
Notoginsenoside R1 Promotes Migration, Adhesin, Spreading, and Osteogenic Differentiation of Human Adipose Tissue-Derived Mesenchymal Stromal Cells


 , and
, and
Abstract
:1. Introduction
2. Results
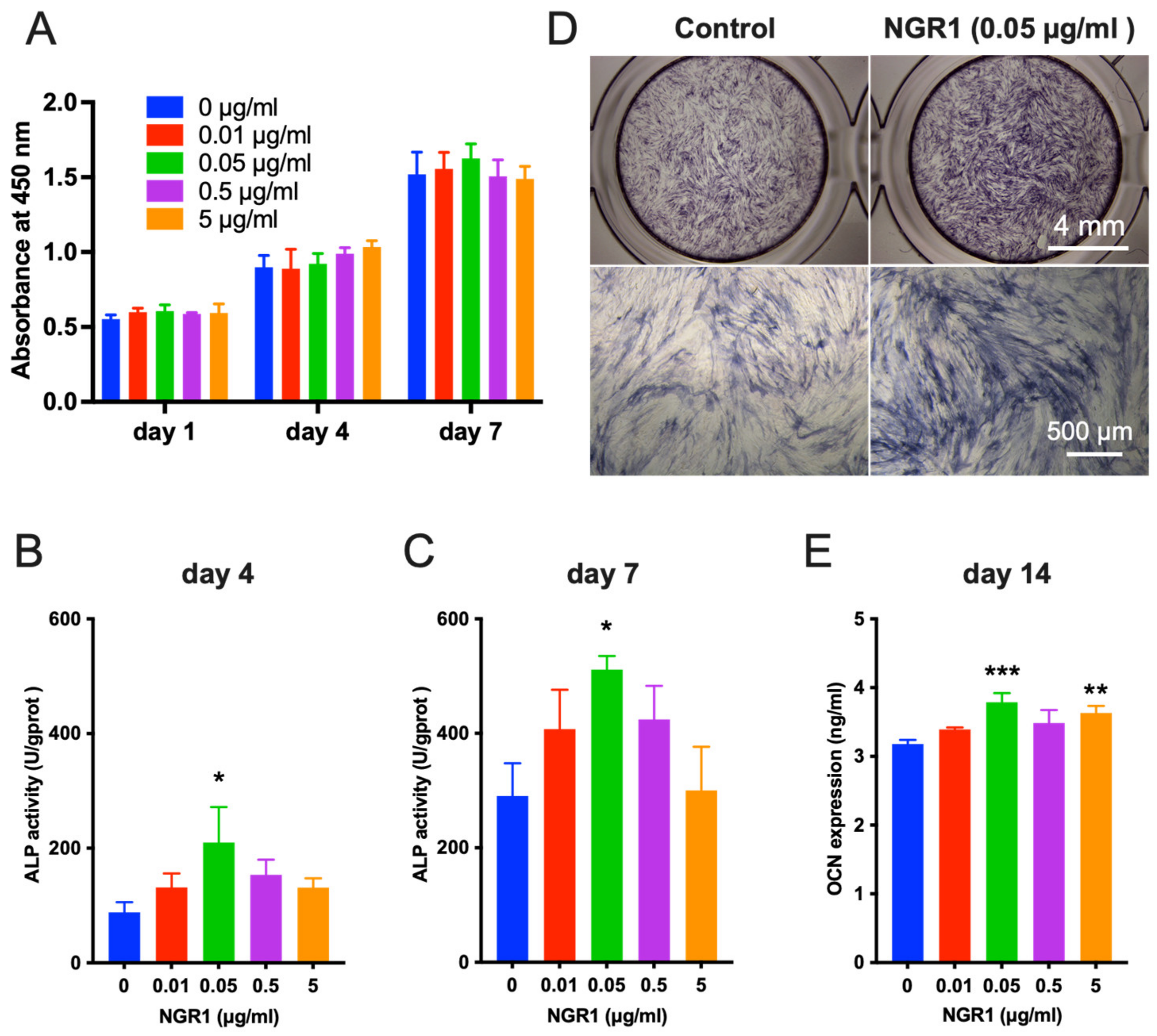
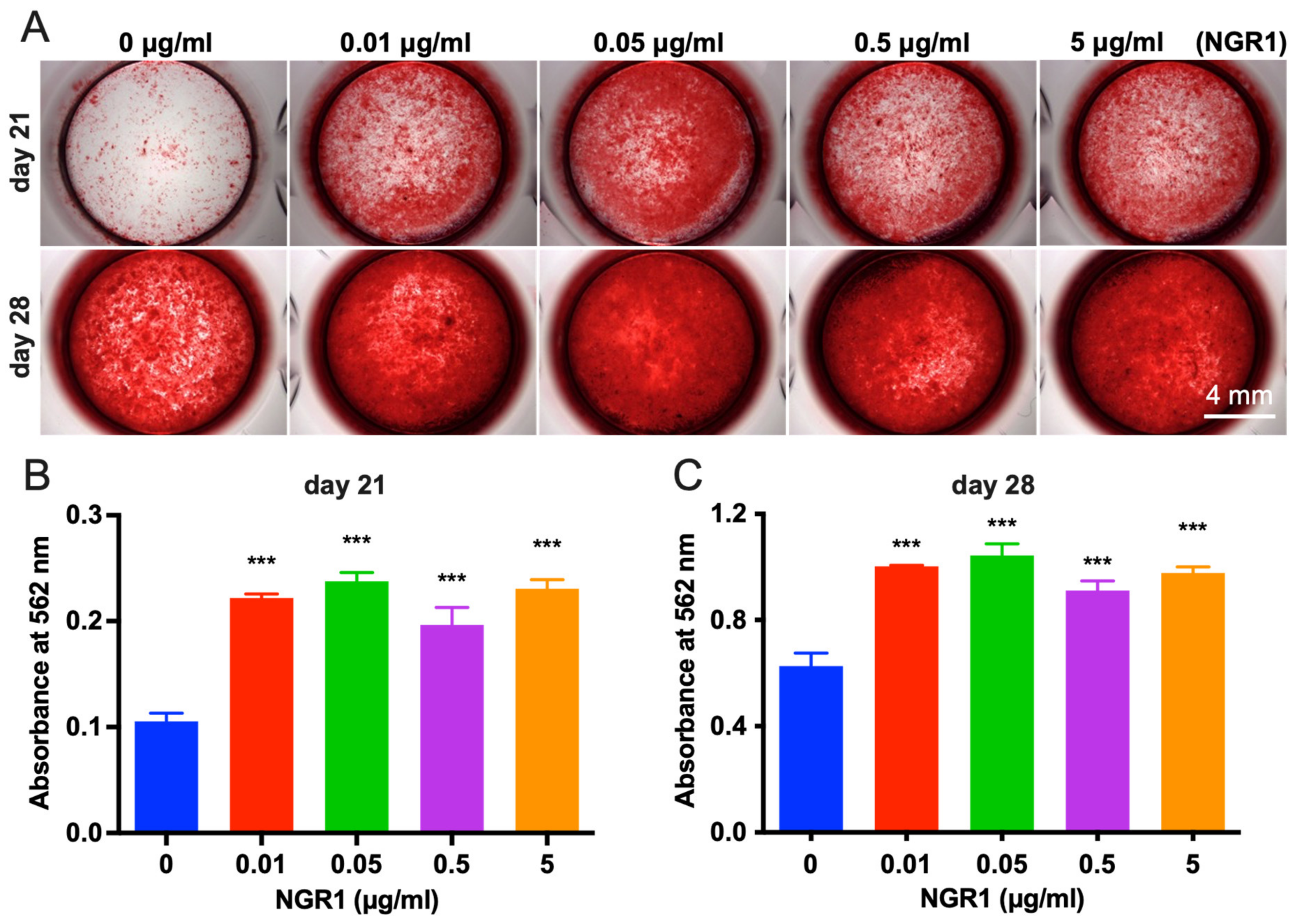
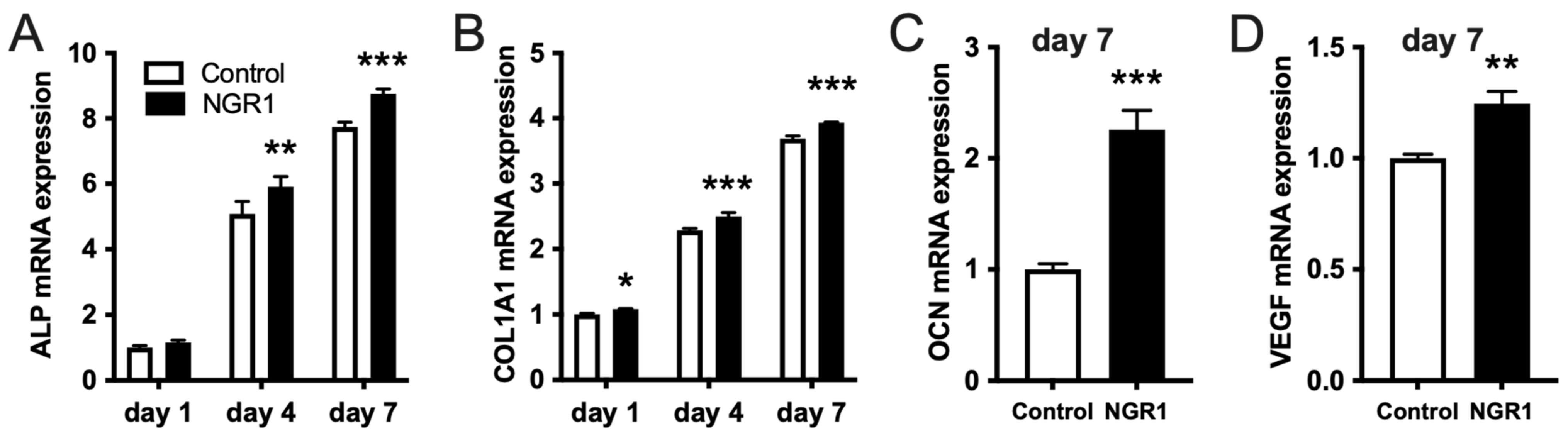
2.1. Biocompatible NGR1 Enhances Osteogenic Differentiation of hASCs
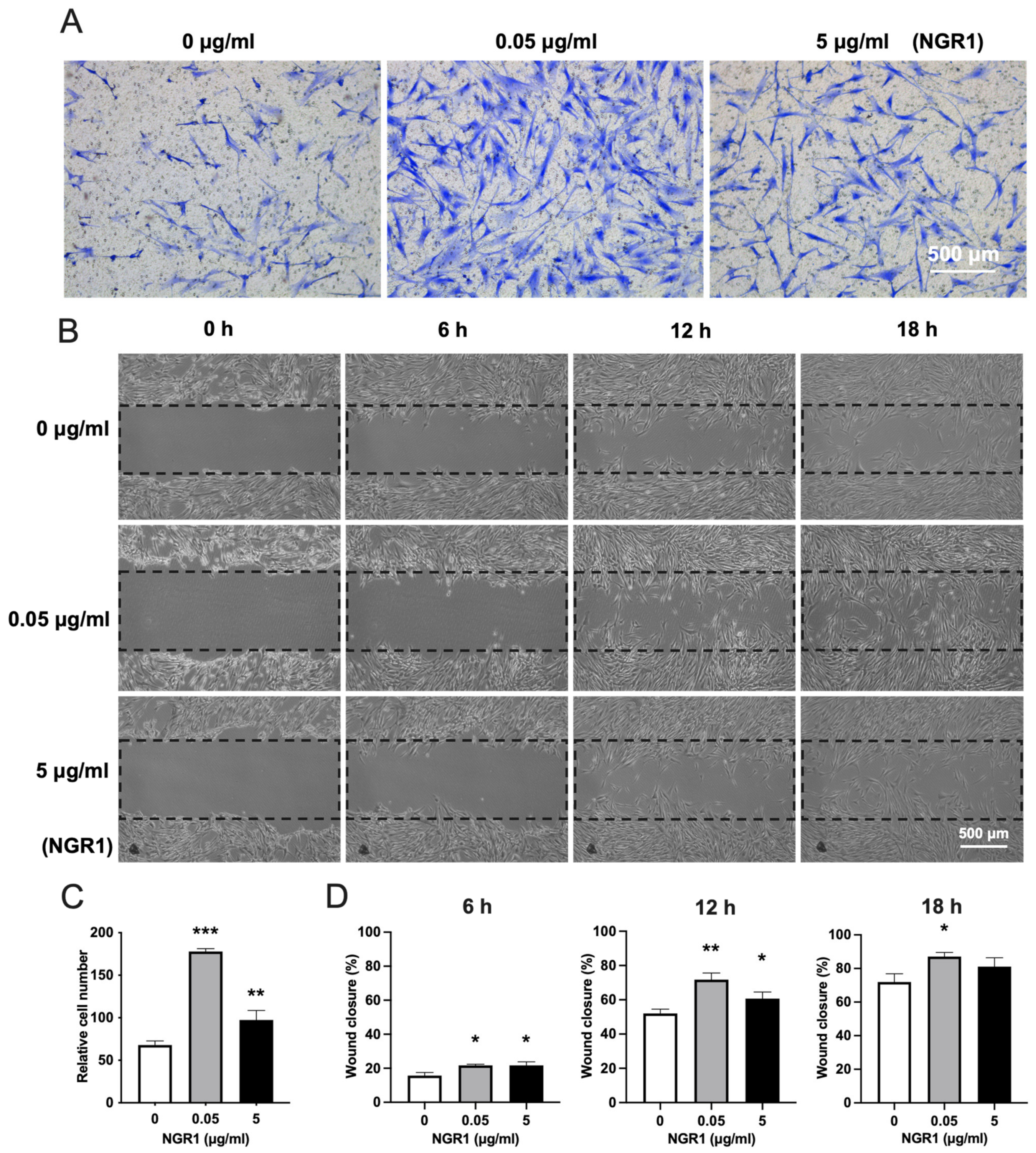
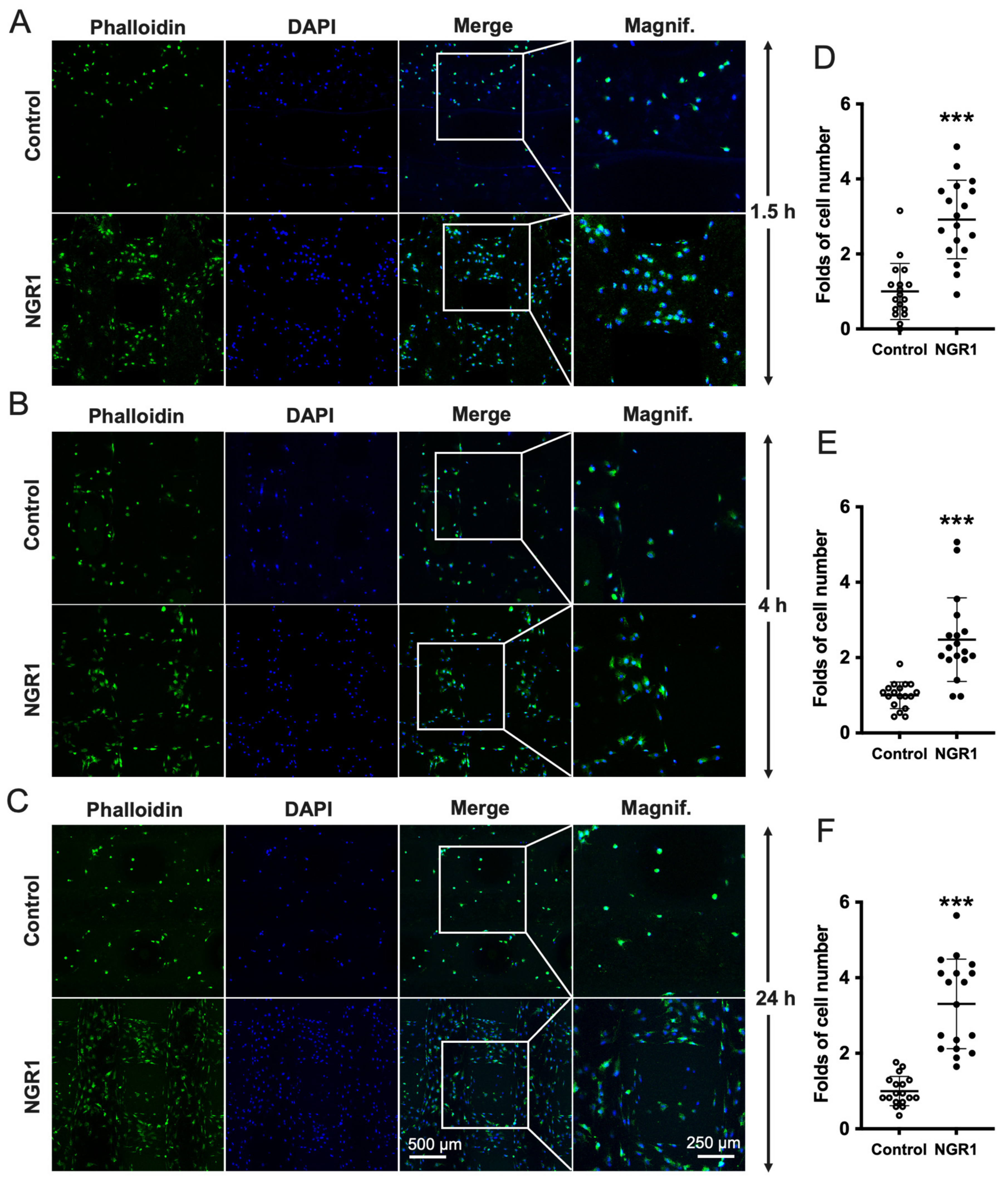
2.2. NGR1 Increases the Adhesion, Spreading, and Migration of hASCs
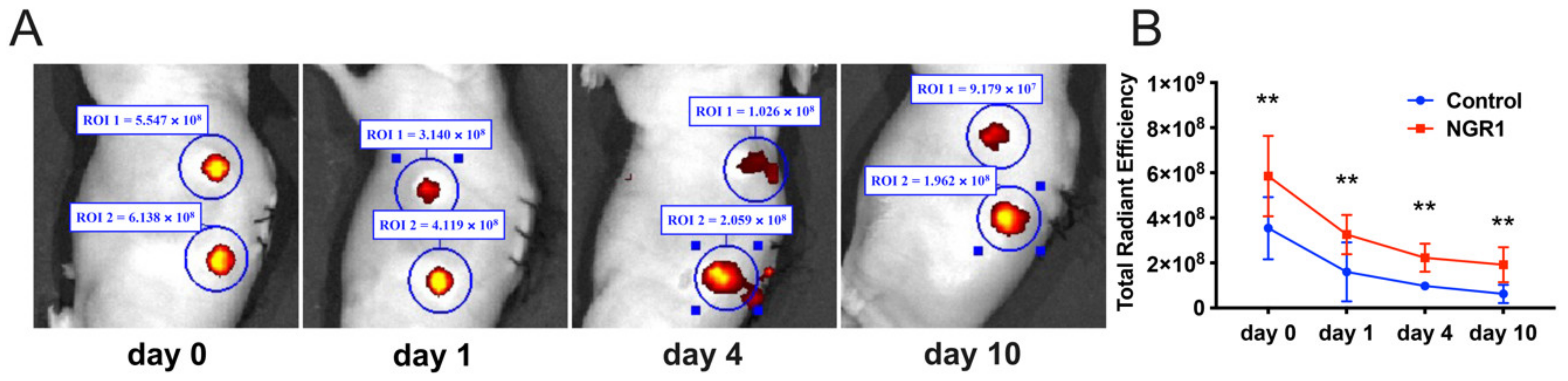
2.3. NGR1 Promotes hASCs Adhesion and Survival in 3D Printed TCP Scaffolds In Vivo
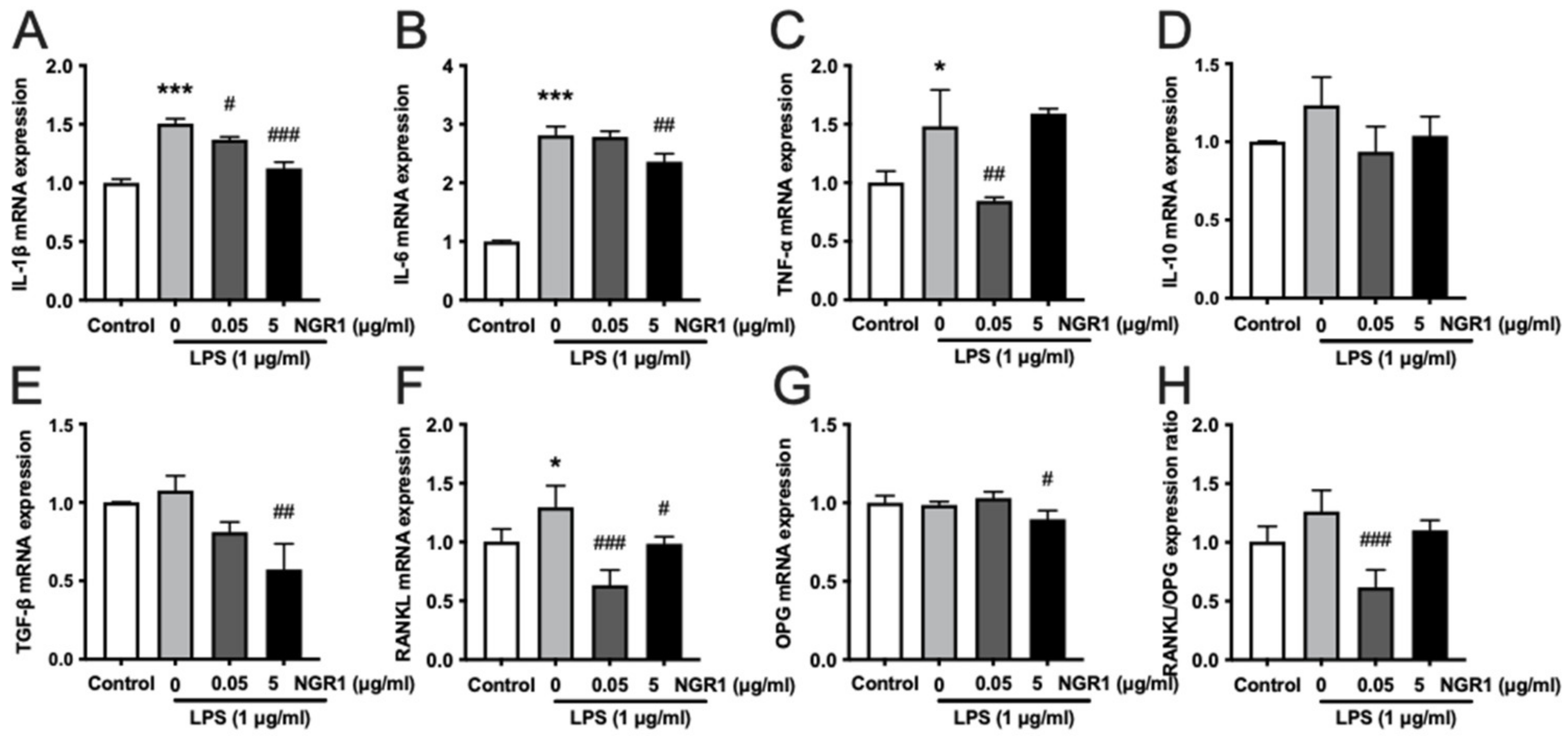
2.4. Anti-Inflammatory Effects of NGR1 in hASCs
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Chemicals
4.2. Cell Viability Assay
4.3. Alkaline Phosphatase (ALP) Staining and Activity Assay
4.4. Osteocalcin (OCN) ELISA
4.5. Alizarin Red Staining
4.6. The induction of the Inflammatory Microenvironment in hASCs
4.7. RT-qPCR Analysis
4.8. Cell Migration Assay
4.9. Analysis of hASCs Adhesion and Spreading
4.10. Animal Study
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Roseti, L.; Parisi, V.; Petretta, M.; Cavallo, C.; Desando, G.; Bartolotti, I.; Grigolo, B. Scaffolds for Bone Tissue Engineering: State of the art and new perspectives. Mater. Sci. Eng. C Mater. Biol. Appl. 2017, 78, 1246–1262. [Google Scholar] [CrossRef] [PubMed]
- Qasim, M.; Chae, D.S.; Lee, N.Y. Advancements and frontiers in nano-based 3D and 4D scaffolds for bone and cartilage tissue engineering. Int. J. Nanomed. 2019, 14, 4333–4351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedrero, S.G.; Llamas-Sillero, P.; Serrano-Lopez, J. A Multidisciplinary Journey towards Bone Tissue Engineering. Materials 2021, 14, 4896. [Google Scholar] [CrossRef]
- Clynes, M.A.; Harvey, N.C.; Curtis, E.M.; Fuggle, N.R.; Dennison, E.M.; Cooper, C. The epidemiology of osteoporosis. Br. Med. Bull. 2020, 133, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Fernandez de Grado, G.; Keller, L.; Idoux-Gillet, Y.; Wagner, Q.; Musset, A.M.; Benkirane-Jessel, N.; Bornert, F.; Offner, D. Bone substitutes: A review of their characteristics, clinical use, and perspectives for large bone defects management. J. Tissue Eng. 2018, 9, 2041731418776819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roddy, E.; DeBaun, M.R.; Daoud-Gray, A.; Yang, Y.P.; Gardner, M.J. Treatment of critical-sized bone defects: Clinical and tissue engineering perspectives. Eur. J. Orthop. Surg. Traumatol. 2018, 28, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Ansari, M. Bone tissue regeneration: Biology, strategies and interface studies. Prog. Biomater. 2019, 8, 223–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Witte, T.M.; Fratila-Apachitei, L.E.; Zadpoor, A.A.; Peppas, N.A. Bone tissue engineering via growth factor delivery: From scaffolds to complex matrices. Regen. Biomater. 2018, 5, 197–211. [Google Scholar] [CrossRef] [Green Version]
- Perić Kačarević, Ž.; Rider, P.; Alkildani, S.; Retnasingh, S.; Pejakic, M.; Schnettler, R.; Gosau, M.; Smeets, R.; Jung, O.; Barbeck, M. An introduction to bone tissue engineering. Int. J. Artif. Organs 2020, 43, 69–86. [Google Scholar] [CrossRef]
- Hinsenkamp, M.; Collard, J.F. Growth factors in orthopaedic surgery: Demineralized bone matrix versus recombinant bone morphogenetic proteins. Int. Orthop. 2015, 39, 137–147. [Google Scholar] [CrossRef] [Green Version]
- Shekaran, A.; Garcia, J.R.; Clark, A.Y.; Kavanaugh, T.E.; Lin, A.S.; Guldberg, R.E.; Garcia, A.J. Bone regeneration using an alpha 2 beta 1 integrin-specific hydrogel as a BMP-2 delivery vehicle. Biomaterials 2014, 35, 5453–5461. [Google Scholar] [CrossRef] [Green Version]
- Zara, J.N.; Siu, R.K.; Zhang, X.; Shen, J.; Ngo, R.; Lee, M.; Li, W.; Chiang, M.; Chung, J.; Kwak, J.; et al. High doses of bone morphogenetic protein 2 induce structurally abnormal bone and inflammation in vivo. Tissue Eng. Part A 2011, 17, 1389–1399. [Google Scholar] [CrossRef] [Green Version]
- Ho-Shui-Ling, A.; Bolander, J.; Rustom, L.E.; Johnson, A.W.; Luyten, F.P.; Picart, C. Bone regeneration strategies: Engineered scaffolds, bioactive molecules and stem cells current stage and future perspectives. Biomaterials 2018, 180, 143–162. [Google Scholar] [CrossRef]
- Singh, R.K.; Kumar, S.; Tomar, M.S.; Verma, P.K.; Kumar, A.; Kumar, S.; Kumar, N.; Singh, J.P.; Acharya, A. Putative role of natural products as Protein Kinase C modulator in different disease conditions. DARU 2021, 29, 397–414. [Google Scholar] [CrossRef]
- Catalogna, G.; Moraca, F.; D’Antona, L.; Dattilo, V.; Perrotti, G.; Lupia, A.; Costa, G.; Ortuso, F.; Iuliano, R.; Trapasso, F.; et al. Review about the multi-target profile of resveratrol and its implication in the SGK1 inhibition. Eur. J. Med. Chem. 2019, 183, 111675. [Google Scholar] [CrossRef]
- Wang, Z.L.; Wang, S.; Kuang, Y.; Hu, Z.M.; Qiao, X.; Ye, M. A comprehensive review on phytochemistry, pharmacology, and flavonoid biosynthesis of Scutellaria baicalensis. Pharm. Biol. 2018, 56, 465–484. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.B.; Ou, J.Y.; Cao, H.H.; Liu, J.S.; Yu, L.Z. Heat-Clearing Chinese Medicines in Lipopolysaccharide-Induced Inflammation. Chin. J. Integr. Med. 2020, 26, 552–559. [Google Scholar] [CrossRef]
- Sun, L.J.; Li, C.; Wen, X.H.; Guo, L.; Guo, Z.F.; Liao, L.Q.; Guo, Y. Icariin Stimulates hFOB 1.19 Osteoblast Proliferation and Differentiation via OPG/RANKL Mediated by the Estrogen Receptor. Curr. Pharm. Biotechnol. 2021, 22, 168–175. [Google Scholar] [CrossRef]
- Murgia, D.; Mauceri, R.; Campisi, G.; De Caro, V. Advance on Resveratrol Application in Bone Regeneration: Progress and Perspectives for Use in Oral and Maxillofacial Surgery. Biomolecules 2019, 9, 94. [Google Scholar] [CrossRef] [Green Version]
- Aravamudhan, A.; Ramos, D.M.; Nip, J.; Subramanian, A.; James, R.; Harmon, M.D.; Yu, X.; Kumbar, S.G. Osteoinductive small molecules: Growth factor alternatives for bone tissue engineering. Curr. Pharm. Des. 2013, 19, 3420–3428. [Google Scholar] [CrossRef]
- Xie, W.; Meng, X.; Zhai, Y.; Zhou, P.; Ye, T.; Wang, Z.; Sun, G.; Sun, X. Panax Notoginseng Saponins: A Review of Its Mechanisms of Antidepressant or Anxiolytic Effects and Network Analysis on Phytochemistry and Pharmacology. Molecules 2018, 23, 940. [Google Scholar] [CrossRef] [Green Version]
- Ji, Z.; Cheng, Y.; Yuan, P.; Dang, X.; Guo, X.; Wang, W. Panax notoginseng stimulates alkaline phosphatase activity, collagen synthesis, and mineralization in osteoblastic MC3T3-E1 cells. In Vitro Cell. Dev. Biol. Anim. 2015, 51, 950–957. [Google Scholar] [CrossRef]
- Liu, H.; Yang, J.; Yang, W.; Hu, S.; Wu, Y.; Zhao, B.; Hu, H.; Du, S. Focus on Notoginsenoside R1 in Metabolism and Prevention Against Human Diseases. Drug Des. Dev. Ther. 2020, 14, 551–565. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Yang, S.; Luo, Y.; Zhang, C.; Rao, Y.; Liu, R.; Feng, Y.; Yu, J. Notoginsenoside R1 inhibits vascular smooth muscle cell proliferation, migration and neointimal hyperplasia through PI3K/Akt signaling. Sci. Rep. 2018, 8, 7595. [Google Scholar] [CrossRef] [Green Version]
- Ge, Z.R.; Xu, M.C.; Huang, Y.U.; Zhang, C.J.; Lin, J.E.; Ruan, C.W. Cardioprotective effect of notoginsenoside R1 in a rabbit lung remote ischemic postconditioning model via activation of the TGF-beta1/TAK1 signaling pathway. Exp. Ther. Med. 2016, 11, 2341–2348. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Liu, B.; Zheng, X.; Wang, D. Notoginsenoside R1 alleviates lipopolysaccharide-triggered PC-12 inflammatory damage via elevating microRNA-132. Artif. Cells Nanomed. Biotechnol. 2019, 47, 1808–1814. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Xiao, J.; Li, X.; Chen, H.; Kang, D.; Shao, Y.; Shen, B.; Zhu, Z.; Yin, X.; Xie, L.; et al. Low Cerebral Exposure Cannot Hinder the Neuroprotective Effects of Panax Notoginsenosides. Drug Metab. Dispos. 2018, 46, 53–65. [Google Scholar] [CrossRef]
- Li, Y.; Li, Z.; Jia, Y.; Ding, B.; Yu, J. In Vitro Anti-hepatoma Activities of Notoginsenoside R1 Through Downregulation of Tumor Promoter miR-21. Dig. Dis. Sci. 2020, 65, 1364–1375. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Z.; Anderson, S.; Yuan, C.S. Phytochemistry and Anticancer Potential of Notoginseng. Am. J. Chin. Med. 2016, 44, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lin, Z.; Guo, J.; Xu, G.; Li, Y.; Xu, T.; Lv, H.; Chen, J.; Wu, G. Notoginsenoside R1 significantly promotes in vitro osteoblastogenesis. Int. J. Mol. Med. 2016, 38, 537–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Sun, H.; Zhong, Y. Notoginsenoside R1 promotes MC3T3-E1 differentiation by up-regulating miR-23a via MAPK and JAK1/STAT3 pathways. Artif. Cells Nanomed. Biotechnol. 2019, 47, 603–609. [Google Scholar] [CrossRef] [Green Version]
- Arthur, A.; Gronthos, S. Clinical Application of Bone Marrow Mesenchymal Stem/Stromal Cells to Repair Skeletal Tissue. Int. J. Mol. Sci. 2020, 21, 9759. [Google Scholar] [CrossRef]
- Toosi, S.; Behravan, N.; Behravan, J. Nonunion fractures, mesenchymal stem cells and bone tissue engineering. J. Biomed. Mater. Res. Part A 2018, 106, 2552–2562. [Google Scholar] [CrossRef]
- Lin, H.; Sohn, J.; Shen, H.; Langhans, M.T.; Tuan, R.S. Bone marrow mesenchymal stem cells: Aging and tissue engineering applications to enhance bone healing. Biomaterials 2019, 203, 96–110. [Google Scholar] [CrossRef]
- Fu, X.R.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, G.B. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8, 784. [Google Scholar] [CrossRef] [Green Version]
- Airuddin, S.S.; Halim, A.S.; Wan Sulaiman, W.A.; Kadir, R.; Nasir, N.A.M. Adipose-Derived Stem Cell: “Treat or Trick”. Biomedicines 2021, 9, 1624. [Google Scholar] [CrossRef]
- Camara, D.A.D.; Shibli, J.A.; Muller, E.A.; De-Sa-Junior, P.L.; Porcacchia, A.S.; Blay, A.; Lizier, N.F. Adipose Tissue-Derived Stem Cells: The Biologic Basis and Future Directions for Tissue Engineering. Materials 2020, 13, 3210. [Google Scholar] [CrossRef]
- Shafaei, H.; Kalarestaghi, H. Adipose-derived stem cells: An appropriate selection for osteogenic differentiation. J. Cell. Physiol. 2020, 235, 8371–8386. [Google Scholar] [CrossRef]
- Klar, A.S.; Zimoch, J.; Biedermann, T. Skin Tissue Engineering: Application of Adipose-Derived Stem Cells. Biomed. Res. Int. 2017, 2017, 9747010. [Google Scholar] [CrossRef] [Green Version]
- Murphy, C.M.; O’Brien, F.J.; Little, D.G.; Schindeler, A. Cell-scaffold interactions in the bone tissue engineering triad. Eur. Cells Mater. 2013, 26, 120–132. [Google Scholar] [CrossRef]
- Newman, H.; Shih, Y.V.; Varghese, S. Resolution of inflammation in bone regeneration: From understandings to therapeutic applications. Biomaterials 2021, 277, 121114. [Google Scholar] [CrossRef]
- Maruyama, M.; Rhee, C.; Utsunomiya, T.; Zhang, N.; Ueno, M.; Yao, Z.; Goodman, S.B. Modulation of the Inflammatory Response and Bone Healing. Front. Endocrinol. 2020, 11, 386. [Google Scholar] [CrossRef]
- Noronha, N.C.; Mizukami, A.; Caliari-Oliveira, C.; Cominal, J.G.; Rocha, J.L.M.; Covas, D.T.; Swiech, K.; Malmegrim, K.C.R. Priming approaches to improve the efficacy of mesenchymal stromal cell-based therapies. Stem Cell Res. Ther. 2019, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.; Li, Q. Notoginsenoside R1 promotes differentiation of human alveolar osteoblasts in inflammatory microenvironment through inhibiting NFkappaB pathway and activating Wnt/betacatenin pathway. Mol. Med. Rep. 2020, 22, 4754–4762. [Google Scholar] [CrossRef]
- Wang, T.; Wan, D.; Shao, L.; Dai, J.; Jiang, C. Notoginsenoside R1 stimulates osteogenic function in primary osteoblasts via estrogen receptor signaling. Biochem. Biophys. Res. Commun. 2015, 466, 232–239. [Google Scholar] [CrossRef]
- Ritz, U.; Nusselt, T.; Sewing, A.; Ziebart, T.; Kaufmann, K.; Baranowski, A.; Rommens, P.M.; Hofmann, A. The effect of different collagen modifications for titanium and titanium nitrite surfaces on functions of gingival fibroblasts. Clin. Oral Investig. 2017, 21, 255–265. [Google Scholar] [CrossRef]
- Van Dijk, I.A.; Beker, A.F.; Jellema, W.; Nazmi, K.; Wu, G.; Wismeijer, D.; Krawczyk, P.M.; Bolscher, J.G.; Veerman, E.C.; Stap, J. Histatin 1 Enhances Cell Adhesion to Titanium in an Implant Integration Model. J. Dent. Res. 2017, 96, 430–436. [Google Scholar] [CrossRef]
- Sun, W.; Shi, A.; Ma, D.; Bolscher, J.G.M.; Nazmi, K.; Veerman, E.C.I.; Bikker, F.J.; Lin, H.; Wu, G. All-trans retinoic acid and human salivary histatin-1 promote the spreading and osteogenic activities of pre-osteoblasts in vitro. FEBS Open Bio 2020, 10, 396–406. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.C.; Guo, Y.L.; Liu, R.H.; Wu, S.Y.; Fang, J.H.; Huang, B.X.; Li, Z.P.; Chen, Z.F.; Chen, Z.T. Tuning surface properties of bone biomaterials to manipulate osteoblastic cell adhesion and the signaling pathways for the enhancement of early osseointegration. Colloids Surf. B Biointerfaces 2018, 164, 58–69. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Chen, H.; Zhao, X.; Chen, Z.; Zhang, P.; Tian, Y.; Wang, Y.; Ding, T.; Wang, L.; Shen, Y. Advances in mesenchymal stem cell conditioned medium-mediated periodontal tissue regeneration. J. Transl. Med. 2021, 19, 456. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Wang, X.; Xu, C.; Asakura, A.; Yoshiyama, M.; From, A.H.; Zhang, J. Xenotransplantation of long-term-cultured swine bone marrow-derived mesenchymal stem cells. Stem Cells 2007, 25, 612–620. [Google Scholar] [CrossRef] [Green Version]
- Mishra, A.K.; Campanale, J.P.; Mondo, J.A.; Montell, D.J. Cell interactions in collective cell migration. Development 2019, 146, dev172056. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yeung, K.W.K. Bone grafts and biomaterials substitutes for bone defect repair: A review. Bioact. Mater. 2017, 2, 224–247. [Google Scholar] [CrossRef]
- Yang, W.; Wang, L.; Mettenbrink, E.M.; DeAngelis, P.L.; Wilhelm, S. Nanoparticle Toxicology. Annu. Rev. Pharmacol. Toxicol. 2021, 61, 269–289. [Google Scholar] [CrossRef]
- Wu, Y.; Cao, L.; Xia, L.; Wu, Q.; Wang, J.; Wang, X.; Xu, L.; Zhou, Y.; Xu, Y.; Jiang, X. Evaluation of Osteogenesis and Angiogenesis of Icariin in Local Controlled Release and Systemic Delivery for Calvarial Defect in Ovariectomized Rats. Sci. Rep. 2017, 7, 5077. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Guo, Y.; Li, D.X.; Wang, R.; Fan, H.S.; Xiao, Y.M.; Zhang, L.; Zhang, X.D. The effect of loading icariin on biocompatibility and bioactivity of porous beta-TCP ceramic. J. Mater. Sci. Mater. Med. 2011, 22, 371–379. [Google Scholar] [CrossRef]
- Lai, Y.; Cao, H.; Wang, X.; Chen, S.; Zhang, M.; Wang, N.; Yao, Z.; Dai, Y.; Xie, X.; Zhang, P.; et al. Porous composite scaffold incorporating osteogenic phytomolecule icariin for promoting skeletal regeneration in challenging osteonecrotic bone in rabbits. Biomaterials 2018, 153, 1–13. [Google Scholar] [CrossRef]
- Ge, Y.W.; Lu, J.W.; Sun, Z.Y.; Liu, Z.Q.; Zhou, J.; Ke, Q.F.; Mao, Y.Q.; Guo, Y.P.; Zhu, Z.A. Ursolic acid loaded-mesoporous bioglass/chitosan porous scaffolds as drug delivery system for bone regeneration. Nanomed. Nanotechnol. Biol. Med. 2019, 18, 336–346. [Google Scholar] [CrossRef]
- Yu, X.J.; Wang, Y.X.; Liu, X.L.; Ge, Y.W.; Zhang, S.Y. Ursolic Acid Loaded-Mesoporous Hydroxylapatite/Chitosan Therapeutic Scaffolds Regulate Bone Regeneration Ability by Promoting the M2-Type Polarization of Macrophages. Int. J. Nanomed. 2021, 16, 5301–5315. [Google Scholar] [CrossRef]
- Garcia, C.F.; Marangon, C.A.; Massimino, L.C.; Klingbeil, M.F.G.; Martins, V.C.A.; Plepis, A.M.G. Development of collagen/nanohydroxyapatite scaffolds containing plant extract intended for bone regeneration. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 123, 111955. [Google Scholar] [CrossRef]
- Thongtham, N.; Chai-in, P.; Unger, O.; Boonrungsiman, S.; Suwantong, O. Fabrication of chitosan/collagen/hydroxyapatite scaffolds with encapsulated Cissus quadrangularis extract. Polym. Adv. Technol. 2020, 31, 1496–1507. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Sugimoto, K.; Kamio, H.; Okabe, K.; Kuroda, K.; Okido, M.; Hibi, H. Kaempferol-immobilized titanium dioxide promotes formation of new bone: Effects of loading methods on bone marrow stromal cell differentiation in vivo and in vitro. Int. J. Nanomed. 2018, 13, 1665–1676. [Google Scholar] [CrossRef] [Green Version]
- Goonoo, N.; Fahmi, A.; Jonas, U.; Gimie, F.; Arsa, I.A.; Benard, S.; Schonherr, H.; Bhaw-Luximon, A. Improved Multicellular Response, Biomimetic Mineralization, Angiogenesis, and Reduced Foreign Body Response of Modified Polydioxanone Scaffolds for Skeletal Tissue Regeneration. ACS Appl. Mater. Interfaces 2019, 11, 5834–5850. [Google Scholar] [CrossRef]
- Hokmabad, V.R.; Davaran, S.; Aghazadeh, M.; Alizadeh, E.; Salehi, R.; Ramazani, A. Effect of incorporating Elaeagnus angustifolia extract in PCL-PEG-PCL nanofibers for bone tissue engineering. Chem. Sci. Eng. 2019, 13, 108–119. [Google Scholar] [CrossRef]
- Li, H.; Shen, S.; Fu, H.; Wang, Z.; Li, X.; Sui, X.; Yuan, M.; Liu, S.; Wang, G.; Guo, Q. Immunomodulatory Functions of Mesenchymal Stem Cells in Tissue Engineering. Stem Cells Int. 2019, 2019, 9671206. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Ma, J. Effect of NGR1 on the Atopic Dermatitis Model and its Mechanisms. Open Med. 2019, 14, 847–853. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.; Guo, J.; Li, M. RANKL-independent modulation of osteoclastogenesis. J. Oral Biosci. 2019, 61, 16–21. [Google Scholar] [CrossRef]
- Tamura, T.; Udagawa, N.; Takahashi, N.; Miyaura, C.; Tanaka, S.; Yamada, Y.; Koishihara, Y.; Ohsugi, Y.; Kumaki, K.; Taga, T.; et al. Soluble interleukin-6 receptor triggers osteoclast formation by interleukin 6. Proc. Natl. Acad. Sci. USA 1993, 90, 11924–11928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razazian, M.; Khosravi, M.; Bahiraii, S.; Uzan, G.; Shamdani, S.; Naserian, S. Differences and similarities between mesenchymal stem cell and endothelial progenitor cell immunoregulatory properties against T cells. World J. Stem Cells 2021, 13, 971–984. [Google Scholar] [CrossRef] [PubMed]
- Naserian, S.; Shamdani, S.; Arouche, N.; Uzan, G. Regulatory T cell induction by mesenchymal stem cells depends on the expression of TNFR2 by T cells. Stem Cell Res. Ther. 2020, 11, 534. [Google Scholar] [CrossRef] [PubMed]
- Beldi, G.; Khosravi, M.; Abdelgawad, M.E.; Salomon, B.L.; Uzan, G.; Haouas, H.; Naserian, S. TNFalpha/TNFR2 signaling pathway: An active immune checkpoint for mesenchymal stem cell immunoregulatory function. Stem Cell Res. Ther. 2020, 11, 281. [Google Scholar] [CrossRef]
- Beldi, G.; Bahiraii, S.; Lezin, C.; Barkestani, M.N.; Abdelgawad, M.E.; Uzan, G.; Naserian, S. TNFR2 Is a Crucial Hub Controlling Mesenchymal Stem Cell Biological and Functional Properties. Cell Dev. Biol. 2020, 8, 1408. [Google Scholar] [CrossRef]
- Zhao, S.; Yan, L.; Li, X.; Zhang, Z.; Sun, Y.; Wang, J. Notoginsenoside R1 suppresses wear particle-induced osteolysis and RANKL mediated osteoclastogenesis In Vivo and In Vitro. Int. Immunopharmacol. 2017, 47, 118–125. [Google Scholar] [CrossRef]
- Cao, W.; Jin, J.; Wu, G.; Bravenboer, N.; Helder, M.N.; Pathak, J.L.; Zandieh-Doulabi, B.; Hogervorst, J.M.A.; Matsukawa, S.; Geonzon, L.C.; et al. K-Carrageenan Stimulates Pre-Osteoblast Proliferation and Osteogenic Differentiation: A Potential Factor for the Promotion of Bone Regeneration? Molecules 2021, 26, 6131. [Google Scholar] [CrossRef]
- Shah, D.; Son, K.N.; Kalmodia, S.; Lee, B.S.; Ali, M.; Balasubramaniam, A.; Shukla, D.; Aakalu, V.K. Wound Healing Properties of Histatin-5 and Identification of a Functional Domain Required for Histatin-5-Induced Cell Migration. Mol. Ther. Methods Clin. Dev. 2020, 17, 709–716. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences |
|---|---|
| ALP | Forward: 5′-GGACCATTCCCACGTCTTCAC-3′ Reverse: 5′-CCTTGTAGCCAGGCCCATTG-3′ |
| COL1A1 | Forward: 5′-GTGCCAAGGGTCTGACTGGAA-3′ Reverse: 5′-ATCACACCAGCCTGACCACG-3′ |
| OCN | Forward: 5′-CTCACACTCCTCGCCCTATTGG-3′ Reverse: 5′-GTAGCGCCTGGGTCTCTTCACT-3′ |
| VEGF | Forward: 5′-GGAGGCAGAGAAAAGAGAAAGTGT-3′ Reverse: 5′-TAAGAGAGCAAGAGAGAGCAAAAGA-3′ |
| RANKL | Forward: 5′- TGATGAAAGGAGGAAGCA-3′ Reverse: 5′- GTAAGGAGGGGTTGGAGA-3′ |
| OPG | Forward: 5′-AACCCCAGAGCGAAATAC-3′ Reverse: 5′-AGCAGGAGACCAAAGACAC-3′ |
| IL-1β | Forward: 5′-ATGATGGCTTATTACAGTGGCAA-3′ Reverse: 5′-GTCGGAGATTCGTAGCTGGA-3′ |
| IL-6 | Forward: 5′-ACTCACCTCTTCAGAACGAATTG-3′ Reverse: 5′-CCATCTTTGGAAGGTTCAGGTTG-3′ |
| TNF-α | Forward: 5′-GAGGCCAAGCCCTGGTATG-3′ Reverse: 5′-CGGGCCGATTGATCTCAGC-3′ |
| IL-10 | Forward: 5′-GACTTTAAGGGTTACCTGGGTTG-3′ Reverse: 5′-TCACATGCGCCTTGATGTCTG-3′ |
| TGF-β | Forward: 5′-CAATTCCTGGCGATACCTCAG-3′ Reverse: 5′-GCACAACTCCGGTGACATCAA-3′ |
| GAPDH | Forward: 5′-GCACCGTCAAGGCTGAGAAC-3′ Reverse: 5′-TGGTGAAGACGCCAGTGGA-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Yan, Y.; Lan, H.; Wei, N.; Zheng, Z.; Wu, L.; Jaspers, R.T.; Wu, G.; Pathak, J.L. Notoginsenoside R1 Promotes Migration, Adhesin, Spreading, and Osteogenic Differentiation of Human Adipose Tissue-Derived Mesenchymal Stromal Cells. Molecules 2022, 27, 3403. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27113403
Wang H, Yan Y, Lan H, Wei N, Zheng Z, Wu L, Jaspers RT, Wu G, Pathak JL. Notoginsenoside R1 Promotes Migration, Adhesin, Spreading, and Osteogenic Differentiation of Human Adipose Tissue-Derived Mesenchymal Stromal Cells. Molecules. 2022; 27(11):3403. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27113403
Chicago/Turabian StyleWang, Haiyan, Yongyong Yan, Haifeng Lan, Nan Wei, Zhichao Zheng, Lihong Wu, Richard T. Jaspers, Gang Wu, and Janak L. Pathak. 2022. "Notoginsenoside R1 Promotes Migration, Adhesin, Spreading, and Osteogenic Differentiation of Human Adipose Tissue-Derived Mesenchymal Stromal Cells" Molecules 27, no. 11: 3403. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27113403