
Production and Purification of Pectinase from Bacillus subtilis 15A-B92 and Its Biotechnological Applications


 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Screening and Identification of Pectinase-Producing Bacterium
2.2. Production of Bacterial Pectinase
2.3. Measuring Enzyme Activity and Protein Content
2.4. Bacterial Pectinase Purification
2.5. Physico-Chemical Properties of Bacterial Pectinase
2.6. Application of Bacterial Pectinase in Apple and Orange Juice Clarification
2.7. Statistical Analysis
3. Results and Discussion
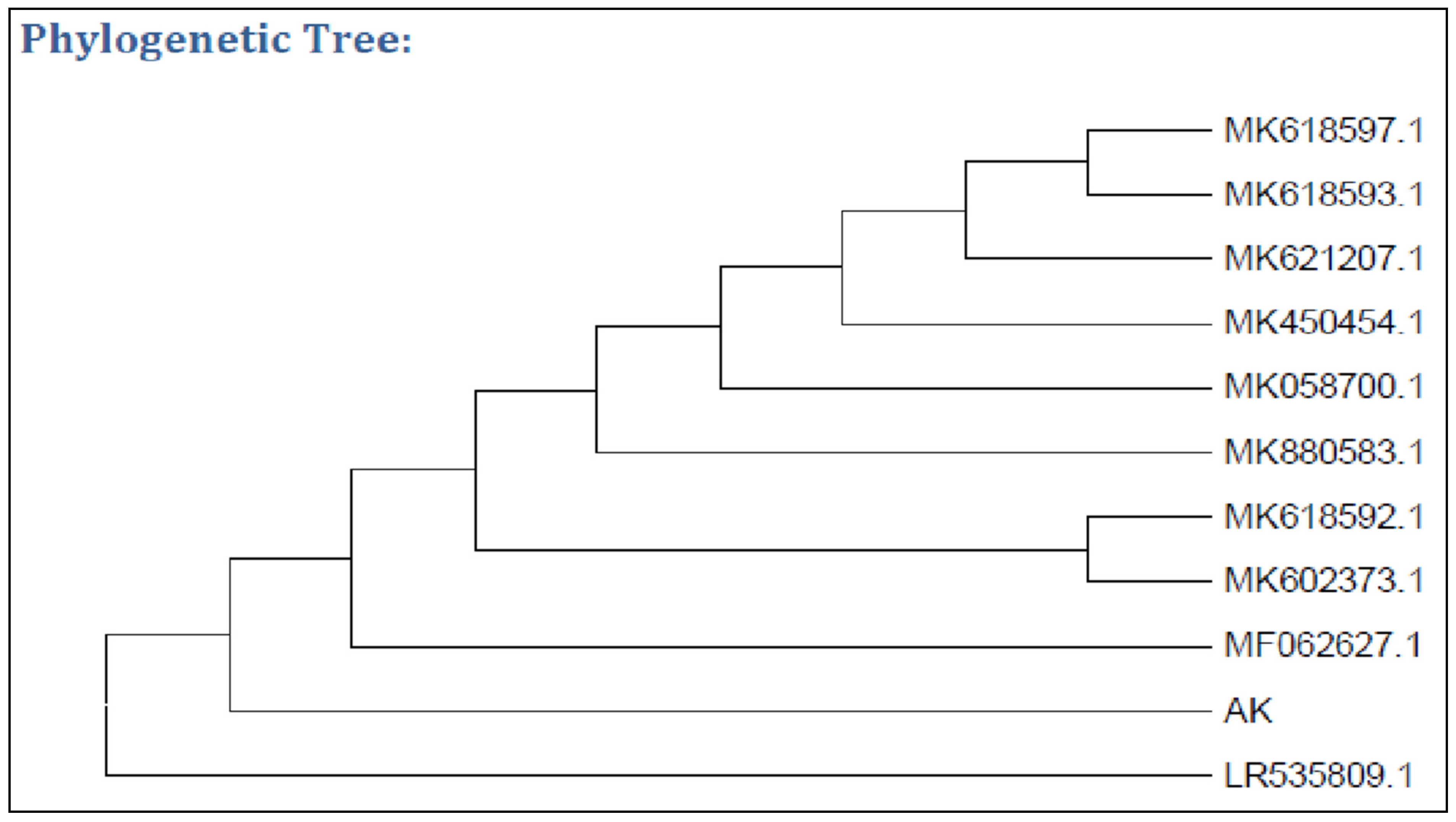
3.1. Isolation and Identification of Pectinase-Producing Bacteria
3.2. Production of Bacterial Pectinase
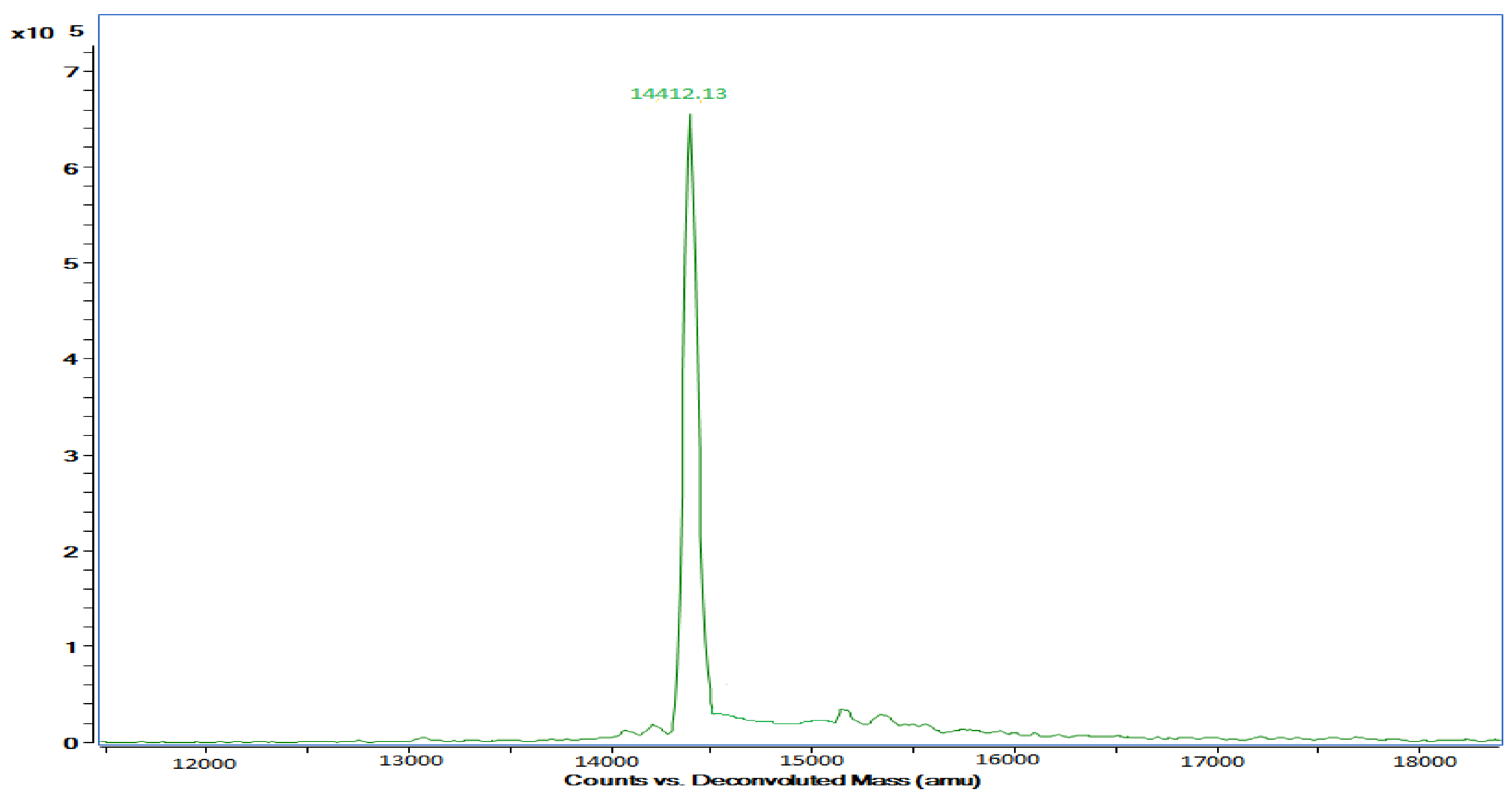
3.3. Pectinase Purification
3.4. Physico-Chemical Properties of Bacterial Pectinase
3.4.1. Effect of Temperature on the Activity and Stability of Pectinase from Bacillus subtilis 15A-B92
3.4.2. Effect of pH on the Activity and Stability of Pectinase from Bacillus subtilis 15A-B92
3.4.3. Effect of Divalent Cations on the Activity of Pectinase from Bacillus subtilis 15A-B92
3.4.4. Effect of Inhibitors and Surfactants on Bacterial Pectinase
3.4.5. Substrate Specificity and Kinetic Determinations of Bacterial Pectinase
3.5. Application of Bacterial Pectinase in Apple and Orange Juice Clarification
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Pedrolli, D.B.; Monteiro, A.C.; Gomes, E.; Carmona, E.C. Pectin and pectinases: Production, characterization and industrial application of microbial pectinolytic enzymes. Open Biotechnol. J. 2009, 3, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Yadav, P.K.; Singh, V.K.; Yadav, S.; Yadav, K.D.S.; Yadav, D. In silico analysis of pectin lyase and pectinase sequences. Biochem. Mosc. 2009, 74, 1049–1055. [Google Scholar] [CrossRef] [PubMed]
- Jayani, R.S.; Saxena, S.; Gupta, R. Microbial pectinolytic enzymes: A review. Process Biochem. 2005, 40, 2931–2944. [Google Scholar] [CrossRef]
- Chaudhri, A.; Suneetha, V. Microbially derived pectinases: A review. J. Pharm. Biol. Sci. 2012, 2, 1–5. [Google Scholar] [CrossRef]
- Kashyap, D.R.; Chandra, S.; Kaul, A.; Tewari, R. Production, purifcation and characterization of pectinase from a Bacillus sp. DT7. World J. Microbiol. Biotechnol. 2000, 16, 277–282. [Google Scholar] [CrossRef]
- Phutela, U.; Dhuna, V.; Sandhu, S.; Chadha, B.S. Pectinase and polygalacturonase production by a thermophilic Aspergillus fumigatus isolated from decomposting orange peels. Braz. J. Microbiol. 2005, 36, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Laha, S.; Sarkar, D.; Chaki, S. Optimization of production and molecular characterization of pectinase enzyme produced from Penicillium chrysogenum. Sch. Acad. J. Biosci. 2014, 2, 326–335. [Google Scholar]
- Kaur, S.; Kaur, H.P.; Prasad, B.; Bharti, T. Production and optimization of pectinase by Bacillus sp. isolated from vegetable waste soil. Indo Am. J. Pharm. Res. 2016, 6, 4185–4190. [Google Scholar]
- Bai, Z.H.; Zhang, H.X.; Qi, H.Y.; Peng, X.W.; Li, B.J. Pectinase production by Aspergillus niger using wastewater in solid state fermentation for eliciting plant disease resistance. Bioresour. Technol. 2004, 95, 49–52. [Google Scholar] [CrossRef]
- Silva, D.; Martins, E.S.; da Silva, R.; Gomes, E. Pectinase production by Penicillium viridicatum RFC3 by solid state fermentation using agricultural wastes and agro-industrial by-products. Braz. J. Microbiol. 2002, 33, 318–324. [Google Scholar] [CrossRef] [Green Version]
- Arotupin, D.J.; Akinyosoye, F.A.; Onifade, A.K. Purification and characterization of pectinmethylesterase from Aspergillus repens isolated from cultivated soil. Afr. J. Biotechnol. 2008, 7, 1991–1998. [Google Scholar] [CrossRef] [Green Version]
- Nadaroglu, H.; Taskın, E.; Adıguzel, A.; Gulluce, M.; Demir, N. Production of a novel pectin lyase from Bacillus pumilus (P9), purification and characterization and fruit juice application. Rom. Biotech. Lett. 2010, 15, 5167–5176. [Google Scholar]
- Zu-ming, L.I.; Bo, J.I.N.; Hong-xun, Z. Purification and characterization of three alkaline endopolygalacturonases from a newly isolated Bacillus gibsonii. Chin. J. Process. Eng. 2008, 8, 4–9. [Google Scholar]
- Amid, M.; Manap, Y.; Zohdi, K. Purifcation and characterization of thermo-alkaline pectinase enzyme from Hylocereus polyrhizus. Eur. Food Res. Technol. 2014, 239, 21–29. [Google Scholar] [CrossRef]
- Celestino, S.M.; de Freitas, S.M.; Medrano, F.J.; de Souza, M.V.; Filho, E.X. Purification and characterization of a novel pectinase from Acrophialophora nainiana with emphasis on its physicochemical properties. J. Biotechnol. 2006, 123, 33–42. [Google Scholar] [CrossRef]
- Oumer, O.J.; Abate, D. Characterization of pectinase from Bacillus subtilis strain Btk 27 and its potential application in removal of mucilage from coffee beans. Enzyme Res. 2017, 2017, 7686904. [Google Scholar] [CrossRef] [Green Version]
- Sandri, I.G.; Lorenzoni, C.M.T.; Fontana, R.C.; Silveira, M.M.D. Use of pectinases produced by a new strain of Aspergillus niger for the enzymatic treatment of apple and blueberry juice. LWT Food Sci. Technol. 2013, 51, 469–475. [Google Scholar] [CrossRef]
- Wang, B.F.; Cheng, F.; Lu, Y.; Ge, W.; Zhang, M.; Yue, B. Immobilization of pectinase from Penicillium oxalicum F67 onto magnetic cornstarch microspheres: Characterization and application in juice production. J. Mol. Catal. B Enzym. 2013, 97, 137–143. [Google Scholar] [CrossRef]
- Bernfeld, P. Amylases, alpha and beta. In Methods in Enzymology; Academic Press: New York, NY, USA, 1955; Volume 1, pp. 149–154. [Google Scholar]
- Lowry, O.H.; Rosenberg, W.J.; Farr, A.L.; Randell, R.J. Quantitation of protein using Folin Ciocalteu reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Spivak, J.L.; Small, D.; Hollenberg, M.D. Erythropoietin: Isolation by affinity chromatography withlectin-agarose derivatives. Proc. Natl. Acad. Sci. USA 1977, 74, 4633–4635. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Blum, H.; Beier, H.; Gross, H.J. Improved staining of plant proteins, RNA and DNA in polyacrylamide gel. Electrophoresis 1987, 8, 93–99. [Google Scholar] [CrossRef]
- Niyonzima, F.N.; More, S.S. Biochemical properties of the alkaline lipase of Bacillus flexus XJU-1 and its detergent compatibility. Biologia 2014, 69, 1108–1117. [Google Scholar] [CrossRef]
- Kumar, G.P.; Suneetha, V. Microbial pectinases: Wonderful enzymes in fruit juice clarification. Int. J. MediPharm. Res. 2016, 2, 119–127. [Google Scholar]
- Logan, N.A.; De Vos, P. Bacillus. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; De Vos, P., Garrity, G., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.-H., Whitman, W.B., Eds.; Springer: New York, NY, USA, 2009; Volume 3, pp. 21–127. [Google Scholar]
- Kareem, S.O.; Adebowale, A.A. Clarification of orange juice by crude fungal pectinase from citrus peel. Niger. Food J. 2007, 25, 130–137. [Google Scholar] [CrossRef] [Green Version]
- Maciel, M.D.H.C.; Ottoni, C.A.; Herculano, P.N.; Porto, T.S.; Porto, A.L.F.; Santos, C.; Lima, N.; Moreira, K.A.; Souza-Motta, C. Purification of polygalacturonases produced by Aspergillus niger using an aqueous two-phase system. Fluid Phase Equilibria 2014, 371, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Kusuma, M.P.; Reddy, D.R. Purification and characterization of polygalacturonase using isolated Bacillus subtilis C4. Res. J. Microbiol. 2014, 9, 95–103. [Google Scholar] [CrossRef] [Green Version]
- Niyonzima, F.N. Detergent-compatible bacterial cellulases. J. Basic Microbiol. 2018, 59, 134–147. [Google Scholar] [CrossRef]
- Alana, A.; Alkorta, I.; Dominguez, J.B.; Liama, M.J.; Serra, J.L. Pectin lyase activity in a Penicillium italicum strain. Appl. Environ. Microbiol. 1990, 56, 3755–3759. [Google Scholar] [CrossRef] [Green Version]
- Beg, Q.N.; Gupta, R. Purification and characterization of an oxidation-stable, thiol-dependent serine alkaline protease from Bacillus mojavensis. Enzym. Microb. Technol. 2003, 32, 294–304. [Google Scholar] [CrossRef]
- Li, S.; Yang, X.; Yang, S.; Zhu, M.; Wang, X. Technology prospecting on enzymes: Application, marketing and engineering. Comput. Struct. Biotechnol. J. 2012, 2, e201209017. [Google Scholar] [CrossRef] [Green Version]
- Diaz, A.B.; Alvarado, O.; De Ory, I.; Caro, I.; Blandino, A. Valorization of grape pomace and orange peels: Improved production of hydrolytic enzymes for the clarification of orange juice. Food Bioprod. Process. 2013, 91, 580–586. [Google Scholar] [CrossRef]
- Dey, T.B.; Adak, S.; Bhattacharya, P.; Banerjee, R. Purification of polygalacturonase from Aspergillus awamori Nakazawa MTCC 6652 and its application in apple juice clarification. LWT Food Sci. Technol. 2014, 59, 591–595. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Activity (U) | Total Protein (mg) | Specific Activity (U/mg) | Purification Fold-Change | Purification Yield |
|---|---|---|---|---|---|
| Crude enzyme | 4352.0 | 506.0 | 8.6 | 1 | 100 |
| Ammonium sulfate precipitation | 1602.5 | 51.9 | 30.9 | 3.6 | 36.8 |
| Affinity chromatography | 816.9 | 8.2 | 99.6 | 11.6 | 18.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqahtani, Y.S.; More, S.S.; R., K.; Shaikh, I.A.; K. J., A.; More, V.S.; Niyonzima, F.N.; Muddapur, U.M.; Khan, A.A. Production and Purification of Pectinase from Bacillus subtilis 15A-B92 and Its Biotechnological Applications. Molecules 2022, 27, 4195. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27134195
Alqahtani YS, More SS, R. K, Shaikh IA, K. J. A, More VS, Niyonzima FN, Muddapur UM, Khan AA. Production and Purification of Pectinase from Bacillus subtilis 15A-B92 and Its Biotechnological Applications. Molecules. 2022; 27(13):4195. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27134195
Chicago/Turabian StyleAlqahtani, Yahya S., Sunil S. More, Keerthana R., Ibrahim Ahmed Shaikh, Anusha K. J., Veena S. More, Francois N. Niyonzima, Uday M. Muddapur, and Aejaz A. Khan. 2022. "Production and Purification of Pectinase from Bacillus subtilis 15A-B92 and Its Biotechnological Applications" Molecules 27, no. 13: 4195. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27134195