
Insecticidal Activity of Organic Extracts of Solidago graminifolia and Its Main Metabolites (Quercetin and Chlorogenic Acid) against Spodoptera frugiperda: An In Vitro and In Silico Approach

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Taxonomic and Molecular Identification
2.2. Organic Extracts
2.3. Insecticidal Activity of the Extracts
2.4. Phytochemical Analysis
2.5. Insecticidal Evaluation of Secondary Metabolites
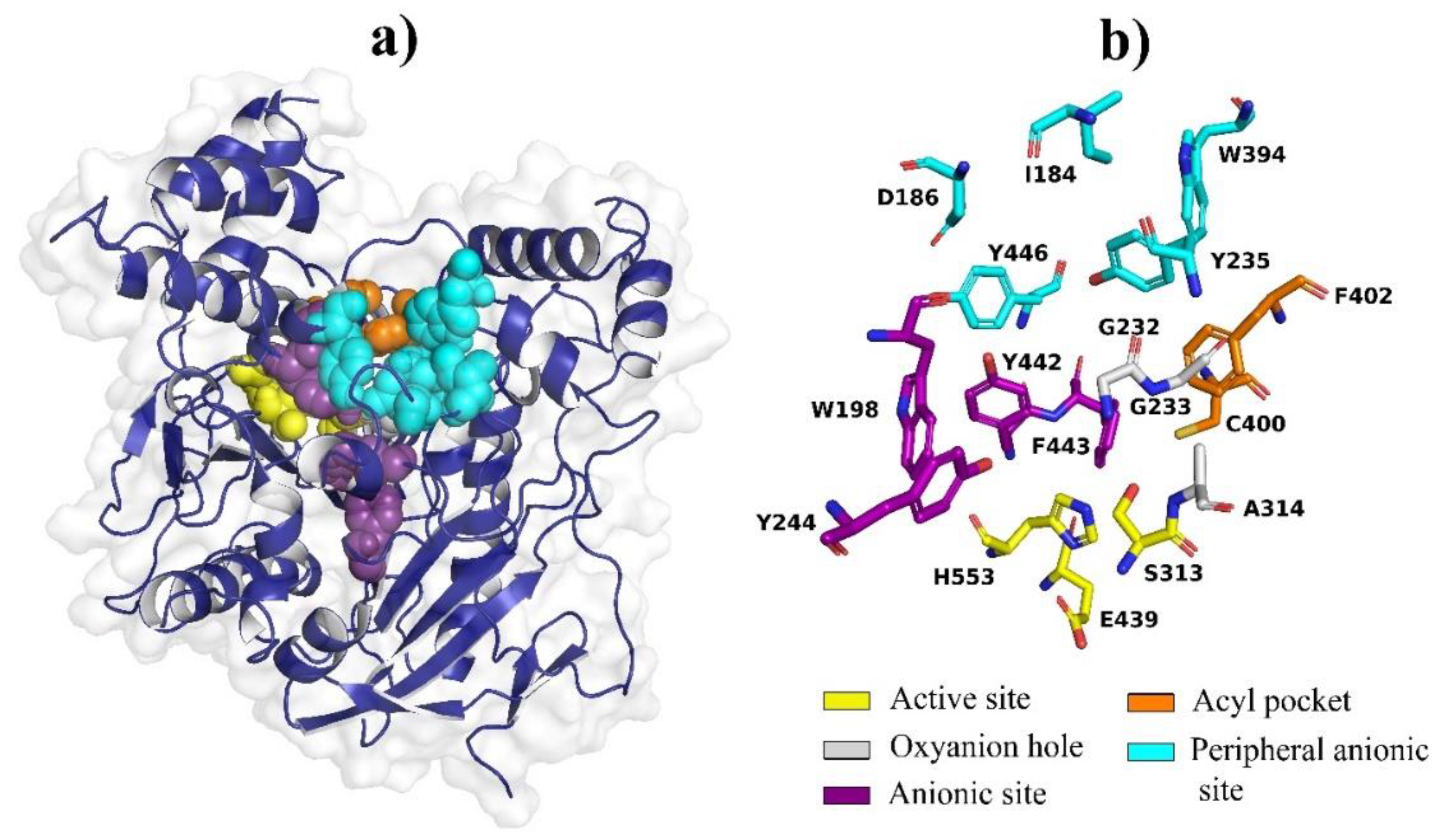
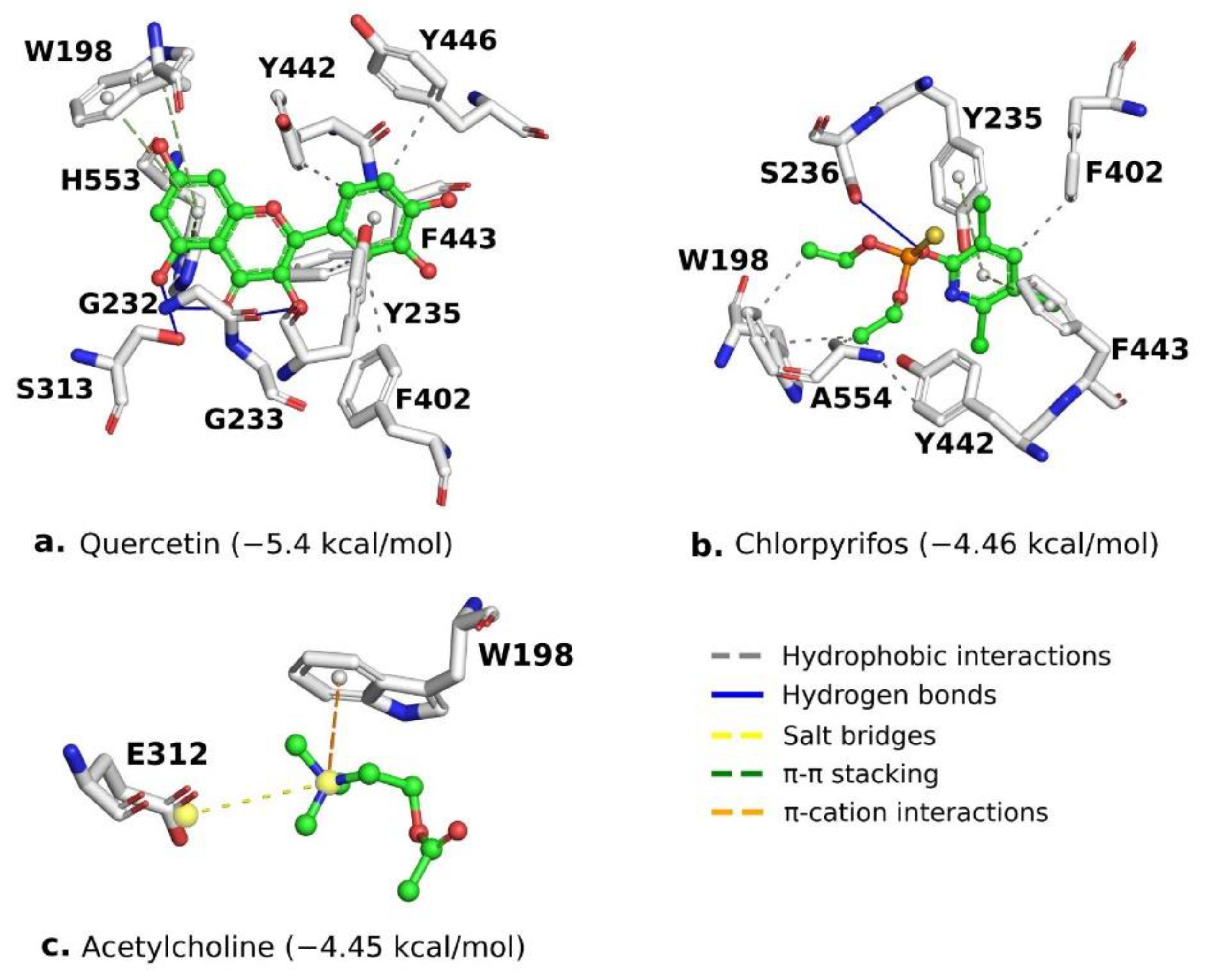
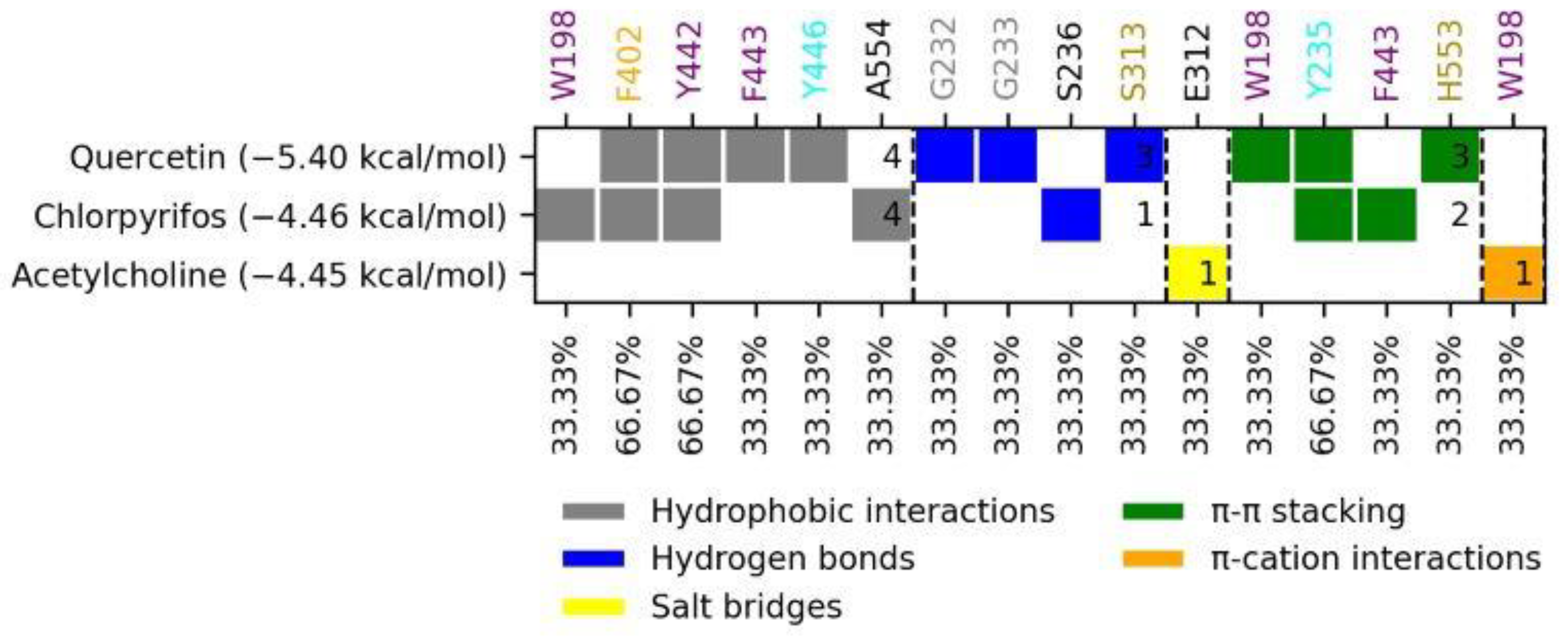
2.6. Molecular Docking Analysis of Quercetin on S. frugiperda Acetylcholinesterase
3. Materials and Methods
3.1. Collection of Plant Material
3.2. Molecular Identification
3.3. Preparation of Organic Extracts
3.4. Ultra-Performance Liquid Chromatography (UPLC)
3.5. Insecticidal Activity
3.6. Molecular Docking Analysis
3.7. Insecticidal Activity of Chlorogenic Acid and Quercetin
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riaz, M.; Rasool, N.; Bukhari, I.H.; Shahid, M.; Zubair, M.; Rizwan, K.; Rashid, U. In Vitro Antimicrobial, Antioxidant, Cytotoxicity and GC-MS Analysis of Mazus goodenifolius. Molecules 2012, 17, 14275–14287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toiu, A.; Vlase, L.; Vodnar, D.C.; Gheldiu, A.-M.; Oniga, I. Solidago graminifolia L. Salisb. (Asteraceae) as a Valuable Source of Bioactive Polyphenols: HPLC Profile, In Vitro Antioxidant and Antimicrobial Potential. Molecules 2019, 24, 2666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krüzselyi, D.; Bakonyi, J.; Ott, P.G.; Darcsi, A.; Csontos, P.; Morlock, G.E.; Móricz, M. Goldenrod Root Compounds Active against Crop Pathogenic Fungi. J. Agric. Food Chem. 2021, 69, 12686–12694. [Google Scholar] [CrossRef] [PubMed]
- Móricz, M.; Jamshidi-Aidji, M.; Krüzselyi, D.; Darcsi, A.; Böszörményi, A.; Csontos, P.; Béni, S.; Ott, P.G.; Morlock, G.E. Distinction and valorization of 30 root extracts of five goldenrod (Solidago) species. J. Chromatogr. A 2020, 1611, 460602. [Google Scholar] [CrossRef]
- Szymura, M.; Szymura, T.H. Interactions between alien goldenrods (Solidago and Euthamia species) and comparison with native species in Central Europe. Flora Morphol. Distrib. Funct. Ecol. Plants 2016, 218, 51–61. [Google Scholar] [CrossRef]
- Fursenco, C.; Calalb, T.; Uncu, L.; Dinu, M.; Ancuceanu, R. Solidago virgaurea L.: A Review of Its Ethnomedicinal Uses, Phytochemistry, and Pharmacological Activities. Biomolecules 2020, 10, 1619. [Google Scholar] [CrossRef]
- Rosado-Solano, D.N.; Barón-Rodríguez, M.A.; Sanabria-Florez, P.L.; Luna-Parada, L.K.; Puerto-Galvis, C.E.; Zor-ro-González, A.F.; Vargas-Méndez, L.Y. Synthesis, Biological Evaluation and In Silico Computational Studies of 7-Chloro-4-(1H-1,2,3-triazol-1-yl) quinoline Derivatives: Search for New Controlling Agents against Spodoptera frugiperda (Lepidoptera: Noctuidae) Larvae. J. Agric. Food Chem. 2019, 33, 9210–9219. [Google Scholar] [CrossRef]
- Calderón, J.S.; Céspedes, C.L.; Rosas, R.; Gómez-Garibay, F.; Salazar, J.R.; Lina, L.; Kubo, I. Acetylcho-linesterase and Insect Growth Inhibitory Activities of Gutierrezia microcephala on Fall Army worm Spodoptera frugiperda JE Smith. Z. Nat. C 2001, 6, 382–394. [Google Scholar]
- Vargas-Méndez, L.Y.; Sanabria-Flórez, P.L.; Saavedra-Reyes, L.M.; Merchan-Arenas, D.R.; Kouznetsov, V.V. Bioactivity of semisynthetic eugenol derivatives against Spodoptera frugiperda (Lepidoptera: Noctuidae) larvae infesting maize in Colombia. Saudi J. Biol. Sci. 2018, 26, 1613–1620. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; De Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Developmental Parameters of Spodoptera frugiperda (Lepidoptera: Noctuidae) Immature Stages Under Controlled and Standardized Conditions. J. Agric. Sci. 2019, 11, 76. [Google Scholar] [CrossRef]
- Guo, J.-M.; Liu, X.-L.; Liu, S.-R.; Wei, Z.-Q.; Han, W.-K.; Guo, Y.; Dong, S.-L. Functional Characterization of Sex Pheromone Receptors in the Fall Armyworm (Spodoptera frugiperda). Insects 2020, 11, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Y.Y.; Liu, J.; Xie, M.C.; Li, Y.H.; Yang, J.J.; Zhang, M.L.; Qiu, K. Observation on law of diffusion damage of Spodoptera frugiperda in China in 2019. Int. J. Plant Prot. 2019, 45, 10–19. [Google Scholar]
- Huang, Y.; Dong, Y.; Huang, W.; Ren, B.; Deng, Q.; Shi, Y.; Bai, J.; Ren, Y.; Geng, Y.; Ma, H. Overwintering Distribution of Fall Armyworm (Spodoptera frugiperda) in Yunnan, China, and Influencing Environmental Factors. Insects 2020, 11, 805. [Google Scholar] [CrossRef]
- Thiem, B.; Wesołowska, M.; Skrzypczak, L.; Budzianowski, J. Phenolic compounds in two Solidago L. species from in vitro culture. Acta Pol. Pharm. Drug Res. 2001, 58, 277–281. [Google Scholar]
- Starks, C.M.; Williams, R.; Goering, M.G.; O’Neil-Johnson, M.; Norman, V.L.; Hu, J.-F.; Garo, E.; Hough, G.W.; Rice, S.M.; Eldridge, G.R. Antibacterial clerodane diterpenes from Goldenrod (Solidago virgaurea). Phytochemistry 2010, 71, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Schmeda-Hirschmann, G.; Rodriguez, J.; Astudillo, L. Gastroprotective activity of the diterpene solidagenone and its de-rivatives on experimentally induced gastric lesions in mice. J. Ethnopharmacol. 2002, 1, 111–115. [Google Scholar] [CrossRef]
- Jaisinghani, R.N. Antibacterial properties of quercetin. Microbiol. Res. 2017, 8, 1. [Google Scholar] [CrossRef] [Green Version]
- Erdogan-Orhan, I.; Altun, M.L.; Yilmaz, B.S.; Saltan, H.G. Anti-Acetylcholinesterase and Antioxidant Assets of the Major Components (Salicin, Amentoflavone, and Chlorogenic Acid) and the Extracts of Viburnum opulus and Viburnum lantana and Their Total Phenol and Flavonoid Contents. J. Med. Food 2011, 14, 434–440. [Google Scholar] [CrossRef]
- Kwon, S.-H.; Lee, H.-K.; Kim, J.-A.; Hong, S.-I.; Kim, H.-C.; Jo, T.-H.; Park, Y.-I.; Lee, C.-K.; Kim, Y.-B.; Lee, S.-Y.; et al. Neuroprotective effects of chlorogenic acid on scopolamine-induced amnesia via anti-acetylcholinesterase and anti-oxidative activities in mice. Eur. J. Pharmacol. 2010, 649, 210–217. [Google Scholar] [CrossRef]
- Xu, D.; Hu, M.-J.; Wang, Y.-Q.; Cui, Y.-L. Antioxidant Activities of Quercetin and Its Complexes for Medicinal Application. Molecules 2019, 24, 1123. [Google Scholar] [CrossRef] [Green Version]
- Hay, W.T.; Behle, R.W.; Berhow, M.A.; Miller, A.C.; Selling, G.W. Biopesticide synergy when combining plant flavonoids and entomopathogenic baculovirus. Sci. Rep. 2020, 10, 6806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olennikov, D.N.; Kashchenko, N.I.; Chirikova, N.K.; Akobirshoeva, A.; Zilfikarov, I.N.; Vennos, C. Isorhamnetin and Quercetin Derivatives as Anti-Acetylcholinesterase Principles of Marigold (Calendula officinalis) Flowers and Preparations. Int. J. Mol. Sci. 2017, 18, 1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Sousa, G.F.; de Aguilar, M.G.; Takahashi, J.A.; Alves, T.M.; Kohlhoff, M.; Filho, S.A.V.; Silva, G.D.; Duarte, L.P. Flavonol triglycosides of leaves from Maytenus robusta with acetylcholinesterase inhibition. Phytochem. Lett. 2017, 19, 34–38. [Google Scholar] [CrossRef]
- Kelly, A.M.; De Oliveira, T.B.; Valverde, S.S. Determination of the metabolic profile of Solidago canadensis using UFLC-PDA-ESI-TOF. Rodriguésia 2020, 71, 1–6. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as Important Molecules of Plant Interactions with the Environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef]
- Dey, A.; Bhattacharya, R.; Mukherjee, A.; Pandey, D.K. Natural products against Alzheimer’s disease: Pharma-co-therapeutics and biotechnological interventions. Biotechnol. Adv. 2017, 2, 178–216. [Google Scholar] [CrossRef]
- Riddick, E. Potential of Quercetin to Reduce Herbivory without Disrupting Natural Enemies and Pollinators. Agriculture 2021, 11, 476. [Google Scholar] [CrossRef]
- Mesbah, H.A.; Saad, A.S.A.; Mourad, A.K.; Taman, F.A.; Mohamed, I.B. Joint action of quercetin with four insecticides on the cotton leaf-worm larvae, Spodoptera littoralis Boisd. (Lep.: Noctuidae) in Egypt. Commun. Agric. Appl. Biol. Sci. 2007, 72, 445–457. [Google Scholar]
- Ahmad, G.; Rasool, N.; Rizwan, K.; Imran, I.; Fawad Zahoor, A.; Zubair, M.; Rashid, U. Synthesis, In-Vitro Cholines-terase Inhibition, In-Vivo Anticonvulsant Activity and In-Silico Exploration of N-(4-Methylpyridin-2-yl) thio-phene-2-carboxamide Analogs. Bioorg. Chem. 2019, 92, 103216. [Google Scholar] [CrossRef]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Klein, B.J.; Vann, K.R.; Andrews, F.H.; Wang, W.W.; Zhang, J.; Zhang, Y.; Beloglazkina, A.A.; Mi, W.; Li, Y.; Li, H.; et al. Structural insights into the π-π-π stacking mechanism and DNA-binding ac-tivity of the YEATS domain. Nat. Commun. 2018, 9, 4574. [Google Scholar] [CrossRef] [PubMed]
- Akher, F.B.; Ebrahimi, A.; Mostafavi, N. Characterization of π-stacking interactions between aromatic amino acids and quercetagetin. J. Mol. Struct. 2017, 1128, 13–20. [Google Scholar] [CrossRef]
- Urbatsch, L.E.; Baldwin, B.G.; Donoghue, M.J.; Donoghue, B.G.B.J. Phylogeny of the Coneflowers and Relatives (Heliantheae: Asteraceae) Based on Nuclear rDNA Internal Transcribed Spacer (ITS) Sequences and Chlorplast DNA Restriction Site Data. Syst. Bot. 2000, 25, 539. [Google Scholar] [CrossRef]
- Egar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acid Res. 2004, 5, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera? A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. Autodock4 and Auto-DockTools4: Automated docking with selective receptor flexiblity. J. Comput. Chem. 2009, 16, 2785–2791. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solvent | Yield (%) | mg per g |
|---|---|---|
| Ethanol | 33.39 | 333 |
| Dichloromethane | 18.34 | 183 |
| Hexane | 5.03 | 50 |
| Extract | Compound | Molecular Formula | Theoretical m/z | Experimental m/z | Reference |
|---|---|---|---|---|---|
| EtOH | Solidagoic acid G | C21H30O5 | 361.19 | 361.13 | [15] |
| Unknown | -- | -- | 542.23 | -- | |
| Quercetin | C15H10O7 | 302.23 | 303.28 | [2] [6] | |
| Solidagoic acid C | C20H28O4 | 331.0 | 331.10 | [15] | |
| Unknown | -- | -- | 104.02 | -- | |
| DCM | Unknown | -- | -- | 377.11 | -- |
| Quercetin | C15H10O7 | 302.23 | 303.28 | [2] [6] | |
| Unknown | -- | -- | 379.25 | -- | |
| Solidagoic acid B | C25H34O5 | 414.5 | 415.18 | [15] | |
| Rosmarinic acid | C18H16O8 | 360.3 | 360.41 | [2] | |
| Chlorogenic acid | C16H18O9 | 355.00 | 356.45 | [2] | |
| Hex | Solidagoic acid B | C25H34O5 | 414.5 | 415.24 | [15] [16] |
| Unknown | -- | -- | 102.04 | -- | |
| Hyperoside | C21H20O12 | 464.4 | 465.27 | [2] [6] | |
| Quercetin | C15H10O7 | 302.23 | 303.21 | [2] [6] | |
| Unknown | -- | -- | 407.28 | -- | |
| Unknown | -- | -- | 389.25 | -- | |
| Chlorogenic acid | C16H18O9 | 355.00 | 356.23 | [2] |
| Compounds | CL50 (mg/mL) | Mixture of Compounds | CL50 (mg/mL) |
|---|---|---|---|
| Quercetin | 0.157 | Quercetin: Chlorogenic acid 1:1 | 0.729 |
| Chlorogenic acid | No insecticidal activity | Quercetin: Chlorogenic acid 1:9 | No insecticidal activity |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera-Mayorga, V.; Guerrero-Sánchez, J.A.; Méndez-Álvarez, D.; Paredes-Sánchez, F.A.; Rodríguez-Duran, L.V.; Niño-García, N.; Paz-González, A.D.; Rivera, G. Insecticidal Activity of Organic Extracts of Solidago graminifolia and Its Main Metabolites (Quercetin and Chlorogenic Acid) against Spodoptera frugiperda: An In Vitro and In Silico Approach. Molecules 2022, 27, 3325. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103325
Herrera-Mayorga V, Guerrero-Sánchez JA, Méndez-Álvarez D, Paredes-Sánchez FA, Rodríguez-Duran LV, Niño-García N, Paz-González AD, Rivera G. Insecticidal Activity of Organic Extracts of Solidago graminifolia and Its Main Metabolites (Quercetin and Chlorogenic Acid) against Spodoptera frugiperda: An In Vitro and In Silico Approach. Molecules. 2022; 27(10):3325. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103325
Chicago/Turabian StyleHerrera-Mayorga, Verónica, José Alfredo Guerrero-Sánchez, Domingo Méndez-Álvarez, Francisco A. Paredes-Sánchez, Luis Víctor Rodríguez-Duran, Nohemí Niño-García, Alma D. Paz-González, and Gildardo Rivera. 2022. "Insecticidal Activity of Organic Extracts of Solidago graminifolia and Its Main Metabolites (Quercetin and Chlorogenic Acid) against Spodoptera frugiperda: An In Vitro and In Silico Approach" Molecules 27, no. 10: 3325. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27103325