
Aquaculture Production of the Brown Seaweeds Laminaria digitata and Macrocystis pyrifera: Applications in Food and Pharmaceuticals




Abstract
:1. Introduction
1.1. Laminaria digitata and Macrocystis pyrifera in the Context of Global Seaweed Aquaculture
1.2. Aquaculture methods for Brown Kelps
1.2.1. Macrocystis pyrifera
1.2.2. Laminaria digitata
2. Current and Future Uses
2.1. Food and Feed Uses of Brown Seaweeds
2.2. Pharmaceutical Uses of Brown Seaweeds
2.3. Other Uses of Brown Seaweeds
2.4. Functional Foods Applications
2.4.1. Protein Content and Applications
2.4.2. Carbohydrate Content and Applications
2.4.3. Tocols Applications
2.4.4. Pigment Applications
2.4.5. Phlorotannins Applications
2.4.6. Polysaccharides—Soluble Fibre Phycocolloids
2.4.7. Polysaccharides—Prebiotic Potential
2.4.8. Polyunsaturated Fatty Acids
3. Current Extraction Strategies
3.1. Seaweed Cell Wall Structure
3.2. Functional Food Extraction
3.2.1. Protein Extraction
3.2.2. Phlorotannins and Polyphenol Extraction
3.2.3. Carbohydrate Extraction
3.3. Pharmaceutical Extraction
3.3.1. Protein Extraction
3.3.2. Phlorotannins and Polyphenol Extraction
3.3.3. Carbohydrate Extraction
3.4. Extraction Improvements—Including Less Environmentally Impactful Strategies
Biorefinery Application—Optimised Biomass Utilisation
4. Regulation and Legislation
5. Delivery Methods and Applications of Seaweeds
5.1. Food and Feed Delivery and Applications
5.2. Pharmaceutical Delivery and Applications
5.3. Other Applications
5.3.1. Fuel
5.3.2. Nutricosmetics and Cosmeceuticals
5.3.3. Bioplastics
5.3.4. Bioremediation
5.3.5. Aquaculture
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- White, L.W.; Wilson, P. Chapter 2—World seaweed utilization. In Seaweed Sustainability; Tiwari, B.K., Troy, D.J., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 7–25. [Google Scholar] [CrossRef]
- Ferdouse, F.; Holdt, S.L.; Smith, R.; Murúa, P.; Yang, Z. The global status of seaweed production, trade and utilization. Globefish Res. Programme 2018, 124, I. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture. Contributing to Food Security and Nutrition for All; FAO: Rome, Italy, 2016; p. 200. [Google Scholar]
- Funge-Smith, S.J. Review of the state of world fishery resources: Inland fisheries. In FAO Fisheries and Aquaculture Circular; No.C942 Revision 3; FAO: Rome, Italy, 2018; p. 397. [Google Scholar]
- Barbier, M.; Charrier, B.; Araujo, R.; Holdt, S.L.; Jacquemin, B.; Rebours, C. Pegasus—Phycomorph European Guidelines for a Sustainable Aquaculture of Seaweeds; SBR: Roscoff, France, 2019. [Google Scholar]
- Hafting, J.T.; Craigie, J.S.; Stengel, D.B.; Loureiro, R.R.; Buschmann, A.H.; Yarish, C.; Edwards, M.D.; Critchley, A.T. Prospects and challenges for industrial production of seaweed bioactives. J. Phycol. 2015, 51, 821–837. [Google Scholar] [CrossRef] [PubMed]
- Suo, R.; Wang, Q. Laminaria culture in China. Infofish Int. 1992, 1, 40–42. [Google Scholar]
- Pereira, R.; Yarish, C. Mass production of marine macroalgae. In Encyclopedia of Ecology; Vol.3; Jørgensen, S.E., Fath, B.D., Eds.; Elsevier: Oxford, UK, 2008; pp. 2236–2247. [Google Scholar]
- Buschmann, A.H.; Camus, C. An introduction to farming and biomass utilisation of marine macroalgae. Phycologia 2019, 58, 443–445. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2018—Meeting the Sustainable Development Goals; FAO: Rome, Italy, 2018. [Google Scholar]
- Indegaard, M.; Minsaas, J. Seaweed Resources in Europe. Uses and Potential. In Animal and Human Nutrition; Guiry, M.D., Blunden, G., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 1991; pp. 21–64. [Google Scholar]
- Adams, N.M. Seaweeds of New Zealand. In An Illustrated Guide; Canterbury University Press: Christchurch, New Zealand, 1994; Chapter 5; p. 65. [Google Scholar]
- Nelson, W.A. New Zealand seaweeds. In An Illustrated Guide; Te Papa Press: Wellington, New Zealand, 2013. [Google Scholar]
- Morrissey, J.; Kraan, S.; Guiry, M.D. A Guide to Commercially Important Seaweeds on the Irish Coast; Irish Sea Fisheries Board: Dublin, Ireland, 2001; p. 67. [Google Scholar]
- Edwards, M.; Watson, L. Aquaculture Explained no. 26: Cultivating Laminaria digitata; Irish Sea Fisheries Board: Dublin, Ireland, 2011. [Google Scholar]
- Vásquez, J.A.; Piaget, N.; Vega, J.M.A. The Lessonia nigrescens fishery in northern Chile: How you harvest is more important than how much you harvest. J. Appl. Phycol. 2012, 24, 417–426. [Google Scholar] [CrossRef]
- Vásquez, J.A. Production, use and fate of Chilean brown seaweeds: Re-sources for a sustainable fishery. In Nineteenth International Seaweed Symposium; Springer: Dordrecht, The Netherlands, 2008; pp. 7–17. [Google Scholar]
- Camus, C.; Infante, J.; Buschmann, A.H. Overview of 3 year precommercial sea farming of Macrocystis pyrifera along the Chilean coast. Rev. Aquac. 2018, 10, 543–559. [Google Scholar] [CrossRef]
- Correa, T.; Gutiérrez, A.; Flores, R.; Buschmann, A.H.; Cornejo, P.; Bucarey, C. Production and economic assessment of giant kelp Macrocystis pyrifera cultivation for abalone feed in the south of Chile. Aquac. Res. 2016, 47, 698–707. [Google Scholar] [CrossRef]
- Tutschulte, T.C.; Connell, J.H. Feeding behavior and algal food of three species of abalones (Haliotis) in southern California. Mar. Ecol. Prog. Ser. 1988, 49, 57–64. [Google Scholar] [CrossRef]
- Castro, N.M.; Valdez, M.C.; Álvarez, A.M.; Ramírez, R.N.Á.; Rodríguez, I.S.; García, L.S. The Kelp Macrocystis pyrifera AS Nutritional Supplement for Goats. Rev. Científica 2009, 19, 63–70. [Google Scholar]
- Gojon-Baez, H.H.; Siqueiros-Beltrones, D.A.; Hernandez-Contreras, H. In situ ruminal digestibility and degradability of Macrocystis pyrifera and Sargassum spp. in bovine livestock. Cienc. Mar. 1998, 24, 463–481. [Google Scholar] [CrossRef] [Green Version]
- Mansilla, A.; Avila, M. Using Macrocystis pyrifera (L.) C. Agardh from southern Chile as a source of applied biological compounds. Rev. Bras. De Farmacogn. J. Pharmacogn. 2011, 21, 262–267. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.K.; Stekoll, M.; Yarish, C. Opportunities, challenges and future directions of open-water seaweed aquaculture in the United States. Phycologia 2019, 58, 446–461. [Google Scholar] [CrossRef]
- McHugh, D.J. A Guide to the Seaweed Industry; FAO Fisheries: Rome, Italy, 2003. [Google Scholar]
- Bailey, J. The History of Alginate Extraction. Available online: https://www.gracesguide.co.uk/The_History_of_Alginate_Extraction_by_Jim_Bailey (accessed on 24 August 2020).
- Kostas, E.T.; White, D.A.; Cook, D.J. Bioethanol Production from UK Seaweeds: Investigating Variable Pre-treatment and Enzyme Hydrolysis Parameters. Bioenergy Res. 2020, 13, 271–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. The State of World Fisheries and Aquaculture Opportunities and Challenges; Food and Agriculture Organization of the United Nations: Rome, Italy, 2014. [Google Scholar]
- Harger, B.W.W.; Neushul, M. Test-Farming of the Giant Kelp, Macrocystis, as a Marine Biomass Producer. J. World Maric. Soc. 1983, 14, 392–403. [Google Scholar] [CrossRef]
- Neushul, M. Marine Farming: Macroalgal Production and Genetics; Report # GRI 87/0070; Gas Research Institute: Chicago, IL, USA, 1986. [Google Scholar]
- Perissinotto, R.; McQuaid, C.D. Deep occurrence of the giant kelp Macrocystis laevis in the Southern Ocean. Mar. Ecol. Prog. Ser. Oldendorf 1992, 81, 89–95. [Google Scholar] [CrossRef]
- Buschmann, A.H.; Prescott, S.; Potin, P.; Faugeron, S.; Vásquez, J.A.; Camus, C.; Infante, J.; Hernández-González, M.C.; Gutíerrez, A.; Varela, D.A. Chapter Six - The Status of Kelp Exploitation and Marine Agronomy, with Emphasis on Macrocystis pyrifera, in Chile. In Advances in Botanical Research; Bourgougnon, N., Ed.; Academic Press Elsevier Ltd.: Cambridge, MA, USA, 2014; Volume 71, pp. 161–188. [Google Scholar] [CrossRef]
- Camus, C.; Infante, J.; Buschmann, A.H. Revisiting the economic profitability of giant kelp Macrocystis pyrifera (Ochrophyta) cultivation in Chile. Aquaculture 2019, 502, 80–86. [Google Scholar] [CrossRef]
- Avila-Peltroche, J.; Padilla-Vallejos, J. The seaweed resources of Peru. Bot. Mar. 2020, 63, 381–394. [Google Scholar] [CrossRef]
- North, W.J. Biology of the Macrocystis Resource in North America. In Case Studies of Seven Commercial Seaweed Resources; FAO Fisheries Technical Paper 281; M.Doty, M.S., Caddy, J.F., Santelices, B., Eds.; Food And Agriculture Organization: Rome, Italy, 1987; Available online: www.fao.org/docrep/X5819E/x5819e0a.htm (accessed on 24 February 2021).
- Petrell, R.J.; Tabrizi, K.M.; Harrison, P.J.; Druehl, L.D. Mathematical model of Laminaria production near a British Columbian salmon sea cage farm. J. Appl. Phycol. 1993, 5, 1–14. [Google Scholar] [CrossRef]
- Buschmann, A.H.; Varela, D.A.; Hernández-González, M.C.; Huovinen, P. Opportunities and challenges for the development of an integrated seaweed-based aquaculture activity in Chile: Determining the physiological capabilities of Macrocystis and Gracilaria as biofilters. J. Appl. Phycol. 2008, 20, 571–577. [Google Scholar] [CrossRef]
- Camus, C.; Buschmann, A.H. Macrocystis pyrifera aquafarming: Production optimization of rope-seeded juvenile sporophytes. Aquaculture 2017, 468, 107–114. [Google Scholar] [CrossRef]
- Sahoo, D.; Yarish, C. Mariculture of seaweeds. In Algal Culturing Techniques; Anderson, R.A., Ed.; Elsevier: Oxford, UK, 2005; pp. 219–237. [Google Scholar]
- Westermeier, R.; Patiño, D.; Piel, M.I.; Maier, I.; Mueller, D.G. A new approach to kelp mariculture in Chile: Production of free-floating sporophyte seedlings from gametophyte cultures of Lessonia trabeculata and Macrocystis pyrifera. Aquac. Res. 2006, 37, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Westermeier, R.; Patiño, D.J.; Murúa, P.; Müller, D.G. Macrocystis mariculture in Chile: Growth performance of heterosis genotype constructs under field conditions. J. Appl. Phycol. 2011, 23, 819–825. [Google Scholar] [CrossRef] [Green Version]
- Lüning, K. Critical levels of light and temperature regulating the gametogenesis of three Laminaria species (Phaeophyceae). J. Phycol. 1980, 16, 1–15. [Google Scholar] [CrossRef]
- Kregting, L.; Blight, A.J.; Elsäßer, B.; Savidge, G. The influence of water motion on the growth rate of the kelp Laminaria digitata. J. Exp. Mar. Biol. Ecol. 2016, 478, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Peteiro, C.; Bidegain, G.; Sánchez, N. Experimental evaluation of the effect of water velocity on the development of string-attached kelp seedlings (Laminariales) with implications for hatchery and nursery production. Algal Res. 2019, 44, 101678. [Google Scholar] [CrossRef]
- Dieck, T.I. Temperature tolerance and survival in darkness of kelp gametophyles (Laminariales, Phaeophyta): Ecological and biogeographical implications. Mar. Ecol. Prog. Ser. 1993, 100, 253–264. [Google Scholar] [CrossRef]
- Martins, N.; Tanttu, H.; Pearson, G.A.; Serrao, E.A.; Bartsch, I. Interactions of daylength, temperature and nutrients affect thresholds for life stage transitions in the kelp Laminaria digitata (Phaeophyceae). Bot. Mar. 2017, 60, 109–121. [Google Scholar] [CrossRef] [Green Version]
- Dieck, T.I. North Pacific and North Atlantic digitate Laminaria species (Phaeophyta): Hybridization experiments and temperature responses. Phycologia 1992, 31, 147–163. [Google Scholar]
- Raybaud, V.; Beaugrand, G.; Goberville, E.; Delebecq, G.; Destombe, C.; Valero, M.; Davoult, D.; Morin, P.; Gevaert, F. Decline in Kelp in West Europe and Climate. PLoS ONE 2013, 8, e66044. [Google Scholar] [CrossRef] [Green Version]
- Fleurence, J.; Morançais, M.; Dumay, J.; Decottignies, P.; Turpin, V.; Munier, M.; Garcia-Bueno, N.; Jaouen, P. What are the prospects for using seaweed in human nutrition and for marine animals raised through aquaculture? Trends Food Sci. Technol. 2012, 27, 57–61. [Google Scholar] [CrossRef]
- Burtin, P. Nutritional Value of Seaweeds. Electron. J. Environ. Agric. Food Chem. 2003, 2, 498–503. [Google Scholar]
- Marsham, S.; Scott, G.W.; Tobin, M.L. Comparison of nutritive chemistry of a range of temperate seaweeds. Food Chem. 2007, 100, 1331–1336. [Google Scholar] [CrossRef]
- Taboada, M.C.; Millán, R.; Miguez, M.I. Nutritional value of the marine algae wakame (Undaria pinnatifida) and nori (Porphyra purpurea) as food supplements. J. Appl. Phycol. 2013, 25, 1271–1276. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Rajauria, G. Chapter 15—Seaweeds: A sustainable feed source for livestock and aquaculture. In Seaweed Sustainability; Tiwari, B.K., Troy, D.J., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 389–420. [Google Scholar] [CrossRef]
- Schiener, P.; Black, K.D.; Stanley, M.S.; Green, D.H. The seasonal variation in the chemical composition of the kelp species Laminaria digitata, Laminaria hyperborea, Saccharina latissima and Alaria esculenta. J. Appl. Phycol. 2015, 27, 363–373. [Google Scholar] [CrossRef]
- Bixler, H.J.; Porse, H. A decade of change in the seaweed hydrocolloids industry. J. Appl. Phycol. 2011, 23, 321–335. [Google Scholar] [CrossRef]
- MacArtain, P.; Gill, C.I.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65, 535–543. [Google Scholar] [CrossRef]
- Food and Drink Federation. Science behind Guideline Daily Amounts; Food and Drink Federation: London, UK, 2009. [Google Scholar]
- Institut de Phytonutrition. Functional, Health and Therapeutic Effects of Algae and Seaweed; Version 1.5; Institut de Phytonutrition Electronic Database: Beausoleil, France, 2004. [Google Scholar]
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological Activities of Fucoidan and the Factors Mediating Its Therapeutic Effects: A Review of Recent Studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef] [Green Version]
- Bor, S.; Kalkan, İ.H.; Çelebi, A.; Dinçer, D.; Akyüz, F.; Dettmar, P.; Özen, H. Alginates: From the ocean to gastroesophageal reflux disease treatment. Turk. J. Gastroenterol. 2019, 30, 109–136. [Google Scholar] [CrossRef]
- Stiger-Pouvreau, V.; Bourgougnon, N.; Deslandes, E. Chapter 8—Carbohydrates From Seaweeds. In Seaweed in Health and Disease Prevention; Fleurence, J., Levine, I., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 223–274. [Google Scholar] [CrossRef]
- Astorga-España, M.S.; Mansilla, A.; Ojeda, J.; Marambio, J.; Rosenfeld, S.; Mendez, F.; Rodriguez, J.P.; Ocaranza, P. Nutritional properties of dishes prepared with sub-Antarctic macroalgae—An opportunity for healthy eating. J. Appl. Phycol. 2017, 29, 2399–2406. [Google Scholar] [CrossRef]
- Kim, H.W.; Choi, J.H.; Choi, Y.S.; Han, D.J.; Kim, H.Y.; Lee, M.A.; Kim, S.Y.; Kim, C.J. Effects of Sea Tangle (Lamina japonica) Powder on Quality Characteristics of Breakfast Sausages. Korean J. Food Sci. Anim. Resour. 2010, 30, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Codex Stan. Codex Alimentarius. Standard for Named Vegetable Oils; FAO/OMS: Rome, Italy, 1999; pp. 1–15. [Google Scholar]
- Ortiz, J.; Uquiche, E.; Robert, P.; Romero, N.; Quitral, V.; Llantén, C. Functional and nutritional value of the Chilean seaweeds Codium fragile, Gracilaria chilensis and Macrocystis pyrifera. Eur. J. Lipid Sci. Technol. 2009, 111, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Cruz-Suarez, L.E.; Tapia-Salazar, M.; Nieto-LÓPez, M.G.; Guajardo-Barbosa, C.; Ricque-Marie, D. Comparison of Ulva clathrata and the kelps Macrocystis pyrifera and Ascophyllum nodosum as ingredients in shrimp feeds. Aquac. Nutr. 2009, 15, 421–430. [Google Scholar] [CrossRef]
- Hansen, H.R.; Hector, B.L.; Feldmann, J. A qualitative and quantitative evaluation of the seaweed diet of North Ronaldsay sheep. Anim. Feed Sci. Technol. 2003, 105, 21–28. [Google Scholar] [CrossRef]
- Tang, Z.L.; Shen, S.F. A study of Laminaria digitata powder on experimental hyperlipoproteinemia and its hemorrheology. Zhong Xi Yi Jie He Za Zhi, 9 April 1989; 223–225, 198. [Google Scholar]
- Gahan, D.A.; Lynch, M.B.; Callan, J.J.; O’Sullivan, J.T.; O’Doherty, J.V. Performance of weanling piglets offered low-, medium- or high-lactose diets supplemented with a seaweed extract from Laminaria spp. Animal 2009, 3, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, Á.R.; Gadicke, P.; Andrades, S.M.; Cubillos, R. Supplementing nursery pig feed with seaweed extracts increases final body weight of pigs. Austral. J. Vet. Sci. 2018, 50, 83–87. [Google Scholar] [CrossRef]
- López, G.; Alujas, A.; Biannic, O.; Gallissot, M.; Laurain, J. Influence of an algae-based complementary feed on the development of the small intestine of piglets during lactation, Journées De La Rech. Porc. En Fr. 2014, 46, 83–84. [Google Scholar]
- Leonard, S.G.; Sweeney, T.; Bahar, B.; O’Doherty, J.V. Effect of maternal seaweed extract supplementation on suckling piglet growth, humoral immunity, selected microflora, and immune response after an ex vivo lipopolysaccharide challenge. J. Anim. Sci. 2012, 90, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Reilly, P.; O’Doherty, J.V.; Pierce, K.M.; Callan, J.J.; O’Sullivan, J.T.; Sweeney, T. The effects of seaweed extract inclusion on gut morphology, selected intestinal microbiota, nutrient digestibility, volatile fatty acid concentrations and the immune status of the weaned pig. Animal 2008, 2, 1465–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, T.; Collins, C.B.; Reilly, P.; Pierce, K.M.; Ryan, M.; O’Doherty, J.V. Effect of purified β-glucans derived from Laminaria digitata, Laminaria hyperborea and Saccharomyces cerevisiae on piglet performance, selected bacterial populations, volatile fatty acids and pro-inflammatory cytokines in the gastrointestinal tract of pigs. Br. J. Nutr. 2012, 108, 1226–1234. [Google Scholar] [CrossRef] [Green Version]
- Moroney, N.C.; O’Grady, M.N.; O’Doherty, J.V.; Kerry, J.P. Addition of seaweed (Laminaria digitata) extracts containing laminarin and fucoidan to porcine diets: Influence on the quality and shelf-life of fresh pork. Meat Sci. 2012, 92, 423–429. [Google Scholar] [CrossRef]
- Moroney, N.C.; O’Grady, M.N.; O’Doherty, J.V.; Kerry, J.P. Effect of a brown seaweed (Laminaria digitata) extract containing laminarin and fucoidan on the quality and shelf-life of fresh and cooked minced pork patties. Meat Sci. 2013, 94, 304–311. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Drug Definition US FDA Drug Approval Process. Available online: https://www.pharmacistspharmajournal.org/2010/11/definitions-of-drug-radioactive-drug_11.html#.Xv3bAUBuJPY (accessed on 2 July 2020).
- Hoppe, H.A. Marine algae and their products and constituents in pharmacy. In Marine Algae and Pharmaceutical Science; Vol.1; Hoppe, H.A., Levring, T., Tanaka, Y., Eds.; W. de Gruyter: New York, NY, USA, 1979; pp. 25–119. [Google Scholar]
- Kılınç, B.; Cirik, S.; Turan, G.; Tekogul, H.; Koru, E. Seaweeds for Food and Industrial Applications. In Food Industry; Muzzalupo, I., Ed.; Intech Open: London, UK, 2013; pp. 735–747. [Google Scholar] [CrossRef] [Green Version]
- Qin, Y. Alginate fibres: An overview of the production processes and applications in wound management. Polym. Int. 2008, 57, 171–180. [Google Scholar] [CrossRef]
- Tønnesen, H.H.; Karlsen, J. Alginate in Drug Delivery Systems. Drug Dev. Ind. Pharm. 2002, 28, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Fertah, M.; Belfkira, A.; Dahmane, E.M.; Taourirte, M.; Brouillette, F. Extraction and characterization of sodium alginate from Moroccan Laminaria digitata brown seaweed. Arab. J. Chem. 2017, 10, S3707–S3714. [Google Scholar] [CrossRef] [Green Version]
- Synytsya, A.; Čopíková, J.; Kim, W.J.; Park, Y.I. Cell wall polysaccharides of marine algae. In Springer Handbook of Marine Biotechnology; Kim, S.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 543–590. [Google Scholar]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.-K.; Jung, U.; Roh, C. Fucoidan from marine brown algae inhibits lipid accumulation. Mar. Drugs 2011, 9, 1359–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S. Therapies from Fucoidan: An Update. Mar. Drugs 2015, 13, 5920–5946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, R.; Azuma, Y.; Ojima, T.; Hashimoto, T.; Mizuno, M.; Nishitani, Y.; Yoshida, M.; Azuma, T.; Kanazawa, K. Modulation of platelet aggregation-related eicosanoid production by dietary F-fucoidan from brown alga Laminaria japonica in human subjects. Br. J. Nutr. 2013, 110, 880–890. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Oda, T.; Yu, Q.; Jin, J.-O. Fucoidan from Macrocystis pyrifera Has Powerful Immune-Modulatory Effects Compared to Three Other Fucoidans. Mar. Drugs 2015, 13, 1084–1104. [Google Scholar] [CrossRef] [Green Version]
- Grundy, S.M. Drug therapy of the metabolic syndrome: Minimizing the emerging crisis in polypharmacy. Nat. Rev. Drug Discov. 2006, 5, 295–309. [Google Scholar] [CrossRef] [PubMed]
- Shan, X.; Liu, X.; Hao, J.; Cai, C.; Fan, F.; Dun, Y.; Zhao, X.; Li, C.; Yu, G. In vitro and in vivo hypoglycemic effects of brown algal fucoidans. Int. J. Biol. Macromol. 2016, 82, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Zheng, Y.; Zhang, Q.; Wang, J.; Wang, L.; Yang, W.; Guo, C.; Gao, W.; Wang, X.; Luo, D. Low-molecular-weight fucoidan protects endothelial function and ameliorates basal hypertension in diabetic Goto-Kakizaki rats. Lab. Invest. 2014, 94, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.W.; Liu, Q.; Thorlacius, H. Inhibition of selectin function and leukocyte rolling protects against dextran sodium sulfate-induced murine colitis. Scand. J. Gastroenterol. 2001, 36, 270–275. [Google Scholar] [CrossRef]
- Matsumoto, S.; Nagaoka, M.; Hara, T.; Kimura-Takagi, I.; Mistuyama, K.; Ueyama, S. Fucoidan derived from Cladosiphon okamuranus Tokida ameliorates murine chronic colitis through the down-regulation of interleukin-6 production on colonic epithelial cells. Clin. Exp. Immunol. 2004, 136, 432–439. [Google Scholar] [CrossRef]
- Cong, Q.; Chen, H.; Liao, W.; Xiao, F.; Wang, P.; Qin, Y.; Dong, Q.; Ding, K. Structural characterization and effect on anti-angiogenic activity of a fucoidan from Sargassum fusiforme. Carbohydr. Polym. 2016, 136, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jónsdóttir, R.; Liu, H.; Gu, L.; Kristinsson, H.G.; Raghavan, S.; Ólafsdóttir, G. Antioxidant Capacities of Phlorotannins Extracted from the Brown Algae Fucus vesiculosus. J. Agric. Food Chem. 2012, 60, 5874–5883. [Google Scholar] [CrossRef] [PubMed]
- Hwang, P.-A.; Hung, Y.-L.; Phan, N.N.; Hieu, B.-T.-N.; Chang, P.-M.; Li, K.-L.; Lin, Y.-C. The in vitro and in vivo effects of the low molecular weight fucoidan on the bone osteogenic differentiation properties. Cytotechnology 2016, 68, 1349–1359. [Google Scholar] [CrossRef]
- Strain, C.R.; Collins, K.C.; Naughton, V.; McSorley, E.M.; Stanton, C.; Smyth, T.J.; Soler-Vila, A.; Rea, M.C.; Ross, P.R.; Cherry, P.; et al. Effects of a polysaccharide-rich extract derived from Irish-sourced Laminaria digitata on the composition and metabolic activity of the human gut microbiota using an in vitro colonic model. Eur. J. Nutr. 2020, 59, 309–325. [Google Scholar] [CrossRef] [Green Version]
- Copp, A.; Glantz, D. Macrocystis pyrifera Derived Health and Wellness Products and Methods of Using the Same. Patent No. WO 2011/044267 Al, 14 April 2011. [Google Scholar]
- SpecialChem2020. Actipone® Laminaria Digitata GW. Available online: https://cosmetics.specialchem.com/product/i-symrise-actipone-laminaria-digitata-gw (accessed on 6 August 2020).
- Boiron—Laminaria Digitata Mother Tincture. Available online: https://www.moncoinsante.co.uk/laminaria-digitata-tm-teinture-mere-boiron-125ml.html (accessed on 6 August 2020).
- Pereira, L. Therapeutic and Nutritional Uses of Algae; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Thomas, N.V.; Kim, S.K. Beneficial effects of marine algal compounds in cosmeceuticals. Mar. Drugs 2013, 11, 146–164. [Google Scholar] [CrossRef] [Green Version]
- Stengel, D.B.; Connan, S.; Popper, Z.A. Algal chemodiversity and bioactivity: Sources of natural variability and implications for commercial application. Biotechnol. Adv. 2011, 29, 483–501. [Google Scholar] [CrossRef]
- Sanjeewa, K.K.A.; Kim, E.A.; Son, K.T.; Jeon, Y.J. Bioactive properties and potentials cosmeceutical applications of phlorotannins isolated from brown seaweeds: A review. J. Photochem. Photobiol. B 2016, 162, 100–105. [Google Scholar] [CrossRef]
- Jacobsen, C.; Sørensen, A.-D.M.; Holdt, S.L.; Akoh, C.C.; Hermund, D.B. Source, Extraction, Characterization, and Applications of Novel Antioxidants from Seaweed. Annu. Rev. Food Sci. Technol. 2019, 10, 541–568. [Google Scholar] [CrossRef]
- Thomas, N.V.; Kim, S.K. Potential pharmacological applications of polyphenolic derivatives from marine brown algae. Env. Toxicol. Pharm. 2011, 32, 325–335. [Google Scholar] [CrossRef]
- Shin, T.; Ahn, M.; Hyun, J.W.; Kim, S.H.; Moon, C. Antioxidant marine algae phlorotannins and radioprotection: A review of experimental evidence. Acta Histochem. 2014, 116, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Ragan, M.A.; Glombitza, K.W. Phlorotannins, brown algal polyphenols. In Progress in Phycological Research; Hellebust, J.A., Craigie, J.S., Eds.; Cambridge University Press: New York, NY, USA, 1986. [Google Scholar]
- Ferreres, F.; Lopes, G.; Gil-Izquierdo, A.; Andrade, P.B.; Sousa, C.; Mouga, T.; Valentão, P. Phlorotannin extracts from fucales characterized by HPLC-DAD-ESI-MSn: Approaches to hyaluronidase inhibitory capacity and antioxidant properties. Mar. Drugs 2012, 10, 2766–2781. [Google Scholar] [CrossRef] [Green Version]
- Vissers, A.M.; Caligiani, A.; Sforza, S.; Vincken, J.-P.; Gruppen, H. Phlorotannin Composition of Laminaria digitata. Phytochem. Anal. 2017, 28, 487–495. [Google Scholar] [CrossRef]
- Yang, H.; Zeng, M.; Dong, S.; Liu, Z.; Li, R. Anti-proliferative activity of phlorotannin extracts from brown algae Laminaria japonica Aresch. Chin. J. Oceanol. Limnol. 2010, 28, 122–130. [Google Scholar] [CrossRef]
- Halm, H.; Lüder, U.H.; Wiencke, C. Induction of phlorotannins through mechanical wounding and radiation conditions in the brown macroalga Laminaria hyperborea. Eur. J. Phycol. 2011, 46, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Leyton, A.; Pezoa-Conte, R.; Barriga, A.; Buschmann, A.H.; Mäki-Arvela, P.; Mikkola, J.P.; Lienqueo, M.E. Identification and efficient extraction method of phlorotannins from the brown seaweed Macrocystis pyrifera using an orthogonal experimental design. Algal Res. 2016, 16, 201–208. [Google Scholar] [CrossRef]
- Myers, S.P.; Mulder, A.M.; Baker, D.G.; Robinson, S.R.; Rolfe, M.I.; Brooks, L.; Fitton, J.H. Effects of fucoidan from Fucus vesiculosus in reducing symptoms of osteoarthritis: A randomized placebo-controlled trial. Biologics 2016, 10, 81–88. [Google Scholar] [PubMed] [Green Version]
- Myers, S.P.; O’Connor, J.; Fitton, J.H.; Brooks, L.; Rolfe, M.; Connellan, P.; Wohlmuth, H.; Cheras, P.A.; Morris, C. A combined phase I and II open label study on the effects of a seaweed extract nutrient complex on osteoarthritis. Biol. Targets. 2010, 4, 33–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horn, S.J.; Aasen, I.M.; Østgaard, K. Ethanol production from seaweed extract. J. Ind. Microbiol. Biotechnol. 2000, 25, 249–254. [Google Scholar] [CrossRef]
- Østgaard, K.; Indergaard, M.; Markussen, S.; Knutsen, S.H.; Jensen, A. Carbohydrate degradation and methane production during fermentation ofLaminaria saccharina (Laminariales, Phaeophyceae). J. Appl. Phycol. 1993, 5, 333–342. [Google Scholar] [CrossRef]
- Stirk, W.A.; van Staden, J. Removal of Heavy Metals from Solution Using Dried Brown Seaweed Material. Bot. Mar. 2000, 43, 467–473. [Google Scholar] [CrossRef]
- Jin-Fen, P.; Rong-Gen, L.; Li, M. A review of heavy metal adsorption by marine algae. Chin. J. Oceanol. Limnol. 2000, 18, 260–264. [Google Scholar] [CrossRef]
- Papageorgiou, S.K.; Kouvelos, E.P.; Katsaros, F.K. Calcium alginate beads from Laminaria digitata for the removal of Cu+2 and Cd+2 from dilute aqueous metal solutions. Desalination 2008, 224, 293–306. [Google Scholar] [CrossRef]
- Malinowska, P. Algae extracts as active cosmetic ingredients. Zesz. Nauk. /Uniw. Ekon. W Pozn. 2011, 212, 123–129. [Google Scholar]
- Dini, I.; Laneri, S. Nutricosmetics: A brief overview. Phytother. Res. 2019, 33, 3054–3063. [Google Scholar] [CrossRef]
- Cerná, M. Seaweed proteins and amino acids as nutraceuticals. Adv. Food Nutr. Res. 2011, 64, 297–312. [Google Scholar]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Seaweeds as nutraceuticals for health and nutrition. Phycologia 2019, 58, 563–577. [Google Scholar] [CrossRef] [Green Version]
- Becker, L.C.; Cherian, P. Safety Assessment of Brown Algae-Derived Ingredients as Used in Cosmetics; Cosmetic Ingredient Review: Washington, DC, USA, 2018; p. 161. [Google Scholar]
- Chojnacka, K.; Saeid, A.; Witkowska, Z.; Tuhy, L. Biologically Active Compounds in Seaweed Extracts—The Prospects for the Application. Open Conf. Proc. J. 2012, 3, 20–28. [Google Scholar] [CrossRef] [Green Version]
- Pangestuti, R.; Kim, S.-K. Chapter 6—Seaweed proteins, peptides, and amino acids. In Seaweed Sustainability; Tiwari, B.K., Troy, D.J., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 125–140. [Google Scholar] [CrossRef]
- Schmidt Hebbel, H.; Pennacchiotti Monti, I.; Masson Salaué, L.; Mella Rojas, M.A. Tabla de Composición Química de Alimentos Chilenos; Universidad de Chile: Santiago, Chile, 1990. [Google Scholar]
- Albanes, D.; Heinonen, O.P.; Huttunen, J.K.; Taylor, P.R.; Virtamo, J.; Edwards, B.K.; Haapakoski, J.; Rautalahti, M.; Hartman, A.M.; Palmgren, J. Effects of alpha-tocopherol and beta-carotene supplements on cancer incidence in the Alpha-Tocopherol Beta-Carotene Cancer Prevention Study. Am. J. Clin. Nutr. 1995, 62, 1427S–1430S. [Google Scholar] [CrossRef]
- Gey, F.K. Vitamins E plus C and interacting conutrients required for optimal health. BioFactors 1998, 7, 113–174. [Google Scholar]
- Albanes, D.; Heinonen, O.P.; Taylor, P.R.; Virtamo, J.; Edwards, B.K.; Rautalahti, M.; Hartman, A.M.; Palmgren, J.; Freedman, L.S.; Haapakoski, J.; et al. α-Tocopherol and β-Carotene Supplements and Lung Cancer Incidence in the Alpha-Tocopherol, Beta-Carotene Cancer Prevention Study: Effects of Base-line Characteristics and Study Compliance. J. Natl. Cancer Inst. 1996, 88, 1560–1570. [Google Scholar] [CrossRef]
- Leo, M.A.; Kim, C.; Lowe, N.; Lieber, C.S. Interaction of ethanol with beta-carotene: Delayed blood clearance and enhanced hepatotoxicity. Hepatology 1992, 15, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.; Leo, M.A.; Lieber, C.S. Interactions between alcohol and beta-carotene in patients with alcoholic liver disease. Am. J. Clin. Nutr. 1994, 60, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Krinsky, N.I. The antioxidant and biological properties of the carotenoids. Ann. N. Y. Acad. Sci. 1998, 854, 443–447. [Google Scholar] [CrossRef]
- Honya, M.; Kinoshita, T.; Ishikawa, M.; Mori, H.; Nisizawa, K. Seasonal variation in the lipid content of culturedLaminaria japonica: Fatty acids, sterols, β-carotene and tocopherol. J. Appl. Phycol. 1994, 6, 25–29. [Google Scholar] [CrossRef]
- Yoon, N.Y.; Lee, S.-H.; Yong, L.; Kim, S.-K. Phlorotannins from Ishige okamurae and their acetyl- and butyrylcholinesterase inhibitory effects. J. Funct. Foods 2009, 1, 331–335. [Google Scholar] [CrossRef]
- Rajapakse, N.; Kim, S.-K. Chapter 2—Nutritional and Digestive Health Benefits of Seaweed. In Advances in Food and Nutrition Research; Kim, S.-K., Ed.; Academic Press: Cambridge, MA, USA, 2011; Volume 64, pp. 17–28. [Google Scholar]
- Brownlee, I.A.; Allen, A.; Pearson, J.P.; Dettmar, P.W.; Havler, M.E.; Atherton, M.R.; Onsøyen, E. Alginate as a source of dietary fiber. Crit. Rev. Food Sci. Nutr. 2005, 45, 497–510. [Google Scholar] [CrossRef]
- El Khoury, D.; Goff, H.D.; Anderson, G.H. The role of alginates in regulation of food intake and glycemia: A gastroenterological perspective. Crit. Rev. Food Sci. Nutr. 2015, 55, 1406–1424. [Google Scholar] [CrossRef]
- Chater, P.I.; Wilcox, M.; Cherry, P.; Herford, A.; Mustar, S.; Wheater, H.; Brownlee, I.; Seal, C.; Pearson, J. Inhibitory activity of extracts of Hebridean brown seaweeds on lipase activity. J. Appl. Phycol. 2016, 28, 1303–1313. [Google Scholar] [CrossRef] [Green Version]
- Congress, S.H.R. 3562—Nutrition Labeling and Education Act of 1990. Available online: https://www.congress.gov/bill/101st-congress/house-bill/3562 (accessed on 11 August 2020).
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucoidan from Sargassum sp. and Fucus vesiculosus reduces cell viability of lung carcinoma and melanoma cells in vitro and activates natural killer cells in mice in vivo. Int. J. Biol. Macromol. 2011, 49, 331–336. [Google Scholar] [CrossRef]
- Kraan, S. Algal polysaccharides, novel applications and outlook. In Carbohydrates-Comprehensive Studies on Glycobiology and Glycotechnology; IntechOpen: London, UK, 2012. [Google Scholar]
- Lahaye, M. Marine algae as sources of fibres: Determination of soluble and insoluble dietary fibre contents in some ‘sea vegetables’. J. Sci. Food Agric. 1991, 54, 587–594. [Google Scholar] [CrossRef]
- Rajapakse, N.; Kim, S.K. Nutritional and digestive health benefits of seaweed. Adv. Food Nutr. Res. 2011, 64, 17–28. [Google Scholar] [PubMed]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Fleury, N.; Lahaye, M. Chemical and physico-chemical characterisation of fibres from Laminaria digitata (kombu breton): A physiological approach. J. Sci. Food Agric. 1991, 55, 389–400. [Google Scholar] [CrossRef]
- Morton, I.D.; Morton, C. Elsevier’s Dictionary of Food Science and Technology; Elsevier Scientific Publishing Co.: Amsterdam, The Netherlands, 1977. [Google Scholar]
- Solomon, H.; William, W. Bioactive Food Components, Encyclopedia of Food & Culture. Accept. Food Politics B Lett. Charles Scribner’s Sons 2003, 1, 201. [Google Scholar]
- Hutkins, R.W.; Krumbeck, J.A.; Bindels, L.B.; Cani, P.D.; Fahey, G.; Goh, Y.J.; Hamaker, B.; Martens, E.C.; Mills, D.A.; Rastal, R.A.; et al. Prebiotics: Why definitions matter. Curr. Opin. Biotechnol. 2016, 37, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devillé, C.; Gharbi, M.; Dandrifosse, G.; Peulen, O. Study on the effects of laminarin, a polysaccharide from seaweed, on gut characteristics. J. Sci. Food Agric. 2007, 87, 1717–1725. [Google Scholar] [CrossRef]
- Spicer, S.E.; Adams, J.M.M.; Thomas, D.S.; Gallagher, J.A.; Winters, A.L. Novel rapid method for the characterisation of polymeric sugars from macroalgae. J. Appl. Phycol. 2017, 29, 1507–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araya, J. Riesgos y Beneficios del Consumo de Grasas y Aceites; En Programa De Educacion a Distancia Departamento de Nutrición; Facultad de Medicina Universidad de Chile: Santiago, Chile, 2008. [Google Scholar]
- Darcy-Vrillon, B. Nutritional aspects of the developing use of marine macroalgae for the human food industry. Int. J. Food Sci. Nutr. 1993, 44, S23–S35. [Google Scholar]
- Cowey, C. Use of synthetic diets and biochemical criteria in the assessment of nutrient requirements of fish. J. Fish. Board Can. 1976, 33, 1040–1045. [Google Scholar] [CrossRef]
- Krauss-Etschmann, S.; Shadid, R.; Campoy, C.; Hoster, E.; Demmelmair, H.; Jiménez, M.; Gil, A.; Rivero, M.; Veszprémi, B.; Decsi, T.; et al. Effects of fish-oil and folate supplementation of pregnant women on maternal and fetal plasma concentrations of docosahexaenoic acid and eicosapentaenoic acid: A European randomized multicenter trial. Am. J. Clin. Nutr. 2007, 85, 1392–1400. [Google Scholar] [PubMed] [Green Version]
- Smith, G.I.; Atherton, P.; Reeds, D.N.; Mohammed, B.S.; Rankin, D.; Rennie, M.J.; Mittendorfer, B. Dietary omega-3 fatty acid supplementation increases the rate of muscle protein synthesis in older adults: A randomized controlled trial. Am. J. Clin. Nutr. 2011, 93, 402–412. [Google Scholar] [CrossRef] [Green Version]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 fatty acids EPA and DHA: Health benefits throughout life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef]
- Freund-Levi, Y.; Eriksdotter-Jönhagen, M.; Cederholm, T.; Basun, H.; Faxen-Irving, G.; Garlind, A.; Vedin, I.; Vessby, B.; Wahlund, L.-O.; Palmblad, J. ω-3 fatty acid treatment in 174 patients with mild to moderate Alzheimer disease: OmegAD study: A randomized double-blind trial. Arch. Neurol. 2006, 63, 1402–1408. [Google Scholar] [CrossRef] [Green Version]
- Judge, M.P.; Harel, O.; Lammi-Keefe, C.J. Maternal consumption of a docosahexaenoic acid–containing functional food during pregnancy: Benefit for infant performance on problem-solving but not on recognition memory tasks at age 9 mo. Am. J. Clin. Nutr. 2007, 85, 1572–1577. [Google Scholar] [CrossRef] [Green Version]
- Conquer, J.A.; Tierney, M.C.; Zecevic, J.; Bettger, W.J.; Fisher, R.H. Fatty acid analysis of blood plasma of patients with Alzheimer's disease, other types of dementia, and cognitive impairment. Lipids 2000, 35, 1305–1312. [Google Scholar] [CrossRef]
- Neff, L.M.; Culiner, J.; Cunningham-Rundles, S.; Seidman, C.; Meehan, D.; Maturi, J.; Wittkowski, K.M.; Levine, B.; Breslow, J.L. Algal docosahexaenoic acid affects plasma lipoprotein particle size distribution in overweight and obese adults. J. Nutr. 2011, 141, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Michel, G.; Tonon, T.; Scornet, D.; Cock, J.M.; Kloareg, B. The cell wall polysaccharide metabolism of the brown alga Ectocarpus siliculosus. Insights into the evolution of extracellular matrix polysaccharides in Eukaryotes. New Phytol. 2010, 188, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Summers, G.; Wong, R. Nutrient and heavy metal content of edible seaweeds in New Zealand. N. Z. J. Crop Hortic. Sci. 2010, 38, 19–28. [Google Scholar] [CrossRef]
- Kloareg, B.; Quatrano, R. Structure of the cell walls of marine algae and ecophysiological functions of the matrix polysaccharides. Oceanogr. Marie Biol. Annu. Rev. 1988, 26, 259–315. [Google Scholar]
- Cronshaw, J.; Myers, A.; Preston, R.D. A chemical and physical investigation of the cell walls of some marine algae. Biochim. Biophys. Acta 1958, 27, 89–103. [Google Scholar] [CrossRef]
- Michel, G.; Tonon, T.; Scornet, D.; Cock, J.M.; Kloareg, B. Central and storage carbon metabolism of the brown alga Ectocarpus siliculosus: Insights into the origin and evolution of storage carbohydrates in Eukaryotes. New Phytol. 2010, 188, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Fleurence, J. Seaweed proteins: Biochemical, nutritional aspects and potential uses. Trends Food Sci. Technol. 1999, 10, 25–28. [Google Scholar] [CrossRef]
- Fleurence, J. The enzymatic degradation of algal cell walls: A useful approach for improving protein accessibility? J. Appl. Phycol. 1999, 11, 313–314. [Google Scholar] [CrossRef]
- Association of Official Agricultural Chemists. Official Methods of Analysis of Association of Official Analytical Chemists International, 16th ed.; AOCA: Gaithersburg, MD, USA, 1995; Volumes I–II, p. 870. [Google Scholar]
- Alaiz, M.; Navarro, J.L.; Girón, J.; Vioque, E. Amino acid analysis by high-performance liquid chromatography after derivatization with diethyl ethoxymethylenemalonate. J. Chromatogr. 1992, 591, 181–186. [Google Scholar] [CrossRef]
- Leyton, A.; Lienqueo, M.E.; Shene, C. Macrocystis pyrifera: Substrate for the production of bioactive compounds. J. Appl. Phycol. 2019. [Google Scholar] [CrossRef]
- Heffernan, N.; Smyth, T.J.; FitzGerald, R.J.; Soler-Vila, A.; Brunton, N. Antioxidant activity and phenolic content of pressurised liquid and solid–liquid extracts from four Irish origin macroalgae. Int. J. Food Sci. Technol. 2014, 49, 1765–1772. [Google Scholar] [CrossRef]
- Zhang, H.; Row, K.H. Extraction and Separation of Polysaccharides from Laminaria japonica by Size-Exclusion Chromatography. J. Chromatogr. Sci. 2014, 53, 498–502. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar] [CrossRef]
- Kadam, S.U.; Álvarez, C.; Tiwari, B.K.; O’Donnell, C.P. Extraction and characterization of protein from Irish brown seaweed Ascophyllum nodosum. Food Res. Int. 2017, 99, 1021–1027. [Google Scholar] [CrossRef]
- Cheung, R.C.; Ng, T.B.; Wong, J.H. Marine Peptides: Bioactivities and Applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef] [PubMed]
- Li-Chan, E.C.Y. Bioactive peptides and protein hydrolysates: Research trends and challenges for application as nutraceuticals and functional food ingredients. Curr. Opin. Food Sci. 2015, 1, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Harnedy, P.A.; FitzGerald, R.J. Bioactive Proteins, Peptides, and Amino Acids from Macroalgae. J. Phycol. 2011, 47, 218–232. [Google Scholar] [CrossRef]
- Bondu, S.; Bonnet, C.; Gaubert, J.; Deslandes, É.; Turgeon, S.L.; Beaulieu, L. Bioassay-guided fractionation approach for determination of protein precursors of proteolytic bioactive metabolites from macroalgae. J. Appl. Phycol. 2015, 27, 2059–2074. [Google Scholar] [CrossRef]
- Vásquez, V.; Martínez, R.; Bernal, C. Enzyme-assisted extraction of proteins from the seaweeds Macrocystis pyrifera and Chondracanthus chamissoi: Characterization of the extracts and their bioactive potential. J. Appl. Phycol. 2019, 31, 1999–2010. [Google Scholar] [CrossRef]
- Leyton, A.; Pezoa-Conte, R.; Mäki-Arvela, P.; Mikkola, J.P.; Lienqueo, M.E. Improvement in carbohydrate and phlorotannin extraction from Macrocystis pyrifera using carbohydrate active enzyme from marine Alternaria sp. as pretreatment. J. Appl. Phycol. 2017, 29, 2039–2048. [Google Scholar] [CrossRef]
- Leyton, A.; Vergara-Salinas, J.R.; Pérez-Correa, J.R.; Lienqueo, M.E. Purification of phlorotannins from Macrocystis pyrifera using macroporous resins. Food Chem. 2017, 237, 312–319. [Google Scholar] [CrossRef]
- Sundberg, A.; Pranovich, A.; Holmbom, B. Chemical characterization of various types of mechanical pulp fines. J. Pulp Pap. Sci. 2003, 29, 173–178. [Google Scholar]
- Ford, L.; Stratakos, A.C.; Theodoridou, K.; Dick, J.T.A.; Sheldrake, G.N.; Linton, M.; Corcionivoschi, N.; Walsh, P.J. Polyphenols from Brown Seaweeds as a Potential Antimicrobial Agent in Animal Feeds. Acs Omega 2020, 5, 9093–9103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larripa, I.B.; de Pargament, M.M.; de Vinuesa, M.L.; Mayer, A.M.S. Biological activity in Macrocystis pyrifera from Argentina: Sodium alginate, fucoidan and laminaran. II. Genotoxicity. Hydrobiologia 1987, 151, 491–496. [Google Scholar] [CrossRef]
- Misra, N.N.; Rai, D.K.; Hossain, M. Chapter 10—Analytical techniques for bioactives from seaweed. In Seaweed Sustainability; Tiwari, B.K., Troy, D.J., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 271–287. [Google Scholar] [CrossRef]
- Galland-Irmouli, A.V.; Fleurence, J.; Lamghari, R.; Luçon, M.; Rouxel, C.; Barbaroux, O.; Bronowicki, J.P.; Villaume, C.; Guéant, J.L. Nutritional value of proteins from edible seaweed Palmaria palmata (dulse). J. Nutr. Biochem. 1999, 10, 353–359. [Google Scholar] [CrossRef]
- Kadam, S.U.; Álvarez, C.; Tiwari, B.K.; O’Donnell, C.P. Chapter 9—Extraction of biomolecules from seaweeds. In Seaweed Sustainability; Tiwari, B.K., Troy, D.J., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 243–269. [Google Scholar] [CrossRef]
- Tierney, M.S.; Smyth, T.J.; Rai, D.K.; Soler-Vila, A.; Croft, A.K.; Brunton, N. Enrichment of polyphenol contents and antioxidant activities of Irish brown macroalgae using food-friendly techniques based on polarity and molecular size. Food Chem. 2013, 139, 753–761. [Google Scholar] [CrossRef]
- Wong, K.; Cheung, P.C. Influence of drying treatment on three Sargassum species. J. Appl. Phycol. 2001, 13, 43–50. [Google Scholar] [CrossRef]
- Maehre, H.K.; Edvinsen, G.K.; Eilertsen, K.-E.; Elvevoll, E.O. Heat treatment increases the protein bioaccessibility in the red seaweed dulse (Palmaria palmata), but not in the brown seaweed winged kelp (Alaria esculenta). J. Appl. Phycol. 2016, 28, 581–590. [Google Scholar] [CrossRef] [Green Version]
- Harrysson, H.; Hayes, M.; Eimer, F.; Carlsson, N.-G.; Toth, G.B.; Undeland, I. Production of protein extracts from Swedish red, green, and brown seaweeds, Porphyra umbilicalis Kützing, Ulva lactuca Linnaeus, and Saccharina latissima (Linnaeus) J. V. Lamouroux using three different methods. J. Appl. Phycol. 2018, 30, 3565–3580. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, J.; Meaney, S.; Williams, G.A.; Hayes, M. Extraction of Protein from Four Different Seaweeds Using Three Different Physical Pre-Treatment Strategies. Molecules 2020, 25, 2005. [Google Scholar]
- Suslick, K.S.; Crum, L.A. Sonochemistry and Sonoluminescence. In Encyclopedia of Acoustics; Crocker, M.J., Ed.; John Wiley and Sons: Hoboken, NJ, USA, 1997; pp. 271–282. [Google Scholar] [CrossRef]
- Chemat, F.; Khan, M.K. Applications of ultrasound in food technology: Processing, preservation and extraction. Ultrason. Sonochemistry 2011, 18, 813–835. [Google Scholar] [CrossRef]
- Vanthoor-Koopmans, M.; Wijffels, R.H.; Barbosa, M.J.; Eppink, M.H. Biorefinery of microalgae for food and fuel. Bioresour. Technol. 2013, 135, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Polikovsky, M.; Fernand, F.; Sack, M.; Frey, W.; Müller, G.; Golberg, A. Towards marine biorefineries: Selective proteins extractions from marine macroalgae Ulva with pulsed electric fields. Innov. Food Sci. Emerg. Technol. 2016, 37, 194–200. [Google Scholar] [CrossRef]
- Goettel, M.; Eing, C.; Gusbeth, C.; Straessner, R.; Frey, W. Pulsed electric field assisted extraction of intracellular valuables from microalgae. Algal Res. 2013, 2, 401–408. [Google Scholar] [CrossRef]
- Coustets, M.; Al-Karablieh, N.; Thomsen, C.; Teissié, J. Flow process for electroextraction of total proteins from microalgae. J. Membr. Biol. 2013, 246, 751–760. [Google Scholar] [CrossRef]
- Lee, J.Y.; Yoo, C.; Jun, S.Y.; Ahn, C.Y.; Oh, H.M. Comparison of several methods for effective lipid extraction from microalgae. Bioresour. Technol. 2010, 101 (Suppl. 1), S75–S77. [Google Scholar] [CrossRef]
- Barba, F.J.; Grimi, N.; Vorobiev, E. New Approaches for the Use of Non-conventional Cell Disruption Technologies to Extract Potential Food Additives and Nutraceuticals from Microalgae. Food Eng. Rev. 2015, 7, 45–62. [Google Scholar] [CrossRef]
- Kumar, P.; Sharma, N.; Ranjan, R.; Kumar, S.; Bhat, Z.F.; Jeong, D.K. Perspective of membrane technology in dairy industry: A review. Asian-Australas. J. Anim. Sci. 2013, 26, 1347–1358. [Google Scholar] [CrossRef] [Green Version]
- Ye, H.; Wang, K.; Zhou, C.; Liu, J.; Zeng, X. Purification, antitumor and antioxidant activities in vitro of polysaccharides from the brown seaweed Sargassum pallidum. Food Chem. 2008, 111, 428–432. [Google Scholar] [CrossRef]
- Rioux, L.E.; Turgeon, S.L.; Beaulieu, M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr. Polym. 2007, 69, 530–537. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; O’Doherty, J.V.; Tiwari, B.K.; Sweeney, T.; Rajauria, G. Enhancing the Extraction of Polysaccharides and Antioxidants from Macroalgae Using Sequential Hydrothermal-Assisted Extraction Followed by Ultrasound and Thermal Technologies. Mar. Drugs 2019, 17, 457. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Vaquero, M.; Rajauria, G.; O'Doherty, J.V.; Sweeney, T. Polysaccharides from macroalgae: Recent advances, innovative technologies and challenges in extraction and purification. Food Res. Int. 2017, 99, 1011–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charoensiddhi, S.; Franco, C.; Su, P.; Zhang, W. Improved antioxidant activities of brown seaweed Ecklonia radiata extracts prepared by microwave-assisted enzymatic extraction. J. Appl. Phycol. 2015, 27, 2049–2058. [Google Scholar] [CrossRef]
- Xiao, X.; Si, X.; Yuan, Z.; Xu, X.; Li, G. Isolation of fucoxanthin from edible brown algae by microwave-assisted extraction coupled with high-speed countercurrent chromatography. J. Sep. Sci. 2012, 35, 2313–2317. [Google Scholar] [CrossRef] [PubMed]
- Praveen, M.A.; Parvathy, K.R.K.; Balasubramanian, P.; Jayabalan, R. An overview of extraction and purification techniques of seaweed dietary fibers for immunomodulation on gut microbiota. Trends Food Sci. Technol. 2019, 92, 46–64. [Google Scholar] [CrossRef]
- Lafarga, T.; Acién-Fernández, F.G.; Garcia-Vaquero, M. Bioactive peptides and carbohydrates from seaweed for food applications: Natural occurrence, isolation, purification, and identification. Algal Res. 2020, 48, 101909. [Google Scholar] [CrossRef]
- Taleuzzaman, M.; Ali, S.; Gilani, S.; Imam, S.; Hafeez, A. Ultra performance liquid chromatography (UPLC)-a review. Austin J. Anal. Pharm. Chem. 2015, 2, 1056. [Google Scholar]
- Adrien, A.; Dufour, D.; Baudouin, S.; Maugard, T.; Bridiau, N. Evaluation of the anticoagulant potential of polysaccharide-rich fractions extracted from macroalgae. Nat. Prod. Res. 2017, 31, 2126–2136. [Google Scholar] [CrossRef]
- Biancarosa, I.; Espe, M.; Bruckner, C.; Heesch, S.; Liland, N.; Waagbø, R.; Torstensen, B.; Lock, E. Amino acid composition, protein content, and nitrogen-to-protein conversion factors of 21 seaweed species from Norwegian waters. J. Appl. Phycol. 2017, 29, 1001–1009. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Anastyuk, S.D.; Ishina, I.A.; Isakov, V.V.; Zvyagintseva, T.N.; Thinh, P.D.; Zadorozhny, P.A.; Dmitrenok, P.S.; Ermakova, S.P. Structural characteristics and anticancer activity in vitro of fucoidan from brown alga Padina boryana. Carbohydr. Polym. 2018, 184, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, H. Exploring the Structure of Oligo-and Polysaccharides: Synthesis and NMR Spectroscopy Studies. Ph.D. Thesis, Department of Organic Chemistry, Stockholm University, Stockholm, Sweden, 2010. [Google Scholar]
- Zollmann, M.; Robin, A.; Prabhu, M.; Polikovsky, M.; Gillis, A.; Greiserman, S.; Golberg, A. Green technology in green macroalgal biorefineries. Phycologia 2019, 58, 516–534. [Google Scholar] [CrossRef]
- Alvarado-Morales, M.; Gunnarsson, I.B.; Fotidis, I.A.; Vasilakou, E.; Lyberatos, G.; Angelidaki, I. Laminaria digitata as a potential carbon source for succinic acid and bioenergy production in a biorefinery perspective. Algal Res. 2015, 9, 126–132. [Google Scholar] [CrossRef]
- Gajaria, T.K.; Suthar, P.; Baghel, R.S.; Balar, N.B.; Sharnagat, P.; Mantri, V.A.; Reddy, C.R.K. Integration of protein extraction with a stream of byproducts from marine macroalgae: A model forms the basis for marine bioeconomy. Bioresour. Technol. 2017, 243, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, L.J.; Zhu, F.X.; Zhu, J.Y.; Chen, X.D.; Zou, L.; Saito, M. In vitro and in vivo studies on the antioxidant activities of the aqueous extracts of Douchi (a traditional Chinese salt-fermented soybean food). Food Chem. 2008, 107, 1421–1428. [Google Scholar] [CrossRef]
- Trivedi, N.; Baghel, R.S.; Bothwell, J.; Gupta, V.; Reddy, C.R.K.; Lali, A.M.; Jha, B. An integrated process for the extraction of fuel and chemicals from marine macroalgal biomass. Sci. Rep. 2016, 6, 30728. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Gao, L.; Zhang, D.; Ye, N.; Chen, S.; Li, D. Enhanced hydrolysis of Macrocystis pyrifera by integrated hydroxyl radicals and hot water pretreatment. Bioresour. Technol. 2015, 179, 490–496. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration. Substances Generally Regarded as Safe (Final Rule). U.S. Food and Drug Administration (HHS). Available online: https://www.fda.gov/media/99659/download (accessed on 11 August 2020).
- Agence Nationale de Sécurité Sanitaire. Opinion of the French Agency for Food Environmental and Occupational Health & Safety on “Maximum Cadmium Levels for Seaweed Intended for Human Consumption”, ANSES Opinion Request No 2017-SA-0070 ed.; French Agency for Food: Maisons-Alfort, France, 2017; p. 57. [Google Scholar]
- Taylor, V.F.; Jackson, B.P. Concentrations and speciation of arsenic in New England seaweed species harvested for food and agriculture. Chemosphere 2016, 163, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Hansen, H.R.; Raab, A.; Francesconi, K.A.; Feldmann, J. Metabolism of Arsenic by Sheep Chronically Exposed to Arsenosugars as a Normal Part of Their Diet. 1. Quantitative Intake, Uptake, and Excretion. Environ. Sci. Technol. 2003, 37, 845–851. [Google Scholar] [CrossRef]
- European Commission. Commission Implementing Regulation (EU) 2017/2470 of 20 December 2017 Establishing the Union List of Novel Foods in Accordance with REGULATION (EU) 2015/2283 of the European Parliament and of the Council on Novel Foods; Text with EEA relevance; European Commission: Brussels, Belgium, 2017; Volume 2017/2470. [Google Scholar]
- Office of Nutrition and Food Labeling. Science Review of Isolated and Synthetic Non-Digestible Carbohydrates. US Department of Health and Human Services; US Food and Drug Administration: College Park, MD, USA, 2016. [Google Scholar]
- EFSA Panel on Food Additives and Nutrient Sources added to Food; Mortensen, A.; Aguilar, F.; Crebelli, R.; Di Domenico, A.; Frutos, M.J.; Dusemund, B. Re-evaluation of agar (E 406) as a food additive. Efsa J. 2016, 14, e04645. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Food Additives and Nutrient Sources added to Food; Younes, M.; Aggett, P.; Aguilar, F.; Crebelli, R.; Filipič, M.; Dusemund, B. Re-evaluation of alginic acid and its sodium, potassium, ammonium and calcium salts (E 400–E 404) as food additives. Efsa J. 2017, 15, e05049. [Google Scholar] [CrossRef] [Green Version]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104 (Suppl. 2), S1–S63. [Google Scholar] [CrossRef] [Green Version]
- Medical Economics Company. PDR for Herbal Medicines, 2nd ed.; Medical Economics Company.: Montvale, NJ, USA, 2000. [Google Scholar]
- Müssig, K. Iodine-induced toxic effects due to seaweed consumption. Compr. Handb. Iodine 2009, 897–908. [Google Scholar]
- Aderibigbe, B.A.; Buyana, B. Alginate in Wound Dressings. Pharmaceutics 2018, 10, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important Determinants for Fucoidan Bioactivity: A Critical Review of Structure-Function Relations and Extraction Methods for Fucose-Containing Sulfated Polysaccharides from Brown Seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [Green Version]
- Wargacki, A.J.; Leonard, E.; Win, M.N.; Regitsky, D.D.; Santos, C.N.S.; Kim, P.B.; Cooper, S.R.; Raisner, R.M.; Herman, A.; Sivitz, A.B.; et al. An engineered microbial platform for direct biofuel production from brown macroalgae. Science 2012, 335, 308–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ly, H.V.; Kim, S.-S.; Choi, J.H.; Woo, H.C.; Kim, J. Fast pyrolysis of Saccharina japonica alga in a fixed-bed reactor for bio-oil production. Energy Convers. Manag. 2016, 122, 526–534. [Google Scholar] [CrossRef]
- Camus, C.; Ballerino, P.; Delgado, R.; Olivera-Nappa, Á.; Leyton, C.; Buschmann, A.H. Scaling up bioethanol production from the farmed brown macroalga Macrocystis pyrifera in Chile. Biofuels Bioprod. Biorefining 2016, 10, 673–685. [Google Scholar] [CrossRef]
- Fitton, J.; Irhimeh, M. Macroalgae in nutricosmetics. Cosmet. Toilet. 2008, 123, 93. [Google Scholar]
- Bommers, M. La-Mer. My Skin—And What It Needs. Available online: https://www.la-mer.com/en/ (accessed on 10 July 2020).
- Making Cosmetics. Sea Kelp Extract, USDA Certified Organic. 2020. Available online: https://www.makingcosmetics.com/Sea-Kelp-Extract-USDA-Certified-Organic_p_22.html?locale=en (accessed on 13 August 2020).
- The Good Scents Company. Macrocystis Pyrifera Juice. Available online: http://www.thegoodscentscompany.com/data/fl1750621.html (accessed on 13 August 2020).
- Barrett, A. Forbes Goes Seaweed Bioplastics. Available online: https://bioplasticsnews.com/2019/04/06/forbes-goes-seaweed-bioplastics/ (accessed on 19 August 2020).
- Chopin, T.; Robinson, S.M.C.; Troell, M.; Neori, A.; Buschmann, A.H.; Fang, J. Multitrophic Integration for Sustainable Marine Aquaculture. In Encyclopedia of Ecology; Jørgensen, S.E., Fath, B.D., Eds.; Academic Press: Oxford, UK, 2008; pp. 2463–2475. [Google Scholar] [CrossRef]
- Abreu, M.H.; Varela, D.A.; Henríquez, L.; Villarroel, A.; Yarish, C.; Sousa-Pinto, I.; Buschmann, A.H. Traditional vs. Integrated Multi-Trophic Aquaculture of Gracilaria chilensis C.J. Bird, J. McLachlan & E.C. Oliveira: Productivity and physiological performance. Aquaculture 2009, 293, 211–220. [Google Scholar]
- Dhargalkar, V.K.; Verlecar, X.N. Southern Ocean seaweeds: A resource for exploration in food and drugs. Aquaculture 2009, 287, 229–242. [Google Scholar] [CrossRef]
- Fernández, P.A.; Gaitán-Espitia, J.D.; Leal, P.P.; Schmid, M.; Revill, A.T.; Hurd, C.L. Nitrogen sufficiency enhances thermal tolerance in habitat-forming kelp: Implications for acclimation under thermal stress. Sci. Rep. 2020, 10, 3186. [Google Scholar] [CrossRef] [Green Version]
- Hafting, J.T.; Critchley, A.T.; Cornish, M.L.; Hubley, S.A.; Archibald, A.F. On-land cultivation of functional seaweed products for human usage. J. Appl. Phycol. 2012, 24, 385–392. [Google Scholar] [CrossRef]
- Alemañ, A.E.; Robledo, D.; Hayashi, L. Development of seaweed cultivation in Latin America: Current trends and future prospects. Phycologia 2019, 58, 462–471. [Google Scholar] [CrossRef] [Green Version]
- Roleda, M.Y.; Hurd, C.L. Seaweed nutrient physiology: Application of concepts to aquaculture and bioremediation. Phycologia 2019, 58, 552–562. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, I.C.; Duarte, P.M.; Marinho, G.S.; Neumann, F.; Sousa-Pinto, I. Growth of Saccharina latissima (Laminariales, Phaeophyceae) cultivated offshore under exposed conditions. Phycologia 2019, 58, 504–515. [Google Scholar] [CrossRef]
- Pilar, G.-J.; Olegario, B.-R.; Rafael, R.R. Occurrence of jasmonates during cystocarp development in the red alga Grateloupia imbricata. J. Phycol. 2016, 52, 1085–1093. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, A.Q.; Neish, I.C.; Critchley, A.T. Phyconomy: The extensive cultivation of seaweeds, their sustainability and economic value, with particular reference to important lessons to be learned and transferred from the practice of eucheumatoid farming. Phycologia 2019, 58, 472–483. [Google Scholar] [CrossRef]
- Hwang, E.K.; Yotsukura, N.; Pang, S.J.; Su, L.; Shan, T.F. Seaweed breeding programs and progress in eastern Asian countries. Phycologia 2019, 58, 484–495. [Google Scholar] [CrossRef]
- Lee, H.J.; Choi, J.I. Enhancing temperature tolerance of Pyropia tenera (Bangiales) by inducing mutation. Phycologia 2019, 58, 496–503. [Google Scholar] [CrossRef] [Green Version]


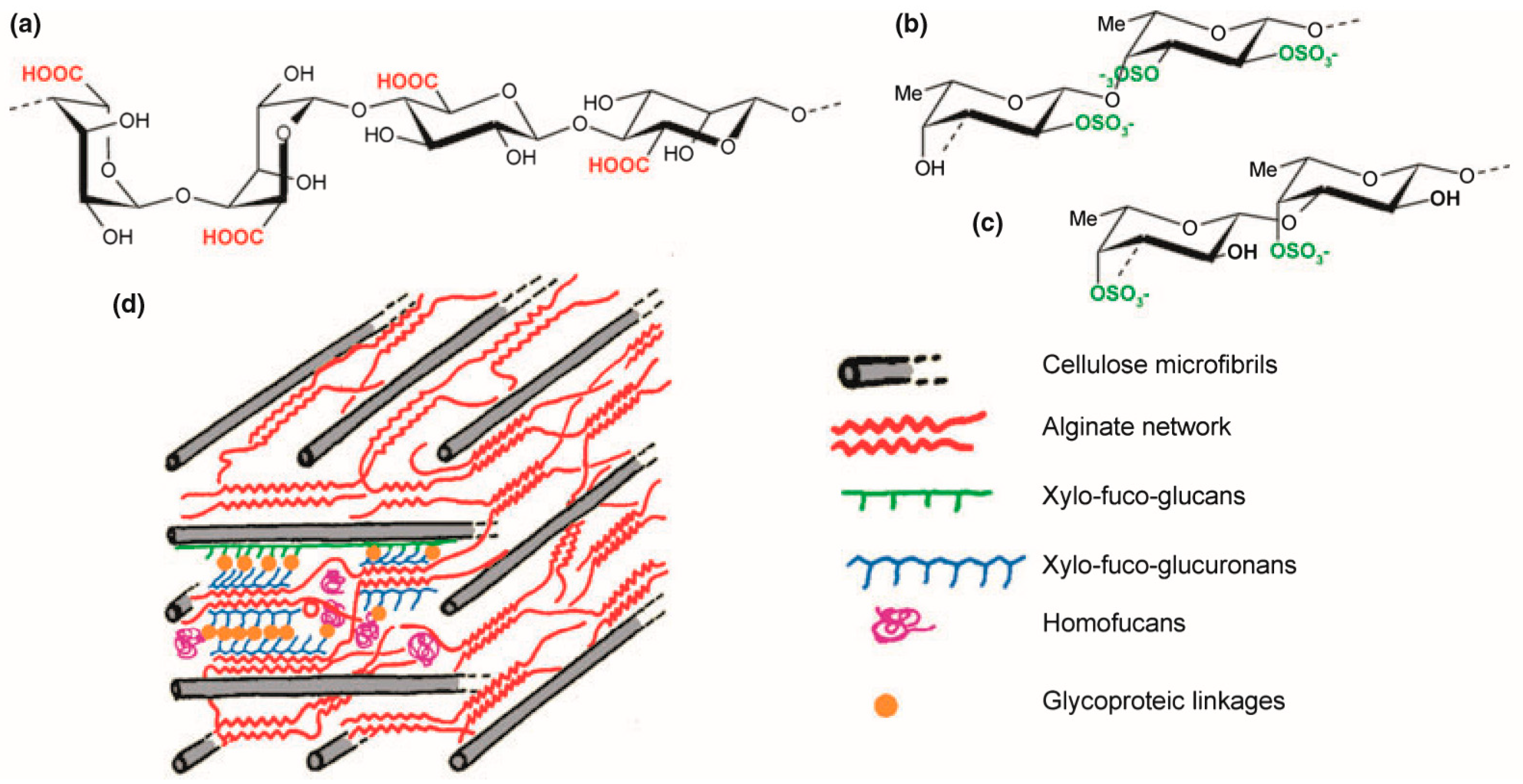{kind=link}
| Brown Seaweed Species | Feed or Food | Food Group | Product Produced/Product Quality | Function of Product | Reference |
|---|---|---|---|---|---|
| M. pyrifera | Feed | Fresh seaweed | Fresh seaweed/ 9–13% Crude Protein | Food supply for abalone | [14,19,20] |
| L. digitata | Feed | Fresh seaweed | Fresh seaweed | Food supply for abalone and sea urchins | [14] |
| L. digitata and L. hyperborea | Feed | Fresh seaweed | 1.4 ± 0.2 kg WW per day/79.6% Digestibility | North Ronaldsay sheep breed complete daily food supply; dry matter degradation (DMD, 71.7%, at 48 h) | [68] |
| M. pyrifera | Feed | Dried seaweed, included in meal | 30% DW of Diet/77% Digestibility | 30%DW of goat daily diet | [21] |
| L. digitata | Feed | Dried seaweed supplement to daily diet | Powder: 0.001 kg/day | Rabbits showed a significant effect of lowering total cholesterol, lipoprotein, especially triglyceride | [69] |
| Saccharina japonica | Food | Dried seaweed supplement | Powder (1, 2, 3, 4%) | Ash content increased; 1% seaweed added to breakfast sausages were the most improved for physiochemical and sensory properties | [64] |
| M. pyrifera | Food | Dried seaweed 3–28% Food recipe ingredient | Huiro fritters/6.9%DW of protein | Protein supplement | |
| M. pyrifera | Food | Dried seaweed 3–28% Food recipe ingredient | Breadsticks/9.5%DW of protein | Protein supplement | |
| M. pyrifera | Feed | Macrocystis meal food supplement | Macrocystis Meal concentration of (33.3 g/kg) based on shrimp weight, was fed (1.6 g over 28 days)/Protein efficiency ratio of 1.7 purified | Dietary supplement for juvenile white shrimp (Litopenaeus vannamei) | [67] |
| M. pyrifera | Feed | Complementary meal | Meal dietary supplement with 8.5% crude protein/85% Digestibility, | Dietary supplement for male bovine zebu bulls | [22] |
| M. pyrifera | Feed | Food supplement in the form of derived flour | 3% DW of daily diet/low digestibility for salmonids fish | 3% DW of daily diet as dietary supplement for minerals and carbohydrates needed by salmonids fish species | [23] |
| M. pyrifera | Food | Carbohydrates | Phycocolloids/ medium or high viscosity alginate | Thickening agent for food products | [25,61] |
| M. pyrifera and Laminaria spp. | Food | Carbohydrates | Phycocolloids/ Alginic acid powder | Ingredients for dietary biscuits, to induce feeling of fullness | [25] |
| Laminaria spp. | Food | Carbohydrates | Phycocolloids/ Alginate: soft to medium strength gel | Thickening agent for food products | [25,61] |
| Macrocystis sp. | Food | Carbohydrates | Phycocolloids/ Alginate | Emulsifying, gelling, stabiliser yoghurts, ice creams | [62] |
| M. pyrifera and Laminaria spp. | Feed | Carbohydrates | Phycocolloids/ crude alginate | Binding agent in salmon and other fish feeds | [25] |
| Laminaria spp. | Feed | Carbohydrates. Prebiotics: Seaweed extract as a dietary supplement | Polysaccharides: (Laminarin and Fucoidan) 1–4 g/kg/day | Supplement to daily diet weanling pigs for 21 days: alleviated the need to use of high-lactose diets for (>60 g/kg) for weanling pigs, and alleviated common problems occurring post-weaning | [70] |
| Laminaria spp. | Feed | Carbohydrates. Food Supplement | Polysaccharides: Laminarin (0.001 kg), Fucoidan (0.0008 kg), and ash (0.0082 kg) = Total supplement weight 0.01 kg/day | Enhanced piglet immune function and colonic microflora at weaning | [73] |
| L. digitata | Feed | Carbohydrates. Seaweed extract: dietary supplement: spray-dried (SD) and wet forms (WS) | Polysaccharides: Laminarin: 0.5 g/kg feed; Fucoidan: 0.42 g/kg Supplement to basal diet (Complete daily basal diet was SD = 1.9 kg/day; WS = 1.8 kg/day) | Reduction in lipid oxidation in the muscle tissue in 75% of pigs consequently improved the quality of pork steaks | [76] |
| L. digitata | Food | Carbohydrates. Seaweed extract: dietary supplement: spray-dried (SD) and wet forms (WS) | Laminarin (9.3%) and fucoidan (7.8%), added to mince pork patties | Reduced the appearance of surface redness of fresh patties, significantly decreased lipid oxidation in cooked patties | [77] |
| L. digitata | Feed | Carbohydrates. Dietary supplement seaweed extract | Polysaccharides: Laminarin and fucoidan: of 1.5 g/kg addition to the basal diet | Reduced the enterobacteria, bifidobacteria, and lactobacilli populations in the caecum and colon, while only marginal effects on the immune response was observed in weaned pigs | [74] |
| L. digitata | Feed | Carbohydrates. Dietary supplement | Structural polysaccharides: Purified β-glucans of 0.25 g/kg addition to the basal diet | Reduced the Enterobacteriaceae population and pro-inflammatory markers in the colon in pigs | [75] |
| Seaweed Types and Species | Compound of Interest | Product Produced | Function of Product | Reference |
|---|---|---|---|---|
| M. pyrifera | Phycocolloids/ medium or high viscosity alginate | Sodium and calcium alginate | Wound dressings | [25,81] |
| M. pyrifera | Phycocolloids/ Alginate | Alginic acid powder | Aid in relieving acid indigestion; treatment of gastroesophageal reflux (GERD) disease | [25,61] |
| M. pyrifera | Phycocolloids/ Alginate | Calcium alginate bead | Controlled release of medicinal drugs and other chemicals | [25,82] |
| Laminaria spp. | Phycocolloids/ Alginate: soft to medium strength gel | Sodium and calcium alginate fibres | Wound dressings | [25,81] |
| Laminaria spp. | Phycocolloids/ Alginate | Alginic acid powder | Aid in relieving acid indigestion; treatment of gastroesophageal reflux (GERD) disease | [25,61] |
| Laminaria spp. | Phycocolloids/ Alginate | Calcium alginate bead | Controlled release of medicinal drugs and other chemicals | [25,82] |
| L. digitata | Phycocolloids/ Alginate | Sodium alginate | Soft and elastic gels; potential drug delivery via microparticles | [83] |
| M. pyrifera | Sulphated polysaccharides | Fucoidan | Immune modulator, causing delays in apoptosis and promoting pro-inflammatory cytokine production | [89] |
| Saccharina japonica | Sulphated polysaccharides | Fucoidan | Significant reduction in thrombus lysis time | [88] |
| Brown seaweeds (including L. digitata, Saccharina japonica; M. pyrifera) | Sulphated polysaccharides | Fucoidan | Anti-inflammatory, antioxidant, antibacterial, and immunological activity; lipid inhibition; obesity prevention or treatment | [84,85,86,87] |
| Saccharina japonica | Secondary metabolite | Phlorotannin | Anti-proliferation of human tumour cells | [112] |
| L. hyperborea | Secondary metabolite | Phlorotannin | Wound sealing and reconstruction during wound healing | [113] |
| M. pyrifera | Secondary metabolite | Phlorotannin: phloroeckol, tetrameric phloroglucinol | Antidiabetic; antioxidant activity; prevention of skin aging | [114] |
| L. digitata | crude polysaccharide-rich seaweed extract | Crude extract and depolymerised extract | Improved gut microbiota composition; increase in short-chain fatty acids | [98] |
| Fucus vesiculosus, M. pyrifera; Saccharina japonica, | Seaweed extract: Maritech ®® extract | Fucus vesiculosus 85%, w/w; M. pyrifera 10%, w/w; Saccharina japonica, 5%, w/w; zinc vitamin B6 and manganese) | Dose-dependent decrease in osteoarthritis in 5 females and 7 males | [115,116] |
| M. pyrifera | Seaweed extract | High purity Fucoidan 75–90% purity; Kelp Oil and/or Kelp Concentrate: Krill oil | Total antioxidant protection | [99] |
| L. digitata | Seaweed Extract: L. digitata thallus prepared in glycerine and water. | Actipone®® | A moisturising agent and stimulant, skin protecting, soothing, and smoothing properties. | [100] |
| L. digitata | Seaweed Extract: 1 DH | Boiron | Homeopathic medicine | [101] |
| Seaweed Types and Species | Compound of interest | Product Produced | Function of Product | Reference |
|---|---|---|---|---|
| M. pyrifera, | Mannitol and alginate | Methane | Fuel | [25] |
| Saccharina latissima | Laminarian and mannitol, alginate | Methane: Natural Gas | Fuel | [118] |
| L. hyperborea | Laminarian and mannitol | Ethanol | Fuel | [117] |
| Macrocystis sp. and Laminaria sp. | Dried seaweed | Dried seaweed | Removal of copper, zinc, and cadmium ions from solution | [119]; [120] |
| L. digitata | Alginate | Beads covered in calcium alginate | Removal of heavy metals, cadmium, and copper from single and binary solutions | [121] |
| Macrocystis sp. and Laminaria sp. | Alginates | Alginates | Cosmetic uses: as gelling colloids, emulsion stabilisers, immunostimulating agents, moisturising, protective phycocolloids | [82,122] |
| Laminaria sp. | Laminarians | Laminarians | Cosmetic uses: antioxidant, anticellulite, and anti-inflammatory agents | [104] |
| M. pyrifera | Seaweed extract | High purity Fucoidan 75–90% purity | Antioxidant and health and wellness benefits (for potential use in cosmetics and nutraceuticals) | [99] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Purcell-Meyerink, D.; Packer, M.A.; Wheeler, T.T.; Hayes, M. Aquaculture Production of the Brown Seaweeds Laminaria digitata and Macrocystis pyrifera: Applications in Food and Pharmaceuticals. Molecules 2021, 26, 1306. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051306
Purcell-Meyerink D, Packer MA, Wheeler TT, Hayes M. Aquaculture Production of the Brown Seaweeds Laminaria digitata and Macrocystis pyrifera: Applications in Food and Pharmaceuticals. Molecules. 2021; 26(5):1306. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051306
Chicago/Turabian StylePurcell-Meyerink, Diane, Michael A. Packer, Thomas T. Wheeler, and Maria Hayes. 2021. "Aquaculture Production of the Brown Seaweeds Laminaria digitata and Macrocystis pyrifera: Applications in Food and Pharmaceuticals" Molecules 26, no. 5: 1306. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules26051306