
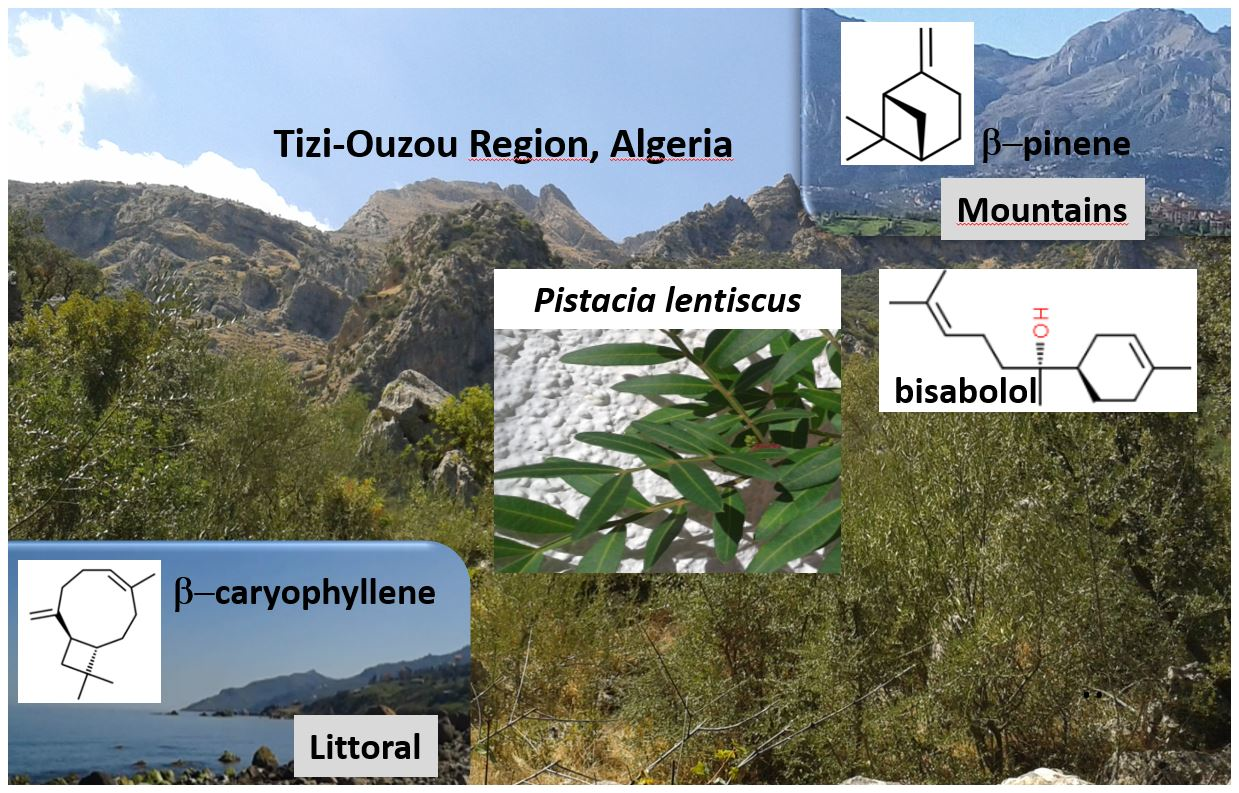
Profiling of Essential Oils from the Leaves of Pistacia lentiscus Collected in the Algerian Region of Tizi-Ouzou: Evidence of Chemical Variations Associated with Climatic Contrasts between Littoral and Mountain Samples

 ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Qualitative and Semi-Quantitative Composition of the Essential Oils
2.2. Quantitative Variations of the Major Compounds and Terpenic Classes
2.3. Multivariate Analysis of All the Components Detected in the EOs
2.4. Antioxidant Activity
2.5. Profiling of the Fatty Acids from the Leaf Acyl Lipids
2.6. Multivariate Analysis of the Fatty Acids Detected (as Methyl Ester: FAMEs) in the Leaf Acyl-Lipids
3. Materials and Methods
3.1. Sampling and Botanical Identification
3.2. Extraction of Essential Oils
3.3. GC-MS Analysis of Essential Oils
3.4. Antioxidant Activity
3.4.1. Radical Scavenging Test (DPPH)
3.4.2. Ferric Reducing Power Test (FRAP)
3.4.3. Radical Cation Reduction Test (ABTS)
3.5. Preparation of Lipidic Extracts from Lentisk Leaves and GC-MS Analysis
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benhammou, N.B.; Belyagoubi, L.; El Zerey-Belaskri, A.; Zitouni, A.; Ghembaza, N.; Benhassaini, H.; Atik-Bekkara, F.; Piras, A.; Falconieri, D.; Rosa, A. Fatty acid composition and antioxidant activity of Pistacia lentiscus L. fruit fatty oil from Algeria. J. Food Meas. Charact. 2018, 12, 1408–1412. [Google Scholar] [CrossRef]
- Zaouali, Y.; Bel HadjYahya, I.; Jaouadi, R.; Messaoud, C.; Boussaid, M. Sex-related differences in essential oil composition, phenol contents and antioxidant activity of aerial parts in Pistacia lentiscus L. during seasons. Ind. Crop. Prod. 2018, 121, 151–159. [Google Scholar]
- Gardeli, C.; Papageorgiou, V.; Mallouchos, A.; Theodosis, K.; Komaitis, M. Essential oilcomposition of Pistacia lentiscus L. and Myrtuscommunis L.: Evaluation of antioxidant capacity of methanolic extracts. Food Chem. 2008, 107, 1120–1130. [Google Scholar] [CrossRef]
- Landau, S.; Muklada, H.; Markovics, A.; Azaizeh, H. Traditional uses of Pistacia lentiscus in veterinary and human medicine. In Medicinal and Aromatic Plants of the Middle-East; Springer: Dordrecht, The Netherlands, 2014; Volume 2, pp. 163–180. [Google Scholar]
- Mehenni, C.; Atmani-Kilani, D.; Dumarçay, S.; Perrin, D.; Gérardin, P.; Atmani, D. Hepatoprotective and antidiabetic effects of Pistacia lentiscus leaf and fruit extracts. J. Food Drug Anal. 2016, 24, 653–669. [Google Scholar] [CrossRef] [PubMed]
- Said, O.; Khalil, K.; Fulder, S.; Azaizeh, H. Ethno-pharmacological survey of medicinal herbs in Israel, the Golan Heights and the West Bank region. J. Ethnopharmacol. 2002, 83, 251–265. [Google Scholar] [CrossRef]
- Jamila, F.; Mostafa, E. Ethnobotanical survey of medicinal plants used by people in Oriental Morocco to manage various ailments. J. Ethnopharmacol. 2014, 154, 76–87. [Google Scholar] [CrossRef]
- Guarrera, P.M.; Salerno, G.; Caneva, G. Folk phytotherapeutical plants from Maratea area (Basilicata, Italy). J. Ethnopharmacol. 2005, 99, 367–378. [Google Scholar] [CrossRef]
- Abdeldjelil, M.C.; Bensegueni, A.; Messaï, A.; Agabou, A.; Benazzouz, H. Medicinal use of Pistacia lentiscus fixed oil in Constantine province, north-east Algeria. J. Nat. Prod. Plant Resour. 2014, 4, 48–51. [Google Scholar]
- Nahida, A.S.; Siddiqui, A.N. Pistacia lentiscus: A review on phytochemistry and pharmacological properties. Int. J. Pharm. Pharm. Sci. 2012, 4, 16–20. [Google Scholar]
- Bozorgi, M.; Memariani, Z.; Mobli, M.; Surmaghi, S.M.H.; Reza Shams-Ardekan, M.; Rahimi, R. Five pistacia species (P. vera, P. atlantica, P. terebinthus, P. khinjuk and P. lentiscus): A review of their traditional uses, phytochemistry, and pharmacology. Sci. World J. 2013, 2013, 219815. [Google Scholar] [CrossRef] [Green Version]
- Gratani, L.; Catoni, R.; Varone, L. Morphological, anatomical and physiological leaf traits of Q. ilex, P. latifolia, P. Lentiscus. And M. communis and their response to Mediterranean climate stress factors. Bot. Stud. 2013, 54, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gourine, N.; Yousfi, M.; Bombarda, I.; Nadjemi, B.; Stocker, P.; Gaydou, E.M. Antioxidant activities and chemical composition of essential oil of Pistacia atlantica from Algeria. Ind. Crop. Prod. 2010, 31, 203–208. [Google Scholar] [CrossRef]
- Ahmed, Z.B.; Yousfi, M.; Viaene, J.; Dejaegher, B.; Demeyer, K.; Vander Heyden, Y. Four Pistacia atlantica subspecies (atlantica, cabulica, kurdica and mutica): A review of their botany, ethnobotany, phytochemistry and pharmacology. J. Ethnopharmacol. 2021, 265, 113329. [Google Scholar] [CrossRef] [PubMed]
- Ben Youssef, E.H.; Charchari, S.; Nacer-Bey, N. The essential oil of Pistacia lentiscus L. from Algeria. J. Essent. Oil Res. 2005, 17, 642–644. [Google Scholar] [CrossRef]
- Mecherara-Idjeri, S.; Hassani, A.; Castola, V.; Casanova, J. Composition and chemical variability of the essential oil from Pistacia lentiscus L. growing wild in Algeria Part 1: Leaf oil. J. Essent. Oil Res. 2008, 20, 32–38. [Google Scholar] [CrossRef]
- Amhamdi, H.; Aouinti, F.; Wathelet, J.P.; Elbachiri, A. Chemical composition of the essential oil of Pistacia lentiscus L. from eastern Morocco. Rec. Nat. Prod. 2009, 3, 90–95. [Google Scholar]
- Hafsé, M.; Benbrahim, K.F.; Saidi, A.; Farah, A. Volatile components and antibacterial profile of essential oils extracted from leaves and twigs of Pistacia lentiscus L. Br. Microbiol. Res. J. 2013, 3, 602–611. [Google Scholar] [CrossRef]
- Aissi, O.; Boussaid, M.; Messaoud, C. Essential oil composition in natural populations of Pistacia lentiscus L. from Tunisia: Effect of ecological factor and incidence on antioxidant and antiacetylcholinesterase activities. Ind. Crop. Prod. 2016, 91, 379–382. [Google Scholar] [CrossRef]
- Lo Presti, M.; Sciarrone, D.; Crupi, M.L.; Costa, R.; Ragusa, S.; Dugo, G.; Mondello, L. Evaluation of the volatile and chiral composition in Pistacia lentiscus L. essential oil. Flavour Fragr. J. 2008, 23, 249–257. [Google Scholar] [CrossRef]
- Djenane, D.; Yanguela, J.; Montanes, L.; Djerbal, M.; Roncales, P. Antimicrobial activity of Pistacia lentiscus and Satureja montana essential oils against Listeria monocytogenes CECT 935 using laboratory media: Efficacy and synergistic potential in minced beef. Food Control 2011, 22, 1046–1053. [Google Scholar] [CrossRef]
- Ismail, A.; Hamrouni, L.; Hanana, M.; Jamoussi, B. Chemical composition and herbicidal effects of Pistacia lentiscus L. essential oil against weeds. Int. J. Med. Aromat. Plants 2012, 2, 558–565. [Google Scholar]
- Haouli, A.; Seridi, R.; Djemli, S.; Bourdjiba, O.; Frih, H. Contribution to the analysis of Pistacia lentiscus extracted oil. Am.-Eur. J. Agric. Environ. Sci. 2015, 15, 1075–1081. [Google Scholar]
- Douissa, F.B.; Hayder, N.; Chekir-Ghedira, L.; Hammami, M.; Ghedira, K.; Mariotte, A.M.; Dijoux-Franca, M.G. New study of the essential oil from leaves of Pistacia lentiscus L. (Anacardiaceae) from Tunisia. Flavour Fragr. J. 2005, 20, 410–414. [Google Scholar] [CrossRef]
- Nahum, S.; Inbar, M.; Ne’eman, G.; Ben-Shlomo, R. Phenotypic plasticity and gene diversity in Pistacia lentiscus L. along environmental gradients in Israel. Tree Genet. Genomes 2008, 4, 777–785. [Google Scholar] [CrossRef]
- Ait Said, S.; Fernandez, C.; Greff, S.; Torre, F.; Derridj, A.; Gauquelin, T.; Mevy, J.P. Inter-population variability of terpenoid composition in leaves of Pistacia lentiscus L. from Algeria: A chemoecological approach. Molecules 2011, 16, 2646–2657. [Google Scholar] [CrossRef]
- Carvalho, S.; Macel, M.; Mulder, P.P.J.; Skidmore, A.; van der Putten, W.H. Chemical variation in Jacobaea vulgaris is influenced by the interaction of season and vegetation successional stage. Phytochemistry 2014, 99, 86–94. [Google Scholar] [CrossRef]
- Juvany, M.; Müller, M.; Pintó-Marijuan, M.; Munné-Bosch, S. Sex-related differences in lipid peroxidation and photoprotection in Pistacia lentiscus. J. Exp. Bot. 2014, 65, 1039–1049. [Google Scholar] [CrossRef] [Green Version]
- Negro, C.; De Bellis, L.; Miceli, A. Chemical composition and antioxidant activity of Pistacia lentiscus essential oil from Southern Italy (Apulia). J. Essent. Oil Res. 2015, 27, 23–29. [Google Scholar] [CrossRef]
- Foddai, M.; Kasabri, V.; Afif, F.U.; Azara, E.; Petretto, G.L.; Pintore, G. In vitro inhibitory effects of Sardinian Pistacia lentiscus L. and Pistacia terebinthus L. on metabolic enzymes Pancreatic lipase, -amylase, and -glucosidase. Starch-Stärke 2014, 67, 204–212. [Google Scholar] [CrossRef]
- Llorens-Molina, J.A.; Gonzalez, S.V.; Martinez, J.S. Essential oil composition of leaves of Pistacia lentiscus L. growing wild in Valencia (Spain). Nat. Volatiles Essent. Oils 2015, 2, 17–26. [Google Scholar]
- Bachrouch, O.; Jemâa, J.M.-B.; AidiWaness, W.; Talou, T.; Marzouk, B.; Abdrerraba, M. Composition and insecticidial activity of essential oil from Pistacia lentiscus L. against Ectomyelois ceratoniae Zeller and Ephestia kuehniella Zeller (Lepidoptera: Pyralidae). J. Stored Prod. Res. 2010, 46, 242–247. [Google Scholar] [CrossRef]
- Klibet, F.; Boumendjel, A.; Khiari, M.; El Feki, A.; Abdenour, C.; Messarah, M. Oxidative stress-related liver dysfunction by sodium arsente: Alleviatio by Pistacia lentiscus oil. Pharm. Biol. 2016, 54, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Maes, C.; Meersmans, J.; Lins, L.; Bouquillon, S.; Fauconnier, M.L. Essential Oil-Based Bioherbicides: Human Health Risks Analysis. Int. J. Mol. Sci. 2021, 22, 9396. [Google Scholar] [CrossRef]
- Damiens, A.; Alebrahim, M.T.; Léonard, E.; Fayeulle, A.; Furman, C.; Hilbert, J.L.; Siah, A.; Billamboz, M. Sesamol–based terpenoids as promising bio-sourced crop protection compounds against the wheat pathogen Zymoseptoriatritici. Pest Manag. Sci. 2021, 77, 2403–2414. [Google Scholar] [CrossRef] [PubMed]
- Vidrich, V.; Fusi, P.; Graziano, A.; Silvestrini, E.; Michelozzi, M.; Marco, F. Chemical composition of the essential oil of Pistacia Lentiscus L. J. Essent. Oil Res. 2004, 16, 223–226. [Google Scholar] [CrossRef]
- El Bishbishy, M.H.; Gad, H.A.; Aborehab, N.M. Chemometric discrimination of three Pistacia species via their metabolic profiling and their possible in vitro effects on memory functions. J. Pharm. Biomed. Anal. 2020, 177, 112840. [Google Scholar] [CrossRef]
- Congiu, R.; Falconieri, D.; Marongiu, B.; Piras, A.; Porcedda, S. Extraction and isolation of Pistacia lentiscus L. essential oil by supercritical CO2. Flavour Fragr. J. 2002, 17, 239–244. [Google Scholar] [CrossRef]
- Bouyahya, A.; Assemian, I.C.C.; Mouzount, H.; Bourais, I.; Et-Touys, A.; Fellah, H.; Benjouad, A.; Dakka, N.; Bakri, Y. Could volatile compounds from leaves and fruits of Pistacia lentiscus constitute a novel source of anticancer, antioxidant, antiparasitic and antibacterial drugs? Ind. Crop. Prod. 2019, 128, 62–69. [Google Scholar] [CrossRef]
- Derwich, E.; Manar, A.; Benziane, Z.; Boukir, A. GC/MS analysis and In vitro antibacterial activity of the essential oil isolated from leaf of Pistacia lentiscus growing in morocoo. World Appl. Sci. J. 2010, 8, 1267–1276. [Google Scholar]
- Aouinti, F.; Zidane, H.; Tahri, M.; Wathelet, J.P.; El Bachiri, A. Chemical composition, mineral contents and antioxidant activity of fruits of Pistacia lentiscus L. from Eastern Morocco. J. Mater. Environ. Sci. 2014, 5, 199–206. [Google Scholar]
- Kıvçak, B.; Akay, S. Quantitative determination of α-tocopherol in Pistacia lentiscus, Pistacia lentiscus var. chia, and Pistacia terebinthus by TLC-densitometry and colorimetry. Fitoterapia 2005, 76, 62–66. [Google Scholar] [CrossRef]
- El Idrissi, M.; Barbouchi, M.; Choukrad, M.; Lhoussain, L. Chemical composition and antimicrobial activity of essential oils isolated from leaves and twigs of Pistacia lentiscus L. Growing wild in Morocco. World J. Pharm. Sci. 2016, 5, 516–524. [Google Scholar]
- Hamiani, A.; Kambouche, N.; Bellahouel, S. Chemical composition and antioxidant potential of Pistacia lentiscus L. essential oil from Oran (Algeria). Adv. Biosci. Biotechnol. 2016, 7, 539–544. [Google Scholar]
- Medini, H.; Elaissi, A.; Khouja, M.L.; Chraief, I.; Farhat, F.; Hammami, M.; Chemli, R.; Harzallah-Skhiri, F. Leaf essential oil of Juniperusoxycedrus L. (Cupressaceae) harvested in northern Tunisia: Composition and intra-Specific variability. Chem. Biodivers. 2010, 7, 1254–1266. [Google Scholar] [CrossRef]
- Lakušić, D.V.; Ristić, M.S.; Slavkovska, V.N.; Šinžar-Sekulić, J.B.; Lakušić, B.S. Environment-related variations of the composition of the essential oils of rosemary (Rosmarinus officinalis L.) in the balkan penninsula. Chem. Biodivers. 2012, 9, 1286–1302. [Google Scholar] [CrossRef]
- Rahimmalek, M.; Mirzakhani, M.; Pirbalouti, A.G. Essential oil variation among 21 wild myrtle (Myrtus communis L.) populations collected from different geographical regions in Iran. Ind. Crop. Prod. 2013, 51, 328–333. [Google Scholar] [CrossRef]
- Djerrad, Z.; Kadik, L.; Djouahri, A. Chemical variability and antioxidant activities among Pinushalepensis Mill. essential oils provenances, depending on geographic variation and environmental conditions. Ind. Crop. Prod. 2015, 74, 440–449. [Google Scholar] [CrossRef]
- Edreva, A.; Velikova, V.; Tsonev, T.; Dagnon, S.; Gurel, A.; Aktaş, L.; Gesheva, E. Stress-protective role of secondary metabolites: Diversity of functions and mechanisms. Gen. Appl. Plant Physiol. 2008, 34, 67–78. [Google Scholar]
- Kofidis, G.; Bosabalidis, A.M. Effects of altitude and season on glandular hairs and leaf structural traits of Nepetanuda L. Bot. Stud. 2008, 49, 363–372. [Google Scholar]
- Abreu, I.N.; Mazzafera, P. Effect of water and temperature stress on the content of active constituents of Hypericum brasiliense Choisy. Plant Physiol. Biochem. 2005, 43, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Cirak, C.; Radusiene, J.; Jakstas, V.; Ivanauskas, L.; Seyis, F.; Yayla, F. Altitudinal changes in secondary metabolite contents of Hypericum androsaemum and Hypericum polyphyllum. Biochem. Syst. Ecol. 2017, 70, 108–115. [Google Scholar] [CrossRef]
- Barra, A.; Coroneo, V.; Dessi, S.; Cabras, P.; Angioni, A. Characterization of the volatile constituents in the essential oil of Pistacia lentiscus L. from different origins and its antifungal and antioxidant activity. J. Agric. Food Chem. 2007, 55, 7093–7098. [Google Scholar] [CrossRef] [PubMed]
- Bampouli, A.; Kyriakopoulou, K.; Papaefstathiou, G.; Louli, V.; Krokida, M.; Magoulas, K. Comparison of different extraction methods of Pistacia lentiscus var. chia leaves: Yield: Antioxidant activity and essential oil chemical composition. J. Appl. Res. Med. Aromat. Plants 2014, 1, 81–91. [Google Scholar] [CrossRef]
- Desmet, S.; Morreel, K.; Dauwe, R. Origin and Function of Structural Diversity in the Plant Specialized Metabolome. Plants 2021, 10, 2393. [Google Scholar] [CrossRef]
- Nguyen, T.K.O.; Nguyen, N.L.; Pham, H.N.; Sawada, Y.; Hirai, M.Y.; Dauwe, R.; Dijoux-Franca, M.G. Development of a Pteris vittata L. compound database by widely targeted metabolomics profiling. Biomed. Chromatogr. BMC 2021, 35, e5110. [Google Scholar] [CrossRef] [PubMed]
- Olszowy, M.; Dawidowicz, A.L. Essential oils as antioxidants: Their evaluation by DPPH, ABTS, FRAP, CUPRAC, and β-carotene bleaching methods. Monatsh. Chem. 2016, 147, 2083–2091. [Google Scholar] [CrossRef]
- Mannion, D.T.; Furey, A.; Kilcawley, K.N. Free fatty acids quantification in dairy products. Int. J. Dairy Technol. 2016, 69, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Diamantoglou, S.; Meletiou-christou, M.S. Das jahres periodische Verhalten des Rohfetts und der Fettsäuren in Rinden und Bläuern von Pistacia lentiscus, Pistacia terebinthus und Pistacia vera: The Lipid Content and Fatty Acid Composition of Barks and Leaves of Pistacia lentiscus, Pistacia terebinthus and Pistacia vera During the Course of a Year. Z. Pflanzenphysiol. 1979, 93, 219–228. [Google Scholar]
- Akdemir, O.F.; Tilka, E.; Onay, A.; Keskin, C.; Bashan, M.; Kilinc, F.M.; Kizmaz, V.; Suzerer, V. Determination of the fatty acid composition of the fruits and different organs of lentisk (Pistacia lentiscus L.). J. Essent. Oil-Bear Plants 2015, 18, 1224–1233. [Google Scholar] [CrossRef]
- Harrat, M.; Benalia, M.; Gourine, N.; Yousfi, M. Variability of the chemical compositions of fatty acids, tocopherols and lipids antioxidant activities, obtained from the leaves of Pistacia lentiscus L. growing in Algeria. Mediterr. J. Nutr. Metab. 2018, 11, 199–215. [Google Scholar] [CrossRef]
- Martz, F.; KiviniemI, S.; Heino, P.; Palva, T.; Sutinen, M.L. Genetic control of fatty acid desaturation during cold acclimation and de-acclimation of birch. Acta Hortic. 2003, 618, 137–146. [Google Scholar] [CrossRef]
- Gontier, E.; Gougeon, S.; Guillot, X.; Thomasset, B.; Méjean, L.; Tran, T.L.M.; Bourgaud, F. Les plantes, sources d’acides gras essentiels oméga 3. Oléagineux Corp. Gras Lipides OCL-J. 2004, 11, 106–111. [Google Scholar] [CrossRef] [Green Version]
- Climate-data.org. Available online: https://fr.climate-data.org/afrique/algerie/tizi-ouzou/ait-irane-693594/ (accessed on 5 February 2022).
- Climate-data.org. Available online: https://fr.climate-data.org/afrique/algerie/tizi-ouzou/tigzirt-321206/ (accessed on 5 February 2022).
- Arabi, A.; Djibaoui, R.; Malihac, C.; Sisbane, I.; Lattab, A.; Bechelaghem, N.; Dahah, H.; Charef, R.; Ettalhi, M.; Taleb, F.; et al. Chemical composition and antibacterial activity of essential oil from leaves and twigs of Pistacia lentiscus growing in Mostaganem Province (Algeria). Int. J. Biosci. 2017, 10, 146–158. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; p. 811. [Google Scholar]
- Doshi, P.; Adsule, P.; Banerjee, K.; Oulkar, D. Phenolic compounds, antioxidant activity and insulinotropic effect of extracts prepared from grape (Vitis vinifera L.) by products. J. Food Sci. Technol. 2015, 52, 181–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messaoud, C.; Laabidi, A.; Boussaid, M. Myrtus communis L. infusions: The effect of infusion time on phytochemical composition antioxidant, and antimicrobial activities. J. Food Sci. 2012, 77, 941–947. [Google Scholar] [CrossRef]
- Choque, E.; Durrieu, V.; Alric, I.; Raynal, J.; Mathieu, F. Impact of spray-drying on biological properties of chitosan matrices supplemented with antioxidant fungal extracts for wine applications. Curr. Microbiol. 2020, 77, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Jousse, C.; Schiltz, S.; Fournez, A.; Guillot, X.; Thomasset, B.; Gougeon, S.; Bourgaud, F.; Gontier, E. Rapid, cost-effective screening of flax genotypes to identify desirable fatty acid compositions. Electron. J. Plant Breed. 2010, 1, 1396–1404. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 21 March 2022).
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
| Means % Content ± Standard Deviation | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mountain | Littoral | Altitude * Season Interaction | ||||||||||
| C | Compounds | IR | Winter | Spring | Summer | Autumn | Winter | Spring | Summer | Autumn | ||
| C1 | MH | tricyclene | 923 | 0.2 ± 0.1 ab | 0.9 ± 0.1 b | 0.8 ± 0.1 ab | 0.5 ± 0.1 a | 0.1 ± 0.1 c | 0.3 ± 0.1 d | 0.2 ± 0.1 d | 0.2 ± 0.1 ab | *** |
| C2 | MH | α-pinene | 935 | 2.6 ± 0.2c | 13.1 ± 0.1b | 12.6 ± 0.3b | 9.4 ± 0.9a | 2.1 ± 0.4c | 5.4 ± 0.4d | 4.9 ± 0.5d | 8.3 ± 0.3a | *** |
| C3 | MH | camphene | 952 | 0.9 ± 0.1 cd | 3.1 ± 0.1 b | 3.2 ± 0.0 d | 1.6 ± 0.0 a | 0.6 ± 0.1 e | 0.8 ± 0.1 f | 1.1 ± 0.1 f | 1.0 ± 0.0 bc | *** |
| C4 | MH | sabinene | 975 | 0.5 ± 0.0 e | 1.4 ± 0.0 c | 1.2 ± 0.1 c | 3.0 ± 0.4 a | 0.3 ± 0.1 f | 1.1 ± 0.1 d | 1.0 ± 0.1 cd | 2.0 ± 0.1 b | * |
| C5 | MH | β-pinene | 981 | 0.9 ± 0.0b | 4.1 ± 0.1a | 7.1 ± 0.0b | 3.6 ± 0.4a | 1.2 ± 0.2b | 1.2 ± 0.1b | 3.7 ± 0.3c | 3.9 ± 0.1a | *** |
| C6 | MH | β-myrcene | 993 | 0.1 ± 0.0c | 0.5 ± 0.0c | 0.3 ± 0.0c | 19.0 ± 0.9b | 0.5 ± 0.1d | 5.0 ± 0.1b | 4.8 ± 0.9ab | 6.1 ± 0.4a | *** |
| C7 | MH | α-phellandrene | 1011 | 0.0 ± 0.0 cd | 0.4 ± 0.0 bc | 0.0 ± 0.0 bc | 0.4 ± 0.0 b | 0.1 ± 0.1 e | 0.1 ± 0.0 de | 0.1 ± 0.0 a | 0.2 ± 0.0 a | *** |
| C8 | Oth | o-cymene | 1032 | 0.2 ± 0.1 c | 0.7 ± 0.1 ab | 0.9 ± 0.1 bc | 0.9 ± 0.1 ab | 0.3 ± 0.1 d | 0.3 ± 0.1 cd | 0.5 ± 0.1 d | 0.6 ± 0.1 a | *** |
| C9 | Oth | p-cymene | 1033 | 0.1 ± 0.1 ac | 0.2 ± 0.1 a | 0.4 ± 0.1 ab | 0.4 ± 0.1 a | 0.1 ± 0.1 bc | 0.1 ± 0.1 a | 0.2 ± 0.1 c | 0.3 ± 0.1 a | |
| C10 | MH | limonene | 1036 | 0.5 ± 0.1 f | 3.8 ± 0.1 f | 2.7 ± 0.0 b | 4.2 ± 0.1 b | 1.4 ± 0.2 e | 4.7 ± 0.1 d | 1.1 ± 0.2 c | 4.9 ± 0.3 a | *** |
| C11 | MH | ɣ-terpinene | 1066 | 0.2 ± 0.0 d | 0.5 ± 0.0 c | 0.3 ± 0.0 c | 0.9 ± 0.0 a | 0.1 ± 0.0 e | 0.3 ± 0.0 d | 0.3 ± 0.0 bc | 0.4 ± 0.0 b | *** |
| C12 | MH | α-terpinolene | 1094 | 0.0 ± 0.0 c | 0.2 ± 0.0 d | 0.0 ± 0.0 bc | 0.2 ± 0.0 b | 0.1 ± 0.0 d | 0.2 ± 0.0 d | 0.1 ± 0.0 a | 0.1± 0.0 a | *** |
| C13 | Oth | 2-nonanaone | 1099 | 0.1 ± 0.0 cd | 0.3 ± 0.0 b | 0.1 ± 0.0 bc | 0.6 ± 0.1 a | 0.1 ± 0.0 d | 0.3 ± 0.0 b | 0.4 ± 0.1 a | 0.5 ± 0.1 a | *** |
| C14 | Oth | nonanol | 1109 | 0.2 ± 0.1 ac | 0.4 ± 0.1 d | 0.3 ± 0.1 bcd | 0.1 ± 0.1 a | 0.1 ± 0.1 ab | 0.5 ± 0.1 cd | 0.4 ± 0.1 ad | 0.3 ± 0.1 ac | |
| C15 | Oth | nonanal | 1114 | 0.0 ± 0.0 a | 0.2 ± 0.0 b | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.0 ± 0.0 a | 0.2 ± 0.0 b | 0.0 ± 0.0 a | 0.0 ± 0.0 a | |
| C16 | OM | menta-2-en-1-ol <E-p> | 1137 | 0.0 ± 0.0 ac | 0.1 ± 0.0 ac | 0.0 ± 0.0 bc | 0.0 ± 0.0 ab | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 c | 0.0 ± 0.0 a | |
| C17 | OM | borneol | 1187 | 0.1 ± 0.0 ab | 0.2 ± 0.0 ab | 0.1 ± 0.0 ab | 0.1 ± 0.0 a | 0.0 ± 0.0 ab | 0.1 ± 0.0 c | 0.1 ± 0.0 ab | 0.1 ± 0.0 b | *** |
| C18 | OM | terpinen-4-ol | 1207 | 1.3 ± 0.1 ce | 1.4 ± 0.0 b | 1.4 ± 0.2 de | 1.7 ± 0.1 a | 0.5 ± 0.0 e | 0.9 ± 0.0 cd | 1.5 ± 0.2 cd | 1.5 ± 0.0 c | *** |
| C19 | OM | α-terpineol | 1209 | 0.4 ± 0.1 d | 0.4 ± 0.1 cd | 0.9 ± 0.1 c | 0.5 ± 0.1 a | 0.3 ± 0.1 c | 0.5 ± 0.1 b | 0.5 ± 0.1 e | 0.6 ± 0.1 ab | *** |
| C20 | OM | bornyl acetate | 1297 | 6.2 ± 0.2a | 7.0 ± 0.1b | 4.9 ± 0.3b | 1.5 ± 0.1c | 2.8 ± 0.1b | 1.9 ± 0.1f | 1.9 ± 0.0d | 0.8 ± 0.0e | *** |
| C21 | Oth | 2-undecanone | 1301 | 1.6 ± 0.0 d | 2.0 ± 0.0 bc | 0.9 ± 0.1 bc | 2.1 ± 0.1 b | 1.6 ± 0.3 cd | 2.0 ± 0.1 c | 1.9 ± 0.2 a | 2.5 ± 0.2 b | *** |
| C22 | OM | α-terpinyl acetate | 1358 | 0.2 ± 0.0 d | 0.3 ± 0.0 f | 0.0 ± 0.0 c | 0.0 ± 0.0 e | 1.0 ± 0.1 a | 1.5 ± 0.1 c | 0.3 ± 0.0 a | 0.5 ± 0.1 b | *** |
| C23 | SH | α-copaene | 1386 | 0.9 ± 0.0 b | 0.8 ± 0.0 c | 1.2 ± 0.0 c | 0.7 ± 0.0 a | 0.3 ± 0.0 d | 0.6 ± 0.0 e | 0.5 ± 0.0 g | 0.4 ± 0.1 f | *** |
| C24 | SH | β-bourbonene | 1395 | 0.0 ± 0.0 ab | 0.1 ± 0.0 de | 0.0 ± 0.0 ac | 0.0 ± 0.0 e | 0.1 ± 0.0 bcd | 0.1 ± 0.0 ce | 0.0 ± 0.0 a | 0.0 ± 0.0 a | *** |
| C25 | SH | β-elemene | 1398 | 1.1 ± 0.0 d | 1.2 ± 0.0 cd | 1.5 ± 0.0 f | 1.0 ± 0.0 e | 1.4 ± 0.1 a | 1.2 ± 0.0 c | 1.5 ± 0.0 ef | 1.3 ± 0.0 b | *** |
| C26 | SH | β-caryophyllene | 1435 | 8.8 ± 0.1e | 10.7 ± 0.2d | 10.8 ± 0.5f | 13.2 ± 0.9f | 24.9 ± 1.0c | 17.2 ± 0.6b | 25.4 ± 0.8b | 21.7 ± 0.1a | *** |
| C27 | SH | β-cubebene | 1444 | 0.8 ± 0.1 b | 0.8 ± 0.0 ab | 0.4 ± 0.1 ab | 0.8 ± 0.0 ab | 0.9 ± 0.1 ab | 0.9 ± 0.1 ab | 0.8 ± 0.1 a | 1.6 ± 0.9 ab | |
| C28 | Oth | isoamyl benzoate | 1455 | 0.8 ± 0.1 ab | 0.7 ± 0.1 ab | 1.0 ± 0.1 a | 0.6 ± 0.1 b | 0.8 ± 0.1 ab | 0.6 ± 0.1 ab | 0.5 ± 0.1 c | 0.6 ± 0.1 b | *** |
| C29 | SH | α-caryophyllene | 1472 | 2.1± 0.0 f | 2.0 ± 0.0 e | 2.3 ± 0.1 h | 1.7 ± 0.1 g | 3.3 ± 0.0 a | 2.5 ± 0.1 b | 3.5 ± 0.0 d | 2.8 ± 0.0 c | *** |
| C30 | SH | allo-aromadendrene | 1477 | 0.8 ± 0.0 a | 0.7 ± 0.0 b | 0.7± 0.0 b | 0.5 ± 0.0 b | 0.6 ± 0.0 a | 0.7 ± 0.0 c | 0.6 ± 0.0 c | 0.4 ± 0.0 d | *** |
| C31 | SH | ɣ-muurolene | 1489 | 2.6 ± 0.0 a | 1.9 ± 0.0 d | 2.8 ± 0.1 c | 1.2 ± 0.1 c | 1.4 ± 0.1 b | 1.7 ± 0.1 e | 1.4 ± 0.0 g | 1.0 ± 0.0 f | *** |
| C32 | SH | germacrene D | 1498 | 2.5 ± 0.6 bc | 2.8 ± 0.1 d | 1.2 ± 0.3 d | 4.0 ± 0.1 d | 4.2 ± 0.3 cd | 4.2 ± 0.3 b | 4.2 ± 0.5 a | 3.2 ± 0.2 b | *** |
| C33 | SH | α-muurolene | 1514 | 1.9 ± 0.2 b | 1.7 ± 0.0 bc | 2.5 ± 0.0 cd | 1.2 ± 0.0 d | 1.8 ± 0.1 a | 1.6 ± 0.0 cd | 1.7 ± 0.1 e | 1.4 ± 0.0 d | *** |
| C34 | SH | β-bisabolene | 1520 | 1.1 ± 0.0 a | 0.8 ± 0.0 a | 0.4 ± 0.0 a | 0.8 ± 0.4 a | 0.4 ± 0.4 a | 0.8 ± 0.2 a | 0.6 ± 0.0 a | 0.6 ± 0.0 a | * |
| C35 | SH | ɣ-cadinene | 1530 | 0.6 ± 0.1 b | 0.8 ± 0.1 b | 0.9 ± 0.0 b | 0.4 ± 0.0 c | 1.1 ± 0.0 a | 0.8 ± 0.0 b | 0.7 ± 0.1 c | 0.6 ± 0.0 b | *** |
| C36 | SH | δ-cadinene | 1534 | 8.5 ± 0.0a | 4.7 ± 0.1b | 3.6 ± 0.4b | 3.0 ± 0.3b | 4.8 ± 0.2a | 4.9 ± 0.1b | 4.6 ± 0.3a | 3.3 ± 0.1c | *** |
| C37 | SH | calamenene | 1540 | 2.3 ± 0.0 a | 1.6 ± 0.2 c | 1.5 ± 0.0 b | 0.7 ± 0.0 b | 0.9 ± 0.0 a | 1.0 ± 0.0 d | 0.9 ± 0.0 d | 0.6 ± 0.0 e | *** |
| C38 | OS | elemol | 1565 | 0.8 ± 0.0 c | 0.2 ± 0.0 f | 0.5 ± 0.0 a | 0.5 ± 0.0 d | 0.4 ± 0.0 e | 0.6 ± 0.0 b | 0.2 ± 0.0 e | 0.3 ± 0.0 g | *** |
| C39 | OS | caryophyllene oxide | 1573 | 8.7 ± 0.2c | 5.7 ± 1.2c | 7.9 ± 0.7bc | 4.5 ± 0.4d | 9.9 ± 0.5a | 7.8 ± 0.1ab | 7.5 ± 0.6c | 7.8 ± 1.0cd | ** |
| C40 | OS | humulene epoxide II | 1633 | 0.9 ± 0.1 bc | 0.6 ± 0.1 df | 0.7 ± 0.1 b | 0.2 ± 0.1 ef | 0.9 ± 0.1 a | 0.8 ± 0.1 bd | 0.5 ± 0.1 cde | 0.5 ± 0.1 f | *** |
| C41 | OS | cubenol | 1648 | 3.0 ± 0.1 a | 1.2 ± 0.0 d | 2.0 ± 0.1 cd | 0.7 ± 0.1 b | 1.0 ± 0.1 a | 1.4 ± 0.0 bc | 1.2 ± 0.1 e | 0.8 ± 0.0 f | *** |
| C42 | OS | epi-Cadinol | 1663 | 2.7 ± 0.0 c | 1.8 ± 0.0 cd | 2.8 ± 0.1 de | 1.3 ± 0.1 e | 2.8 ± 0.1 a | 2.5 ± 0.1 b | 2.7 ± 0.1 e | 2.3 ± 0.0 de | *** |
| C43 | OS | δ-cadinol | 1668 | 2.3 ± 0.1 b | 1.1 ± 0.1 cd | 1.3 ± 0.2 cd | 0.6 ± 0.0 d | 1.5 ± 0.1 a | 1.4 ± 0.1 bc | 1.2 ± 0.1 cd | 0.9 ± 0.2 e | *** |
| C44 | OS | α-cadinol | 1679 | 8.0 ± 0.1cd | 5.0 ± 0.9bc | 8.0 ± 0.2bc | 3.7 ± 0.3d | 11.9 ± 0.4a | 7.9 ± 0.1ab | 7.6 ± 0.8bc | 9.5 ± 3.2bc | ** |
| C45 | OS | aromadendrene oxide | 1684 | 1.0 ± 0.1 a | 0.6± 0.0 a | 1.1 ± 0.4 a | 0.3 ± 0.0 a | 0.5 ± 0.0 a | 0.5 ± 0.0 a | 0.4 ± 0.0 b | 0.3 ± 0.0 b | ** |
| C46 | OS | bisabolol | 1704 | 9.7 ± 0.3ab | 2.6 ± 0.0b | 1.2 ± 0.0a | 1.4 ± 0.1ab | 1.3 ± 0.1ab | 1.5 ± 0.0c | 1.1 ± 0.1ab | 1.2 ± 0.0d | *** |
| C47 | Oth | benzyl benzoate | 1797 | 0.4 ± 0.0 ab | 0.2 ± 0.0 c | 0.5 ± 0.0 b | 0.3 ± 0.1 c | 0.8 ± 0.2 ab | 0.9 ± 0.0 a | 0.4 ± 0.0 b | 0.4 ± 0.0 ab | *** |
| Total (sum of C1-47) | 88.0 ± 0.6 | 91.1 ± 1.4 | 94.8 ± 0.6 | 94.0 ± 0.3 | 90.9 ± 0.8 | 90.9 ± 0.4 | 94.9 ± 0.3 | 96.6 ± 3.0 | ||||
| Terpene Classes | ||||||||||||
| MH | monoterpene hydrocarbons | 5.8 ± 0.3 c | 28.0 ± 0.1 b | 28.2 ± 0.3 b | 42.8 ± 2.6 a | 6.4 ± 1.1 d | 19.1 ± 1.0 c | 17.4 ± 1.5 c | 27.1 ± 1.2 a | *** | ||
| SH | sesquiterpene hydrocarbons | 33.8 ± 0.5 c | 30.5 ± 0.6 c | 29.8 ± 1.3 d | 29.1 ± 1.4 d | 45.9 ± 0.9 a | 37.9 ± 0.9 a | 46.5 ± 1.7 a | 38.3 ± 0.4 b | *** | ||
| OM | oxygenated monoterpenes | 8 ± 0.3 a | 9.3 ± 0.1 a | 7.3 ± 0.5 a | 3.8 ± 0.2 a | 4.6 ± 0.4 a | 4.9 ± 0.2 a | 4.3 ± 0.3 a | 3.4 ± 0.1 a | |||
| OS | oxygenated Sesquiterpenes | 37.0 ± 0.5 bc | 18.8 ± 1.1 c | 25.4 ± 0.6 bc | 13.2 ± 1.0 d | 30.3 ± 0.9 a | 24.3 ± 0.4 b | 22.4 ± 0.9 c | 23.6 ± 4.4 e | *** | ||
| Oth | Others | 3.3 ± 0.1 d | 4.5 ± 0.2 cd | 4.1 ± 0.2 bc | 5.1 ± 0.0 ab | 3.7 ± 0.5 d | 4.8 ± 0.1 cd | 4.3 ± 0.1 bc | 5.1 ± 0.1 a | |||
| Antioxidant Assays | ||||
|---|---|---|---|---|
| Essential Oil Samples | FRAP (mg TE/g EO) | ABTS (mg TE/g EO) | DPPH (mgTE/g EO) | |
| Mountain | Winter | 11.3 ± 0.1 c | 0.28 ± 0.01 b | 0.12 ± 0.0 a |
| Spring | 9.6 ± 0.3 a | 0.32 ± 0.0 c | 0.44 ± 0.01 a | |
| Summer | 22.3 ± 0.2 c | 0.09 ± 0.01 e | 0.08 ± 0.01 b | |
| Autumn | 14.3 ± 0.1 a | 0.24 ± 0.01 a | 0.11 ± 0.0 a | |
| Littoral | Winter | 6.4 ± 0.9 c | 0.09 ± 0 c | 0.06 ± 0.0 c |
| Spring | 6.6 ± 1.1 b | 0.23 ± 0.01 e | 0.06 ± 0.0 d | |
| Summer | 15.7 ± 0.5 d | 0.30 ± 0.01 a | 0.08 ± 0.0 b | |
| Autumn | 14.8 ± 0.4 b | 0.13 ± 0.01 d | 0.06 ± 0.0 c | |
| Altitude * Season Interaction | *** | *** | *** | |
| Vitamin C (mg TE/g vit C) | 1080 ± 144 | 894 ± 88 | 1340 ± 192 | |
| Major Compounds | FRAP | ABTS | DPPH |
|---|---|---|---|
| α-Pinene | 0.37 | 0.09 | 0.39 |
| ß-Pinene | 0.62 ** | −0.09 | 0.17 |
| β-Myrcene | 0.07 | 0.02 | −0.33 |
| Bornyl acetate | −0.18 | 0.27 | 0.56 ** |
| β-Caryophyllene | −0.07 | −0.18 | −0.67 *** |
| δ-Cadinene | −0.54 ** | 0.33 | 0.15 |
| Caryophyllene oxide | −0.24 | −0.45 * | −0.46 * |
| α-Cadinol | −0.11 | −0.54 ** | −0.56 ** |
| Bisabolol | −0.63 *** | 0.36 | 0.43 |
| Oxygenated monoterpenes | −0.21 | 0.30 | 0.55 ** |
| Oxygenated sesquiterpenes | −0.17 | −0.40 | −0.27 |
| Mountain (Site M) | Littoral (Site L) | Altitude * Season Interaction | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Fatty Acid | Corresponding acyl | Winter | Spring | Summer | Autumn | Winter | Spring | Summer | Autumn | |
| Methyl Ester (ME) | ||||||||||
| Myristic acid ME | C14:0 | 6.6 ± 1.7 ac | 4.0 ± 0.2 bc | 1.8 ± 0.1 ac | 3.4 ± 0.1 ab | 2.5 ± 0.1 bc | 3.2 ± 0.1 c | 2.7 ± 0.3 a | 2.7 ± 0.5 d | *** |
| Hexadecenoic acid ME | C16:1 | 1.2 ± 0.3 c | 0.9 ± 0.1 b | 1.2 ± 0.1 bc | 1.1 ± 0.1 a | 0.5 ± 0.1 b | 1.1 ± 0.1 ab | 1.3 ± 0.1 bc | 1.5 ± 0.2 bc | *** |
| Palmitic acid ME | C16:0 | 24.8 ± 2.3 c | 28.3 ± 0.3 c | 27.4 ± 0.5 c | 30.3 ± 0.9 a | 26.3 ± 1.1 bc | 31.5 ± 0.4 ac | 31.4 ± 1.0 ab | 31.7 ± 2.0 a | |
| Margaric acid ME | C17:0 | 0.7 ± 0.1 c | 0.8 ± 0.1 b | 0.8 ± 0.1 bc | 0.9 ± 0.1 a | 0.6 ± 0.1 b | 0.9 ± 0.1 ab | 0.9 ± 0.1 ab | 1.1 ± 0.1 ab | ** |
| Linoleic acid ME | C18:2 | 20.1 ± 1.7 ab | 20.0 ± 0.2 c | 28.2 ± 8.7 a | 13.9 ± 0.6 c | 26.2 ± 0.9 ab | 22.7 ± 0.5 bc | 9.7 ± 0.2 c | 14.1 ± 1.2 bc | *** |
| Oleic acid ME | C18:1 | 2.8 ± 0.2 c | 5.7 ± 0.4 bc | 15.3 ± 8.3 d | 10.8 ± 0.3 c | 14.4 ± 0.3 bc | 10.4 ± 0.6 ab | 30.2 ± 1.2 c | 15.3 ± 1.5 a | ** |
| Linolenic acid ME | C18:3 | 35.5 ± 1.8 d | 34.0 ±1.2 bc | 18.5 ± 0.2 a | 31.8 ± 1.1 c | 23.4 ± 0.2 e | 21.7 ± 0.6 e | 17.6 ± 0.7 ab | 27.2 ± 2.8 e | *** |
| Stearic acid ME | C18:0 | 1.9 ± 0.2 bc | 2.3 ± 0.1 ab | 2.6 ± 0.1 c | 2.6 ± 0.1 a | 2.1 ± 0.1 bc | 2.2 ± 0.2 ab | 2.8 ± 0.1 bc | 2.6 ± 0.3 a | |
| Docosanoic acid ME | C22:0 | 0.9 ± 0.1 ac | 1.2 ± 0.1 c | 1.1 ± 0.1 bc | 1.2 ± 0.1 a | 0.9 ± 0.1 bc | 1.3 ± 0.1 c | 1.2 ± 0.2 ac | 1.2 ± 0.2 ab | |
| Total identified | 94.3 ± 3.0 | 96.7 ± 0.7 | 96.8 ± 0.7 | 95.9 ± 0.6 | 96.9 ± 0.3 | 94.8 ± 0.9 | 97.8 ± 0.2 | 97.3 ± 0.3 | ||
| C14; C16; C17; C18; C22 | Mean (ΣSFA) | 34.8 ± 1.0 d | 36.6 ± 0.4 d | 33.6 ± 0.7 d | 38.3 ± 1.1 a | 32.4 ± 1.2 cd | 39.0 ± 0.7 bd | 39.0 ± 1.5 ab | 39.6 ± 2.9 abc | ** |
| C16:1; C18:1 | Mean (ΣMUFA) | 3.9 ± 0.5 c | 6.6 ± 0.4 ac | 16.5 ± 8.3 d | 11.9 ± 0.4 bc | 14.9 ± 0.2 ac | 11.5 ± 0.5 ab | 31.5 ± 1.2 c | 16.8 ± 1.7 a | * |
| C18:2; C18:3 | Mean (ΣPUFA) | 55.5 ± 2.7 b | 54.0 ±1.1 bc | 46.7 ± 8.6 a | 45.7± 0.5 bd | 49.6 ± 0.9 bd | 44.3 ± 1.1 cd | 27.3 ± 0.5 bd | 41.2 ± 1.8 d | ** |
| Compounds | C18:3 | C18:2 | C18:1 | C16:0 | C14:0 |
|---|---|---|---|---|---|
| β-Caryophyllene | −0.58 * | −0.19 | 0.77 *** | 0.48 * | 0.31 |
| β-Myrcene | 0.15 | −0.55 *** | 0.49 * | 0.76 *** | 0.08 |
| α-Pinene | 0.10 | −0.08 | −0.18 | 0.21 | 0.009 |
| Bornyl acetate | 0.29 | 0.44 * | −0.57 * | −0.68 ** | 0.3 |
| δ-Cadinene | 0.15 | 0.36 | −0.42 * | −0.34 | 0.39 |
| Bisabolol | 0.81 *** | 0.29 | −0.83 *** | −0.37 | 0.80 *** |
| ß-Pinene | −0.26 * | −0.10 | 0.18 | 0.21 | −0.41 * |
| Caryophyllene oxide | −0.14 | 0.40 | 0.09 | −0.46 * | −0.32 |
| α-Cadinol | −0.30 | 0.34 | 0.21 | −0.33 | −0.48 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sehaki, C.; Jullian, N.; Choque, E.; Dauwe, R.; Fontaine, J.X.; Molinie, R.; Ayati, F.; Fernane, F.; Gontier, E. Profiling of Essential Oils from the Leaves of Pistacia lentiscus Collected in the Algerian Region of Tizi-Ouzou: Evidence of Chemical Variations Associated with Climatic Contrasts between Littoral and Mountain Samples. Molecules 2022, 27, 4148. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27134148
Sehaki C, Jullian N, Choque E, Dauwe R, Fontaine JX, Molinie R, Ayati F, Fernane F, Gontier E. Profiling of Essential Oils from the Leaves of Pistacia lentiscus Collected in the Algerian Region of Tizi-Ouzou: Evidence of Chemical Variations Associated with Climatic Contrasts between Littoral and Mountain Samples. Molecules. 2022; 27(13):4148. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27134148
Chicago/Turabian StyleSehaki, Chabha, Nathalie Jullian, Elodie Choque, Rebecca Dauwe, Jean Xavier Fontaine, Roland Molinie, Fadila Ayati, Farida Fernane, and Eric Gontier. 2022. "Profiling of Essential Oils from the Leaves of Pistacia lentiscus Collected in the Algerian Region of Tizi-Ouzou: Evidence of Chemical Variations Associated with Climatic Contrasts between Littoral and Mountain Samples" Molecules 27, no. 13: 4148. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27134148