
Unveiling the Hidden Therapeutic Potential of Carnosine, a Molecule with a Multimodal Mechanism of Action: A Position Paper


1
Department of Drug and Health Sciences, University of Catania, 95125 Catania, Italy
2
Unit of Neuropharmacology and Translational Neurosciences, Oasi Research Institute—IRCCS, 94018 Troina, Italy
Molecules 2022, 27(10), 3303; https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103303
Submission received: 28 April 2022
/
Accepted: 19 May 2022
/
Published: 20 May 2022
(This article belongs to the Special Issue The Therapeutic Potential of Naturally Occurring Compounds in Counteracting Oxidative Stress and Inflammation: From Basic Science to Therapy)
{kind=link}
{kind=link}
Abstract
:Carnosine (β-alanyl-L-histidine) is a naturally occurring endogenous dipeptide and an over-the-counter food supplement with a well-demonstrated multimodal mechanism of action that includes the detoxification of reactive oxygen and nitrogen species, the down-regulation of the production of pro-inflammatory mediators, the inhibition of aberrant protein formation, and the modulation of cells in the peripheral (macrophages) and brain (microglia) immune systems. Since its discovery more than 100 years ago, a plethora of in vivo preclinical studies have been carried out; however, there is still substantial heterogeneity regarding the route of administration, the dosage, the duration of the treatment, and the animal model selected, underlining the urgent need for “coordinated/aligned” preclinical studies laying the foundations for well-defined future clinical trials. The main aim of the present position paper is to critically and concisely consider these key points and open a discussion on the possible “alignment” for future studies, with the goal of validating the full therapeutic potential of this intriguing molecule.
1. Carnosine: History and Biological Activities
Carnosine (β-alanyl-L-histidine) was discovered by Gulewitsch and Amiradžibi (Laboratorium der Universität Charkow, Charkow, Ukraine) more than 100 years ago during a study in which they were analyzing a meat extract [1]. They isolated and characterized several unidentified (at that time) nitrogen-containing compounds, one of which was carnosine. Since this molecule was isolated from minced meat, it was given the name “carnosine”, from the Latin caro, carnis (meat).
Carnosine is a naturally occurring endogenous dipeptide synthesized by the enzyme carnosine synthase 1 (CARNS1) from its constituent amino acids, β-alanine (synthesized in the liver) and L-histidine (external source) [2,3]. The levels of this dipeptide are very high (millimolar order of magnitude) in cardiac and skeletal muscles (∼99% of the carnosine in the body) [4,5] as well as in the brain [6]. Carnosine levels in human tissues and biological fluids are regulated by the activity of two carnosinases, enzymes that are able to decompose it into β-alanine and L-histidine: serum-circulating carnosine dipeptidase 1 (CNDP1) [7] and cytosolic carnosine dipeptidase 2 (CNDP2) [8], which belong to the M20 metalloprotease family [9]. A variety of other histidine-containing dipeptides such as anserine and balenine, methylated analogues of carnosine, can be found in the tissues of several animal species [10].
The biological activities and potential applications of carnosine are widespread and noteworthy. Although the muscles and brain contain the highest levels of carnosine, this dipeptide also performs biological activities in other areas of the body. Over the last 70 years, more than a thousand papers have been published describing the structure of carnosine and its biological activities in different systems; in particular, numerous studies have investigated the physiological role played by carnosine in muscles, probably due to its predominant localization in these areas, and the benefits of its exogenous supplementation for athletic performance enhancement. In this context, the supplementation of carnosine or β-alanine has been shown to improve the strength of muscle contraction and the mechanical work produced (“Severin’s phenomenon”) [11]; reduce the accumulation of lactate in active muscles, thus preventing intramuscular acidification [12]; increase the contraction and relaxation rates of muscles [13]; activate contractile proteins and stabilize muscles’ energy metabolism [14]; and improve physical performance and executive function following endurance exercise [15,16,17,18].
As previously mentioned, the biological effects and potential benefits of carnosine are not limited to muscle tissue. In fact, this dipeptide has shown the ability to act as a neurotransmitter [19], an enhancer of cell energy metabolism [20,21] and the immune system [22], a modulator of the metabolism of nitric oxide (NO) and related species [23,24,25], an anti-glycation and anti-aging agent [26,27], and a chelator of heavy metals [28,29]. Additionally, carnosine can modulate the glutamatergic system through the up-regulation of glutamate transporter 1 and the reduction of glutamate levels in the central nervous system (CNS) [30].
Taking into consideration all the above-mentioned activities of carnosine, it is clear why numerous research groups are working on carnosine and believe in its high therapeutic potential. In this regard, I would like to give credit to the enormous efforts made by Alan Hipkiss during the last three decades in describing the potential of carnosine and, in particular, how some of its activities could be useful for the treatment of cancer [10], Parkinson’s disease (PD) [31], depression, diabetes, dementia [32], Alzheimer’s disease (AD) [33], and COVID-19 [34]. Several other authors have contributed significantly to the advancement of our knowledge on the role of carnosine, including Giancarlo Aldini [4,35,36,37], Wim Derave [4,38,39], Craig Sale [40,41,42], Barbora De Courten [37,38,43], Guilherme Artioli [40,41,42], and Alexander Boldyrev [4,44].
2. The Multimodal Mechanism of Action of Carnosine: Contribution from In Vitro Studies
Sometimes it seems that there is a clear separation between those who believe in in vitro studies and those who believe only in in vivo research and consider the latter to be the most important step for the transition to studies on human beings; in vitro studies have developed a reputation for being “less translatable” to humans. The truth is that both are interrelated and indispensable to fully understanding the therapeutic potential of a molecule of interest. In vitro studies have several advantages over in vivo research; for example, they allow the tight control of the chemical and physical environment; reduce the research costs; provide the opportunity to obtain a higher throughput; and minimize the use of animals, which is nowadays strongly limited by the growing ethical concerns. Additionally, as has been demonstrated in carnosine research but is applicable to scientific research as a whole, in vitro experiments allow the in-depth study of the different mechanisms of action related to a specific phenomenon, which is often very difficult or only partially possible in vivo.
There is continually increasing evidence that inflammation [45,46,47], oxidative stress [48,49,50], and aberrant aggregation and accumulation of proteins [51,52] significantly contribute to numerous systemic and neurodegenerative disorders such as diabetes mellitus type 2 (T2DM) [53,54], PD [55,56], and AD [57,58,59]. In this context, the well-known and frequently cited anti-oxidant, anti-inflammatory, and anti-aggregant activities of carnosine, which underline its multimodal mechanism of action, have been considered.
A plethora of in vitro studies have shown the various protective activities of carnosine in multiple heterogeneous cell types, such as macrophages/microglia [21,60,61,62], myocytes [63], skeletal muscle myoblasts [64], podocytes [65], endothelial cells [66], pancreatic β-cells [67], chondrocytes [68], fibroblasts [69], hepatic cells [70], lymphocytes [71], erythrocytes [72], astrocytes [30,73], neuron-like cells [74,75], and stem cells [76]. The “coverage” of the very wide range of cell types demonstrated in vitro, representative of different body districts, strengthens the idea that carnosine has the potential to exert therapeutic effects in a broad spectrum of pathological conditions.
Despite the considerable number of in vitro experiments that have been performed and the diversity of the cell types considered, similar concentrations of carnosine (millimolar order of magnitude) and durations of treatment (often within 24 h) have been used in most studies, allowing the results to be reproduced and compared and establishing a common basis for in vivo experiments.
I will now discuss the therapeutic potential of carnosine as demonstrated in numerous clinical trials, in which greater heterogeneity, specifically in terms of methodology, can be observed compared to the in vitro studies.
3. Clinical Trials of Carnosine: What Is the Basis for the Heterogeneity?
According to www.clinicaltrials.gov (accessed on 15 April 2022), a service provided by the U.S. National Institute of Health that shares information on current clinical trials, there are currently 32 studies at different stages (not yet recruiting, recruiting, completed, unknown, etc.) using carnosine, its analog zinc L-carnosine (polaprezinc [77]), β-alanine, or carnosine-rich foods for the treatment of various diseases (peripheral arterial disease, bipolar I disorder, schizophrenia, AD, multiple sclerosis, etc.).
In addition to the above, numerous clinical trials have been conducted to explore the therapeutic effects of carnosine in a wide range of diseases/pathological conditions (including age-related conditions). For example, Gulf War illness (carnosine 1500 mg/day; 12 weeks (dose-escalation study)) [78]; mild cognitive impairment (MCI) (anserine:carnosine 750 mg:250 mg/day; 12 weeks) [79]; health status of elderly people treated with a pill-based nutraceutical (NT-020) containing carnosine (unspecified dosage; 2 months) [80], a formulation (formula F) containing carnosine (100 mg; 6 months) [81], or carnosine:anserine (250–350:650–750 mg/day; 13 weeks) [82,83,84]; depression (carnosine 400 mg twice daily; 6 weeks) [85]; T2DM (carnosine 1 g/day; 12 weeks) [86]; binge eating disorder and bulimia nervosa (polaprezinc 150 mg/day: 34 mg zinc and 116 mg L-carnosine; 16 weeks); autism spectrum disorder (carnosine 10–15 mg/kg/day plus standard care treatment; 2 months) [87]; and attention-deficit/hyperactivity disorder (carnosine 800 mg/day; 8 weeks [88]. As can be seen, carnosine has been co-administered with anserine in multiple studies. This natural derivative of carnosine, with equivalent physiological functions [4], is usually adopted because it is not cleaved by human carnosinase, which is abundant in human serum and is known to strongly reduce carnosine bioavailability [89].
One of the reasons why so many studies have been carried out on carnosine may be because both preclinical [90] and clinical [91,92] studies have demonstrated that this dipeptide is essentially non-toxic and well-tolerated, without any known drug interactions or dangerous side effects.
A detailed analysis of the clinical trials reveals substantial heterogeneity among the studies carried out on carnosine, alone or in combination with other molecules. Inevitably, studies will differ in terms of both clinical heterogeneity, due to the variability of participants, interventions, and outcomes, and methodological heterogeneity, due to the differences in the study designs and risk of bias. As a concrete example, my colleagues and I recently published a systematic review with meta-analysis on the therapeutic potential of carnosine/anserine supplementation against cognitive decline [93]. Despite the reasonably high number of studies that resulted from the systematic search (516), only 36 were still considered after an initial (i.e., title and abstract) evaluation. Unfortunately, 31 of these 36 studies did not meet the pre-specified inclusion criteria; we were forced to exclude most of the articles because they: (a) reported the acute effects of carnosine supplementation; (b) included results from children; (c) reported other outcomes (e.g., quality of life or cognitive performance); (d) did not explore the outcomes of interest; or (e) were partially conducted on the same patients. In the end, we included only five studies [79,81,83,94,95] in the systematic review, and only three provided sufficient statistical data to be included in the quantitative analysis.
At this point, we should carefully and critically consider what lies “between” in vitro studies and clinical trials, that is, in vivo research, which is the truly pivotal stage.
4. In Vivo Preclinical Studies: Administration Route, Dosage, Treatment Duration, and Selected Animal Model. Are We All Converging in the Same Direction?
Despite the limits imposed by ethical concerns, the coupling of in vivo and in vitro experiments is of utmost importance for fully understanding the therapeutic potential of a candidate molecule. The use of animals provides an opportunity to addresses many of the shortcomings of in vitro studies, particularly by allowing a more accurate evaluation of the safety, toxicity, and efficacy in a complex model. Additionally, advances in the “modulation of animal genotypes” have helped researchers to replicate human diseases with very high accuracy.
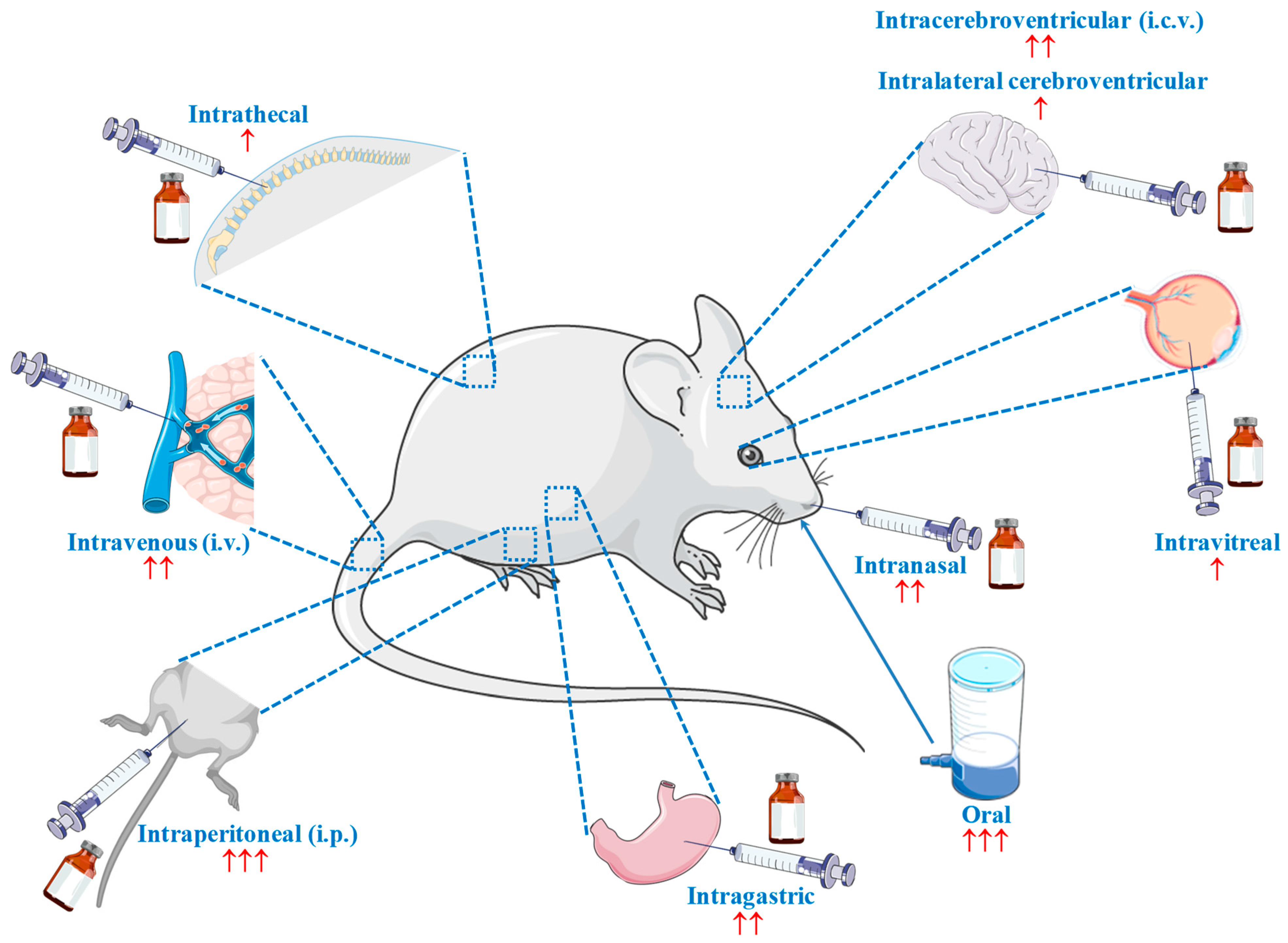
Since the publication of Tomonaga et al. in 2004 [96], which investigated the effect of the central administration of carnosine and its constituents on the behavior of chicks, more than 140 in vivo research studies have been published describing the therapeutic potential of carnosine for numerous diseases, such as stroke [97], diabetes [98], depression [99], liver injury [100], hypoxia–ischemia [101], AD [102], dyslipidaemia [103], atherosclerosis [104], myocardial infarction [105], PD [106], septic shock [107], autism spectrum disorder [108], and autoimmune encephalomyelitis [109]. This incomplete list of disorders is enough to clearly illustrate the substantial attention that this molecule has garnered. At the same time, the great variability of the route of carnosine administration stands out as a crucial point that deserves to be discussed. Researchers have tested many different administration routes (and related formulations), including intranasal, oral, intracerebroventricular (i.c.v.), intraperitoneal (i.p.), intravenous (i.v.), intralateral cerebroventricular, intravitreal, intragastric, and intrathecal (Figure 1), with oral and i.p. being the most widely employed.
One might assume that this heterogeneity arose due to the “physiological variability” of the different animal models; however, substantial discrepancies can be found not only within the same animal model, but also within the same strain, as in the case of C57BL/6 mice, for which i.p. [110], oral [111], and i.v. [112] administration routes have been used. Furthermore, this situation highlights a major unmet need in the in vivo research on carnosine—the lack of studies on its bioavailability after a specific administration process. Often, we are only able to observe the endpoint effects, which cannot be correlated to a specific carnosine concentration in a specific area of the body. The implementation of pharmacokinetic studies showing how a specific administration route influences the quantity of the molecule that reaches specific organs and tissues will help us understand more comprehensively the therapeutic potential of carnosine and evaluate more accurately the results already obtained by research groups. Supposing that we manage to administer a certain amount of carnosine into the brain, the next question is “how much carnosine will be able to overcome a selective barrier such as the blood–brain barrier (BBB)?”.
Two additional critical factors, characterized by substantial variability and related “comparison/translational” issues, are dosage and treatment duration. It can be expected that some of the differences observed according to dosage are caused by the administration route selected; for example, “mg/kg of body weight” is often used for i.p. [100,113,114], while “g/L” is frequently used for oral administration through drinking water [115,116,117]. Nevertheless, as observed in the case of animal strains, there is also a source of “internal” variability that complicates the comparison of results between different studies. In studies involving oral (drinking water) administration, one of the two “often used” (Figure 1) administration routes, the dosage amounts vary widely, ranging from 0.5 g/L [98,116] to 30 g/L (60 times higher!) [109,118]. Furthermore, in the context of oral administration, at least five different units of measurement can be found within the research studies: mg or g/Kg [119,120,121], mg or g/L [98,115,116], mmol/L or Kg [99,122,123,124], mM [102,125,126], or % [127,128,129]. This, along with the wide range of treatment durations (from 1 [120] to 50 weeks [130]), reinforces the need for the alignment of future studies, which will likely lead to even more surprising results with regard to the therapeutic potential of carnosine.
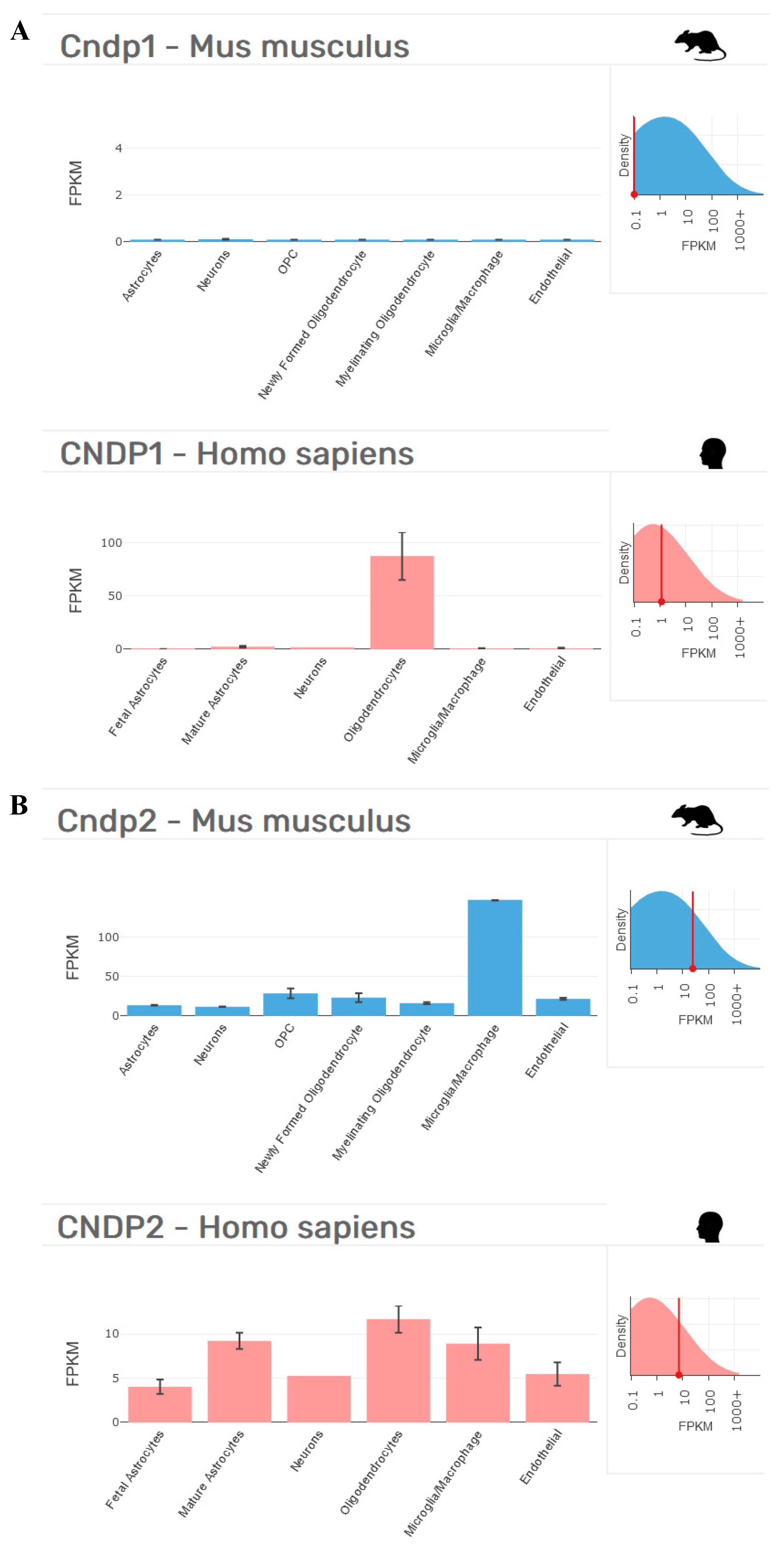
When performing in vivo studies, an additional drawback could be represented by the animal model selected. In this context, it is worth recalling that the administration of carnosine in humans only leads to a small increase in circulating carnosine. When we consider both the bioavailability and high therapeutic potential of carnosine in rodents (mice and rats) and humans, it is important to consider that rodents lack the signal peptide in the CNDP1 gene (CTG)5 and consequently do not have circulating CNDP1 enzymes [4] (see Figure 2 for an example).
These interspecies differences could explain why the levels of carnosine increase in rodents (>120 studies carried out since 2005) after oral carnosine supplementation [125], a phenomenon that does not occur in humans, who only experience a small increase in circulating carnosine [133]. The results obtained in rodents could represent an overestimation of the therapeutic potential of carnosine that will not be confirmed by a translational approach, moving from mice/rats to humans.
Consequently, different research groups are currently working on the development of new approaches and new formulations of carnosine that are able to improve its bioavailability and/or reach a specific target (drug delivery systems). One potential approach to increasing carnosine bioavailability is the use of selective inhibitors of carnosinases (especially CNDP1), as this has been achieved using carnostatine in combination with carnosine [134]. Similarly, Anderson and collaborators described the rational design, characterization, and pharmacological evaluation of carnosinol, a derivative of carnosine with high oral bioavailability that is resistant to carnosinases [135]. By employing a rodent model of diet-induced obesity and metabolic syndrome, the authors demonstrated the ability of carnosinol to dose-dependently attenuate 4-hydroxynonenal adduct formation in liver and skeletal muscle, while simultaneously mitigating inflammation, dyslipidemia, insulin resistance, and steatohepatitis. As described by Grasso et al. [136], alternative approaches to increase carnosine delivery and its bioavailability include the use of carnosine derivatives [137,138,139]; vesicular systems (nanoliposomes, niosomes, and polymerosomes) [140,141,142,143]; and nanoparticulate systems [144,145,146,147]. The currently “under-used” intranasal administration route might also represent an innovative approach, since it can purportedly bypass the BBB and first-pass metabolism [148,149]; the vasodilatory activity of carnosine [150] also makes this endogenous dipeptide an attractive candidate for this kind of delivery. The intranasal administration of carnosine has been successfully adopted in two recent studies carried out by Bermúdez et al. in a Thy1-aSyn mouse model of PD characterized by the overexpression of human alpha-synuclein [148], the aberrant aggregation of which has been recognized as a key contributor to the neurodegenerative process observed in PD [151,152].
As previously mentioned, numerous in vivo studies have been carried out to demonstrate how exogenously administered carnosine can improve pathological conditions. However, the physiological role of carnosine and related histidine-containing dipeptides is not fully understood and should receive more attention. In this regard, substantial improvements could arise from the use of transgenic and knockout (KO) animal models. Very recently, Gonçalves et al. used a novel CARNS1 KO rat model to demonstrate that histidine dipeptides, including carnosine, are key regulators of excitation–contraction coupling in cardiac muscle [129]. Eckhardt and co-workers produced a mouse line deficient in CARNS1 and showed that CARNS1 deficiency is compatible with normal skeletal muscle and olfactory function but causes reduced olfactory sensitivity [153].
During the drafting of this position paper, a final point that drew my attention and that I would like to focus on relates to the use of anserine in combination with carnosine. What really surprised me is that, despite the fact that a large number of clinical trials have considered (or are still considering) this combination of molecules [79,82,83,84,92,94,154], only a few studies (I would say “close to zero”; see Qi et al. for an example almost impossible to find [155]) have investigated this kind of treatment at the preclinical level. Now, the question is “should clinical trials not be guided by preclinical ones?”. The obvious answer to this question should encourage us to think deeply about how we can more effectively connect and align future studies.
5. Concluding Remarks
Since the 1950s, more than a thousand research studies have been published on the structure, role, function, and biological activities of carnosine under numerous experimental and clinical conditions. As discussed above, a plethora of in vitro, in vivo, and clinical studies have been carried out showing the multimodal mechanism of action of carnosine, including anti-aggregant, anti-oxidant, and anti-inflammatory activities, which are all of great interest for numerous systemic and neurodegenerative disorders, such T2DM, PD, and AD. Despite this, as I attempted to underline in this position paper, advances need to be made in order to fully unveil the enormous therapeutic potential of this dipeptide, specifically in the context of in vivo studies, which are currently characterized by substantial heterogeneity regarding administration route, dosage, treatment duration, and animal model. The most urgent need is to perform pharmacokinetic studies to determine how the bioavailability of carnosine is connected to specific administration routes, dosages, and treatment durations. These studies will help to evaluate more accurately the existing results and plan future studies. Finally, there is also a need for “coordinated/aligned” preclinical studies laying the foundations for well-defined future clinical trials.
Funding
This research was funded by the Italian Ministry of Health Research Program 2018, grant number RC: 2635256.
Data Availability Statement
Not applicable.
Acknowledgments
I would like to thank Anna Privitera from the Department of Drug and Health Sciences, University of Catania (Italy) for her valuable assistance during the literature search for in vivo studies. Part of Figure 1 was generated using Servier Medical Art, available at smart.servier.com (accessed on 20 April 2022). Figure 2 was generated using the Barres group transcriptomes, available at www.brainrnaseq.org (accessed on 17 April 2022).
Conflicts of Interest
The author declares no conflict of interest.
References
- Gulewitsch, W.; Amiradžibi, S. Ueber das carnosin, eine neue organische base des fleischextractes. Ber. Der Dtsch. Chem. Ges. 1900, 33, 1902–1903. [Google Scholar] [CrossRef] [Green Version]
- Kalyankar, G.D.; Meister, A. Enzymatic synthesis of carnosine and related β-alanyl and γ-aminobutyryl peptides. J. Biol. Chem. 1959, 234, 3210–3218. [Google Scholar] [CrossRef]
- Winnick, R.; Winnick, T. Carnosine-anserine synthetase of muscle i. Preparation and properties of a soluble enyzme from chick muscle. Biochim. Biophys. Acta 1959, 31, 47–55. [Google Scholar] [CrossRef]
- Boldyrev, A.A.; Aldini, G.; Derave, W. Physiology and pathophysiology of carnosine. Physiol. Rev. 2013, 93, 1803–1845. [Google Scholar] [CrossRef] [PubMed]
- Gariballa, S.E.; Sinclair, A.J. Carnosine: Physiological properties and therapeutic potential. Age Ageing 2000, 29, 207–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hipkiss, A.R.; Preston, J.E.; Himsworth, D.T.; Worthington, V.C.; Keown, M.; Michaelis, J.; Lawrence, J.; Mateen, A.; Allende, L.; Eagles, P.A.; et al. Pluripotent protective effects of carnosine, a naturally occurring dipeptide. Ann. N. Y. Acad. Sci. 1998, 854, 37–53. [Google Scholar] [CrossRef]
- Lenney, J.F.; George, R.P.; Weiss, A.M.; Kucera, C.M.; Chan, P.W.; Rinzler, G.S. Human serum carnosinase: Characterization, distinction from cellular carnosinase, and activation by cadmium. Clin. Chim. Acta 1982, 123, 221–231. [Google Scholar] [CrossRef]
- Lenney, J.F.; Peppers, S.C.; Kucera-Orallo, C.M.; George, R.P. Characterization of human tissue carnosinase. Biochem. J. 1985, 228, 653–660. [Google Scholar] [CrossRef] [Green Version]
- Teufel, M.; Saudek, V.; Ledig, J.P.; Bernhardt, A.; Boularand, S.; Carreau, A.; Cairns, N.J.; Carter, C.; Cowley, D.J.; Duverger, D.; et al. Sequence identification and characterization of human carnosinase and a closely related non-specific dipeptidase. J. Biol. Chem. 2003, 278, 6521–6531. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.D.; Sale, C.; Garner, A.C.; Hipkiss, A.R. Anti-cancer actions of carnosine and the restoration of normal cellular homeostasis. Biochim. Biophys. Acta Mol. Cell. Res. 2021, 1868, 119117. [Google Scholar] [CrossRef]
- Severin, S.E.; Kirzon, M.V.; Kaftanova, T.M. Effect of carnosine and anserine on action of isolated frog muscles. Dokl. Akad. Nauk SSSR 1953, 91, 691–694. [Google Scholar] [PubMed]
- Stvolinskiĭ, S.L.; Dobrota, D.; Mezeshova, V.; Liptaĭ, T.; Pronaĭova, N.; Zalibera, L.; Boldyrev, A.A. Carnosine and anserine in working muscles—study using proton nmr spectroscopy. Biokhimiia 1992, 57, 1317–1323. [Google Scholar] [PubMed]
- Boldyrev, A.A.; Petukhov, V.B. Localization of carnosine effect on the fatigued muscle preparation. Gen. Pharmacol. 1978, 9, 17–20. [Google Scholar] [CrossRef]
- Rubtsov, A.M. Molecular mechanisms of regulation of the activity of sarcoplasmic reticulum ca-release channels (ryanodine receptors), muscle fatigue, and severin’s phenomenon. Biochemistry 2001, 66, 1132–1143. [Google Scholar]
- Brisola, G.M.P.; de Souza Malta, E.; Santiago, P.R.P.; Vieira, L.H.P.; Zagatto, A.M. B-alanine supplementation’s improvement of high-intensity game activities in water polo. Int. J. Sports Physiol. Perform. 2018, 13, 1208–1214. [Google Scholar] [CrossRef]
- De Andrade Kratz, C.; de Salles Painelli, V.; de Andrade Nemezio, K.M.; da Silva, R.P.; Franchini, E.; Zagatto, A.M.; Gualano, B.; Artioli, G.G. Beta-alanine supplementation enhances judo-related performance in highly-trained athletes. J. Sci. Med. Sport 2017, 20, 403–408. [Google Scholar] [CrossRef] [Green Version]
- Furst, T.; Massaro, A.; Miller, C.; Williams, B.T.; LaMacchia, Z.M.; Horvath, P.J. B-alanine supplementation increased physical performance and improved executive function following endurance exercise in middle aged individuals. J. Int. Soc. Sports Nutr. 2018, 15, 32. [Google Scholar] [CrossRef] [Green Version]
- Glenn, J.M.; Smith, K.; Moyen, N.E.; Binns, A.; Gray, M. Effects of acute beta-alanine supplementation on anaerobic performance in trained female cyclists. J. Nutr. Sci. Vitam. 2015, 61, 161–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiedje, K.; Stevens, K.; Barnes, S.; Weaver, D. B-alanine as a small molecule neurotransmitter. Neurochem. Int. 2010, 57, 177–188. [Google Scholar] [CrossRef]
- Fresta, C.G.; Fidilio, A.; Lazzarino, G.; Musso, N.; Grasso, M.; Merlo, S.; Amorini, A.M.; Bucolo, C.; Tavazzi, B.; Lazzarino, G.; et al. Modulation of pro-oxidant and pro-inflammatory activities of m1 macrophages by the natural dipeptide carnosine. Int. J. Mol. Sci. 2020, 21, 776. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Fresta, C.G.; Fidilio, A.; O’Donnell, F.; Musso, N.; Lazzarino, G.; Grasso, M.; Amorini, A.M.; Tascedda, F.; Bucolo, C.; et al. Carnosine decreases pma-induced oxidative stress and inflammation in murine macrophages. Antioxidants 2019, 8, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mal’tseva, V.V.; Sergienko, V.V.; Stvolinskii, S.L. The effect of carnosine on hematopoietic stem cell activity in irradiated animals. Biokhimiia 1992, 57, 1378–1382. [Google Scholar] [PubMed]
- Fresta, C.G.; Chakraborty, A.; Wijesinghe, M.B.; Amorini, A.M.; Lazzarino, G.; Lazzarino, G.; Tavazzi, B.; Lunte, S.M.; Caraci, F.; Dhar, P.; et al. Non-toxic engineered carbon nanodiamond concentrations induce oxidative/nitrosative stress, imbalance of energy metabolism, and mitochondrial dysfunction in microglial and alveolar basal epithelial cells. Cell Death Dis. 2018, 9, 245. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Fresta, C.G.; Martinez-Becerra, F.; Antonio, L.; Johnson, R.T.; de Campos, R.P.S.; Siegel, J.M.; Wijesinghe, M.B.; Lazzarino, G.; Lunte, S.M. Carnosine modulates nitric oxide in stimulated murine raw 264.7 macrophages. Mol. Cell. Biochem. 2017, 431, 197–210. [Google Scholar] [CrossRef]
- Caruso, G.; Benatti, C.; Musso, N.; Fresta, C.G.; Fidilio, A.; Spampinato, G.; Brunello, N.; Bucolo, C.; Drago, F.; Lunte, S.M.; et al. Carnosine protects macrophages against the toxicity of aβ1-42 oligomers by decreasing oxidative stress. Biomedicines 2021, 9, 477. [Google Scholar] [CrossRef]
- Pepper, E.D.; Farrell, M.J.; Nord, G.; Finkel, S.E. Antiglycation effects of carnosine and other compounds on the long-term survival of escherichia coli. Appl. Env. Microbiol. 2010, 76, 7925–7930. [Google Scholar] [CrossRef] [Green Version]
- Boldyrev, A.A.; Gallant, S.C.; Sukhich, G.T. Carnosine, the protective, anti-aging peptide. Biosci. Rep. 1999, 19, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Hasanein, P.; Felegari, Z. Chelating effects of carnosine in ameliorating nickel-induced nephrotoxicity in rats. Can. J. Physiol. Pharm. 2017, 95, 1426–1432. [Google Scholar] [CrossRef]
- Brown, C.E.; Antholine, W.E. Chelation chemistry of carnosine. Evidence that mixed complexes may occur in vivo. J. Phys. Chem. 1979, 83, 3314–3319. [Google Scholar] [CrossRef]
- Ouyang, L.; Tian, Y.; Bao, Y.; Xu, H.; Cheng, J.; Wang, B.; Shen, Y.; Chen, Z.; Lyu, J. Carnosine decreased neuronal cell death through targeting glutamate system and astrocyte mitochondrial bioenergetics in cultured neuron/astrocyte exposed to ogd/recovery. Brain Res. Bull. 2016, 124, 76–84. [Google Scholar] [CrossRef]
- Hipkiss, A.R. Glycotoxins: Dietary and metabolic origins; possible amelioration of neurotoxicity by carnosine, with special reference to parkinson’s disease. Neurotox. Res. 2018, 34, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Hipkiss, A.R. Depression, diabetes and dementia: Formaldehyde may be a common causal agent; could carnosine, a pluripotent peptide, be protective? Aging Dis. 2017, 8, 128–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hipkiss, A.R. Could carnosine or related structures suppress alzheimer’s disease? J. Alzheimers Dis. 2007, 11, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Hipkiss, A.R. COVID-19 and senotherapeutics: Any role for the naturally-occurring dipeptide carnosine? Aging Dis. 2020, 11, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Aldini, G.; de Courten, B.; Regazzoni, L.; Gilardoni, E.; Ferrario, G.; Baron, G.; Altomare, A.; D’Amato, A.; Vistoli, G.; Carini, M. Understanding the antioxidant and carbonyl sequestering activity of carnosine: Direct and indirect mechanisms. Free Radic. Res. 2021, 55, 321–330. [Google Scholar] [CrossRef]
- Vistoli, G.; Aldini, G.; Fumagalli, L.; Dallanoce, C.; Angeli, A.; Supuran, C.T. Activation effects of carnosine- and histidine-containing dipeptides on human carbonic anhydrases: A comprehensive study. Int. J. Mol. Sci. 2020, 21, 1761. [Google Scholar] [CrossRef] [Green Version]
- Baye, E.; Ukropcova, B.; Ukropec, J.; Hipkiss, A.; Aldini, G.; de Courten, B. Physiological and therapeutic effects of carnosine on cardiometabolic risk and disease. Amino Acids 2016, 48, 1131–1149. [Google Scholar] [CrossRef]
- Derave, W.; De Courten, B.; Baba, S.P. An update on carnosine and anserine research. Amino Acids 2019, 51, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Blancquaert, L.; Everaert, I.; Derave, W. Beta-alanine supplementation, muscle carnosine and exercise performance. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 63–70. [Google Scholar] [CrossRef]
- Matthews, J.J.; Artioli, G.G.; Turner, M.D.; Sale, C. The physiological roles of carnosine and β-alanine in exercising human skeletal muscle. Med. Sci. Sports Exerc. 2019, 51, 2098–2108. [Google Scholar] [CrossRef]
- Artioli, G.G.; Sale, C.; Jones, R.L. Carnosine in health and disease. Eur. J.Sport Sci. 2019, 19, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Saunders, B.; Elliott-Sale, K.; Artioli, G.G.; Swinton, P.A.; Dolan, E.; Roschel, H.; Sale, C.; Gualano, B. B-alanine supplementation to improve exercise capacity and performance: A systematic review and meta-analysis. Br. J. Sports Med. 2017, 51, 658–669. [Google Scholar] [CrossRef] [PubMed]
- Schön, M.; Mousa, A.; Berk, M.; Chia, W.L.; Ukropec, J.; Majid, A.; Ukropcová, B.; de Courten, B. The potential of carnosine in brain-related disorders: A comprehensive review of current evidence. Nutrients 2019, 11, 1196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boldyrev, A.A. Carnosine: New concept for the function of an old molecule. Biochemistry 2012, 77, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Marsland, A.L.; Gianaros, P.J.; Kuan, D.C.; Sheu, L.K.; Krajina, K.; Manuck, S.B. Brain morphology links systemic inflammation to cognitive function in midlife adults. Brain Behav. Immun. 2015, 48, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caruso, G.; Fresta, C.G.; Grasso, M.; Santangelo, R.; Lazzarino, G.; Lunte, S.M.; Caraci, F. Inflammation as the common biological link between depression and cardiovascular diseases: Can carnosine exert a protective role? Curr. Med. Chem. 2020, 27, 1782–1800. [Google Scholar] [CrossRef]
- Bettcher, B.M.; Kramer, J.H. Longitudinal inflammation, cognitive decline, and alzheimer’s disease: A mini-review. Clin. Pharmacol. Ther. 2014, 96, 464–469. [Google Scholar] [CrossRef] [Green Version]
- Morrison, C.D.; Pistell, P.J.; Ingram, D.K.; Johnson, W.D.; Liu, Y.; Fernandez-Kim, S.O.; White, C.L.; Purpera, M.N.; Uranga, R.M.; Bruce-Keller, A.J.; et al. High fat diet increases hippocampal oxidative stress and cognitive impairment in aged mice: Implications for decreased nrf2 signaling. J. Neurochem. 2010, 114, 1581–1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Droge, W.; Schipper, H.M. Oxidative stress and aberrant signaling in aging and cognitive decline. Aging Cell 2007, 6, 361–370. [Google Scholar] [CrossRef]
- Fresta, C.G.; Fidilio, A.; Caruso, G.; Caraci, F.; Giblin, F.J.; Leggio, G.M.; Salomone, S.; Drago, F.; Bucolo, C. A new human blood-retinal barrier model based on endothelial cells, pericytes, and astrocytes. Int. J. Mol. Sci. 2020, 21, 1636. [Google Scholar] [CrossRef] [Green Version]
- Irvine, G.B.; El-Agnaf, O.M.; Shankar, G.M.; Walsh, D.M. Protein aggregation in the brain: The molecular basis for alzheimer’s and parkinson’s diseases. Mol. Med. 2008, 14, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Espa, E.; Clemensson, E.K.H.; Luk, K.C.; Heuer, A.; Björklund, T.; Cenci, M.A. Seeding of protein aggregation causes cognitive impairment in rat model of cortical synucleinopathy. Mov. Disord. 2019, 34, 1699–1710. [Google Scholar] [CrossRef]
- Caruso, G.; Distefano, D.A.; Parlascino, P.; Fresta, C.G.; Lazzarino, G.; Lunte, S.M.; Nicoletti, V.G. Receptor-mediated toxicity of human amylin fragment aggregated by short- and long-term incubations with copper ions. Mol. Cell. Biochem. 2017, 425, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Oguntibeju, O.O. Type 2 diabetes mellitus, oxidative stress and inflammation: Examining the links. Int. J. Physiol. Pathophysiol. Pharm. 2019, 11, 45–63. [Google Scholar]
- Hald, A.; Lotharius, J. Oxidative stress and inflammation in parkinson’s disease: Is there a causal link? Exp. Neurol. 2005, 193, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Niranjan, R. The role of inflammatory and oxidative stress mechanisms in the pathogenesis of parkinson’s disease: Focus on astrocytes. Mol. Neurobiol. 2014, 49, 28–38. [Google Scholar] [CrossRef]
- Agostinho, P.; Cunha, R.A.; Oliveira, C. Neuroinflammation, oxidative stress and the pathogenesis of alzheimer’s disease. Curr. Pharm. Des. 2010, 16, 2766–2778. [Google Scholar] [CrossRef]
- Torrisi, S.A.; Geraci, F.; Tropea, M.R.; Grasso, M.; Caruso, G.; Fidilio, A.; Musso, N.; Sanfilippo, G.; Tascedda, F.; Palmeri, A.; et al. Fluoxetine and vortioxetine reverse depressive-like phenotype and memory deficits induced by aβ(1-42) oligomers in mice: A key role of transforming growth factor-β1. Front. Pharm. 2019, 10, 693. [Google Scholar] [CrossRef]
- Caruso, G.; Grasso, M.; Fidilio, A.; Torrisi, S.A.; Musso, N.; Geraci, F.; Tropea, M.R.; Privitera, A.; Tascedda, F.; Puzzo, D.; et al. Antioxidant activity of fluoxetine and vortioxetine in a non-transgenic animal model of alzheimer’s disease. Front. Pharm. 2021, 12, 809541. [Google Scholar] [CrossRef]
- Caruso, G.; Musso, N.; Grasso, M.; Costantino, A.; Lazzarino, G.; Tascedda, F.; Gulisano, M.; Lunte, S.M.; Caraci, F. Microfluidics as a novel tool for biological and toxicological assays in drug discovery processes: Focus on microchip electrophoresis. Micromachines 2020, 11, 593. [Google Scholar] [CrossRef]
- Caruso, G.; Fresta, C.G.; Siegel, J.M.; Wijesinghe, M.B.; Lunte, S.M. Microchip electrophoresis with laser-induced fluorescence detection for the determination of the ratio of nitric oxide to superoxide production in macrophages during inflammation. Anal. Bioanal. Chem. 2017, 409, 4529–4538. [Google Scholar] [CrossRef] [PubMed]
- Fleisher-Berkovich, S.; Abramovitch-Dahan, C.; Ben-Shabat, S.; Apte, R.; Beit-Yannai, E. Inhibitory effect of carnosine and n-acetyl carnosine on lps-induced microglial oxidative stress and inflammation. Peptides 2009, 30, 1306–1312. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Posa, D.K.; Kumar, V.; Hoetker, D.; Kumar, A.; Ganesan, S.; Riggs, D.W.; Bhatnagar, A.; Wempe, M.F.; Baba, S.P. Carnosine protects cardiac myocytes against lipid peroxidation products. Amino Acids 2019, 51, 123–138. [Google Scholar] [CrossRef] [PubMed]
- Lavilla, C.J.; Billacura, M.P.; Hanna, K.; Boocock, D.J.; Coveney, C.; Miles, A.K.; Foulds, G.A.; Murphy, A.; Tan, A.; Jackisch, L.; et al. Carnosine protects stimulus-secretion coupling through prevention of protein carbonyl adduction events in cells under metabolic stress. Free Radic. Biol. Med. 2021, 175, 65–79. [Google Scholar] [CrossRef]
- Zhao, K.; Li, Y.; Wang, Z.; Han, N.; Wang, Y. Carnosine protects mouse podocytes from high glucose induced apoptosis through pi3k/akt and nrf2 pathways. Biomed. Res. Int. 2019, 2019, 4348973. [Google Scholar] [CrossRef]
- Zhang, L.; Yao, K.; Fan, Y.; He, P.; Wang, X.; Hu, W.; Chen, Z. Carnosine protects brain microvascular endothelial cells against rotenone-induced oxidative stress injury through histamine h1 and h2 receptors in vitro. Clin. Exp. Pharm. Physiol. 2012, 39, 1019–1025. [Google Scholar] [CrossRef]
- Miceli, V.; Pampalone, M.; Frazziano, G.; Grasso, G.; Rizzarelli, E.; Ricordi, C.; Casu, A.; Iannolo, G.; Conaldi, P.G. Carnosine protects pancreatic beta cells and islets against oxidative stress damage. Mol. Cell. Endocrinol. 2018, 474, 105–118. [Google Scholar] [CrossRef]
- Ponist, S.; Drafi, F.; Kuncirova, V.; Mihalova, D.; Rackova, L.; Danisovic, L.; Ondrejickova, O.; Tumova, I.; Trunova, O.; Fedorova, T.; et al. Effect of carnosine in experimental arthritis and on primary culture chondrocytes. Oxid. Med. Cell. Longev. 2016, 2016, 8470589. [Google Scholar] [CrossRef] [Green Version]
- Ooi, T.C.; Chan, K.M.; Sharif, R. Zinc l-carnosine protects ccd-18co cells from l-buthionine sulfoximine-induced oxidative stress via the induction of metallothionein and superoxide dismutase 1 expression. Biol. Trace Elem. Res. 2020, 198, 464–471. [Google Scholar] [CrossRef]
- Schwank-Xu, C.; Forsberg, E.; Bentinger, M.; Zhao, A.; Ansurudeen, I.; Dallner, G.; Catrina, S.B.; Brismar, K.; Tekle, M. L-carnosine stimulation of coenzyme q10 biosynthesis promotes improved mitochondrial function and decreases hepatic steatosis in diabetic conditions. Antioxidants 2021, 10, 793. [Google Scholar] [CrossRef]
- Alpsoy, L.; Akcayoglu, G.; Sahin, H. Anti-oxidative and anti-genotoxic effects of carnosine on human lymphocyte culture. Hum. Exp. Toxicol. 2011, 30, 1979–1985. [Google Scholar] [CrossRef] [PubMed]
- Oppermann, H.; Elsel, S.; Birkemeyer, C.; Meixensberger, J.; Gaunitz, F. Erythrocytes prevent degradation of carnosine by human serum carnosinase. Int. J. Mol. Sci. 2021, 22, 12802. [Google Scholar] [CrossRef] [PubMed]
- Calabrese, V.; Colombrita, C.; Guagliano, E.; Sapienza, M.; Ravagna, A.; Cardile, V.; Scapagnini, G.; Santoro, A.M.; Mangiameli, A.; Butterfield, D.A.; et al. Protective effect of carnosine during nitrosative stress in astroglial cell cultures. Neurochem. Res. 2005, 30, 797–807. [Google Scholar] [CrossRef] [PubMed]
- Tabakman, R.; Lazarovici, P.; Kohen, R. Neuroprotective effects of carnosine and homocarnosine on pheochromocytoma pc12 cells exposed to ischemia. J. Neurosci. Res. 2002, 68, 463–469. [Google Scholar] [CrossRef]
- Kulikova, O.I.; Fedorova, T.N.; Lopachev, A.V.; Orlova, V.S.; Grachev, V.A. Effects of antioxidants on the viability of the human neuroblastoma sh-sy5y cell culture under the conditions of heavy-metal toxicity. Biol. Med. 2016, 8, 1. [Google Scholar] [CrossRef]
- Zhou, J.Y.; Lin, H.L.; Qin, Y.C.; Li, X.G.; Gao, C.Q.; Yan, H.C.; Wang, X.Q. L-carnosine protects against deoxynivalenol-induced oxidative stress in intestinal stem cells by regulating the keap1/nrf2 signaling pathway. Mol. Nutr. Food Res. 2021, 65, e2100406. [Google Scholar] [CrossRef]
- Li, M.; Sun, Z.; Zhang, H.; Liu, Z. Recent advances on polaprezinc for medical use (review). Exp. Ther. Med. 2021, 22, 1445. [Google Scholar] [CrossRef]
- Baraniuk, J.N.; El-Amin, S.; Corey, R.; Rayhan, R.; Timbol, C. Carnosine treatment for gulf war illness: A randomized controlled trial. Glob. J. Health Sci. 2013, 5, 69–81. [Google Scholar] [CrossRef]
- Masuoka, N.; Yoshimine, C.; Hori, M.; Tanaka, M.; Asada, T.; Abe, K.; Hisatsune, T. Effects of anserine/carnosine supplementation on mild cognitive impairment with apoe4. Nutrients 2019, 11, 1626. [Google Scholar] [CrossRef] [Green Version]
- Small, B.J.; Rawson, K.S.; Martin, C.; Eisel, S.L.; Sanberg, C.D.; McEvoy, C.L.; Sanberg, P.R.; Shytle, R.D.; Tan, J.; Bickford, P.C. Nutraceutical intervention improves older adults’ cognitive functioning. Rejuvenation Res. 2014, 17, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Cornelli, U. Treatment of alzheimer’s disease with a cholinesterase inhibitor combined with antioxidants. Neurodegener. Dis. 2010, 7, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Rokicki, J.; Li, L.; Imabayashi, E.; Kaneko, J.; Hisatsune, T.; Matsuda, H. Daily carnosine and anserine supplementation alters verbal episodic memory and resting state network connectivity in healthy elderly adults. Front. Aging Neurosci. 2015, 7, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szcześniak, D.; Budzeń, S.; Kopeć, W.; Rymaszewska, J. Anserine and carnosine supplementation in the elderly: Effects on cognitive functioning and physical capacity. Arch. Gerontol. Geriatr. 2014, 59, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Hisatsune, T.; Kaneko, J.; Kurashige, H.; Cao, Y.; Satsu, H.; Totsuka, M.; Katakura, Y.; Imabayashi, E.; Matsuda, H. Effect of anserine/carnosine supplementation on verbal episodic memory in elderly people. J. Alzheimers Dis. 2016, 50, 149–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araminia, B.; Shalbafan, M.; Mortezaei, A.; Shirazi, E.; Ghaffari, S.; Sahebolzamani, E.; Mortazavi, S.H.; Shariati, B.; Ardebili, M.E.; Aqamolaei, A.; et al. L-carnosine combination therapy for major depressive disorder: A randomized, double-blind, placebo-controlled trial. J. Affect. Disord. 2020, 267, 131–136. [Google Scholar] [CrossRef]
- Houjeghani, S.; Kheirouri, S.; Faraji, E.; Jafarabadi, M.A. L-carnosine supplementation attenuated fasting glucose, triglycerides, advanced glycation end products, and tumor necrosis factor-alpha levels in patients with type 2 diabetes: A double-blind placebo-controlled randomized clinical trial. Nutr. Res. 2018, 49, 96–106. [Google Scholar] [CrossRef]
- Ann Abraham, D.; Narasimhan, U.; Christy, S.; Muhasaparur Ganesan, R. Effect of l-carnosine as adjunctive therapy in the management of children with autism spectrum disorder: A randomized controlled study. Amino Acids 2020, 52, 1521–1528. [Google Scholar] [CrossRef]
- Ghajar, A.; Aghajan-Nashtaei, F.; Afarideh, M.; Mohammadi, M.R.; Akhondzadeh, S. L-carnosine as adjunctive therapy in children and adolescents with attention-deficit/hyperactivity disorder: A randomized, double-blind, placebo-controlled clinical trial. J. Child Adolesc. Psychopharmacol. 2018, 28, 331–338. [Google Scholar] [CrossRef]
- Kubomura, D.; Matahira, Y.; Masui, A.; Matsuda, H. Intestinal absorption and blood clearance of l-histidine-related compounds after ingestion of anserine in humans and comparison to anserine-containing diets. J. Agric. Food Chem. 2009, 57, 1781–1785. [Google Scholar] [CrossRef]
- Boldyrev, A.A. Carnosine and Oxidative Stress in Cells and Tissues; Nova Publishers: Hapog, NY, USA, 2007. [Google Scholar]
- Gardner, M.L.; Illingworth, K.M.; Kelleher, J.; Wood, D. Intestinal absorption of the intact peptide carnosine in man, and comparison with intestinal permeability to lactulose. J. Physiol. 1991, 439, 411–422. [Google Scholar] [CrossRef]
- Goto, K.; Maemura, H.; Takamatsu, K.; Ishii, N. Hormonal responses to resistance exercise after ingestion of carnosine and anserine. J. Strength Cond. Res. 2011, 25, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Godos, J.; Castellano, S.; Micek, A.; Murabito, P.; Galvano, F.; Ferri, R.; Grosso, G.; Caraci, F. The therapeutic potential of carnosine/anserine supplementation against cognitive decline: A systematic review with meta-analysis. Biomedicines 2021, 9, 253. [Google Scholar] [CrossRef] [PubMed]
- Katakura, Y.; Totsuka, M.; Imabayashi, E.; Matsuda, H.; Hisatsune, T. Anserine/carnosine supplementation suppresses the expression of the inflammatory chemokine ccl24 in peripheral blood mononuclear cells from elderly people. Nutrients 2017, 9, 1199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirotsuki, K.; Nonaka, Y.; Abe, K.; Adachi, S.I.; Adachi, S.; Kuboki, T.; Nakao, M. The effect for japanese workers of a self-help computerized cognitive behaviour therapy program with a supplement soft drink. Biopsychosoc. Med. 2017, 11, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomonaga, S.; Tachibana, T.; Takagi, T.; Saito, E.S.; Zhang, R.; Denbow, D.M.; Furuse, M. Effect of central administration of carnosine and its constituents on behaviors in chicks. Brain Res. Bull. 2004, 63, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Zemke, D.; Krishnamurthy, R.; Majid, A. Carnosine is neuroprotective in a mouse model of stroke. J. Cereb. Blood Flow Metab. 2005, 25, S313. [Google Scholar] [CrossRef]
- Lee, Y.T.; Hsu, C.C.; Lin, M.H.; Liu, K.S.; Yin, M.C. Histidine and carnosine delay diabetic deterioration in mice and protect human low density lipoprotein against oxidation and glycation. Eur. J. Pharm. 2005, 513, 145–150. [Google Scholar] [CrossRef]
- Tomonaga, S.; Yamane, H.; Onitsuka, E.; Yamada, S.; Sato, M.; Takahata, Y.; Morimatsu, F.; Furuse, M. Carnosine-induced antidepressant-like activity in rats. Pharm. Biochem Behav. 2008, 89, 627–632. [Google Scholar] [CrossRef]
- Mehmetçik, G.; Ozdemirler, G.; Koçak-Toker, N.; Cevikbaş, U.; Uysal, M. Role of carnosine in preventing thioacetamide-induced liver injury in the rat. Peptides 2008, 29, 425–429. [Google Scholar] [CrossRef]
- Zhang, X.; Song, L.; Cheng, X.; Yang, Y.; Luan, B.; Jia, L.; Xu, F.; Zhang, Z. Carnosine pretreatment protects against hypoxia-ischemia brain damage in the neonatal rat model. Eur. J. Pharm. 2011, 667, 202–207. [Google Scholar] [CrossRef]
- Corona, C.; Frazzini, V.; Silvestri, E.; Lattanzio, R.; La Sorda, R.; Piantelli, M.; Canzoniero, L.M.; Ciavardelli, D.; Rizzarelli, E.; Sensi, S.L. Effects of dietary supplementation of carnosine on mitochondrial dysfunction, amyloid pathology, and cognitive deficits in 3xtg-ad mice. PLoS ONE 2011, 6, e17971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldini, G.; Orioli, M.; Rossoni, G.; Savi, F.; Braidotti, P.; Vistoli, G.; Yeum, K.J.; Negrisoli, G.; Carini, M. The carbonyl scavenger carnosine ameliorates dyslipidaemia and renal function in zucker obese rats. J. Cell. Mol. Med. 2011, 15, 1339–1354. [Google Scholar] [CrossRef] [PubMed]
- Menini, S.; Iacobini, C.; Ricci, C.; Scipioni, A.; Blasetti Fantauzzi, C.; Giaccari, A.; Salomone, E.; Canevotti, R.; Lapolla, A.; Orioli, M.; et al. D-carnosine octylester attenuates atherosclerosis and renal disease in apoe null mice fed a western diet through reduction of carbonyl stress and inflammation. Br. J. Pharm. 2012, 166, 1344–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evran, B.; Karpuzoğlu, H.; Develi, S.; Kalaz, E.B.; Soluk-Tekkeşin, M.; Olgaç, V.; Doğru-Abbasoğlu, S.; Uysal, M. Effects of carnosine on prooxidant-antioxidant status in heart tissue, plasma and erythrocytes of rats with isoproterenol-induced myocardial infarction. Pharm. Rep. 2014, 66, 81–86. [Google Scholar] [CrossRef]
- Afshin-Majd, S.; Khalili, M.; Roghani, M.; Mehranmehr, N.; Baluchnejadmojarad, T. Carnosine exerts neuroprotective effect against 6-hydroxydopamine toxicity in hemiparkinsonian rat. Mol. Neurobiol. 2015, 51, 1064–1070. [Google Scholar] [CrossRef]
- Sahin, S.; Burukoglu Donmez, D. Effects of carnosine (beta-alanyl-l-histidine) in an experimental rat model of acute kidney injury due to septic shock. Med. Sci. Monit. 2018, 24, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Tsuji, T.; Furuhara, K.; Gerasimenko, M.; Shabalova, A.; Cherepanov, S.M.; Minami, K.; Higashida, H.; Tsuji, C. Oral supplementation with l-carnosine attenuates social recognition deficits in cd157ko mice via oxytocin release. Nutrients 2022, 14, 803. [Google Scholar] [CrossRef]
- Spaas, J.; Van Noten, P.; Keytsman, C.; Nieste, I.; Blancquaert, L.; Derave, W.; Eijnde, B.O. Carnosine and skeletal muscle dysfunction in a rodent multiple sclerosis model. Amino Acids 2021, 53, 1749–1761. [Google Scholar] [CrossRef]
- Ma, J.; Xiong, J.Y.; Hou, W.W.; Yan, H.J.; Sun, Y.; Huang, S.W.; Jin, L.; Wang, Y.; Hu, W.W.; Chen, Z. Protective effect of carnosine on subcortical ischemic vascular dementia in mice. CNS Neurosci. Ther. 2012, 18, 745–753. [Google Scholar] [CrossRef]
- Brown, B.E.; Kim, C.H.; Torpy, F.R.; Bursill, C.A.; McRobb, L.S.; Heather, A.K.; Davies, M.J.; van Reyk, D.M. Supplementation with carnosine decreases plasma triglycerides and modulates atherosclerotic plaque composition in diabetic apo e(-/-) mice. Atherosclerosis 2014, 232, 403–409. [Google Scholar] [CrossRef]
- Kamal, M.A.; Jiang, H.; Hu, Y.; Keep, R.F.; Smith, D.E. Influence of genetic knockout of pept2 on the in vivo disposition of endogenous and exogenous carnosine in wild-type and pept2 null mice. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R986–R991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouad, A.A.; El-Rehany, M.A.; Maghraby, H.K. The hepatoprotective effect of carnosine against ischemia/reperfusion liver injury in rats. Eur. J. Pharm. 2007, 572, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Pekcetin, C.; Kiray, M.; Ergur, B.U.; Tugyan, K.; Bagriyanik, H.A.; Erbil, G.; Baykara, B.; Camsari, U.M. Carnosine attenuates oxidative stress and apoptosis in transient cerebral ischemia in rats. Acta Biol. Hung. 2009, 60, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.L.; Wu, S.T.; Yin, M.C.; Chen, H.T.; Chen, H.C. Protective effects from carnosine and histidine on acetaminophen-induced liver injury. J. Food Sci. 2009, 74, H259–H265. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.H.; Liu, T.C.; Yin, M.C. Beneficial effects of histidine and carnosine on ethanol-induced chronic liver injury. Food Chem. Toxicol. 2008, 46, 1503–1509. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.J.; Kuo, W.W.; Liu, W.H.; Yin, M.C. Antioxidative and anti-inflammatory protection from carnosine in the striatum of mptp-treated mice. J. Agric. Food Chem. 2010, 58, 11510–11516. [Google Scholar] [CrossRef] [PubMed]
- Spaas, J.; Franssen, W.M.A.; Keytsman, C.; Blancquaert, L.; Vanmierlo, T.; Bogie, J.; Broux, B.; Hellings, N.; van Horssen, J.; Posa, D.K.; et al. Carnosine quenches the reactive carbonyl acrolein in the central nervous system and attenuates autoimmune neuroinflammation. J. Neuroinflamm. 2021, 18, 255. [Google Scholar] [CrossRef]
- Fedorova, T.; Macletsova, M.; Kulikov, A.; Stepanova, M.; Boldyrev, A. Carnosine protects from the oxidative stress induced by prenatal hypoxia. Dokl. Biol. Sci. 2006, 408, 207–210. [Google Scholar] [CrossRef]
- Cuzzocrea, S.; Genovese, T.; Failla, M.; Vecchio, G.; Fruciano, M.; Mazzon, E.; Di Paola, R.; Muià, C.; La Rosa, C.; Crimi, N.; et al. Protective effect of orally administered carnosine on bleomycin-induced lung injury. Am. J. Physiol. Lung Cell. Mol. Physiol. 2007, 292, L1095–L1104. [Google Scholar] [CrossRef]
- Mahmoud, A.H. Comparative study between carnosine and fluvastatin in hypercholesterolemic rabbits. J. Appl. Sci. 2006, 6, 1725–1730. [Google Scholar] [CrossRef]
- Sauerhöfer, S.; Yuan, G.; Braun, G.S.; Deinzer, M.; Neumaier, M.; Gretz, N.; Floege, J.; Kriz, W.; van der Woude, F.; Moeller, M.J. L-carnosine, a substrate of carnosinase-1, influences glucose metabolism. Diabetes 2007, 56, 2425–2432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamei, J.; Ohsawa, M.; Miyata, S.; Tanaka, S. Preventive effect of l-carnosine on changes in the thermal nociceptive threshold in streptozotocin-induced diabetic mice. Eur. J. Pharm. 2008, 600, 83–86. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.Y.; Zheng, X.J.; Wang, J. Effects of carnosine on the evoked potentials in hippocampal ca1 region. J. Zhejiang Univ. Sci. B 2009, 10, 505–511. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, T.; Schilperoort, M.; Zhang, S.; Braun, J.D.; Qiu, J.; Rodriguez, A.; Pastene, D.O.; Krämer, B.K.; Köppel, H.; Baelde, H.; et al. Carnosine attenuates the development of both type 2 diabetes and diabetic nephropathy in btbr ob/ob mice. Sci. Rep. 2017, 7, 44492. [Google Scholar] [CrossRef]
- Everaert, I.; He, J.; Hanssens, M.; Stautemas, J.; Bakker, K.; Albrecht, T.; Zhang, S.; van der Stede, T.; Vanhove, K.; Hoetker, D.; et al. Carnosinase-1 overexpression, but not aerobic exercise training, affects the development of diabetic nephropathy in btbr ob/ob mice. Am. J. Physiol. Ren. Physiol. 2020, 318, F1030–F1040. [Google Scholar] [CrossRef]
- Everaert, I.; Stegen, S.; Vanheel, B.; Taes, Y.; Derave, W. Effect of beta-alanine and carnosine supplementation on muscle contractility in mice. Med. Sci. Sports Exerc. 2013, 45, 43–51. [Google Scholar] [CrossRef]
- Stegen, S.; Stegen, B.; Aldini, G.; Altomare, A.; Cannizzaro, L.; Orioli, M.; Gerlo, S.; Deldicque, L.; Ramaekers, M.; Hespel, P.; et al. Plasma carnosine, but not muscle carnosine, attenuates high-fat diet-induced metabolic stress. Appl. Physiol. Nutr. Metab. 2015, 40, 868–876. [Google Scholar] [CrossRef]
- Gonçalves, L.S.; Sales, L.P.; Saito, T.R.; Campos, J.C.; Fernandes, A.L.; Natali, J.; Jensen, L.; Arnold, A.; Ramalho, L.; Bechara, L.R.G.; et al. Histidine dipeptides are key regulators of excitation-contraction coupling in cardiac muscle: Evidence from a novel carns1 knockout rat model. Redox Biol. 2021, 44, 102016. [Google Scholar] [CrossRef]
- Derave, W.; Jones, G.; Hespel, P.; Harris, R.C. Creatine supplementation augments skeletal muscle carnosine content in senescence-accelerated mice (samp8). Rejuvenation Res. 2008, 11, 641–647. [Google Scholar] [CrossRef]
- Zhang, Y.; Sloan, S.A.; Clarke, L.E.; Caneda, C.; Plaza, C.A.; Blumenthal, P.D.; Vogel, H.; Steinberg, G.K.; Edwards, M.S.; Li, G.; et al. Purification and characterization of progenitor and mature human astrocytes reveals transcriptional and functional differences with mouse. Neuron 2016, 89, 37–53. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Chen, K.; Sloan, S.A.; Bennett, M.L.; Scholze, A.R.; O’Keeffe, S.; Phatnani, H.P.; Guarnieri, P.; Caneda, C.; Ruderisch, N.; et al. An rna-sequencing transcriptome and splicing database of glia, neurons, and vascular cells of the cerebral cortex. J. Neurosci. 2014, 34, 11929–11947. [Google Scholar] [CrossRef] [PubMed]
- Everaert, I.; Mooyaart, A.; Baguet, A.; Zutinic, A.; Baelde, H.; Achten, E.; Taes, Y.; De Heer, E.; Derave, W. Vegetarianism, female gender and increasing age, but not cndp1 genotype, are associated with reduced muscle carnosine levels in humans. Amino Acids 2011, 40, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Hauske, S.J.; Zhang, S.; Rodriguez-Niño, A.; Albrecht, T.; Pastene, D.O.; van den Born, J.; van Goor, H.; Ruf, S.; Kohlmann, M.; et al. Identification and characterisation of carnostatine (san9812), a potent and selective carnosinase (cn1) inhibitor with in vivo activity. Amino Acids 2019, 51, 7–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, E.J.; Vistoli, G.; Katunga, L.A.; Funai, K.; Regazzoni, L.; Monroe, T.B.; Gilardoni, E.; Cannizzaro, L.; Colzani, M.; De Maddis, D.; et al. A carnosine analog mitigates metabolic disorders of obesity by reducing carbonyl stress. J. Clin. Investig. 2018, 128, 5280–5293. [Google Scholar] [CrossRef]
- Grasso, M.; Caruso, G.; Godos, J.; Bonaccorso, A.; Carbone, C.; Castellano, S.; Currenti, W.; Grosso, G.; Musumeci, T.; Caraci, F. Improving cognition with nutraceuticals targeting tgf-β1 signaling. Antioxidants 2021, 10, 1075. [Google Scholar] [CrossRef]
- Vecchio, G.; La Mendola, D.; Rizzarelli, E. The synthesis and conformation of β-cyclodextrins functionalized with enantiomers of boc-carnosine. J. Supramol. Chem. 2001, 1, 87–95. [Google Scholar] [CrossRef]
- Lanza, V.; Bellia, F.; D’Agata, R.; Grasso, G.; Rizzarelli, E.; Vecchio, G. New glycoside derivatives of carnosine and analogs resistant to carnosinase hydrolysis: Synthesis and characterization of their copper(ii) complexes. J. Inorg. Biochem. 2011, 105, 181–188. [Google Scholar] [CrossRef]
- Bellia, F.; Oliveri, V.; Rizzarelli, E.; Vecchio, G. New derivative of carnosine for nanoparticle assemblies. Eur. J. Med. Chem. 2013, 70, 225–232. [Google Scholar] [CrossRef]
- Maherani, B.; Arab-Tehrany, E.; Kheirolomoom, A.; Cleymand, F.; Linder, M. Influence of lipid composition on physicochemical properties of nanoliposomes encapsulating natural dipeptide antioxidant l-carnosine. Food Chem 2012, 134, 632–640. [Google Scholar] [CrossRef]
- Maestrelli, F.; Landucci, E.; De Luca, E.; Nerli, G.; Bergonzi, M.C.; Piazzini, V.; Pellegrini-Giampietro, D.E.; Gullo, F.; Becchetti, A.; Tadini-Buoninsegni, F.; et al. Niosomal formulation of a lipoyl-carnosine derivative targeting trpa1 channels in brain. Pharmaceutics 2019, 11, 669. [Google Scholar] [CrossRef] [Green Version]
- Moulahoum, H.; Sanli, S.; Timur, S.; Zihnioglu, F. Potential effect of carnosine encapsulated niosomes in bovine serum albumin modifications. Int. J. Biol. Macromol. 2019, 137, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.S.; Kim, D.; Nyberg, S.; Poma, A.; Cecchin, D.; Jain, S.A.; Kim, K.A.; Shin, Y.J.; Kim, E.H.; Kim, M.; et al. Lrp-1 functionalized polymersomes enhance the efficacy of carnosine in experimental stroke. Sci. Rep. 2020, 10, 699. [Google Scholar] [CrossRef] [PubMed]
- Farid, R.M.; Gaafar, P.M.E.; Hazzah, H.A.; Helmy, M.W.; Abdallah, O.Y. Chemotherapeutic potential of l-carnosine from stimuli-responsive magnetic nanoparticles against breast cancer model. Nanomedicine 2020, 15, 891–911. [Google Scholar] [CrossRef] [PubMed]
- Khramtsov, P.; Barkina, I.; Kropaneva, M.; Bochkova, M.; Timganova, V.; Nechaev, A.; Byzov, I.; Zamorina, S.; Yermakov, A.; Rayev, M. Magnetic nanoclusters coated with albumin, casein, and gelatin: Size tuning, relaxivity, stability, protein corona, and application in nuclear magnetic resonance immunoassay. Nanomaterials 2019, 9, 1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durmus, Z.; Kavas, H.; Baykal, A.; Sozeri, H.; Alpsoy, L.; Çelik, S.; Toprak, M. Synthesis and characterization of l-carnosine coated iron oxide nanoparticles. J. Alloy. Compd. 2011, 509, 2555–2561. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, Y.; Wang, L.; Li, G.; Gao, J.; Wang, Y. Development of l-carnosine functionalized iron oxide nanoparticles loaded with dexamethasone for simultaneous therapeutic potential of blood brain barrier crossing and ischemic stroke treatment. Drug Deliv. 2021, 28, 380–389. [Google Scholar] [CrossRef]
- Bermúdez, M.L.; Skelton, M.R.; Genter, M.B. Intranasal carnosine attenuates transcriptomic alterations and improves mitochondrial function in the thy1-asyn mouse model of parkinson’s disease. Mol. Genet. Metab. 2018, 125, 305–313. [Google Scholar] [CrossRef]
- Jadhav, K.R.; Gambhire, M.N.; Shaikh, I.M.; Kadam, V.J.; Pisal, S.S. Nasal drug delivery system-factors affecting and applications. Curr. Drug Ther. 2007, 2, 27–38. [Google Scholar] [CrossRef]
- Ririe, D.G.; Roberts, P.R.; Shouse, M.N.; Zaloga, G.P. Vasodilatory actions of the dietary peptide carnosine. Nutrition 2000, 16, 168–172. [Google Scholar] [CrossRef]
- Caruso, G.; Godos, J.; Privitera, A.; Lanza, G.; Castellano, S.; Chillemi, A.; Bruni, O.; Ferri, R.; Caraci, F.; Grosso, G. Phenolic acids and prevention of cognitive decline: Polyphenols with a neuroprotective role in cognitive disorders and alzheimer’s disease. Nutrients 2022, 14, 819. [Google Scholar] [CrossRef]
- Caruso, G.; Torrisi, S.A.; Mogavero, M.P.; Currenti, W.; Castellano, S.; Godos, J.; Ferri, R.; Galvano, F.; Leggio, G.M.; Grosso, G.; et al. Polyphenols and neuroprotection: Therapeutic implications for cognitive decline. Pharmacol. Ther. 2022, 232, 108013. [Google Scholar] [CrossRef] [PubMed]
- Wang-Eckhardt, L.; Bastian, A.; Bruegmann, T.; Sasse, P.; Eckhardt, M. Carnosine synthase deficiency is compatible with normal skeletal muscle and olfactory function but causes reduced olfactory sensitivity in aging mice. J. Biol. Chem. 2020, 295, 17100–17113. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Tanigawa, K.; Kaneko, J.; Totsuka, M.; Katakura, Y.; Imabayashi, E.; Matsuda, H.; Hisatsune, T. Anserine/carnosine supplementation preserves blood flow in the prefrontal brain of elderly people carrying apoe e4. Aging Dis. 2018, 9, 334–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Z.; Yu, X.; Xu, P.; Hao, Y.; Pan, X.; Zhang, C. L-homocarnosine, l-carnosine, and anserine attenuate brain oxidative damage in a pentylenetetrazole-induced epilepsy model of ovariectomized rats. 3 Biotech 2018, 8, 363. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Different routes employed for the administration of carnosine in vivo. ↑ = rarely used (1 to 2 times); ↑↑ = infrequently used (3 to 6 times); ↑↑↑ = often used (>50 times). Part of the figure was generated using Servier Medical Art, available at smart.servier.com (accessed on 20 April 2022).
Figure 1.
Different routes employed for the administration of carnosine in vivo. ↑ = rarely used (1 to 2 times); ↑↑ = infrequently used (3 to 6 times); ↑↑↑ = often used (>50 times). Part of the figure was generated using Servier Medical Art, available at smart.servier.com (accessed on 20 April 2022).

Figure 2.
Cellular distribution in the brains of Mus musculus and Homo sapiens of (A) CNDP1 and (B) CNDP2, according to transcriptome studies [131,132]. Fragments per kilobase of transcript per million mapped reads (FPKM) data collected from http://www.brainrnaseq.org on 17 April 2022.
Figure 2.
Cellular distribution in the brains of Mus musculus and Homo sapiens of (A) CNDP1 and (B) CNDP2, according to transcriptome studies [131,132]. Fragments per kilobase of transcript per million mapped reads (FPKM) data collected from http://www.brainrnaseq.org on 17 April 2022.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Caruso, G. Unveiling the Hidden Therapeutic Potential of Carnosine, a Molecule with a Multimodal Mechanism of Action: A Position Paper. Molecules 2022, 27, 3303. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103303
AMA Style
Caruso G. Unveiling the Hidden Therapeutic Potential of Carnosine, a Molecule with a Multimodal Mechanism of Action: A Position Paper. Molecules. 2022; 27(10):3303. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103303
Chicago/Turabian StyleCaruso, Giuseppe. 2022. "Unveiling the Hidden Therapeutic Potential of Carnosine, a Molecule with a Multimodal Mechanism of Action: A Position Paper" Molecules 27, no. 10: 3303. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103303