
Seed Priming with Devosia sp. Cell-Free Supernatant (CFS) and Citrus Bioflavonoids Enhance Canola and Soybean Seed Germination


Abstract
:1. Introduction
2. Results
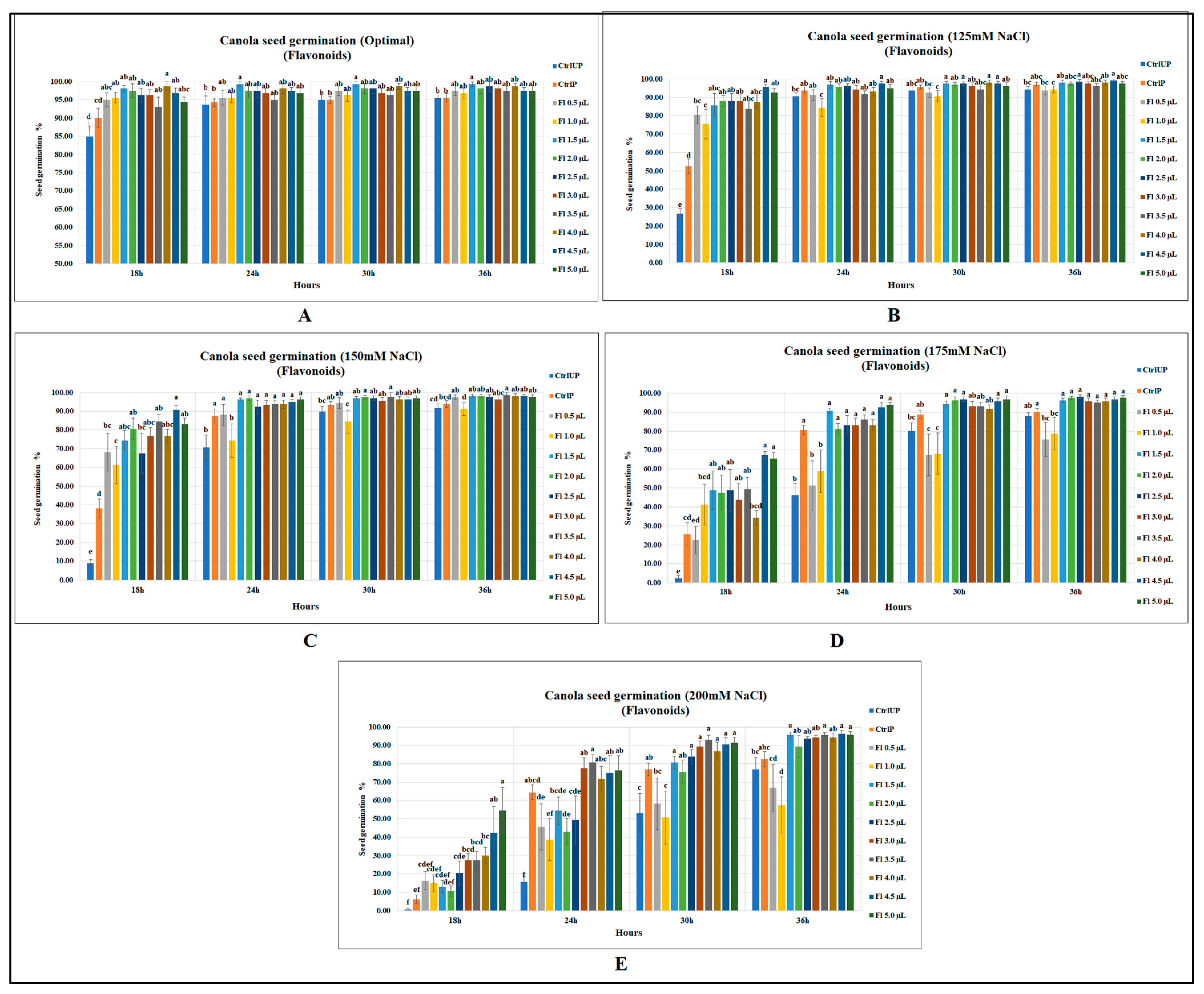
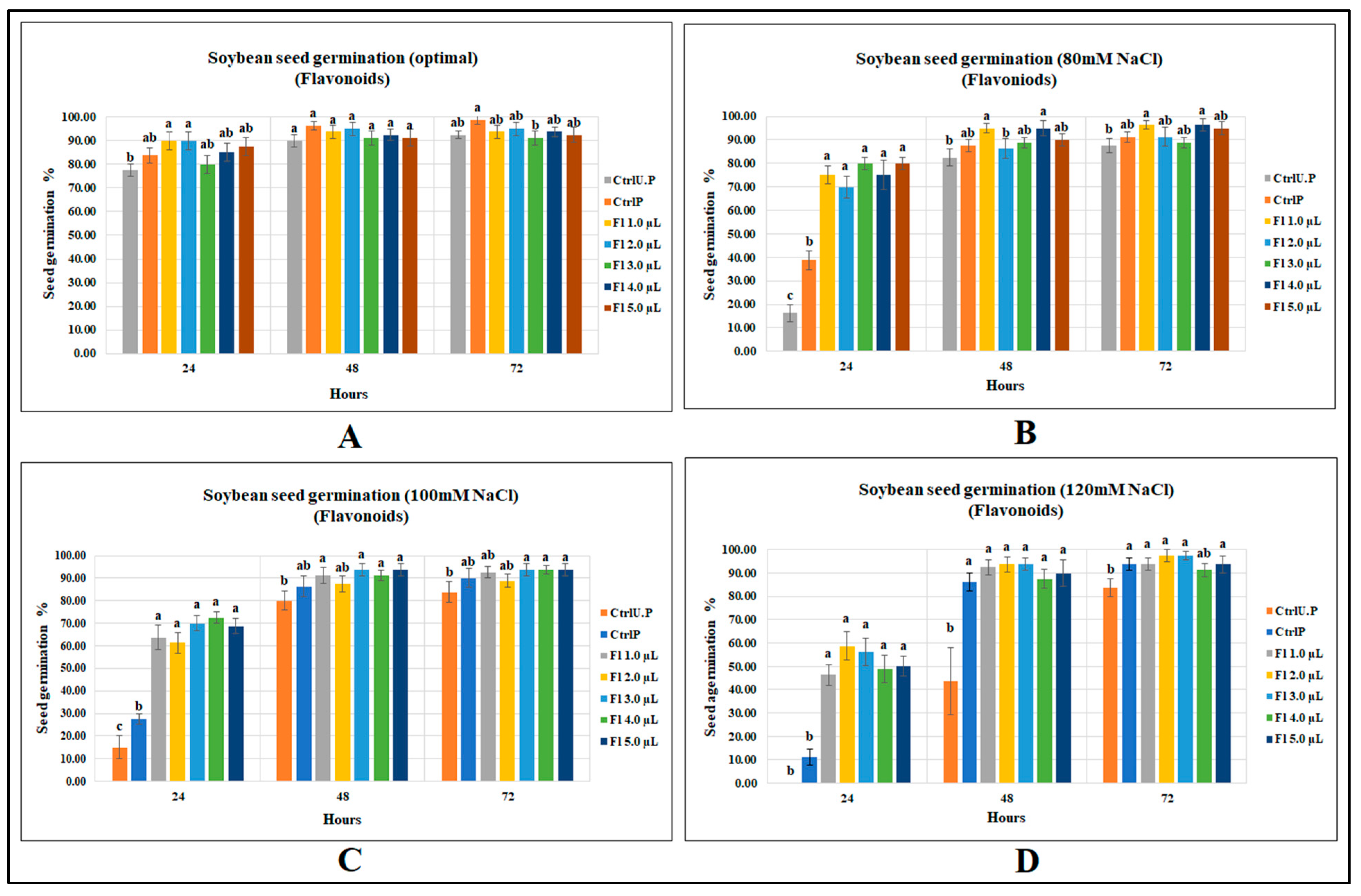
2.1. Effect of Flavonoids on Seed Germination of Canola and Soybean
2.1.1. Canola Seed Germination
2.1.2. Soybean Seed Germination
2.2. Effects of Cell-Free Supernatant on Seed Germination of Canola and Soybean
2.2.1. Canola Seed Germination
2.2.2. Soybean SEED Germination
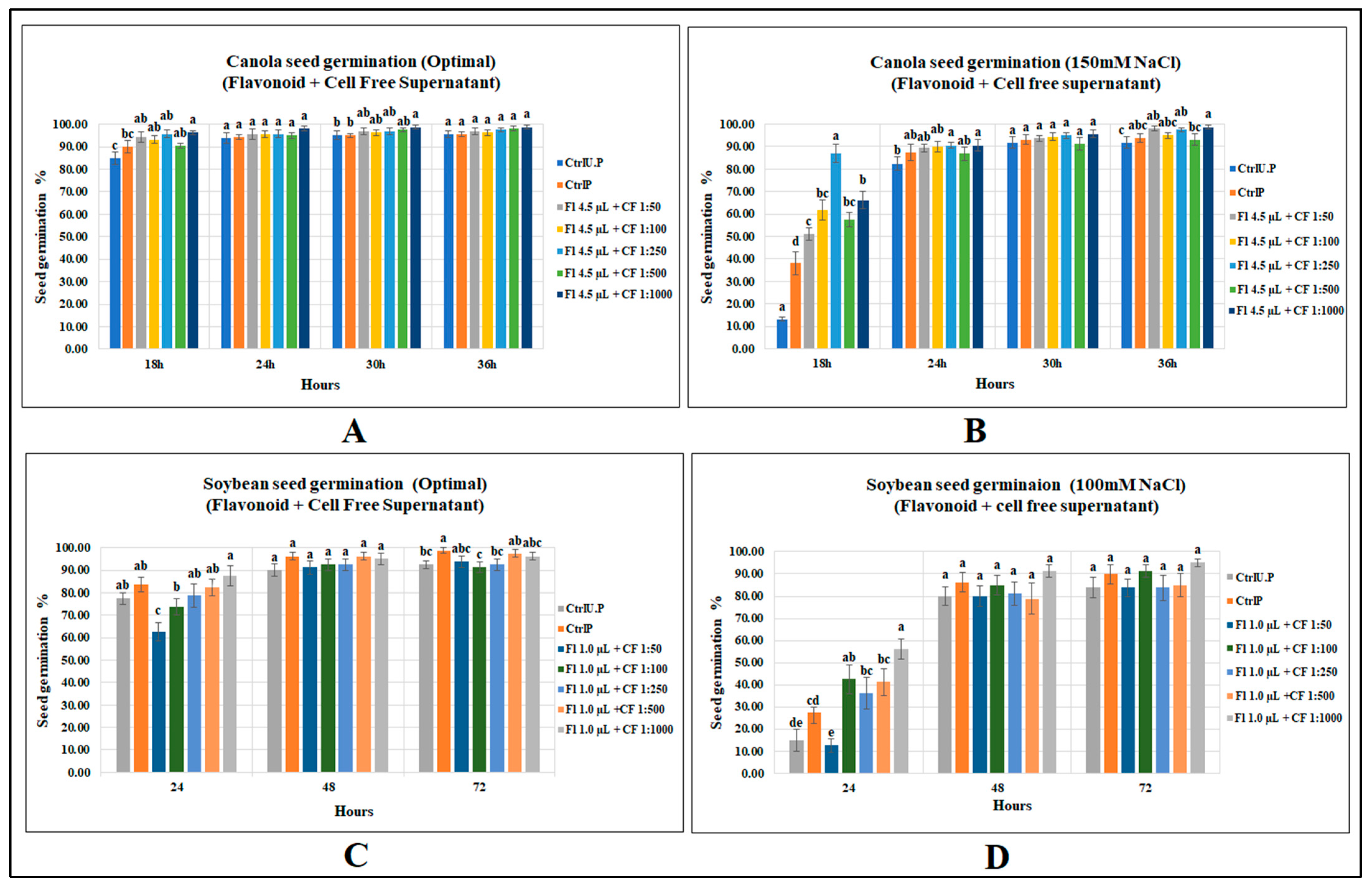
2.3. Effects of Flavonoids and CFS Mixtures on Seed Germination of Canola and Soybean
2.3.1. Canola Seed Germination
2.3.2. Soybean Seed Germination
3. Discussion
3.1. Flavopriming
3.2. Cell-Free Supernatant
3.3. Combined Application of Flavonoids and CFS
4. Material and Methods
4.1. Flavonoids
4.1.1. Preparation of Flavonoid Solution for Seed Treatment
4.1.2. Seed Preparation and Priming
4.1.3. Experimental Design and Treatments
4.2. Cell-Free Supernatant
4.2.1. Propagation of Bacteria and CFS Extraction
4.2.2. Experimental Design and Treatments
4.3. Flavonoids and Cell-Free Supernatant Combined
4.4. Data Collection
4.5. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pathan, S.; Lee, J.-D.; Shannon, J.G.; Nguyen, H.T. Recent advances in breeding for drought and salt stress tolerance in soybean. In Advances in Molecular Breeding toward Drought and Salt Tolerant Crops; Springer: Berlin/Heidelberg, Germany, 2007; pp. 739–773. [Google Scholar] [CrossRef]
- Rengasamy, P. World salinization with emphasis on Australia. J. Exp. Bot. 2006, 57, 1017–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaleem, F.; Shabir, G.; Aslam, K.; Rasul, S.; Manzoor, H.; Shah, S.M.; Khan, A.R. An Overview of the Genetics of Plant Response to Salt Stress: Present Status and the Way Forward. Appl. Biochem. Biotechnol. 2018, 186, 306–334. [Google Scholar] [CrossRef]
- Yildiz, M.; Poyraz, İ.; Çavdar, A.; Özgen, Y.; Beyaz, R. Plant Responses to Salt Stress. In Plant Breeding—Current and Future Views; Abdurakhmonov, I.Y., Ed.; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Yari, L.; Aghaalikani, M.; Khazaei, F. Effect of seed priming duration and temperature on seed germination behavior of bread wheat (Triticum aestivum L.). ARPN J. Agric. Biol. Sci. 2010, 5, 1–6. [Google Scholar]
- Kan, G.; Zhang, W.; Yang, W.; Ma, D.; Zhang, D.; Hao, D.; Hu, Z.; Yu, D. Association mapping of soybean seed germination under salt stress. Mol. Genet. Genom. 2015, 290, 2147–2162. [Google Scholar] [CrossRef]
- Miransari, M.; Smith, D. Alleviating salt stress on soybean (Glycine max (L.) Merr.)–Bradyrhizobium japonicum symbiosis, using signal molecule genistein. Eur. J. Soil Biol. 2009, 45, 146–152. [Google Scholar] [CrossRef]
- Mahmood, T.; Basra, S.M. Invigoration of low vigor sunflower hybrids by seed priming. Int. J. Agric. Biol. 2009, 11, 521–528. [Google Scholar]
- Govindaraj, M.; Masilamani, P.; Albert, V.A.; Bhaskaran, M. Role of antioxidant in seed quality-A review. Agric. Rev. 2017, 38, 180–190. [Google Scholar] [CrossRef]
- Hosseini, M.K.; Powell, A.A.; Bingham, I.J. Comparison of the seed germination and early seedling growth of soybean in saline conditions. Seed Sci. Res. 2002, 12, 165. [Google Scholar] [CrossRef]
- Bano CAmist, N.; Singh, N.B. Morphological and Anatomical Modifications of Plants for Environmental Stresses. In Molecular Plant Abiotic Stress; Roychoudhury, A., Tripathi, D., Eds.; Wiley Online Library: Hoboken, NJ, USA, 2019. [Google Scholar] [CrossRef]
- Treutter, D. Significance of flavonoids in plant resistance: A review. Environ. Chem. Lett. 2006, 4, 147–157. [Google Scholar] [CrossRef]
- Shah, A.; Smith, D.L. Flavonoids in agriculture: Chemistry and roles in, biotic and abiotic stress responses, and microbial associations. Agronomy 2020, 10, 1209. [Google Scholar] [CrossRef]
- Antar, M.; Lyu, D.; Nazari, M.; Shah, A.; Zhou, X.; Smith, D.L. Biomass for a sustainable bioeconomy: An overview of world biomass production and utilization. Renew. Sustain. Energy Rev. 2021, 139, 110691. [Google Scholar] [CrossRef]
- Subramanian, S.; Smith, D.L. Bacteriocins from the rhizosphere microbiome–from an agriculture perspective. Front. Plant Sci. 2015, 6, 909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, S.; Souleimanov, A.; Smith, D.L. Proteomic studies on the effects of lipo-chitooligosaccharide and thuricin 17 under unstressed and salt stressed conditions in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 1314. [Google Scholar] [CrossRef] [PubMed]
- Nazari, M.; Smith, D.L. A PGPR-produced bacteriocin for sustainable agriculture: A review of thuricin 17 characteristics and applications. Front. Plant Sci. 2020, 11, 916. [Google Scholar] [CrossRef]
- Shirley, B.W. Flavonoids in seeds and grains: Physiological function, agronomic importance and the genetics of biosynthesis. Seed Sci. Res. 1998, 8, 415–422. [Google Scholar] [CrossRef]
- Song, C.; Xiang, D.-B.; Yan, L.; Song, Y.; Zhao, G.; Wang, Y.-H.; Zhang, B.-L. Changes in seed growth, levels and distribution of flavonoids during tartary buckwheat seed development. Plant Prod. Sci. 2016, 19, 518–527. [Google Scholar] [CrossRef] [Green Version]
- Falcone Ferreyra, M.L.; Rius, S.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef] [Green Version]
- Graham, T.L. Flavonoid and Isoflavonoid Distribution in Developing Soybean Seedling Tissues and in Seed and Root Exudates 1. Plant Physiol. 1991, 95, 594–603. [Google Scholar] [CrossRef] [Green Version]
- Palma-Tenango, M.; Soto-Hernández, M.; Aguirre-Hernández, E. Flavonoids in agriculture. In Flavonoids-From Biosynthesis to Human Health; IntechOpen: London, UK, 2017. [Google Scholar]
- Nandakumar, L.; Rangaswamy, N. Effect of some flavonoids and phenolic acids on seed germination and rooting. J. Exp. Bot. 1985, 36, 1313–1319. [Google Scholar] [CrossRef]
- Yoshioka, T.; Inokuchi, T.; Fujioka, S.; Kimura, Y. Phenolic compounds and flavonoids as plant growth regulators from fruit and leaf of Vitex rotundifolia. Z. Für Nat. C 2004, 59, 509–514. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Fan, R.; Guo, S.; Wang, P.; Zhu, X.; Fan, Y.; Chen, Y.; He, K.; Kumar, A.; Shi, J.; et al. The Arabidopsis MYB transcription factor, MYB111 modulates salt responses by regulating flavonoid biosynthesis. Environ. Exp. Bot. 2019, 166, 103807. [Google Scholar] [CrossRef]
- Kaur, S.; Gupta, A.K.; Kaur, N. Effect of osmo-and hydropriming of chickpea seeds on seedling growth and carbohydrate metabolism under water deficit stress. Plant Growth Regul. 2002, 37, 17–22. [Google Scholar] [CrossRef]
- Mohammadi, G. The influence of NaCl priming on seed germination and seedling growth of canola (Brassica napus L.) under salinity conditions. Am. Eurasian J. Agric. Environ. Sci. 2009, 5, 696–700. [Google Scholar]
- Arif, M.; Jan, M.T.; Marwat, K.B.; Khan, M.A. Seed priming improves emergence and yield of soybean. Pak. J. Bot. 2008, 40, 1169–1177. [Google Scholar]
- Kaymak, H.Ç.; Güvenç, İ.; Yarali, F.; Dönmez, M.F. The effects of bio-priming with PGPR on germination of radish (Raphanus sativus L.) seeds under saline conditions. Turk. J. Agric. For. 2009, 33, 173–179. [Google Scholar]
- Moeinzadeh, A.; Sharif-Zadeh, F.; Ahmadzadeh, M.; Tajabadi, F.H. Biopriming of Sunflower (‘Helianthus annuus’ L.) Seed with “Pseudomonas fluorescens” for Improvement of Seed Invigoration and Seedling Growth. South. Cross J. 2010, 4, 564–570. [Google Scholar]
- Yadav, J.; Verma, J.P.; Tiwari, K.N. Effect of plant growth promoting rhizobacteria on seed germination and plant growth chickpea (Cicer arietinum L.) under in vitro conditions. Biol. Forum. 2010, 2, 15–18. [Google Scholar]
- Nezarat, S.; Gholami, A. Screening plant growth promoting rhizobacteria for improving seed germination, seedling growth and yield of maize. Pak. J. Biol. Sci. 2009, 12, 26. [Google Scholar] [CrossRef]
- Mia, M.B.; Shamsuddin, Z.; Mahmood, M. Effects of rhizobia and plant growth promoting bacteria inoculation on germination and seedling vigor of lowland rice. Afr. J. Biotechnol. 2012, 11, 3758–3765. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.S.; Kosanke, J.; Gygi, B.; Reed, P.; Habib, A. LCO applications provide improved responses with legumes and non-legumes. In Plant Growth-Promoting Rhizobacteria (PGPR) for Sustainable Agriculture, Proceedings of the 2nd Asian PGPR Conference, Beijing, China, 21–24 August 2011; Asian PGPR Society: Telangana, India, 2011; pp. 54–58. [Google Scholar]
- Prithiviraj, B.; Zhou, X.; Souleimanov, A.; Kahn, W.; Smith, D. A host-specific bacteria-to-plant signal molecule (Nod factor) enhances germination and early growth of diverse crop plants. Planta 2003, 216, 437–445. [Google Scholar] [CrossRef]
- Miransari, M.; Smith, D. Rhizobial lipo-chitooligosaccharides and gibberellins enhance barley (Hordeum vulgare L.) seed germination. Biotechnology 2009, 8, 270–275. [Google Scholar] [CrossRef]
- Gautam, K.; Schwinghamer, T.D.; Smith, D.L. The response of soybean to nod factors and a bacteriocin. Plant Signal. Behav. 2016, 11, e1241934. [Google Scholar] [CrossRef] [PubMed]
- Hung, R.; Lee, S.; Rodriguez-Saona, C.; Bennett, J.W. Common gas phase molecules from fungi affect seed germination and plant health in Arabidopsis thaliana. AMB Express 2014, 4, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gfeller, V.; Huber, M.; Förster, C.; Huang, W.; Köllner, T.; Erb, M. Root volatiles in plant–plant interactions I: High root sesquiterpene release is associated with increased germination and growth of plant neighbours. Plant Cell Environ. 2019, 42, 1950–1963. [Google Scholar] [CrossRef] [Green Version]
- Fincheira, P.; Parada, M.; Quiroz, A. Volatile organic compounds stimulate plant growing and seed germination of Lactuca sativa. J. Soil Sci. Plant Nutr. 2017, 17, 853–867. [Google Scholar] [CrossRef] [Green Version]
- Ilangumaran, G.; Schwinghamer, T.D.; Smith, D.L. Rhizobacteria from Root Nodules of an Indigenous Legume Enhance Salinity Stress Tolerance in Soybean. Front. Sustain. Food Syst. 2021, 4, 308. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Canola | Soybean | ||
|---|---|---|---|
| Flavonoid Levels | Salt Stress (NaCl) Levels | Flavonoid Levels | Salt Stress (NaCl) Levels |
|
|
|
|
| Experimental Units (Seed Germination-Cell-Free Supernatant) | |||
|---|---|---|---|
| Canola | Soybean | ||
| Unstressed | Stressed (NaCl) | Unstressed | Stressed (NaCl) |
| Control—I | Control—I | Control—I | Control—I |
| (Unprimed) | 150 mM NaCl (Unprimed) | (Unprimed) | 100 mM NaCl (Unprimed) |
| Control—II | Control—II | Control—II | Control—II |
| (hydroprimed) | 150 mM NaCl (hydroprimed) | (hydroprimed) | 100mM NaCl (hydroprimed) |
| CFS 1:50 | CFS 1:50 + 150 mM NaCl | CFS 1:50 | CFS 1:50 + 100 mM NaCl |
| CFS 1:100 | CFS 1:100 + 150 mM NaCl | CFS 1:100 | CFS 1:100 + 100 mM NaCl |
| CFS 1:250 | CFS 1:250 + 150 mM NaCl | CFS 1:250 | CFS 1:250 + 100 mM NaCl |
| CFS 1:500 | CFS 1:500 + 150 mM NaCl | CFS 1:500 | CFS 1:500 + 100 mM NaCl |
| CFS 1:1000 | CFS 1:1000 + 150 mM NaCl | CFS 1:1000 | CFS 1:1000 + 100 mM NaCl |
| Experimental Units (Seed Germination-Flavonoids and Cell-Free Supernatant Combined) | |||
|---|---|---|---|
| Canola | Soybean | ||
| Unstressed | Stressed (NaCl) | Unstressed | Stressed (NaCl) |
| Control—I | Control—I | Control—I | Control—I |
| (Unprimed) | 150 mM NaCl (Unprimed) | (Unprimed) | 100 mM NaCl (Unprimed) |
| Control—II | Control—II | Control—II | Control—II |
| (hydroprimed) | 150 mM NaCl (hydroprimed) | (hydroprimed) | 100 mM NaCl (hydroprimed) |
| 4.5 µL + CFS 1:50 | 4.5 µL + CFS 1:50 (150 mM NaCl) | 1.0 µL + CFS 1:50 | 1.0 µL + CFS 1:50 + 100 mM NaCl |
| 4.5 µL + CFS 1:100 | 4.5 µL + CFS 1:100 (150 mM NaCl) | 1.0 µL + CFS 1:100 | 1.0 µL + CFS 1:100 + 100 mM NaCl |
| 4.5 µL + CFS 1:250 | 4.5 µL + CFS 1:250 (150 mM NaCl) | 1.0 µL + CFS 1:250 | 1.0 µL + CFS 1:250 + 100 mM NaCl |
| 4.5 µL + CFS 1:500 | 4.5 µL + CFS 1:500 (150 mM NaCl) | 1.0 µL + CFS 1:500 | 1.0 µL + CFS 1:500 + 100 mM NaCl |
| 4.5 µL + CFS 1:1000 | 4.5 µL + CFS 1:1000 (150 mM NaCl) | 1.0 µL + CFS 1:1000 | 1.0 µL + CFS 1:1000 + 100 mM NaCl |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shah, A.; Subramanian, S.; Smith, D.L. Seed Priming with Devosia sp. Cell-Free Supernatant (CFS) and Citrus Bioflavonoids Enhance Canola and Soybean Seed Germination. Molecules 2022, 27, 3410. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27113410
Shah A, Subramanian S, Smith DL. Seed Priming with Devosia sp. Cell-Free Supernatant (CFS) and Citrus Bioflavonoids Enhance Canola and Soybean Seed Germination. Molecules. 2022; 27(11):3410. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27113410
Chicago/Turabian StyleShah, Ateeq, Sowmyalakshmi Subramanian, and Donald L. Smith. 2022. "Seed Priming with Devosia sp. Cell-Free Supernatant (CFS) and Citrus Bioflavonoids Enhance Canola and Soybean Seed Germination" Molecules 27, no. 11: 3410. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27113410