Oridonin sensitizes cisplatin-induced apoptosis via AMPK/Akt/mTOR-dependent autophagosome accumulation in A549 cells. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bu, H.; Liu, D.; Zhang, G.; Chen, L.; Song, Z. AMPK/mTOR/ULK1 Axis-Mediated Pathway Participates in Apoptosis and Autophagy Induction by Oridonin in Colon Cancer DLD-1 Cells. OncoTargets Ther. 2020, 13, 8533. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Wan, Q.; Zhang, L.; Li, Y.; Zhang, P.; Li, D.; Feng, C.; Yi, F.; Zhang, L.; Ding, X. Analysis of LncRNA expression in cell differentiation. RNA Biol. 2018, 15, 413–422. [Google Scholar] [CrossRef]
- Gibb, E.A.; Brown, C.J.; Lam, W.L. The functional role of long non-coding RNA in human carcinomas. Mol. Cancer 2011, 10, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalbantoglu, S.; Abu-Asab, M.; Tan, M.; Zhang, X.; Cai, L.; Amri, H. Study of clinical survival and gene expression in a sample of pancreatic ductal adenocarcinoma by parsimony phylogenetic analysis. OMICS A J. Integr. Biol. 2016, 20, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Muller, P.A.J.; Vousden, K.H.; Norman, J.C. p53 and its mutants in tumor cell migration and invasion. J. Cell Biol. 2011, 192, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-P.; Wang, W.-L.; Chang, Y.-L.; Wu, C.-T.; Chao, Y.-C.; Kao, S.-H.; Yuan, A.; Lin, C.-W.; Yang, S.-C.; Chan, W.-K. p53 controls cancer cell invasion by inducing the MDM2-mediated degradation of Slug. Nat. Cell Biol. 2009, 11, 694–704. [Google Scholar] [CrossRef] [PubMed]
- Shi, D.; Gu, W. Dual roles of MDM2 in the regulation of p53: Ubiquitination dependent and ubiquitination independent mechanisms of MDM2 repression of p53 activity. Genes Cancer 2012, 3, 240–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busuttil, R.A.; Zapparoli, G.V.; Haupt, S.; Fennell, C.; Wong, S.Q.; Pang, J.-M.B.; Takeno, E.A.; Mitchell, C.; Di Costanzo, N.; Fox, S. Role of p53 in the progression of gastric cancer. Oncotarget 2014, 5, 12016. [Google Scholar] [CrossRef] [Green Version]
- Fenoglio-Preiser, C.M.; Wang, J.; Stemmermann, G.N.; Noffsinger, A. TP53 and gastric carcinoma: A review. Hum. Mutat. 2003, 21, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.-Q.; Zhang, C.; Guo, Z.-Y.; Yang, J.-M.; Guo, J.-H.; Chen, C.; Yao, Q.-H.; Liu, F.; Zhang, Q.-W.; Gao, F.-H. Oridonin induces Mdm2-p60 to promote p53-mediated apoptosis and cell cycle arrest in neuroblastoma. Cancer Med. 2019, 8, 5313–5326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.-H.; Pi, J.; Jin, H.; Cai, J.-Y. Oridonin-induced mitochondria-dependent apoptosis in esophageal cancer cells by inhibiting PI3K/AKT/mTOR and Ras/Raf pathways. J. Cell. Biochem. 2019, 120, 3736–3746. [Google Scholar] [CrossRef] [PubMed]
- Vaupel, P.; Mayer, A.; Höckel, M. Tumor hypoxia and malignant progression. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2004; Volume 381, pp. 335–354. ISBN 0076-6879. [Google Scholar]
- Jögi, A.; Ehinger, A.; Hartman, L.; Alkner, S. Expression of HIF-1α is related to a poor prognosis and tamoxifen resistance in contralateral breast cancer. PLoS ONE 2019, 14, e0226150. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.-H.; Wu, K.-J. TWIST activation by hypoxia inducible factor-1 (HIF-1): Implications in metastasis and development. Cell Cycle 2008, 7, 2090–2096. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shi, X.; Peng, Y.; Wu, M.; Zhang, P.; Xie, R.; Wu, Y.; Yan, Q.; Liu, S.; Wang, J. HIF-1α promotes epithelial-mesenchymal transition and metastasis through direct regulation of ZEB1 in colorectal cancer. PLoS ONE 2015, 10, e0129603. [Google Scholar] [CrossRef] [PubMed]
- Krock, B.L.; Skuli, N.; Simon, M.C. Hypoxia-induced angiogenesis: Good and evil. Genes Cancer 2011, 2, 1117–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, Y.-P.; Wu, K.-J. Hypoxia-regulated target genes implicated in tumor metastasis. J. Biomed. Sci. 2012, 19, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, X.; Yang, F.; Shao, C.; Wei, K.; Xie, M.; Shen, H.; Shu, Y. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol. Cancer 2019, 18, 157. [Google Scholar] [CrossRef] [Green Version]
- Guise, C.P.; Mowday, A.M.; Ashoorzadeh, A.; Yuan, R.; Lin, W.-H.; Wu, D.-H.; Smaill, J.B.; Patterson, A.V.; Ding, K. Bioreductive prodrugs as cancer therapeutics: Targeting tumor hypoxia. Chin. J. Cancer 2014, 33, 80. [Google Scholar] [CrossRef] [PubMed]
- Marcu, L.; Olver, I. Tirapazamine: From bench to clinical trials. Curr. Clin. Pharmacol. 2006, 1, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.M.; Hendriks, H.R.; Peters, G.J.; on Behalf of the EORTC-Pharmacology; Molecular Mechanism Group. EO9 (Apaziquone): From the clinic to the laboratory and back again. Br. J. Pharmacol. 2013, 168, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Chen, K.; Ye, J.; Qi, L.; Liao, Y.; Li, R.; Song, S.; Zhou, C.; Feng, R.; Zhai, W. Oridonin inhibits hypoxia-induced epithelial-mesenchymal transition and cell migration by the hypoxia-inducible factor-1α/matrix metallopeptidase-9 signal pathway in gallbladder cancer. Anti Cancer Drugs 2019, 30, 925–932. [Google Scholar] [CrossRef]
- Ding, Y.; Ding, C.; Ye, N.; Liu, Z.; Wold, E.A.; Chen, H.; Wild, C.; Shen, Q.; Zhou, J. Discovery and development of natural product oridonin-inspired anticancer agents. Eur. J. Med. Chem. 2016, 122, 102–117. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Yun, E.-J.; Chen, W.; Ding, Y.; Wu, K.; Wang, B.; Ding, C.; Hernandez, E.; Santoyo, J.; Pong, R.-C.; et al. Targeting 3-phosphoinositide-dependent protein kinase 1 associated with drug-resistant renal cell carcinoma using new oridonin analogs. Cell Death Dis. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Yang, J.; Ran, Q.; Wang, L.; Liu, J.; Wang, Z.; Wu, X.; Hua, W.; Yuan, S.; Zhang, L.; et al. Synthesis and biological evaluation of novel 1-O- and 14-O-derivatives of oridonin as potential anticancer drug candidates. Bioorg. Med. Chem. Lett. 2008, 18, 4741–4744. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| EMT Marker | Cancer Type | Cell Line | Concentration and Treatment Time | Effect | References |
|---|---|---|---|---|---|
| ZEB1 | Pancreatic | BxPC-3 | 87.8–95 µM (24 h) | Downregulation of ZEB1 protein expression | [52,53] |
| PANC-1 | 55.8 μM–95 µM (24 h) | ||||
| E-cadherin | Pancreatic | BxPC-3 | 95 µM (24 h) | Enhances of E-cadherin protein expression | [53] |
| PANC-1 | 95 µM (24 h) | ||||
| SW1990 | 15 µM (24 h) | Enhances of E-cadherin protein expression | [61] | ||
| Breast | MDA-MB-231 | 4–16 µM (24 h) | Upregulation of E-cadherin mRNA and protein expression | [40] | |
| 4T1 | |||||
| Bone | MG-63 | 0.8–2 µM (24 h) | Upregulation of E-cadherin mRNA and protein expression | [54] | |
| 143B | |||||
| Lung | H1688 | 5–10 µM (24 h) | Upregulation of E-cadherin mRNA and protein expression | [16] | |
| N-cadherin | Pancreatic | BxPC-3 | 87.8–95 µM (24 h) | Downregulation of N-cadherin protein expression | [52,53] |
| PANC-1 | 55.8 μM–95 µM (24 h) | ||||
| Breast | MDA-MB-231 | 4–16 µM (24 h) | Downregulation of N-cadherin mRNA and protein expression | [40] | |
| 4T1 | |||||
| Bone | MG-63 | 0.8–2 µM (24 h) | Downregulation N-cadherin mRNA and protein expression | [54] | |
| 143B | |||||
| Fibronectin | Pancreatic | BxPC-3 | 87.8 µM (24 h) | Downregulation of fibronectin protein expression | [52] |
| PANC-1 | 55.8 μM (24 h) | ||||
| Vimentin | Breast | MDA-MB-231 | 4–16 µM (24 h) | Downregulation of vimentin mRNA and protein expression | [40] |
| 4T1 | |||||
| Pancreatic | SW1990 | 15 µM (24 h) | Downregulation of vimentin mRNA levels | [61] | |
| Bone | MG-63 | 0.8–2 µM (24 h) | Downregulation of vimentin mRNA and protein expression | [54] | |
| 143B | |||||
| Lung | H1688 | 5–10 µM (24 h) | Downregulation of vimentin mRNA and protein expression | [16] | |
| Snail | Breast | MDA-MB-231 | 4–16 µM (24 h) | Downregulation of snail mRNA and protein expression | [40] |
| 4T1 | |||||
| Pancreatic | SW1990 | 15 µM (24 h) | Decreases snail mRNA levels | [61] | |
| BxPC-3 | 95 µM (24 h) | Downregulation of snail protein expression | [53] | ||
| PANC-1 | |||||
| Bone | MG-63 | 0.8–2 µM (24 h) | Downregulation of snail mRNA and protein expression | [54] | |
| 143B | |||||
| Lung | H1688 | 5–10 µM (24 h) | Downregulation of snail mRNA and protein expression | [16] | |
| Skin | A375 | 20 µM (12 h) | Downregulation of snail protein expression | [62] | |
| MDA-MB-435S | |||||
| Slug | Pancreatic | SW1990 | 15 µM (24 h) | Decreases slug mRNA levels | [61] |
| BxPC-3 | 95 µM (24 h) | Downregulation of slug protein expression | [53] | ||
| PANC-1 | |||||
| Bone | MG-63 | 0.8–2 µM (24 h) | Downregulation of slug mRNA and protein expression | [54] | |
| 143B | |||||
| Lung | H1688 | 5–10 µM (24 h) | Downregulation of slug mRNA and protein expression | [16] | |
| MMP-2 | Breast | MDA-MB-231 | 1–5 µM (24 h) | Downregulation of MMP-2 protein expression | [13] |
| AML | MV4-11/DDP | 10–80 µM (48 h) | Downregulation of MMP-2 protein expression | [59] | |
| Bone | U2OS | 15–60 µM (48 h) | Downregulation of MMP-2 protein expression | [58] | |
| Ovarian | A2780/DDP | 10–80 µM (48 h) | Downregulation of MMP-2 protein expression | [57] | |
| MMP-3 | Bone | U2OS | 15–60 µM (48 h) | Downregulation of MMP-3 protein expression | [58] |
| MMP-9 | Breast | MDA-MB-231 | 1–5 µM (24 h) | Downregulation of MMP-9 protein expression | [13] |
| AML | MV4-11/DDP | 10–80 µM (48 h) | Downregulation of MMP-9 protein expression | [59] | |
| Bone | U2OS | 15–60 µM (48 h) | Downregulation of MMP-9 protein expression | [58] | |
| Ovarian | A2780/DDP | 10–80 µM (48 h) | Downregulation of MMP-9 protein expression | [57] | |
| MMP-12 | Lung | H1975 | 10–20 µM (24 h) | Downregulation of MMP-12 protein expression | [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdullah, N.A.; Md Hashim, N.F.; Ammar, A.; Muhamad Zakuan, N. An Insight into the Anti-Angiogenic and Anti-Metastatic Effects of Oridonin: Current Knowledge and Future Potential. Molecules 2021, 26, 775. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040775
Abdullah NA, Md Hashim NF, Ammar A, Muhamad Zakuan N. An Insight into the Anti-Angiogenic and Anti-Metastatic Effects of Oridonin: Current Knowledge and Future Potential. Molecules. 2021; 26(4):775. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040775
Chicago/Turabian StyleAbdullah, Nurul Akmaryanti, Nur Fariesha Md Hashim, Aula Ammar, and Noraina Muhamad Zakuan. 2021. "An Insight into the Anti-Angiogenic and Anti-Metastatic Effects of Oridonin: Current Knowledge and Future Potential" Molecules 26, no. 4: 775. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040775