
Anti-Malignant Ascites Effect of Total Diterpenoids from Euphorbiae ebracteolatae Radix Is Attributable to Alterations of Aquaporins via Inhibiting PKC Activity in the Kidney

 ,
,  ,
,  ,
,  , , , and
, , , and 
Abstract
:1. Introduction
2. Results
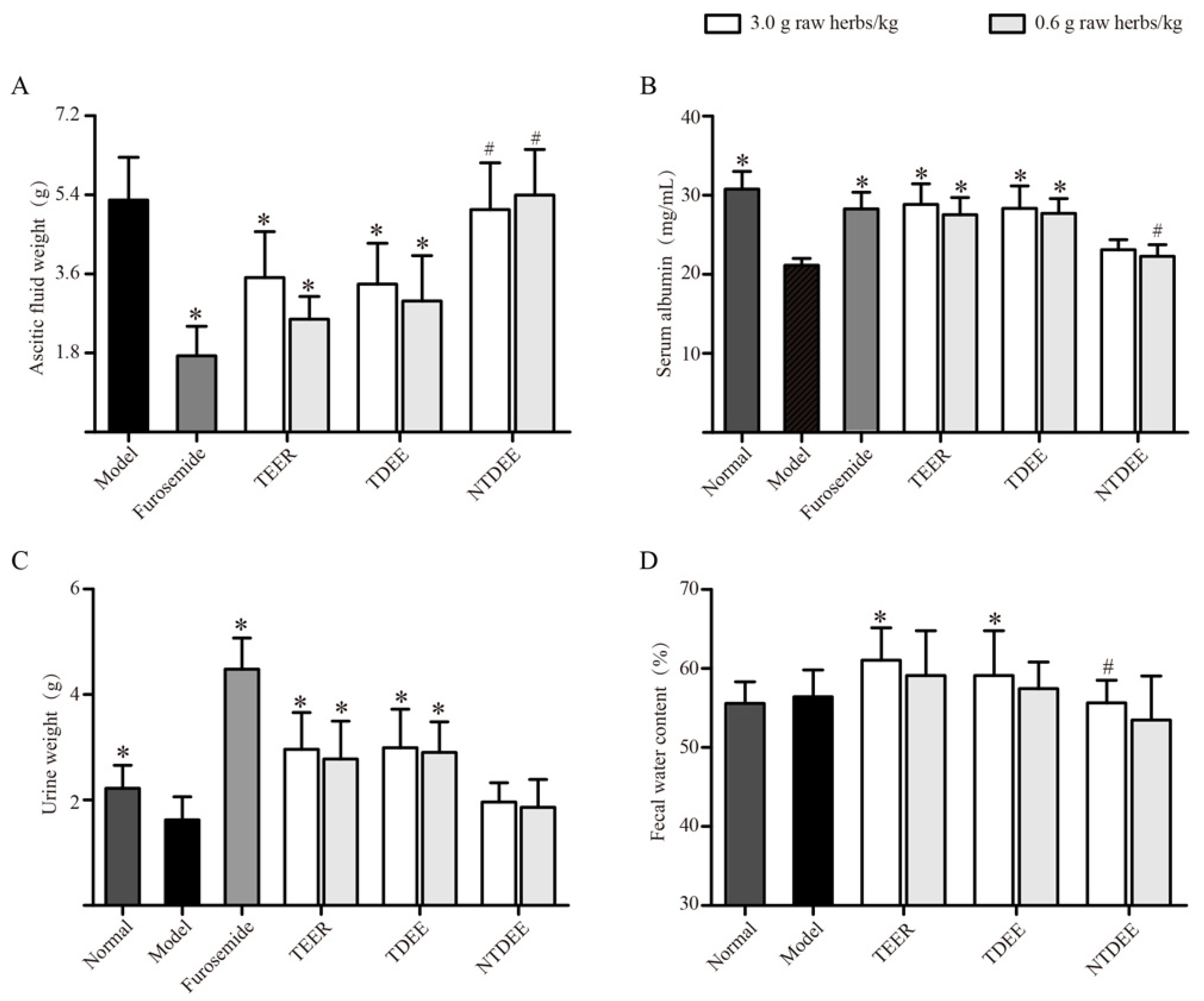
2.1. Effects of TDEE on Ascitic Fluid Weight, Urine Weight, and Fecal Water Content
2.2. Effects of TDEE on Serum Albumin Levels in Ascitic Mice
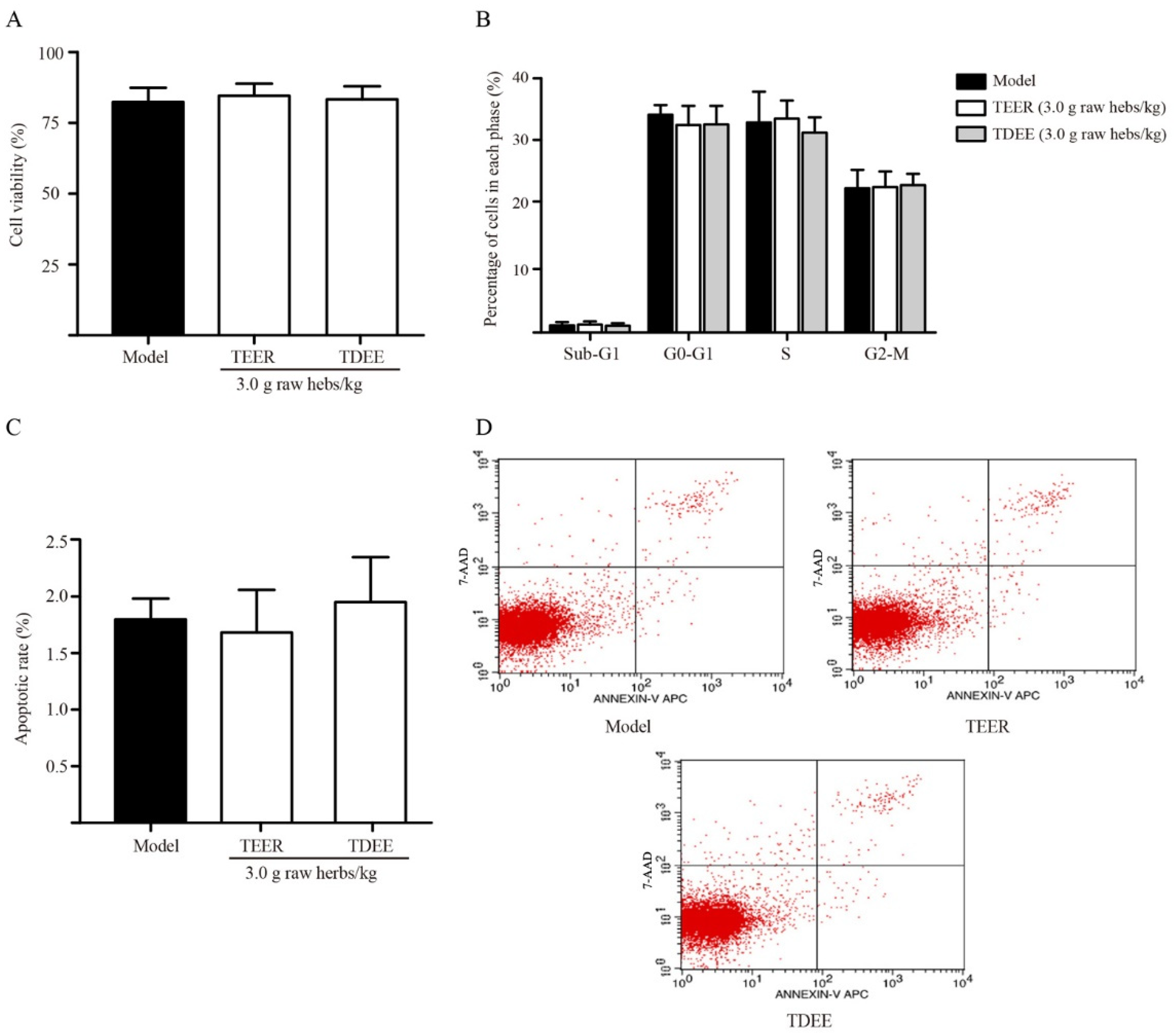
2.3. Effects of TDEE on Tumor Cell Viability, Cycle, and Apoptosis in Ascitic Mice
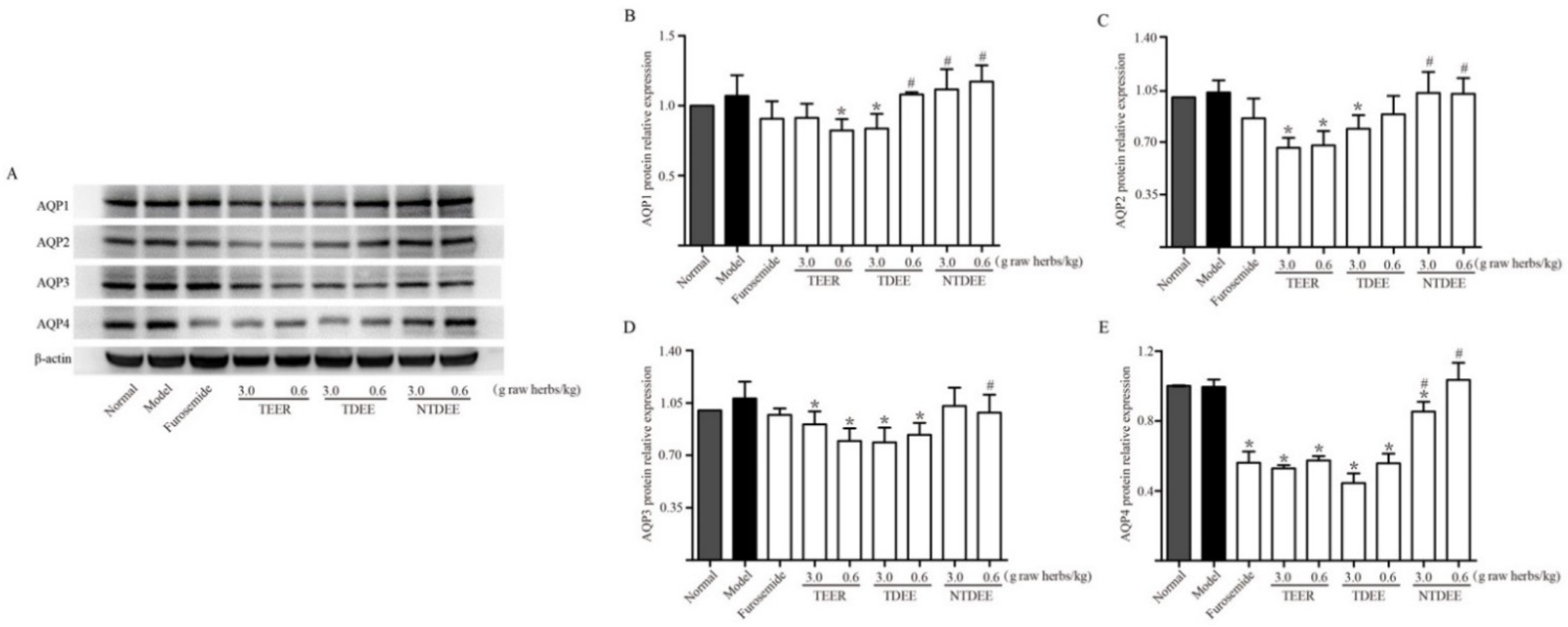
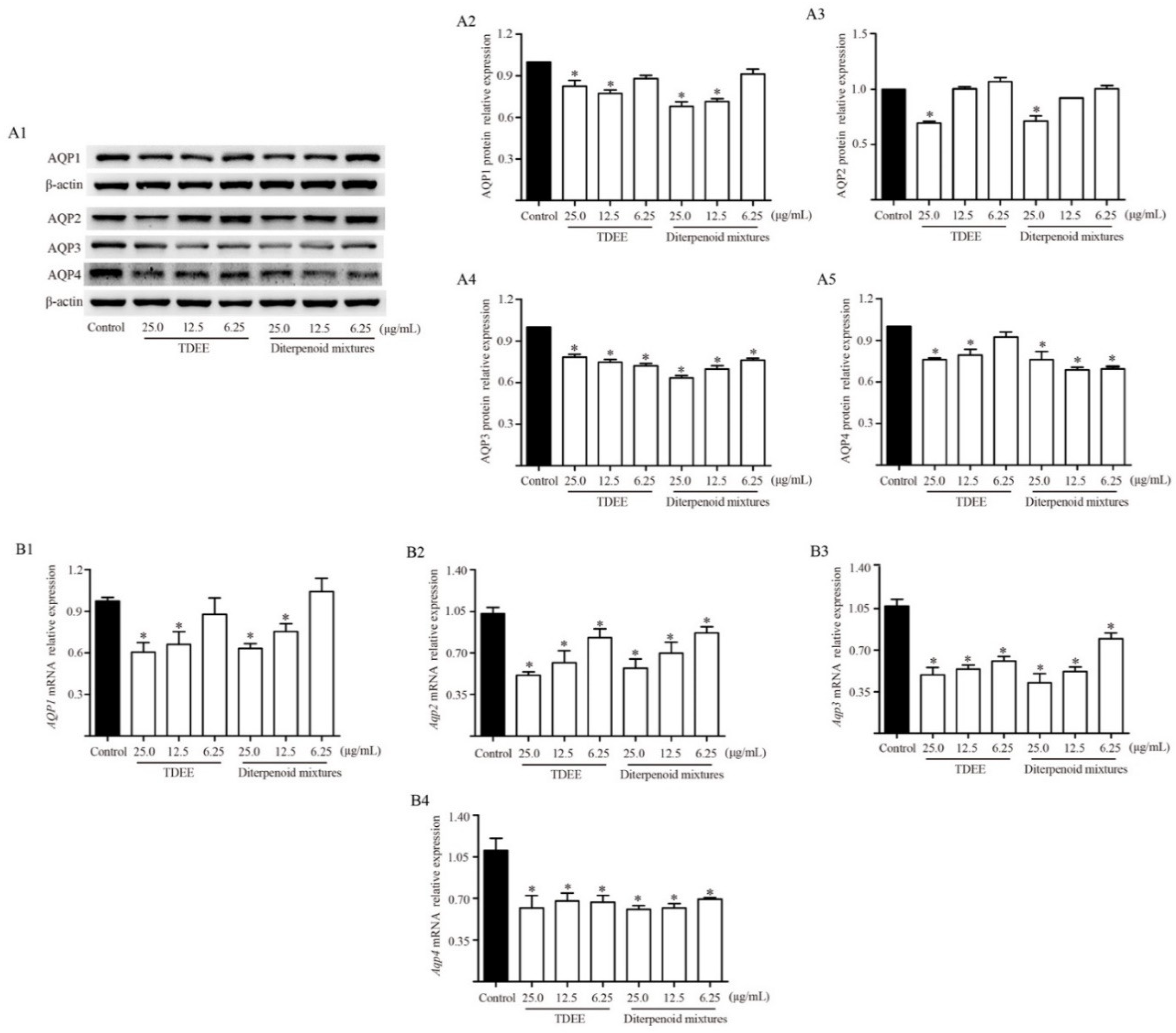
2.4. Effects of TDEE on AQPs Expression in the Kidneys of Ascitic Mice
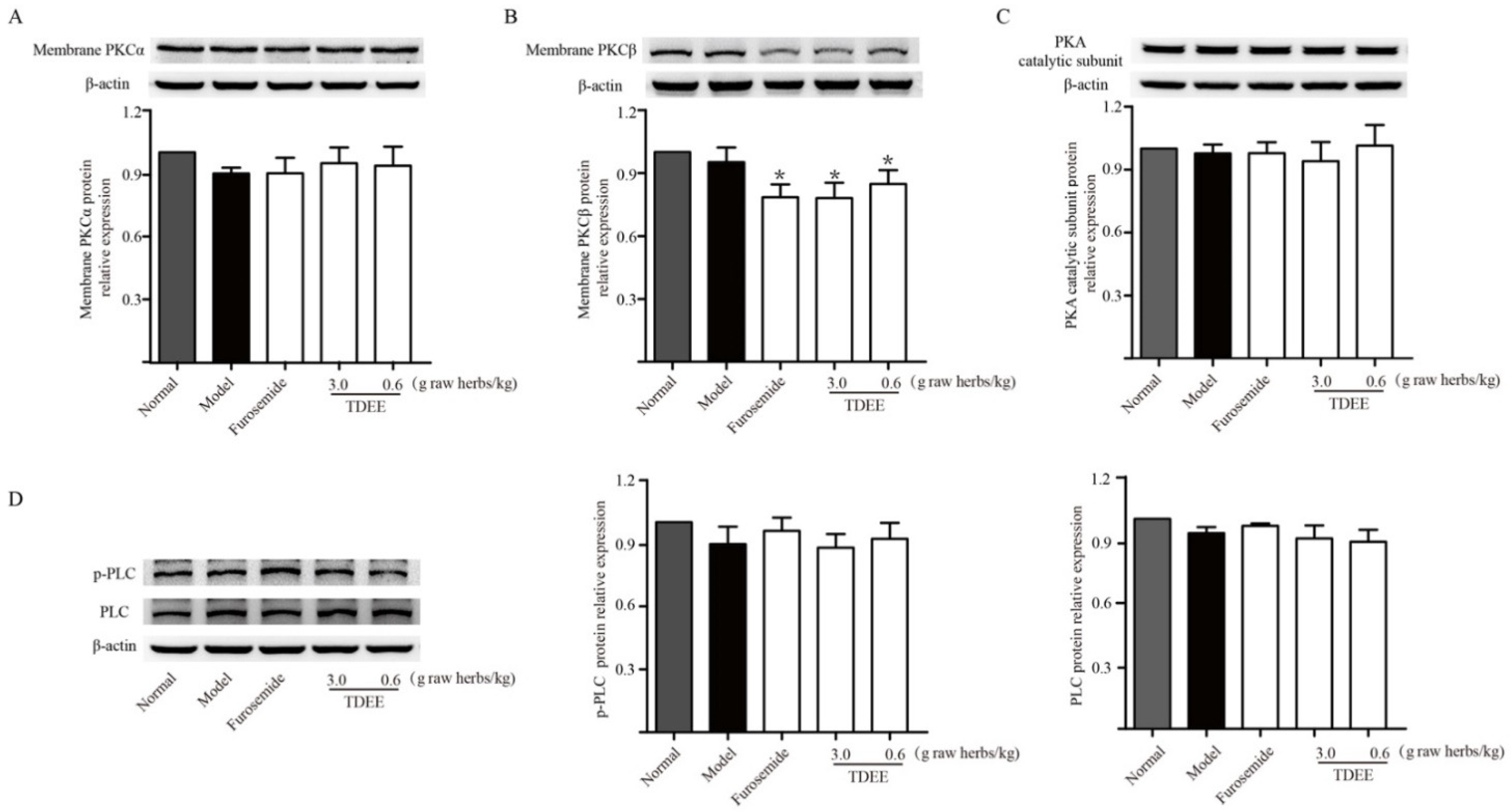
2.5. Effects of TDEE on PKC and PKA Activation in the Kidneys of Ascitic Mice
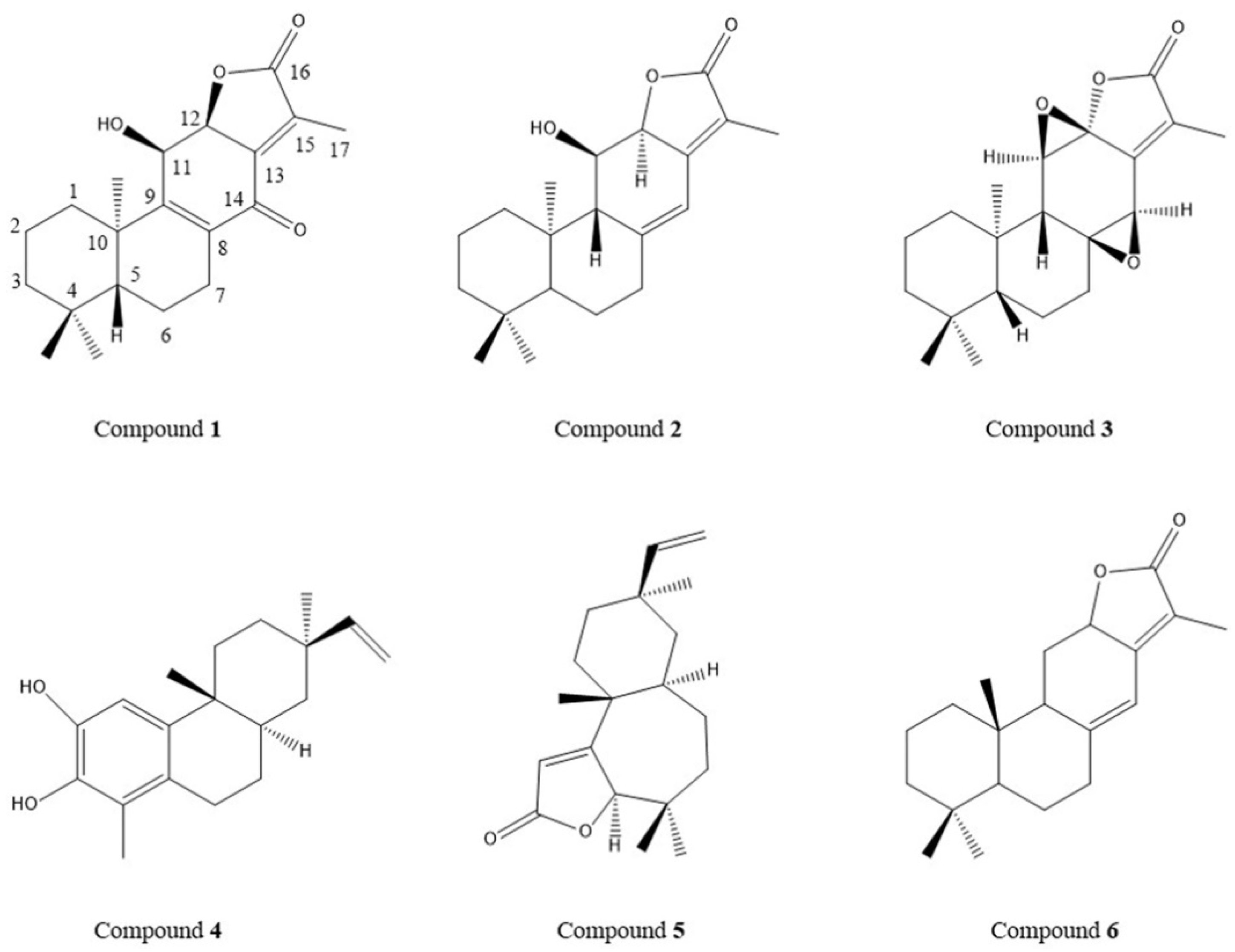
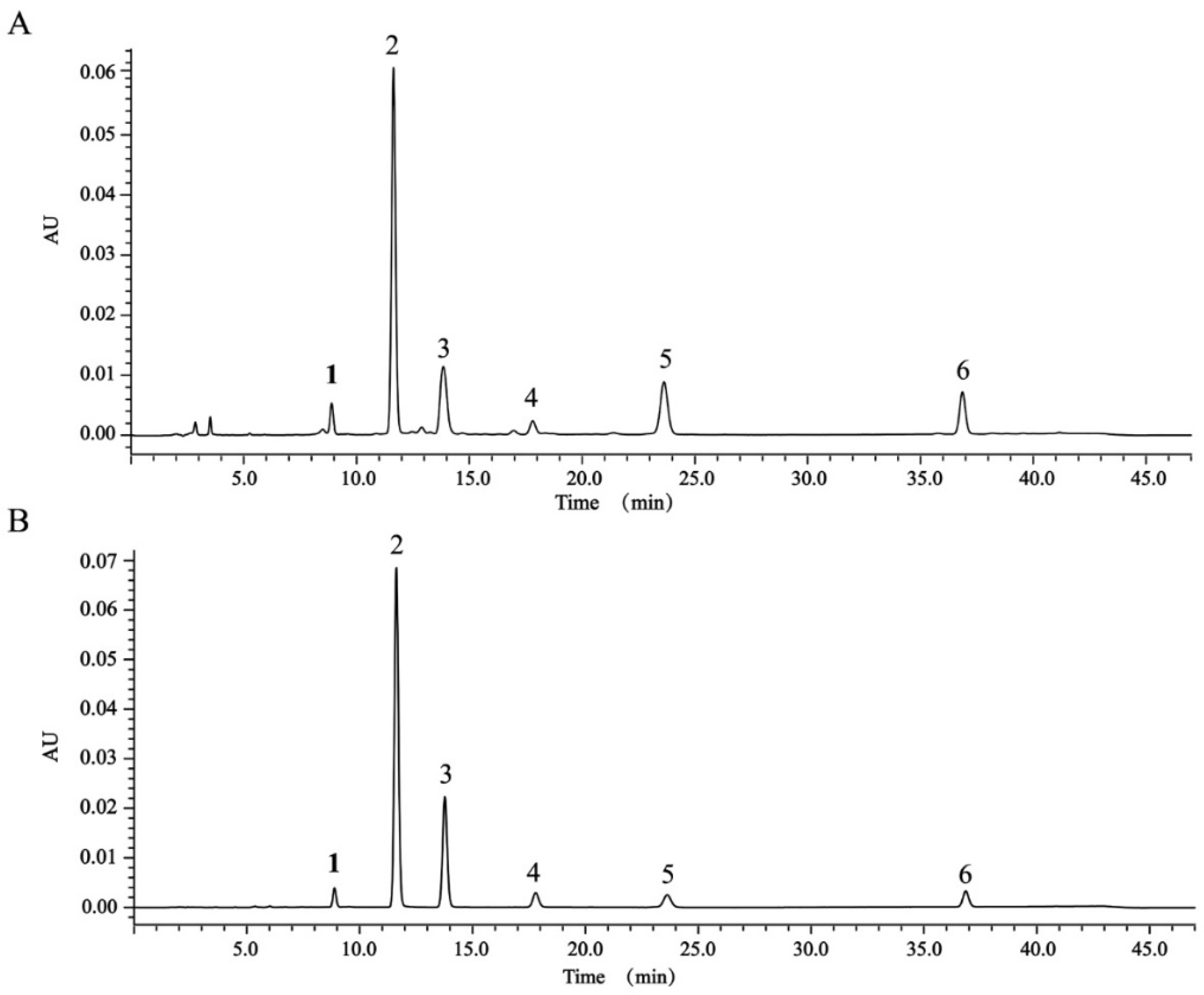
2.6. Determination of Content of Six Diterpenoids in TDEE by High-Performance Liquid Chromatography (HPLC)
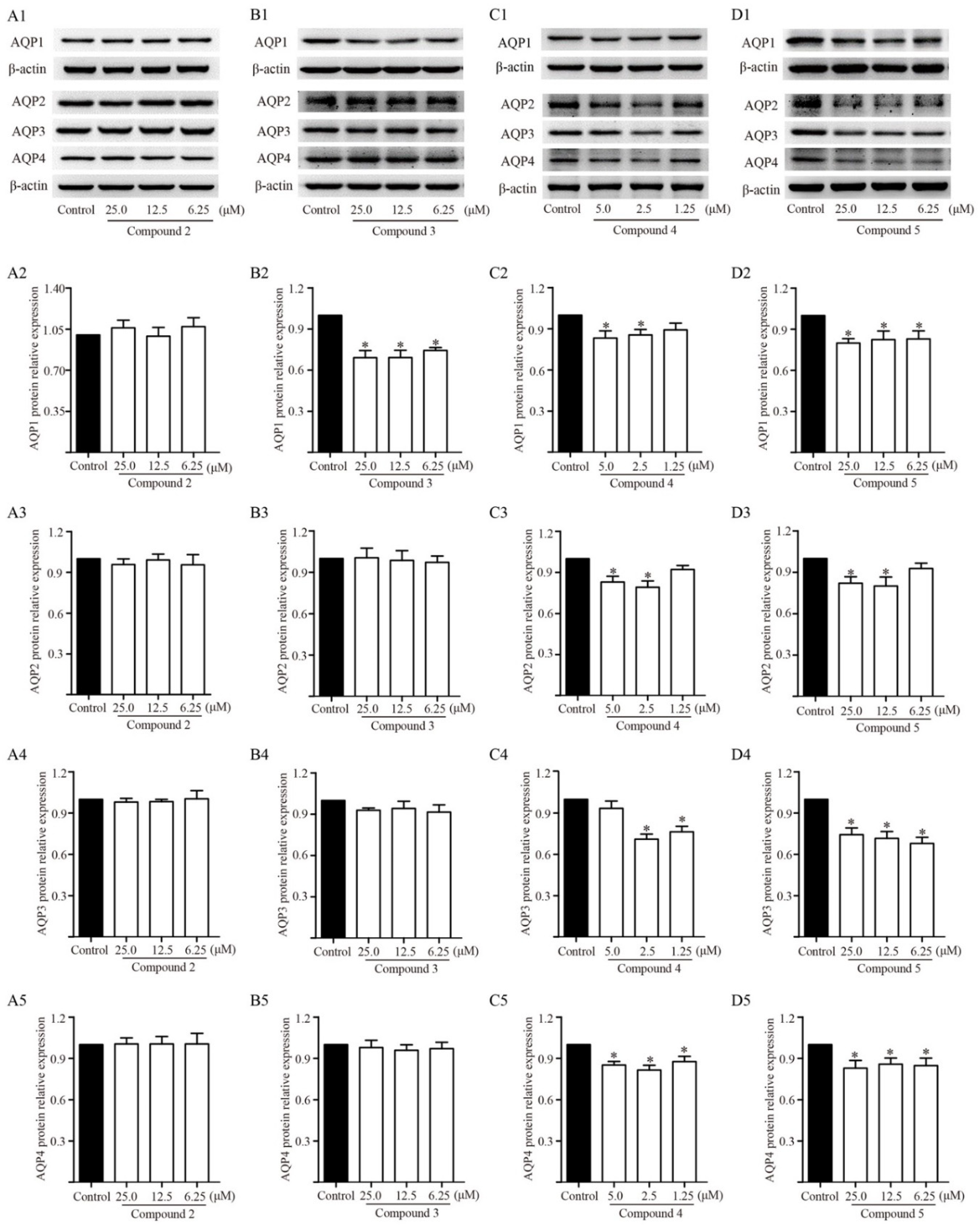
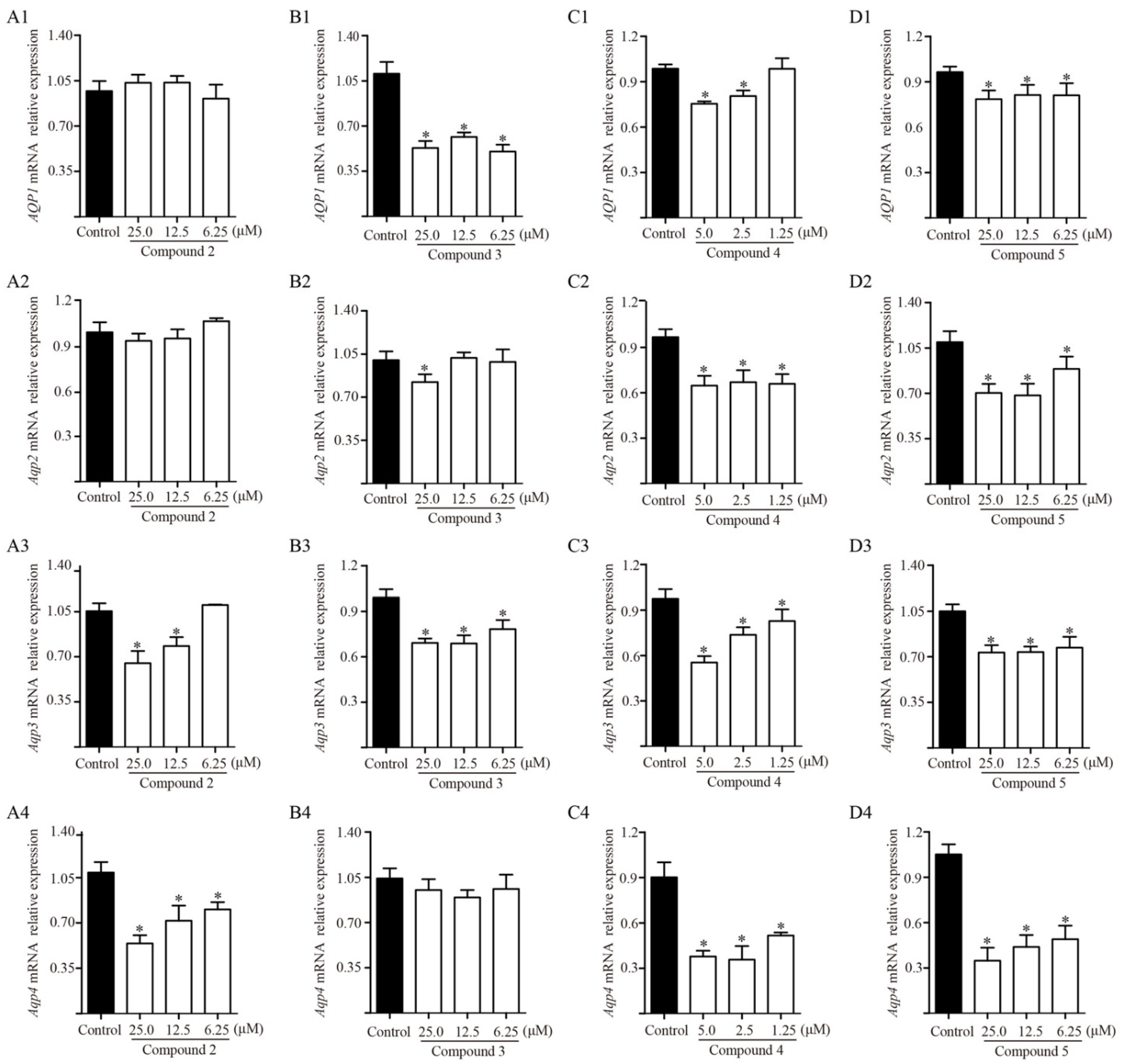
2.7. Effects of Diterpenoids from TDEE on AQPs Expression in Kidney Cells
2.8. Effects of Activator of PKC on Diterpenoid-Induced Inhibition of AQPs
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Plant Material
4.3. Extraction and Isolation of TDEE and Purification of Diterpenoids
4.4. In Vivo Experiments
4.4.1. Animals and Experimental Design
4.4.2. Ascitic Fluid Weight, Urine Weight, and Fecal Water Content Assays
4.4.3. Serum Albumin Analysis
4.4.4. Tumor Cell Viability, Cycle, and Apoptosis Assays
4.4.5. Western Blot Analysis of AQPs, PKC, PKA and PLC in the Kidneys
4.5. Determination of Six Major Diterpenoids in TDEE
4.6. In Vitro Experiments
4.6.1. Cell Culture
4.6.2. Western Blot and Quantitative Reverse Transcription-PCR (qRT-PCR) Analyses of AQPs
4.6.3. Effect of PKC agonist PMA on the AQPs Protein Expression in Kidney Cells
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Ye, D.; Yuan, S. Dictionary of Traditional Chinese Medicine Processing; Shanghai Scientific & Technical Publishers: Shanghai, China, 2005; p. 298. [Google Scholar]
- Yu, Z.; Wei, Y.; Tian, X.; Yan, Q.; Yan, Q.; Huo, X.; Wang, C.; Sun, C.; Zhang, B.; Ma, X. Diterpenoids from the roots of Euphorbia ebracteolata and their anti-tuberculosis effects. Bioorg. Chem. 2018, 77, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.S.; Lu, Y.; Chen, C.H.; Lee, K.H.; Chen, D.F. Potent Anti-HIV Ingenane Diterpenoids from Euphorbia ebracteolata. J. Nat. Prod. 2019, 82, 1587–1592. [Google Scholar] [CrossRef]
- Bai, J.; Huang, X.Y.; Liu, Z.G.; Gong, C.; Li, X.Y.; Li, D.H.; Hua, H.M.; Li, Z.L. Four new compounds from the roots of Euphorbia ebracteolata and their inhibitory effect on LPS-induced NO production. Fitoterapia 2018, 125, 235–239. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Peng, Y.; Wang, Y.; Huo, X.; Zhang, B.; Li, D.; Leng, A.; Zhang, H.; Ma, X.; Wang, C. Cytotoxic ent-Abietane-type diterpenoids from the roots of Euphorbia ebracteolata. Bioorg. Chem. 2018, 81, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Deng, T.; Wang, Y. Chinese Medical Classics; Hunan Electronic Audio & Video Publishers: Changsha, China, 2014. [Google Scholar]
- Liu, L.; Yu, H.L.; Wang, K.L.; Wu, H.; Wang, W.; Zhang, Y.B.; Yu, S.; Yin, H.Q. Effects of toxic fractions of Euphorbiae ebracteolatae Radix on toxicity of mice intestinal tract and colon aquaporins expression level before and after vinegar processing. China J. Chin. Mater. Med. 2018, 43, 2516–2521. [Google Scholar]
- Lenz, K.; Buder, R.; Kapun, L.; Voglmayr, M. Treatment and management of ascites and hepatorenal syndrome: An update. Ther. Adv. Gastroenter. 2015, 8, 83–100. [Google Scholar] [CrossRef] [Green Version]
- Gantt, C.L.; Ecklund, R.E. Significance of aldosterone antagonism in the treatment of edema and ascites. Am. J. Med. 1962, 33, 490–500. [Google Scholar] [CrossRef]
- Piano, S.; Tonon, M.; Angeli, P. Management of ascites and hepatorenal syndrome. Hepatol. Int. 2018, 12, 122–134. [Google Scholar] [CrossRef]
- Mobasheri, A.; Barrett-Jolley, R. Aquaporin Water Channels in the Mammary Gland: From Physiology to Pathophysiology and Neoplasia. J. Mammary Gland Biol. Neoplasia 2013, 19, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Esteva-Font, C.; Baccaro, M.E.; Fernández-Llama, P.; Sans, L.; Guevara, M.; Ars, E.; Jiménez, W.; Arroyo, V.; Ballarín, J.A.; Ginès, P. Aquaporin-1 and aquaporin-2 urinary excretion in cirrhosis: Relationship with ascites and hepatorenal syndrome. Hepatology 2006, 44, 1555–1563. [Google Scholar] [CrossRef]
- Ma, J.; Zhou, C.; Yang, J.; Ding, X.; Zhu, Y.; Chen, X. Expression of AQP6 and AQP8 in epithelial ovarian tumor. J. Mol. Histol. 2016, 47, 129–134. [Google Scholar] [CrossRef]
- Frede, J.; Fraser, S.P.; Oskay-Ozcelik, G.; Hong, Y.; Ioana Braicu, E.; Sehouli, J.; Gabra, H.; Djamgoz, M.B.A. Ovarian cancer: Ion channel and aquaporin expression as novel targets of clinical potential. Eur. J. Cancer 2013, 49, 2331–2344. [Google Scholar] [CrossRef] [PubMed]
- Sun, J. The Expression of Aquaporins in the Cirrhotic Rats Complicated with Ascites. Clin. Med. J. China 2004, 11, 1020–1022. [Google Scholar]
- Su, W.; Cao, R.; Zhang, X.Y.; Guan, Y. Aquaporins in the kidney: Physiology and pathophysiology. Am. J. Physiol. Renal Physiol. 2020, 318, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Patil, R.V. Protein kinase A-dependent phosphorylation of aquaporin-1. Biochem. Bioph. Res. Co. 2000, 273, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Shaw, S.; Marples, D. A rat kidney tubule suspension for the study of vasopressin-induced shuttling of AQP2 water channels. Am. J. Physiol. Renal Physiol. 2002, 283, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Li, X.D.; Yang, Y.J.; Geng, Y.J.; Cheng, Y.T.; Zhang, H.T.; Zhao, J.L.; Yuan, J.Q.; Gao, R.L. The cardioprotection of simvastatin in reperfused swine hearts relates to the inhibition of myocardial edema by modulating aquaporins via the PKA pathway. Int. J. Cardiol. 2013, 167, 2657–2666. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Yao, X.; Wang, T.X.; Jin, W.M.; Ji, Q.Q.; Yang, X.; Duan, Q.H.; Yao, L.J. PKCα regulates vasopressin-induced aquaporin-2 trafficking in mouse kidney collecting duct cells in vitro via altering microtubule assembly. Acta Pharmacol. Sin. 2012, 33, 230–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patra, S.; Muthuraman, M.S.; Prabhu, A.R.; Priyadharshini, R.R.; Parthiban, S. Evaluation of Antitumor and Antioxidant Activity of Sargassum tenerrimum against Ehrlich Ascites Carcinoma in Mice. Asian Pac. J. Cancer Prev. 2015, 16, 915–921. [Google Scholar] [CrossRef] [Green Version]
- Lou, J.W.; Cao, L.L.; Zhang, Q.; Jiang, D.J.; Yao, W.F.; Bao, B.H.; Cao, Y.D.; Tang, Y.P.; Zhang, L.; Wang, K. The toxicity and efficacy evaluation of different fractions of Kansui fry-baked with vinegar on Walker-256 tumor-bearing malignant ascites effusion rats and normal rats. J. Ethnopharmacol. 2018, 219, 257–268. [Google Scholar] [CrossRef]
- Yuan, W.J.; Ding, X.; Wang, Z.; Yang, B.J.; Li, X.N.; Zhang, Y.; Chen, D.Z.; Li, S.L.; Di, Y.T.; Hao, X.J.; et al. Two novel diterpenoid heterodimers, Bisebracteolasins A and B, from Euphorbia ebracteolata Hayata, and the cancer chemotherapeutic potential of Bisebracteolasin A. Sci. Rep. 2017, 7, 14507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, J.; Yan, S.; Chen, Y.; Han, L.; Wen, L.; Guo, X.; Wen, Y.; Li, Y.; He, X.; Han, Z.; et al. Aquaporin1-3 expression in normal and hydronephrotic kidneys in the human fetus. Pediatr. Res. 2019, 86, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Holmes, R.P. The role of renal water channels in health and disease. Mol. Asp. Med. 2012, 33, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Colombo, D.; Tringali, C.; Franchini, L.; Cirillo, F.; Venerando, B. Glycoglycerolipid analogues inhibit PKC translocation to the plasma membrane and downstream signaling pathways in PMA-treated fibroblasts and human glioblastoma cells, U87MG. Eur. J. Med. Chem. 2011, 46, 1827–1834. [Google Scholar] [CrossRef]
- Pany, S.; You, Y.; Das, J. Curcumin Inhibits Protein Kinase Cα Activity by Binding to Its C1 Domain. Biochemistry 2016, 55, 6327–6336. [Google Scholar] [CrossRef] [Green Version]
- Hadley, S.H.; Bahia, P.K.; Taylor-Clark, T.E. Sensory nerve terminal mitochondrial dysfunction induces hyperexcitability in airway nociceptors via protein kinase C. Mol. Pharm. 2014, 85, 839–848. [Google Scholar] [CrossRef] [Green Version]
- Theile, D.; Geng, S.; Denny, E.C.; Momand, J.; Kane, S.E. t-Darpp stimulates protein kinase A activity by forming a complex with its RI regulatory subunit. Cell. Signal. 2017, 40, 53–61. [Google Scholar] [CrossRef]
- Chen, Y. Basic Research of TCM Incompatibility Theory BASED on “Eighteen Incompatible Medicament”-Toxicity Effect Characterization and Basic Material Research on the Compatibility of Genkwa Flos and Glycyrrhizae Radix et Rhizoma. Ph.D. Thesis, Nanjing University Chinese Medicine, Nanjing, China, 2014. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′→3′) | Reverse (5′→3′) |
|---|---|---|
| Human AQP1 | TGCCATCGGCCTCTCTGTAG | AAGGACCGAGCAGGGTTAATC |
| Mouse Aqp2 | TGGCTGTCAATGCTCTCCAC | GGAGCAGCCGGTGAAATAGA |
| Mouse Aqp3 | GAATCGTTGTGGGGAGATGC | CAAGATGCCAAGGGTGACAG |
| Mouse Aqp4 | ATCAGCATCGCTAAGTCCGTC | GAGGTGTGACCAGGTAGAGGA |
| Human β-actin | GACCCAGATCATGTTTGAGAC | GTAGCCACGCTCGGTCAG |
| Mouse β-actin | TGGCTCCTAGCACCATGAAG | CCTGCTTGCTGATCCACATC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Liu, D.; Xue, F.; Yu, H.; Wu, H.; Cui, X.; Zhang, X.; Wang, H. Anti-Malignant Ascites Effect of Total Diterpenoids from Euphorbiae ebracteolatae Radix Is Attributable to Alterations of Aquaporins via Inhibiting PKC Activity in the Kidney. Molecules 2021, 26, 942. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040942
Zhang Y, Liu D, Xue F, Yu H, Wu H, Cui X, Zhang X, Wang H. Anti-Malignant Ascites Effect of Total Diterpenoids from Euphorbiae ebracteolatae Radix Is Attributable to Alterations of Aquaporins via Inhibiting PKC Activity in the Kidney. Molecules. 2021; 26(4):942. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040942
Chicago/Turabian StyleZhang, Yuanbin, Dongfang Liu, Fan Xue, Hongli Yu, Hao Wu, Xiaobing Cui, Xingde Zhang, and Hepeng Wang. 2021. "Anti-Malignant Ascites Effect of Total Diterpenoids from Euphorbiae ebracteolatae Radix Is Attributable to Alterations of Aquaporins via Inhibiting PKC Activity in the Kidney" Molecules 26, no. 4: 942. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26040942