
Impact of the Hydrolysis and Methanolysis of Bidesmosidic Chenopodium quinoa Saponins on Their Hemolytic Activity

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
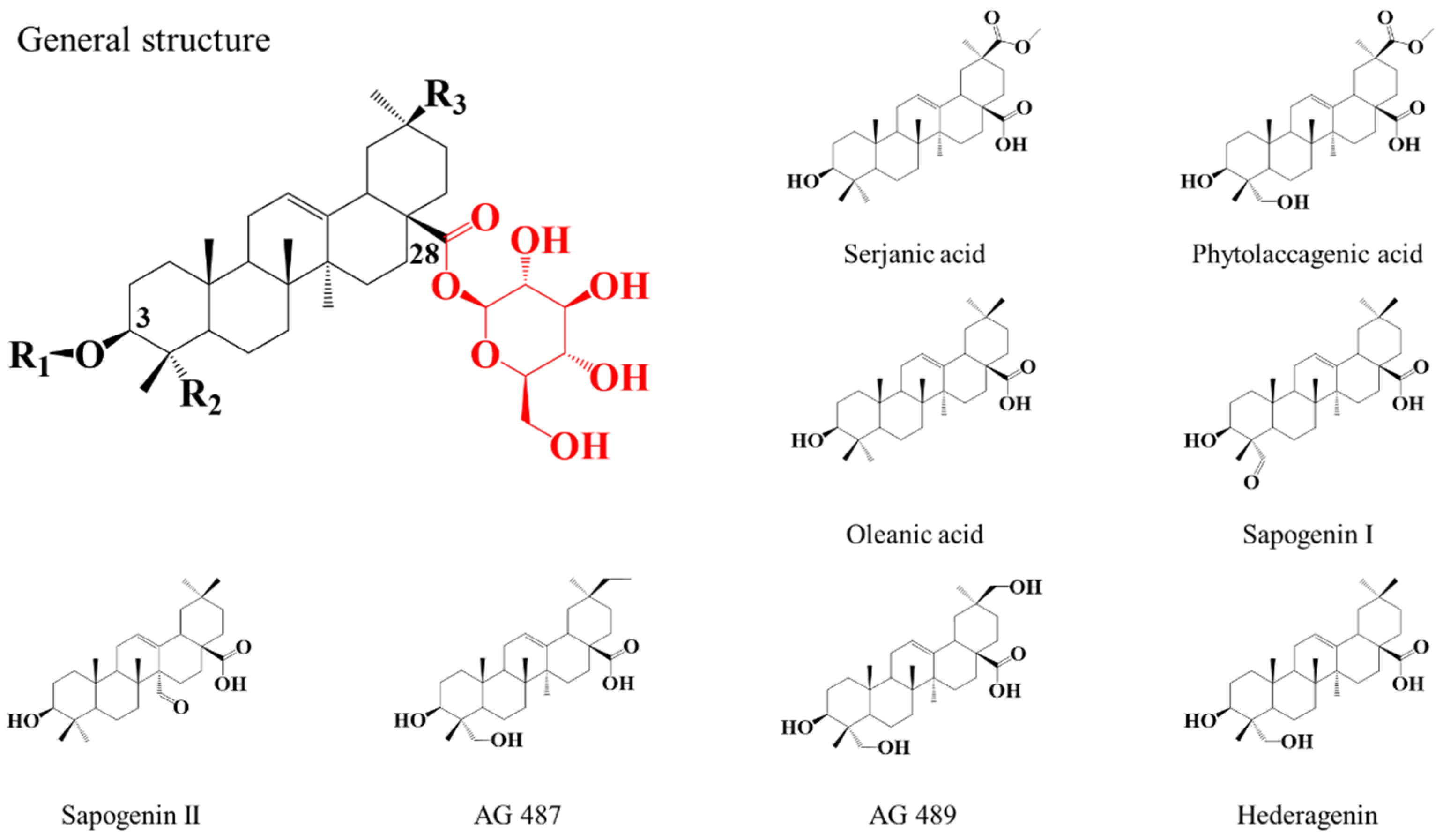
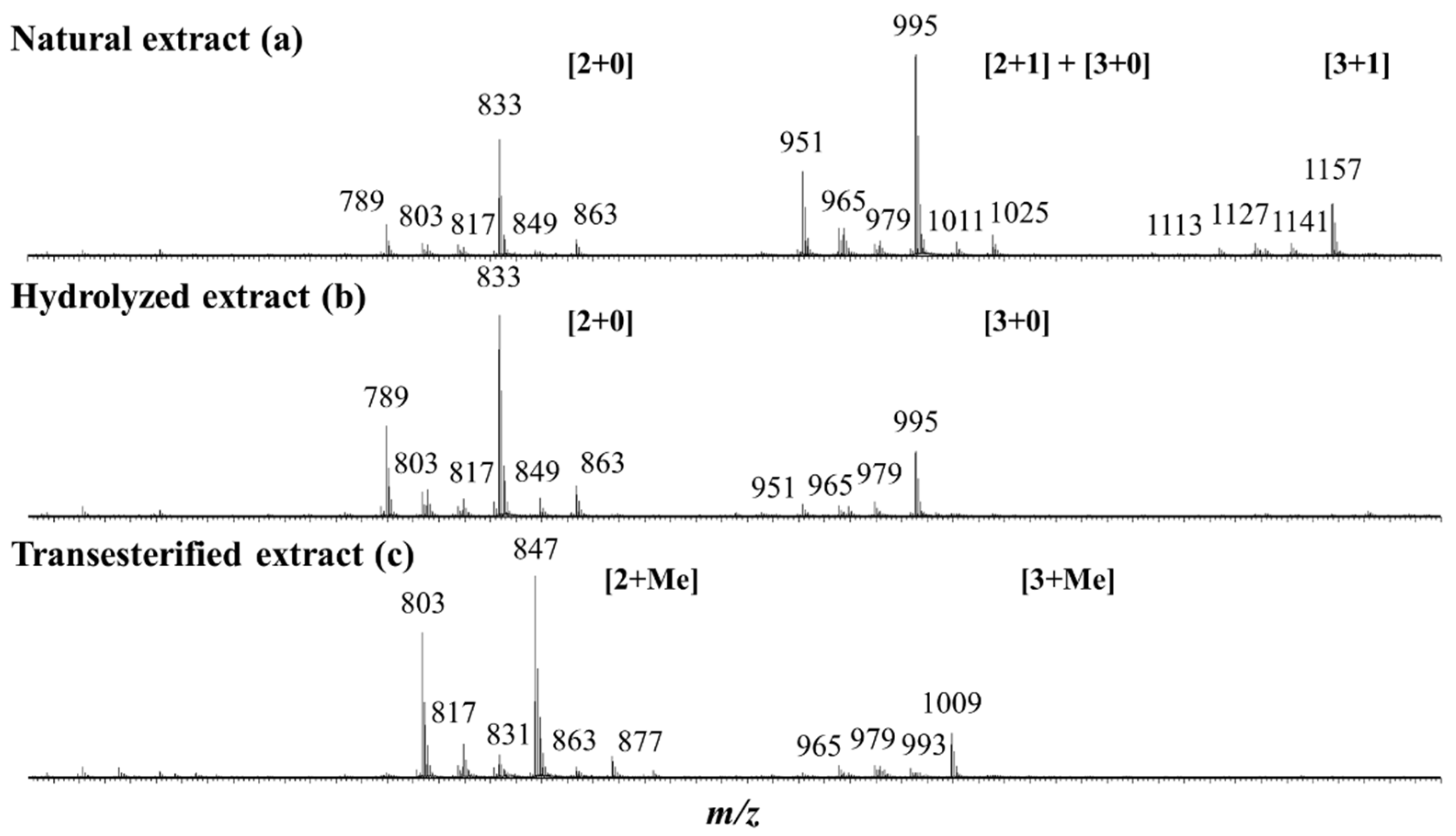
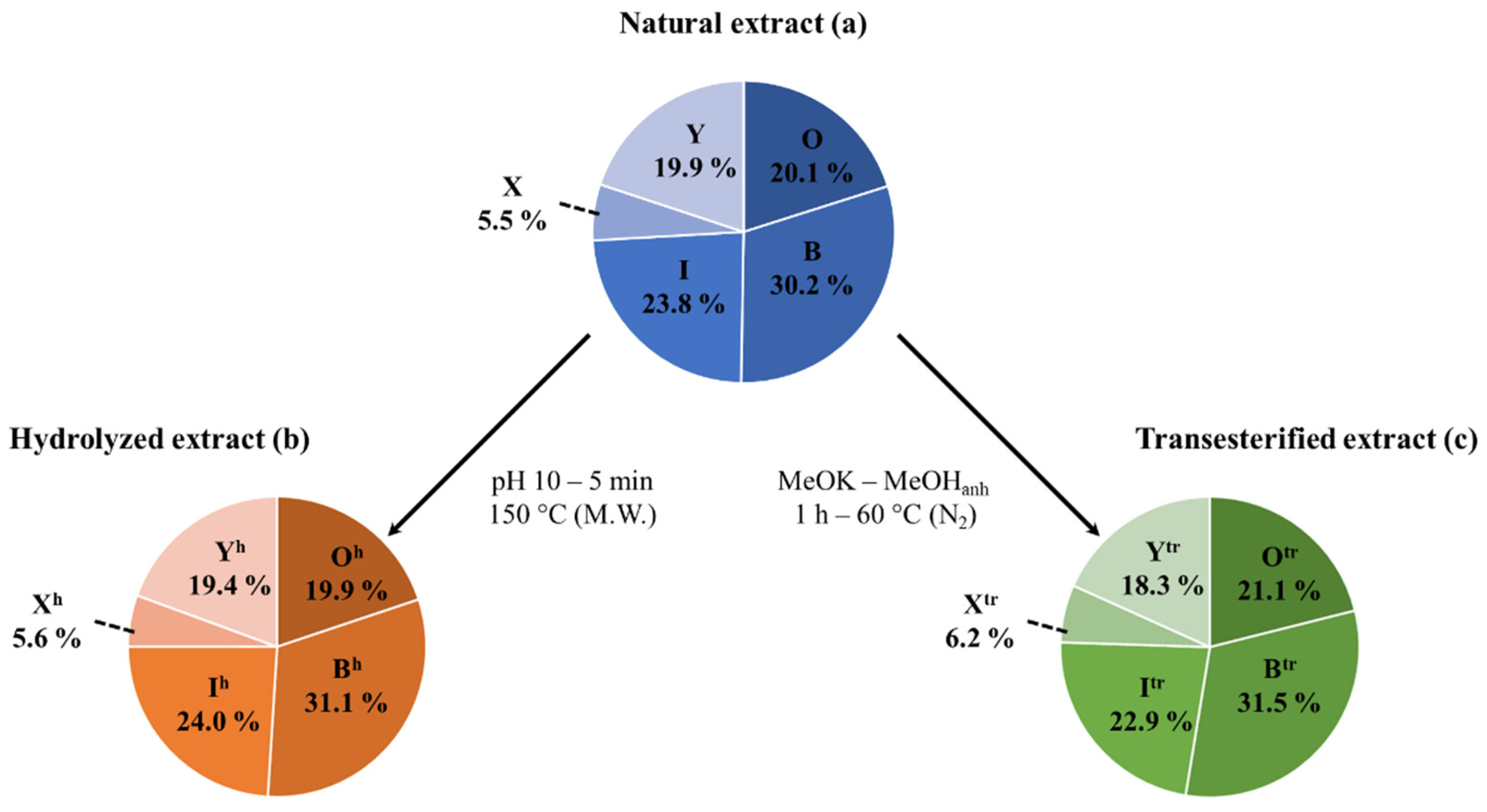
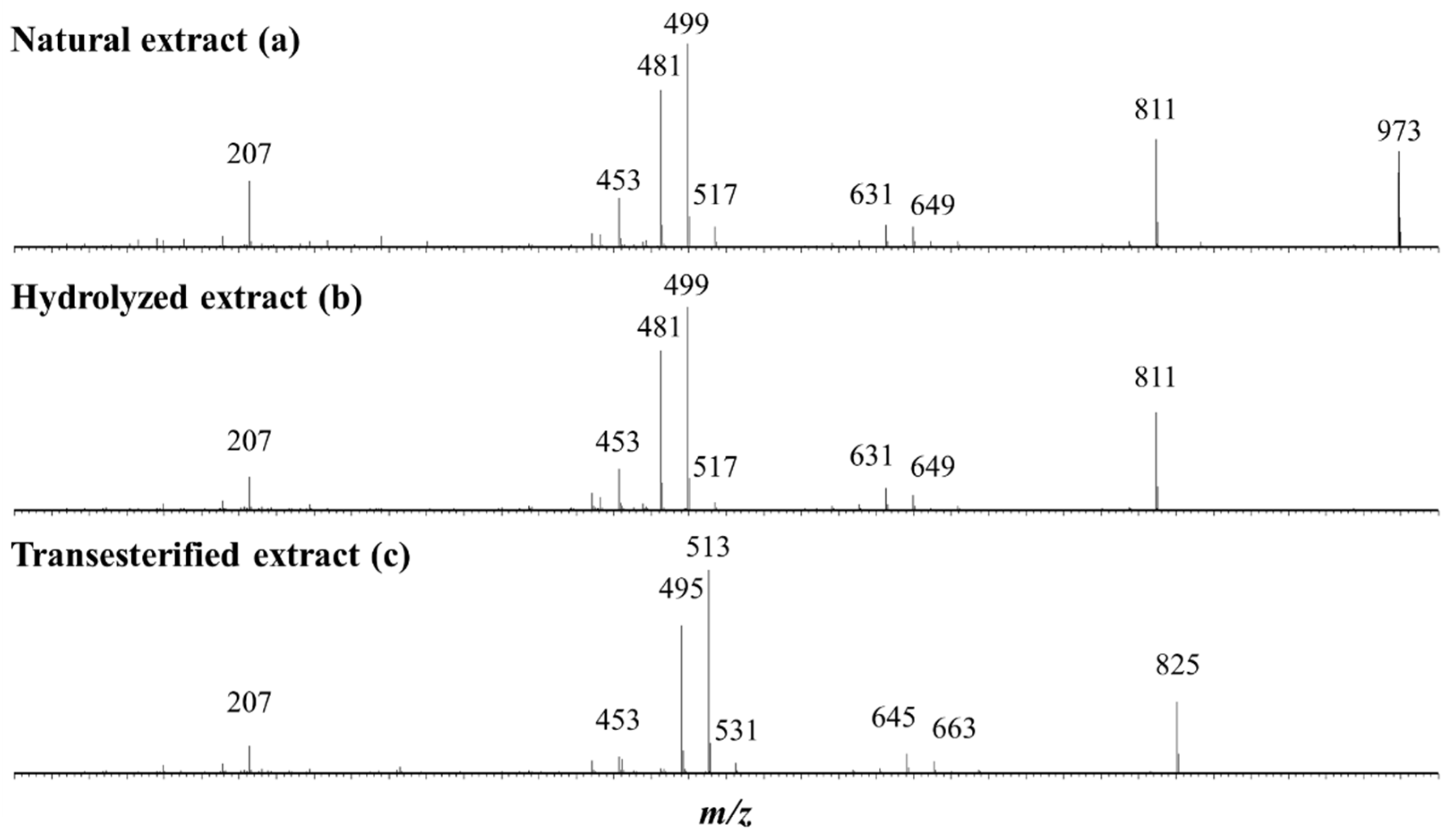
2.1. Saponin Identification and Quantification in the Natural Extract (NE)
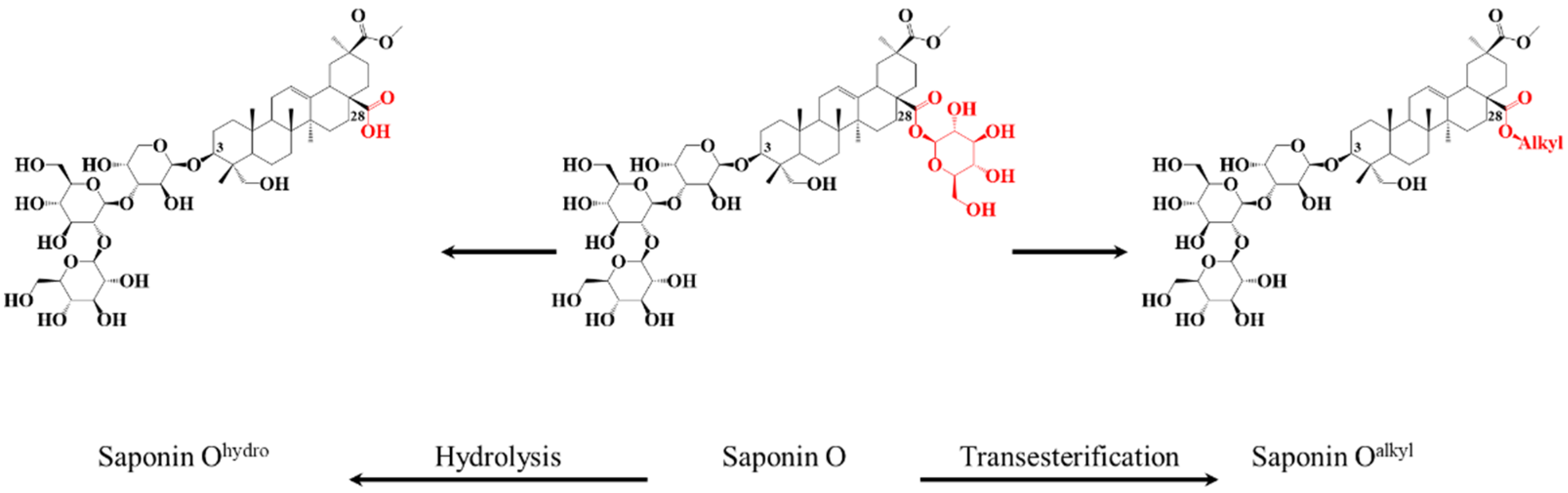
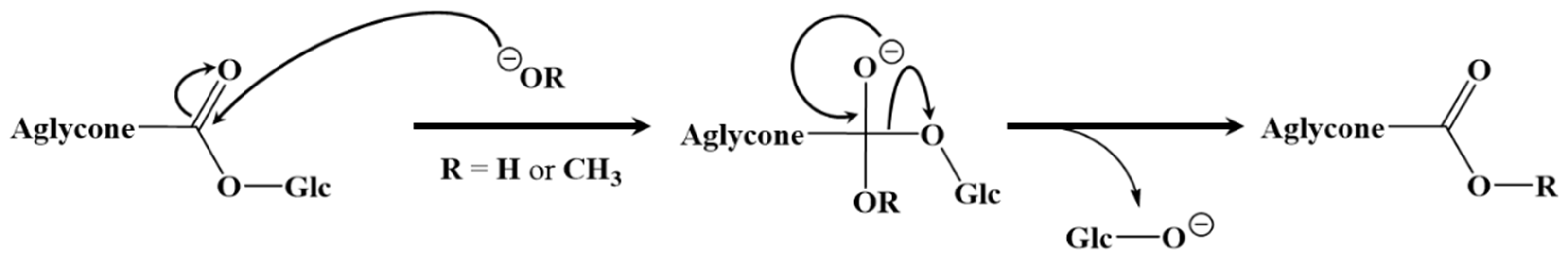
2.2. Selective Hydrolysis and Transesterification of the Quinoa Husk Bidesmosidic Saponins at C28
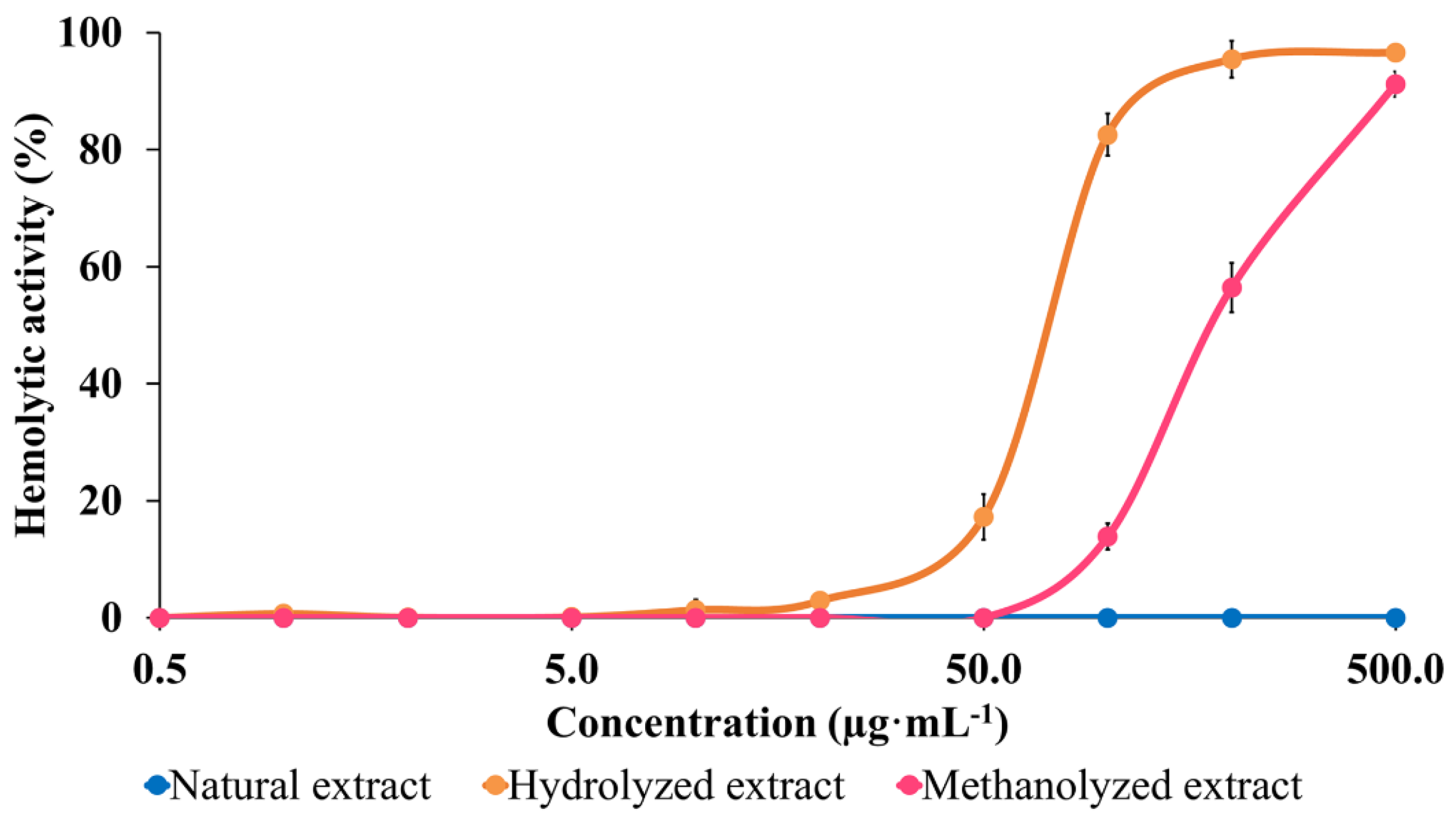
2.3. Hemolytic Activity (HA) Modulation
3. Materials and Methods
3.1. Chemicals
3.2. Extraction
3.3. Microwave-Assisted Alkaline Hydrolysis
3.4. Methanolysis
3.5. Mass Spectrometry (MS) Analyses
3.6. Hemolytic Activity Experiments
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Rohit Dutt, A.K.; Sharma, R.K.; Keservani, V.G. Promising Drug Molecules of Natural Origin; CRC Press: Coca Raton, FL, USA, 2020; ISBN 9781771888868. [Google Scholar]
- Hostettmann, K.; Marston, A. Chemistry Pharmacology of Natural Products-Saponins; Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Seca, A.M.L.; Pinto, D.C.G.A. Biological Potential and Medical Use of Secondary Metabolites. Medicines 2019, 6, 66. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Cao, Y.; Jørgensen, L.V.G.; Strobel, B.W.; Hansen, H.C.B.; Cedergreen, N. Where does the toxicity come from in saponin extract? Chemosphere 2018, 204, 243–250. [Google Scholar] [CrossRef]
- Deacon, B.J.W.; Mitchell, R.T. Toxicity of oat roots, oat root extracts, and saponins to zoospores of Pythium spp. and other fungi. Trans. Br. Mycol. Soc. 1985, 84, 479–487. [Google Scholar] [CrossRef]
- Caulier, G.; Flammang, P.; Gerbaux, P.; Eeckhaut, I. When a repellent becomes an attractant: Harmful saponins are kairomones attracting the symbiotic Harlequin crab. Sci. Rep. 2013, 3, 2639. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, S.; Ichiba, T.; Reimer, J.D.; Tanaka, J. Chemoattraction of the pearlfish Encheliophis vermicularis to the sea cucumber Holothuria leucospilota. Chemoecology 2014, 24, 121–126. [Google Scholar] [CrossRef]
- Francis, G.; Kerem, Z.; Makkar, H.P.S.; Becker, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef]
- Desai, S.D.; Desai, D.G.; Kaur, H. Saponins and their biological activities. Pharma Times 2009, 41, 13–16. [Google Scholar]
- Voogt, P.A.; Huiskamp, R. Sex-dependence and seasonal variation of saponins in the gonads of the starfish Asterias rubens: Their relation to reproduction. Comp. Biochem. Physiol. Part A Physiol. 1979, 62, 1049–1055. [Google Scholar] [CrossRef]
- Kassem, A.S.; Ahmed, A.M.; Tariq, M.R. Study of saponins in methanol extract of the leaves of Acacia etbaica subspecies etbaica. Res. J. Pharm. Biol. Chem. Sci. 2014, 5, 803–810. [Google Scholar]
- Demeyer, M.; De Winter, J.; Caulier, G.; Eeckhaut, I.; Flammang, P.; Gerbaux, P. Molecular diversity and body distribution of saponins in the sea star Asterias rubens by mass spectrometry. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2014, 168, 1–11. [Google Scholar] [CrossRef]
- Bahrami, Y.; Zhang, W.; Franco, C.M.M. Distribution of Saponins in the Sea Cucumber Holothuria lessoni; the Body Wall Versus the Viscera, and Their Biological Activities. Mar. Drugs 2018, 16, 423. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Laval, S.; Yu, B. Chemical Synthesis of Saponins, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2014; Volume 71, ISBN 9780128001288. [Google Scholar]
- Maier, M.S. Biological Activities of Sulfated Glycosides from Echinoderms. In Studies in Natural Products Chemistry; Elsevier B.V.: Amsterdam, The Netherlands, 2008; Volume 35, pp. 311–354. ISBN 9780444531810. [Google Scholar]
- Tantry, M.A.; Khan, I.A. Saponins from Glycine max Merrill (soybean). Fitoterapia 2013, 87, 49–56. [Google Scholar] [CrossRef]
- Yoshikawa, M.; Harada, E.; Murakami, T.; Matsuda, H.; Wariishi, N.; Yamahara, J.; Murakami, N.; Kitagawa, I. Escins-Ia, Ib, IIa, IIb, and IIIa, Bioactive Triterpene Oligoglycosides From the Seeds of Aesculus hippocastanum L.: Their Inhibitory Effects on Ethanol Absorption and Hypoglycemic Activity on Glucose Tolerance Test. Chem. Pharm. Bull. Tokyo 1994, 42, 1357–1359. [Google Scholar]
- Jæger, D.; Ndi, C.P.; Crocoll, C.; Simpson, B.S.; Khakimov, B.; Guzman-Genuino, R.M.; Hayball, J.D.; Xing, X.; Bulone, V.; Weinstein, P.; et al. Isolation and Structural Characterization of Echinocystic Acid Triterpenoid Saponins from the Australian Medicinal and Food Plant Acacia ligulata. J. Nat. Prod. 2017, 80, 2692–2698. [Google Scholar] [CrossRef]
- Honey-Escandón, M.; Arreguín-Espinosa, R.; Solís-Marín, F.A.; Samyn, Y. Biological and taxonomic perspective of triterpenoid glycosides of sea cucumbers of the family Holothuriidae (Echinodermata, Holothuroidea). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2015, 180, 16–39. [Google Scholar] [CrossRef]
- Caulier, G.; Mezali, K.; Soualili, D.L.; Decroo, C.; Demeyer, M.; Eeckhaut, I.; Gerbaux, P.; Flammang, P. Chemical characterization of saponins contained in the body wall and the Cuvierian tubules of the sea cucumber Holothuria (Platyperona) sanctori (Delle Chiaje, 1823). Biochem. Syst. Ecol. 2016, 68, 119–127. [Google Scholar] [CrossRef]
- Mohammadizadeh, F.; Ehsanpor, M.; Afkhami, M.; Mokhlesi, A.; Khazaali, A.; Montazeri, S. Evaluation of antibacterial, antifungal and cytotoxic effects of Holothuria scabra from the North Coast of the Persian Gulf. J. Mycol. Med. 2013, 23, 225–229. [Google Scholar] [CrossRef]
- Kashani, H.H.; Hoseini, E.S.; Nikzad, H.; Aarabi, M.H. Pharmacological properties of medicinal herbs by focus on secondary metabolites. Life Sci. J. 2012, 9, 509–520. [Google Scholar]
- Lee, S.-J.; Sung, J.-H.; Lee, S.-J.; Moon, C.-K.; Lee, B.-H. Antitumor activity of a novel ginseng saponin metabolite in human pulmonary adenocarcinoma cells resistant to cisplatin. Cancer Lett. 1999, 144, 39–43. [Google Scholar] [CrossRef]
- Mert-Türk, F. Saponins versus plant fungal pathogens. J. Cell Mol. Biol. 2006, 5, 13–17. [Google Scholar]
- Bordbar, S.; Anwar, F.; Saari, N. High-Value Components and Bioactives from Sea Cucumbers for Functional Foods—A Review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef] [Green Version]
- Lorent, J.H.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.P. The amphiphilic nature of saponins and their effects on artificial and biological membranes and potential consequences for red blood and cancer cells. Org. Biomol. Chem. 2014, 12, 8803–8822. [Google Scholar] [CrossRef]
- Lorent, J.; Lins, L.; Domenech, Ò.; Quetin-Leclercq, J.; Brasseur, R.; Mingeot-Leclercq, M.-P. Domain Formation and Permeabilization Induced by the Saponin α-Hederin and Its Aglycone Hederagenin in a Cholesterol-Containing Bilayer. Langmuir 2014, 30, 4556–4569. [Google Scholar] [CrossRef]
- Lorent, J.; Le Duff, C.S.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.-P. Induction of Highly Curved Structures in Relation to Membrane Permeabilization and Budding by the Triterpenoid Saponins, α- and δ-Hederin. J. Biol. Chem. 2013, 288, 14000–14017. [Google Scholar] [CrossRef] [Green Version]
- Böttger, S.; Melzig, M.F. The influence of saponins on cell membrane cholesterol. Bioorg. Med. Chem. 2013, 21, 7118–7124. [Google Scholar] [CrossRef]
- Keukens, E.A.J.; De Vrije, T.; Van Den Boom, C.; De Waard, P.; Plasman, H.H.; Thiel, F.; Chupin, V.; Jongen, W.M.F.; De Kruijff, B. Molecular basis of glycoalkaloid induced membrane disruption. Biochim. Biophys. Acta 1995, 1240, 216–228. [Google Scholar] [CrossRef] [Green Version]
- Qu, C.; Yang, L.; Yu, S.; Wang, S.; Bai, Y.; Zhang, H. Investigation of the interactions between ginsenosides and amino acids by mass spectrometry and theoretical chemistry. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2009, 74, 478–483. [Google Scholar] [CrossRef]
- Zelepuga, E.A.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I. Structure-activity relationships of holothuroid’s triterpene glycosides and some in silico insights obtained by molecular dynamics study on the mechanisms of their membranolytic action. Mar. Drugs 2021, 19, 604. [Google Scholar] [CrossRef]
- Armah, C.N.; Mackie, A.R.; Roy, C.; Price, K.; Osbourn, A.E.; Bowyer, P.; Ladha, S. The Membrane-Permeabilizing Effect of Avenacin A-1 Involves the Reorganization of Bilayer Cholesterol. Biophys. J. 1999, 76, 281–290. [Google Scholar] [CrossRef]
- Claereboudt, E.J.S.; Eeckhaut, I.; Lins, L.; Deleu, M. How different sterols contribute to saponin tolerant plasma membranes in sea cucumbers. Sci. Rep. 2018, 8, 10845. [Google Scholar] [CrossRef]
- Kalinin, V.I.; Prokofieva, N.G.; Likhatskaya, G.N.; Schentsova, E.B.; Agafonova, I.G.; Avilov, S.A.; Drozdova, O.A. Hemolytic activities of triterpene glycosides from the holothurian order dendrochirotida: Some trends in the evolution of this group of toxins. Toxicon 1996, 34, 475–483. [Google Scholar] [CrossRef]
- Reim, V.; Rohn, S. Characterization of saponins in peas (Pisum sativum L.) by HPTLC coupled to mass spectrometry and a hemolysis assay. Food Res. Int. 2015, 76, 3–10. [Google Scholar] [CrossRef]
- Madl, T.; Sterk, H.; Mittelbach, M.; Rechberger, G.N. Tandem Mass Spectrometric Analysis of a Complex Triterpene Saponin Mixture of Chenopodium quinoa. J. Am. Soc. Mass Spectrom. 2006, 17, 795–806. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Yang, X.; Xue, P.; Zhang, Z.; Ren, G. Improved antibacterial effects of alkali-transformed saponin from quinoa husks against halitosis-related bacteria. BMC Complement. Altern. Med. 2019, 19, 46–55. [Google Scholar] [CrossRef]
- Kuljanabhagavad, T.; Thongphasuk, P.; Chamulitrat, W.; Wink, M. Triterpene saponins from Chenopodium quinoa Willd. Phytochemistry 2008, 69, 1919–1926. [Google Scholar] [CrossRef]
- Colson, E.; Savarino, P.; Claereboudt, E.J.S.; Cabrera-Barjas, G.; Deleu, M.; Lins, L.; Eeckhaut, I.; Flammang, P.; Gerbaux, P. Enhancing the Membranolytic Activity of Chenopodium quinoa Saponins by Fast Microwave Hydrolysis. Molecules 2020, 25, 1731–1753. [Google Scholar] [CrossRef]
- Savarino, P.; Colson, E.; Caulier, G.; Eeckhaut, I.; Flammang, P.; Gerbaux, P. Microwave-Assisted Desulfation of the Hemolytic Saponins Extracted from Holothuria scabra Viscera. Molecules 2022, 27, 537. [Google Scholar] [CrossRef]
- Feng, J.; Chen, Y.; Liu, X.; Liu, S. Efficient improvement of surface activity of tea saponin through Gemini-like modification by straightforward esterification. Food Chem. 2015, 171, 272–279. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, W.-X.; Yan, M.-C.; Yu, Y.; Ikejima, T.; Cheng, M.-S. Synthesis and Tumor Cytotoxicity of Novel Amide Derivatives of β-Hederin. Molecules 2010, 15, 7871–7883. [Google Scholar] [CrossRef] [Green Version]
- Ding, N.; Chen, Q.; Zhang, W.; Ren, S.; Guo, Y.; Li, Y. Structure–activity relationships of saponin derivatives: A series of entry inhibitors for highly pathogenic H5N1 influenza virus. Eur. J. Med. Chem. 2012, 53, 316–326. [Google Scholar] [CrossRef]
- Abugoch James, L.E. Chapter 1 Quinoa (Chenopodium quinoa Willd.). In Advances in Food and Nutrition Research; Elsevier Inc.: Amsterdam, The Netherlands, 2009; Volume 58, pp. 1–31. [Google Scholar]
- Savarino, P.; Demeyer, M.; Decroo, C.; Colson, E.; Gerbaux, P. Mass spectrometry analysis of saponins. Mass Spectrom. Rev. 2021, 1–30. [Google Scholar] [CrossRef]
- Van Dyck, S.; Gerbaux, P.; Flammang, P. Elucidation of molecular diversity and body distribution of saponins in the sea cucumber Holothuria forskali (Echinodermata) by mass spectrometry. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2009, 152, 124–134. [Google Scholar] [CrossRef]
- Turhanen, P.A.; Leppanen, J.; Vepsalaïnen, J.J. Green and efficient esterification method using dried Dowex H+/NAI approach. ACS Omega 2019, 4, 8974–8984. [Google Scholar] [CrossRef] [Green Version]
- Leggio, A.; Belsito, E.L.; De Luca, G.; Di Gioia, M.L.; Leotta, V.; Romio, E.; Siciliano, C.; Liguori, A. One-pot synthesis of amides from carboxylic acids activated using thionyl chloride. RSC Adv. 2016, 6, 34468–34475. [Google Scholar] [CrossRef]
- Chung, J.W.; Noh, E.J.; Zhao, H.L.; Sim, J.-S.; Ha, Y.W.; Shin, E.M.; Lee, E.B.; Cheong, C.S.; Kim, Y.S. Anti-inflammatory Activity of Prosapogenin Methyl Ester of Platycodin D via Nuclear Factor-kappaB Pathway Inhibition. Biol. Pharm. Bull. 2008, 31, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Vo, N.N.Q.; Fukushima, E.O.; Muranaka, T. Structure and hemolytic activity relationships of triterpenoid saponins and sapogenins. J. Nat. Med. 2017, 71, 50–58. [Google Scholar] [CrossRef]
- Voutquenne, L.; Lavaud, C.; Massiot, G.; Men-Olivier, L. Le Structure-Activity Relationships of Haemolytic Saponins. Pharm. Biol. 2002, 40, 253–262. [Google Scholar] [CrossRef]
- Takechi, M.; Yasuo, T. Structure-activity relationships of the saponin α-hederin. Phytochemistry 1990, 29, 451–452. [Google Scholar] [CrossRef]
- Lorenz, P.; Conrad, J.; Klaiber, I.; Bunse, M.; Pfeiffer, T.; Stintzing, F.C.; Kammerer, D.R. Monodesmosidic oleanene-type saponins from kidney vetch (Anthyllis vulneraria L.) with hemolytic activity. Phytochem. Lett. 2021, 46, 21–28. [Google Scholar] [CrossRef]
- Mackie, A.M.; Grant, P.T.; Lasker, R. Avoidance reactions of a mollusc Buccinum undatum to saponin-like surface-active substances in extracts of the starfish Asterias rubens and Marthasterias glacialis. Comp. Biochem. Physiol. 1968, 26, 415–428. [Google Scholar] [CrossRef]
- Domanski, D.; Zegrocka-Stendel, O.; Perzanowska, A.; Dutkiewicz, M.; Kowalewska, M.; Grabowska, I.; Maciejko, D.; Fogtman, A.; Dadlez, M.; Koziak, K. Molecular Mechanism for Cellular Response to β-Escin and Its Therapeutic Implications. PLoS ONE 2016, 11, e0164365. [Google Scholar] [CrossRef]
- Muramatsu, N.; Kawasaki, H.; Onoe, F.; Ohshima, H.; Kondo, T. Hemolysis by Amphoteric Surfactants. J. Jpn. Oil Chem. Soc. 1990, 39, 555–559. [Google Scholar] [CrossRef]
- Stuardo, M.; San Martin, R. Antifungal properties of quinoa (Chenopodium quinoa Willd) alkali treated saponins against Botrytis cinerea. Ind. Crop. Prod. 2008, 27, 296–302. [Google Scholar] [CrossRef]
- Van Dyck, S.; Gerbaux, P.; Flammang, P. Qualitative and Quantitative Saponin Contents in Five Sea Cucumbers from the Indian Ocean. Mar. Drugs 2010, 8, 173–189. [Google Scholar] [CrossRef]
- Moses, T.; Papadopoulou, K.K.; Osbourn, A. Metabolic and functional diversity of saponins, biosynthetic intermediates and semi-synthetic derivatives. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 439–462. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Saponins | Elemental Compositions (M) | m/z (Δ ppm) [M + Na]+ | Aglycone | 3-O Glycan (R1) | %-Weights in Extract (%) | Mass Fractions in Husk (mg·g−1) | Retention Times (min) | Molar Proportions (%) |
|---|---|---|---|---|---|---|---|---|
| O a | C54H86O25 | 1157.5356 (1.6) | PA | Glu-Glu-Ara- | 20.01 ± 0.11 | 3.101 | 5.34 | 20.11 ± 0.13 |
| G a | C54H86O24 | 1141.5407 (0.6) | SA | Glu-Glu-Ara- | 0.29 ± 0.03 | 0.045 | 6.31 | 0.34 ± 0.05 |
| 32 a | C53H84O24 | 1127.5250 (3.2) | PA | Glu-Ara-Ara- | 3.46 ± 0.02 | 0.536 | 4.89 | 3.62 ± 0.03 |
| 61 a | C53H86O23 | 1113.5458 (1.2) | Hed | Glu-Glu-Ara- | 1.98 ± 0.06 | 0.307 | 6.51 | 2.03 ± 0.07 |
| N a | C49H78O21 | 1025.4933 (3.2) | PA | Glu-Gal- | 3.89 ± 0.10 | 0.603 | 4.77 | 4.00 ± 0.13 |
| 4 a | C48H76O21 | 1011.4777 (1.1) | AG533 | Glu-Ara- | 0.07 ± 0.03 | 0.011 | 5.43 | 0.09 ± 0.03 |
| B a | C48H76O20 | 995.4828 (0.1) | PA | Glu-Ara- | 29.50 ± 0.09 | 4.572 | 5.39 | 30.16 ± 0.09 |
| Q a | C48H78O19 | 981.5035 (3.3) | Hed | Glu-Gal- | 1.87 ± 0.10 | 0.290 | 5.92 | 1.97 ± 0.11 |
| H a | C48H76O19 | 979.4878 (4.1) | SA | Glu-Ara- | 0.29 ± 0.09 | 0.045 | 6.31 | 0.33 ± 0.12 |
| 19 a | C47H76O19 | 967.4878 (1.6) | AG489 | Glu-Ara- | 5.11 ± 0.07 | 0.792 | 3.72 | 5.83 ± 0.10 |
| F a | C47H74O19 | 965.4722 (1.4) | Hed | Xyl-GlcA- | 7.01 ± 0.10 | 1.086 | 4.92 | 7.69 ± 0.05 |
| I a | C47H76O18 | 951.4929 (2.0) | Hed | Glu-Ara- | 22.43 ± 0.11 | 3.476 | 6.58 | 23.83 ± 0.14 |
| / b | C43H68O16 | 863.4405 (0.6) | / | / | / | / | / | / |
| / b | C42H66O16 | 849.4249 (1.9) | / | / | / | / | / | / |
| / b | C42H66O15 | 833.4299 (0.1) | / | / | / | / | / | / |
| / b | C42H68O14 | 819.4507 (2.1) | / | / | / | / | / | / |
| / b | C42H66O14 | 817.4350 (0.5) | / | / | / | / | / | / |
| / b | C41H66O14 | 805.4350 (5.0) | / | / | / | / | / | / |
| / b | C41H64O14 | 803.4194 (0.7) | / | / | / | / | / | / |
| / b | C41H66O13 | 789.4401 (0.1) | / | / | / | / | / | / |
| Saponins | Elemental Compositions (M) | m/z [M + Na]+ | Mass Errors (Δ ppm) | Retention Times (min) | Molar Proportions (%) |
|---|---|---|---|---|---|
| Oh | C48H76O20 | 995.4828 | 2.8 | 7.02 | 20.06 ± 0.08 |
| Gh | C48H76O19 | 979.4878 | 0.8 | 8.71 | 0.23 ± 0.01 |
| 32h | C47H74O19 | 965.4746 | 2.3 | 6.51 | 3.35 ± 0.03 |
| 61h | C47H76O18 | 951.4929 | 3.0 | 6.71 | 1.98 ± 0.03 |
| Nh | C43H68O16 | 863.4405 | 0.6 | 6.78 | 4.43 ± 0.04 |
| 4h | C42H66O16 | 849.4249 | 1.9 | 7.42 | 0.05 ± 0.01 |
| Bh | C42H66O15 | 833.4299 | 0.1 | 7.23 | 30.31 ± 0.02 |
| Qh | C42H68O14 | 819.4507 | 2.1 | 8.76 | 1.95 ± 0.03 |
| Hh | C42H66O14 | 817.4350 | 0.5 | 8.81 | 0.35 ± 0.03 |
| 19h | C41H66O14 | 805.4350 | 5.0 | 5.28 | 5.75 ± 0.04 |
| Fh | C41H64O14 | 803.4194 | 0.7 | 6.67 | 7.95 ± 0.03 |
| Ih | C41H66O13 | 789.4401 | 0.1 | 10.14 | 23.59 ± 0.10 |
| Saponin | Elemental Composition (M) | m/z [M + Na]+ | Mass Error (Δ ppm) | Retention Time (min) | Composition Molar Proportion (%) |
|---|---|---|---|---|---|
| Otr | C49H78O20 | 1009.4984 | 1.6 | 9.47 | 20.09 ± 0.06 |
| Gtr | C49H78O19 | 993.5035 | 3.5 | 11.45 | 0.21 ± 0.01 |
| 32tr | C48H76O19 | 979.4878 | 0.2 | 8.74 | 3.34 ± 0.05 |
| 61tr | C48H78O18 | 965.5086 | 1.5 | 11.81 | 1.91 ± 0.03 |
| Ntr | C44H70O16 | 877.4198 | 0.2 | 9.89 | 4.46 ± 0.11 |
| 4tr | C43H68O16 | 863.4405 | 2.2 | 9.33 | 0.06 ± 0.01 |
| Btr | C43H68O15 | 847.4456 | 1.7 | 10.72 | 30.26 ± 0.11 |
| Qtr | C43H70O14 | 833.4663 | 3.1 | 7.00 | 1.88 ± 0.03 |
| Htr | C43H68O14 | 831.4507 | 1.7 | 7.86 | 0.32 ± 0.04 |
| 19tr | C42H68O14 | 819.4507 | 4.8 | 7.34 | 5.70 ± 0.06 |
| Ftr | C42H66O14 | 817.4350 | 5.0 | 9.57 | 7.94 ± 0.03 |
| Itr | C42H68O13 | 803.4558 | 1.0 | 11.99 | 23.83 ± 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Savarino, P.; Contino, C.; Colson, E.; Cabrera-Barjas, G.; De Winter, J.; Gerbaux, P. Impact of the Hydrolysis and Methanolysis of Bidesmosidic Chenopodium quinoa Saponins on Their Hemolytic Activity. Molecules 2022, 27, 3211. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103211
Savarino P, Contino C, Colson E, Cabrera-Barjas G, De Winter J, Gerbaux P. Impact of the Hydrolysis and Methanolysis of Bidesmosidic Chenopodium quinoa Saponins on Their Hemolytic Activity. Molecules. 2022; 27(10):3211. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103211
Chicago/Turabian StyleSavarino, Philippe, Carolina Contino, Emmanuel Colson, Gustavo Cabrera-Barjas, Julien De Winter, and Pascal Gerbaux. 2022. "Impact of the Hydrolysis and Methanolysis of Bidesmosidic Chenopodium quinoa Saponins on Their Hemolytic Activity" Molecules 27, no. 10: 3211. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103211