
Bridging the Chemical Profiles and Biological Effects of Spathodea campanulata Extracts: A New Contribution on the Road from Natural Treasure to Pharmacy Shelves

 ,
,  ,
, 
 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemical Characterization of the Tested Extracts

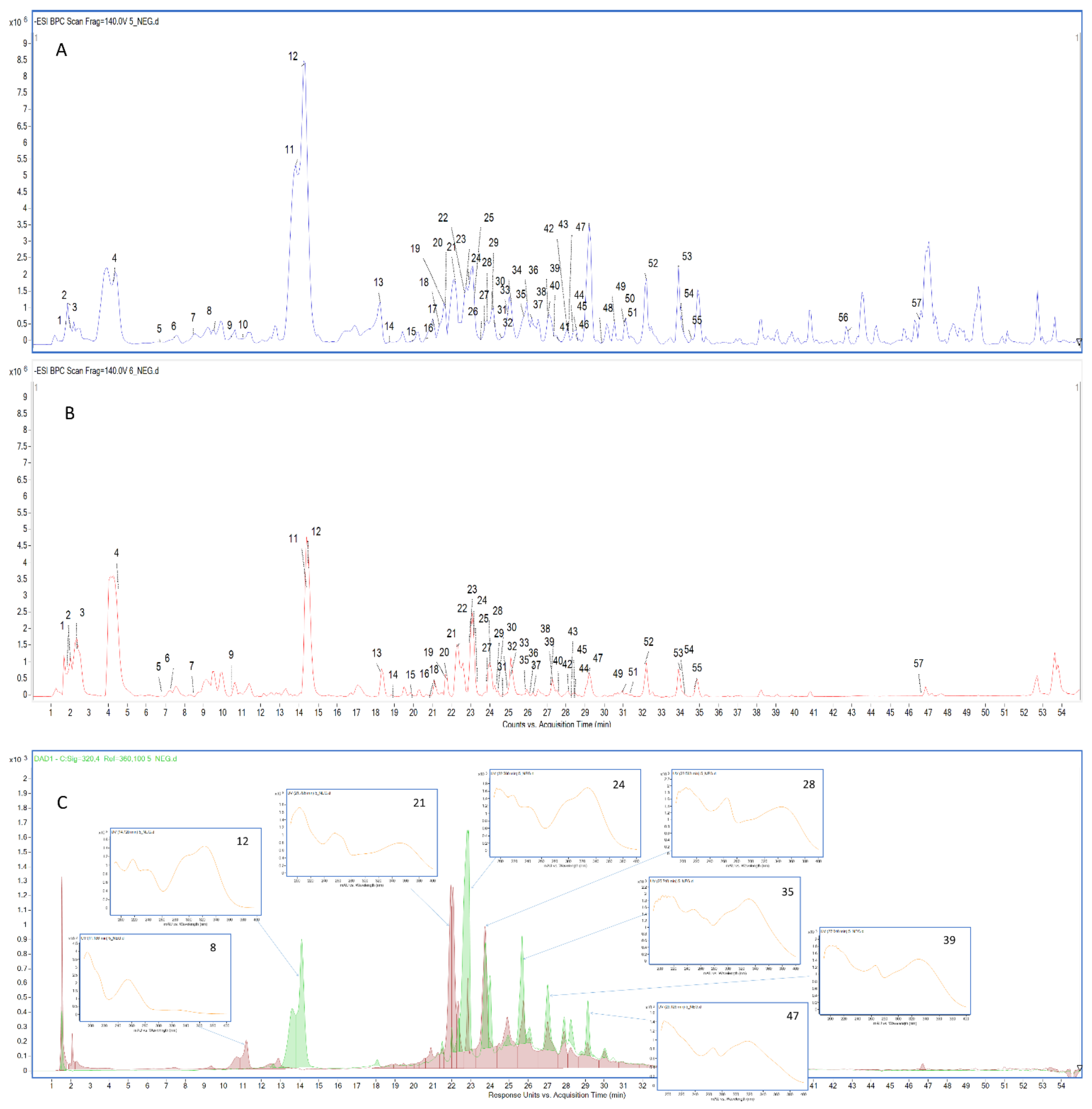{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comp. No. | Tentative Identification | Rt (min) | Molecular Formula | [M−H]− (m/z) | Product Ions (m/z) | Extracts | References |
|---|---|---|---|---|---|---|---|
| 1. | Sucrose | 1.569 | C12H22O11 | 341.1035 | 179.0530; 161.0270; 135.0455; 119.0314; 89.0226 | Leaves-MeOH Leaves-infusion | [43] |
| 2. | Malic acid | 2.049 | C4H6O5 | 133.0110 | 115.0040; 89.0235; 71.0135 | Leaves-MeOH Leaves-infusion | [44] |
| 3. | Citric acid | 2.409 | C6H8O7 | 191.0187 | 173.0064; 154.9955; 111.0084; 87.0090 | Leaves-MeOH Leaves-infusion Stem bark-infusion | [42] |
| 4. | Quinic acid | 4.206 | C7H12O6 | 191.0540 | 173.0425; 101.0591; 85.0649 | Leaves-MeOH Leaves-infusion Stem bark-infusion | [44] |
| 5. | Dihydroxybenzoic acid | 7.59 | C7H6O4 | 153.0143 | 109.0302; 108.0225; 91.0172 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [44] |
| 6. | Ajugol | 7.802 | C15H24O9 | 347.1358 | 303.1342; 123.0783; 185.0829; 167.0704 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [30] |
| 7. | Loganic acid | 9.121 | C16H24O10 | 375.1251 | 213.0758; 194.8742; 169.0859; 151.0761; 125.0603 | Leaves-MeOH Leaves-infusion | [30] |
| 8. | Hydroxybenzoic acid | 9.840 | C7H6O3 | 137.0228 | 108.0218; 109.0287 | Leaves-MeOH Stem bark-MeOH | [44] |
| 9. | Caffeoyl-glucopyranoside | 10.080 | C15H18O9 | 341.0877 | 281.0675; 179.0364; 161.0247; 133.0282; 135.0445 | Leaves-MeOH Leaves-infusion | [31] |
| 10. | Methylgallate | 11.110 | C8H8O5 | 183.0279 | 168.0064; 124.0160; 78.0117 | Leaves-MeOH | [45] |
| 11. | Iridoid compound | 13.721 | - | 459.1586 | 281.0622; 279.1173; 179.0330; 135.0459 | Leaves-MeOH Leaves-infusion | |
| 12. | Caffeic acid | 14.694 | C9H8O4 | 179.0323 | 135.0424 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [45] |
| 13. | 6-O-trans-caffeoyldecinnamoyl Globularimin (spatheoside A) | 18.698 | C24H30O14 | 541.1554 | 179.0352; 161.0242; 135.0451 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [30] |
| 14. | 6′-O-Caffeoylcatalpol | 19.189 | C24H28O13 | 523.1473 | 323.0679; 281.0643; 221.0368; 179.0359; 161.0301; 135.0421 | Leaves-MeOH Leaves-infusion | [32] |
| 15. | Derivative of spatheoside A | 20.484 | - | 657.1810 | 541.1523; 179.0358; 135.0459 | Leaves-MeOH Leaves-infusion | |
| 16. | Quercetin diglucoside | 20.927 | C27H30O17 | 625.1344 | 301.0269; 300.0225; 271.0181; 255.0193; 178.9903; 151.0008 | Leaves-MeOH Leaves-infusion | [44] |
| 17. | 6-O-trans-caffeoyl-5,7-bisdeoxycynanchoside (spatheoside C) | 21.347 | C24H30O13 | 525.1595 | 345.0935; 179.0313; 161.0217; 135.0423 | Leaves-MeOH Stem bark-MeOH Stem bark-infusion | [30] |
| 18. | Quercetin-3-O-apiosylrutinoside | 21.551 | C32H38O20 | 741.1808 | 609.1364; 591.1358; 475.0774; 343.0367; 300.0235; 271.0228; 178.9960; 150.9969 | Leaves-MeOH Leaves-infusion | [44] |
| 19. | Iridoid compound 2 | 21.651 | - | 535.1568 | 491.1581; 341.0895; 323.0786; 179.0363; 161.0256; 135.0471; 133.0299 | Leaves-MeOH Leaves-infusion | |
| 20. | 6-O-trans-caffeoyl-asystasioside E (spatheoside B) isomer 1 | 21.730 | C24H29ClO13 | 559.1241 | 523.1395; 361.0853; 179.0300; 161.0202; 135.0401 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [30] |
| 21. | Quercetin-3-O-(2-O-β-d-xylopyranosyl)-β-d-galactopyranoside | 22.186 | C26H28O16 | 595.1271 | 523.1400; 445.0642; 300.0233; 271.0206; 255.0262; 178.9963; 151.0000 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [13,30] |
| 22. | 6ʹ-O-trans-caffeoyl-loganic acid | 22.785 | C25H30O13 | 537.1563 | 323.0662; 179.0309; 161.0247 | Leaves-MeOH Leaves-infusion | [30] |
| 23. | 6-O-caffeoylcatalpol (verminoside) | 22.845 | C24H28O13 | 523.1431 | 361.0851; 343.0808; 179.0337;161.0241; 135.0458;133.0311 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [30,31] |
| 24. | 6-O-trans-caffeoyl-asystasioside E (spatheoside B) isomer 2 | 22.941 | C24H29ClO13 | 559.1268 | 523.1422; 361.0940; 179.0320; 161.0229; 135.0443; 133.0367 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [3] |
| 25. | Rutin | 23.324 | C27H30O16 | 609.1439 | 300.0225; 271.0202; 151.0040 | Leaves-MeOH Leaves-infusion | [38,46] |
| 26. | Isoquercetin | 23.742 | C21H20O12 | 463.0861 | 300.0247; 271.0164; 255.0283; 151.0001 | Leaves-MeOH | [38,46] |
| 27. | Quercetin-3-O-pentosyl-pentoside | 23.744 | C25H25O15 | 565.1154 | 300.0223; 271.0215; 178.9841; 151.0006; 116.9266 | Leaves-MeOH Leaves-infusion | [44] |
| 28. | Kaempferol 3-O-(2-O-β-d-xylopyranosyl)-β-d-galactopyranoside | 23.984 | C26H28O15 | 579.1306 | 285.0349; 284.0282; 255.0252; 178.9957; 151.0011 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [30] |
| 29. | Luteolin-O-hexoside | 23.924 | C21H20O11 | 447.0888 | 285.0359; 284.0282; 151.0022; 133.0258 | Leaves-MeOH Leaves-infusion | [36] |
| 30. | 6-O-trans-caffeoyl-asystasioside E (spatheoside B) isomer 3 | 24.163 | C24H29ClO13 | 559.1226 | 523.1405; 179.0313; 161.0212; 135.0421 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [30] |
| 31. | Kaempferol-O-rutinoside | 24.463 | C27H30O15 | 593.1475 | 284.0274; 255.0288; 150.9984 | Leaves-MeOH Leaves-infusion | [46] |
| 32. | Kaempferol-O-sophoroside-O-glucoside | 24.643 | C33H40O21 | 771.1692 | 609.1393; 285.0369; 255.0227; 150.9955; | Leaves-MeOH Leaves-infusion | [44] |
| 33. | 6-O-trans-caffeoyl-asystasioside E (spatheoside B) isomer 4 | 24.163 | C24H29ClO13 | 559.1214 | 523.1433; 361.0930; 179.0342; 161.0238; 135.0453 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [30] |
| 34. | Ferulic acid | 25.003 | C10H10O4 | 193.0473 | 178.0242; 161.0214; 149.0535; 134.0355 | Leaves-MeOH Stem bark-MeOH | [28] |
| 35. | Kaempferol-O-glucuronide | 25.123 | C21H18O12 | 461.0702 | 285.0360; 151.0008 | Leaves-MeOH Leaves-infusion | [44] |
| 36. | Quercetin-O-(pentoside-hexoside)-O- hexoside | 26.225 | C32H38O21 | 757.1526 | 595.1228; 301.0280; 300.0236; 179.9978; 161.0208; 151.0013 | Leaves-MeOH Leaves-infusion | |
| 37. | Caffeoyl dihexoside | 26.333 | C21H28O14 | 503.1141 | 341.0839; 281.0627; 251.0531; 21.0442; 179.0324; 161.0220; 135.0414 | Leaves-MeOH Leaves-infusion | [47] |
| 38. | Caffeoyl/glucosyl spatheoside A | 27.160 | C30H40O19 | 703.1859 | 541.1500; 179.0317; 161.0219; 135.0415 | Leaves-MeOH Leaves-infusion | |
| 39. | Kaempferol-O-caffeoyl-pentoside-O-hexoside | 27.190 | C35H34018 | 741.1812 | 579.1459; 455.1247; 285.0438; 184.0367; 179.0368; 161.0271; 151.0063; 135.0468 | Leaves-MeOH Leaves-infusion | |
| 40. | Trihydroxyflanon-O-glucuronide | 27.400 | C21H18O11 | 445.0733 | 269.0406 | Leaves-MeOH Leaves-infusion | [44] |
| 41. | Quercetin-O-dihexoside | 27.520 | C30H26O15 | 625.1165 | 463.0899; 301.0303; 300.0196; 271.0178; 178.9977; 150.9979 | Leaves-MeOH | [38] |
| 42. | Quercetin-O- arabinoside-glucoside-O-rhamnoside | 28.110 | C32H38O20 | 741.1812 | 595.1311; 301.0324; 300.0280; 178.9983; 151.0041 | Leaves-MeOH Leaves-infusion | |
| 43. | Quercetin-O-arabinoside-glucoside-O-glucuronide | 28.119 | C32H36O22 | 771.1725 | 595.1144; 300.0237; 271.0192; 255.0354; 178.9916; 150.9941 | Leaves-MeOH Leaves-infusion | |
| 44. | Kaempferol-O-caffeoylglucoside | 28.444 | C30H26O14 | 609.1353 | 447.0937; 285.0407; 179.0356; 161.0241; 151.0049; 135.0452 | Leaves-MeOH Leaves-infusion | [44] |
| 45. | Kaempferol-O-(caffeoylglucoside)-O-rhamnoside | 28.503 | C36H36018 | 755.1733 | 609.1363; 285.0346; 284.0283; 255.0262; 227.0300; 178.9987; 150.9968 | Leaves-MeOH Leaves-infusion | [48] |
| 46. | Di-O-caffeoylcatalpol isomer 1 | 28.766 | C33H34O16 | 685.1722 | 523.1403; 323.0718; 179.0304; 161.0213 | Leaves-MeOH | [33] |
| 47. | Kaempferol-O-(pentoside-hexoside)-O-deoxyhexoside | 29.306 | C32H38O19 | 725.1650 | 579.1321; 284.0289; 145.0291 | Leaves-MeOH Leaves-infusion | [49] |
| 48. | Di-O-caffeoylcatalpol isomer 2 | 29.997 | C33H34O16 | 685.1712 | 523.1397; 343.0764; 179.0345; 161.0159 | Leaves-MeOH | [33] |
| 49. | 6-O-(E)-caffeoylajugol | 30.552 | C24H30O12 | 509.1973 | 347.1728; 179.0338; 161.0230 | Leaves-MeOH Leaves-infusion | [31] |
| 50. | Quercetin | 31.148 | C15H10O7 | 301.0324 | 178.9964; 151.0009; 121.0308; 107.0146 | Leaves-MeOH | [43] |
| 51. | Luteolin | 31.176 | C15H10O6 | 285.0428 | 267.0399; 241.0546; 175.0375; 151.0058; 133.0311 | Leaves-MeOH Leaves-infusion | [36,38] |
| 52. | Trihydroxy-octadecadienoic acid | 32.36 | C18H32O5 | 327.2116 | 291.1989; 229.1460; 211.1336; 171.1031 | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [45] |
| 53. | Trihydroxy-octadecenoic acid | 33.91 | C18H34O5 | 329.2288 | 311.2203; 293.1239; 229.1450; 211.1334; 171.1011; | Leaves-MeOH Leaves-infusion Stem bark-MeOH Stem bark-infusion | [45] |
| 54. | Apigenin | 34.088 | C15H10O5 | 269.0417 | 227.0342; 151.0027; 117.0349; 107.0126 | Leaves-MeOH Leaves-infusion | [40,43] |
| 55. | Tetrahydroxyflavone | 34.652 | C15H10O6 | 285.0365 | 151.0018; 133.0281; 117.0336 | Leaves-MeOH Leaves-infusion | [44] |
| 56. | Spathodic acid | 43.761 | C30H48O5 | 487.3429 | 469.2759; 443.2390 | Leaves-MeOH Stem bark-MeOH Stem bark-infusion | [5,41] |
| 57. | Hydroxy octadecatrienoic acid | 47.369 | C18H30O3 | 293.2078 | 275.2015; 224.1403; 195.1388 | Leaves-MeOH Leaves-infusion Stem bark-MeOH | [38] |
2.2. Antioxidant Capacity
2.3. Enzyme Inhibitory Properties
2.4. Antineoplastic and Antiviral Ability
2.5. Molecular Docking
3. Materials and Methods
3.1. Plant Materials
3.2. Total Phenolic and Flavonoid Content
3.3. Chemical Characterization
3.4. Antioxidant and Enzyme Inhibitory Assays
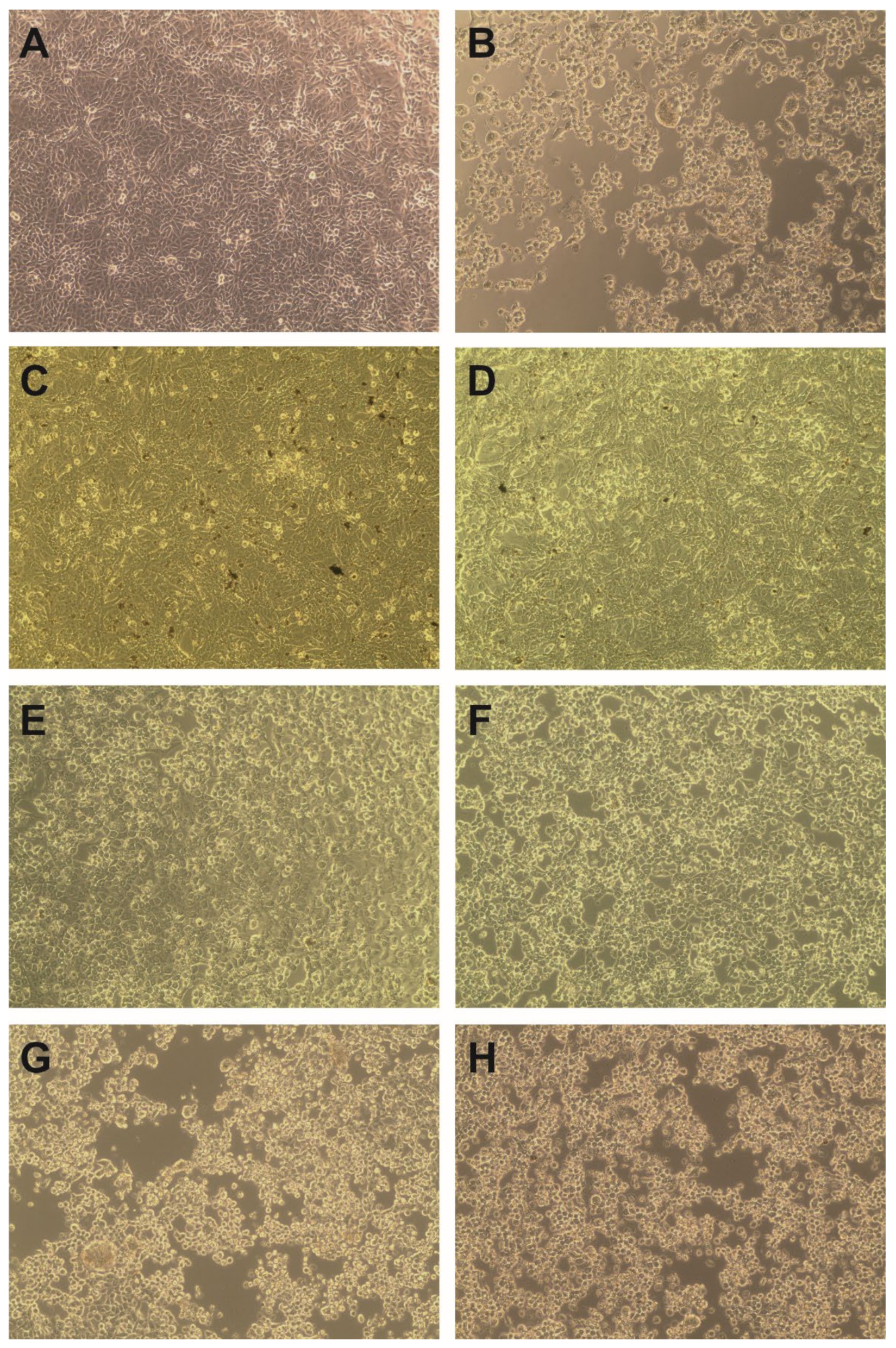
3.5. Evaluation of Cytotoxicity and Anticancer Selectivity
3.6. Evaluation of Antiviral Effects
3.7. Molecular Docking
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Mahomoodally, M.F. Traditional medicines in Africa: An appraisal of ten potent African medicinal plants. Evid.-Based Complement. Altern. Med. 2013, 2013, 617459. [Google Scholar] [CrossRef] [Green Version]
- Wagh, A.; Butle, S.; Raut, D. Isolation, identification, and cytotoxicity evaluation of phytochemicals from chloroform extract of Spathodea campanulata. Future J. Pharm. Sci. 2021, 7, 58. [Google Scholar] [CrossRef]
- Bajaj, J.; Dwivedi, J.; Sahu, R.; Dave, V.; Verma, K.; Joshi, S.; Sati, B.; Sharma, S.; Seidel, V.; Mishra, A.P. Antidepressant activity of Spathodea campanulata in mice and predictive affinity of spatheosides towards type A monoamine oxidase. Cell. Mol. Biol. 2021, 67, 1–8. [Google Scholar] [CrossRef]
- VB, A.B.; VT, A.; Thomas, B. Ethno-pharmacological potentialities of some ornamental plants in the family Bignoniaceae. J. Med. Plants 2015, 3, 135–140. [Google Scholar]
- Ngnameko, C.R.; Marchetti, L.; Zambelli, B.; Quotadamo, A.; Roncarati, D.; Bertelli, D.; Njayou, F.N.; Smith, S.I.; Moundipa, P.F.; Costi, M.P. New Insights into Bioactive Compounds from the Medicinal Plant Spathodea campanulata P. Beauv. and Their Activity against Helicobacter pylori. Antibiotics 2020, 9, 258. [Google Scholar] [CrossRef]
- Choudhury, S.; Datta, S.; Talukdar, A.D.; Choudhury, M.D. Phytochemistry of the Family Bignoniaceae—A Review. Assam Univ. J. Sci. Technol. 2011, 7, 145–150. [Google Scholar]
- Boniface, P.K. Advances on ethnomedicinal uses, phytochemistry, and pharmacology of Spathodea campanulata P. Beauv. EC Pharmacol. Toxicol. 2017, 5, 51–62. [Google Scholar]
- Vastrad, J.V.; Goudar, G. Evaluation of phenolic compounds and development of chromatographic profiles in Spathodea campanulata inflorescence by HPTLC. Asian J. Chem. 2016, 28, 497. [Google Scholar] [CrossRef]
- Kolodziej, H. Studies on Bignoniaceae: Newbouldiosides D-F, Minor Phenylethanoid Glycosides from Newbouldia laevis, and New Flavonoids from Markhamia zanzibarica and Spathodea campanulata. Planta Med. 2021, 87, 989–997. [Google Scholar] [CrossRef]
- Beg, M.; Maji, A.; Islam, M.; Hossain, M. Elucidating the interaction of Spathodea campanulata leaf extracts mediated potential bactericidal gold nanoparticles with human serum albumin: Spectroscopic analysis. J. Biomol. Struct. Dyn. 2019, 37, 3536–3549. [Google Scholar] [CrossRef]
- Kuete, V.; Tchinda, C.F.; Mambe, F.T.; Beng, V.P.; Efferth, T. Cytotoxicity of methanol extracts of 10 Cameroonian medicinal plants towards multi-factorial drug-resistant cancer cell lines. BMC Complement. Altern. Med. 2016, 16, 267. [Google Scholar] [CrossRef] [Green Version]
- Navarro, B.; Barrera, S.; Calvillo, T.; Hernández, A. Antioxidant capacity in vitro of leaves and flowers extracts of Spathodea campanulata. Planta Med. 2021, 87, 1307. [Google Scholar]
- Santos, V.H.; Minatel, I.O.; Lima, G.P.; Silva, R.M.; Chen, C.-Y.O. Antioxidant capacity and phytochemical characterization of Spathodea campanulata growing in different climatic zones in Brazil. Biocatal. Agric. Biotechnol. 2020, 24, 101536. [Google Scholar] [CrossRef]
- Villarreal, S.; Moreno, S.; Jaimez, D.; Rojas-Fermin, L.; Lucena, M.; Diaz, L.; Diaz, T.; Carmona, J. Antibacterial and Antioxidant Activity of Raw Extracts of Plants Belonging to the Bignoniaceae Family. Acta Bioclin. 2017, 7, 205–222. [Google Scholar]
- Mbosso Teinkela, J.E.; Assob Nguedia, J.C.; Meyer, F.; Vouffo Donfack, E.; Lenta Ndjakou, B.; Ngouela, S.; Tsamo, E.; Adiogo, D.; Guy Blaise Azebaze, A.; Wintjens, R. In vitro antimicrobial and anti-proliferative activities of plant extracts from Spathodea campanulata, Ficus bubu, and Carica papaya. Pharm. Biol. 2016, 54, 1086–1095. [Google Scholar] [CrossRef]
- Saranya, M.; Mohanraj, R.S.; Dhanakkodi, B. Larvicidal, pupicidal activities and morphological deformities of Spathodea campanulata aqueous leaf extract against the dengue vector Aedes aegypti. Eur. J. Exp. Biol. 2013, 3, 205–213. [Google Scholar]
- Boniface, P.K.; Verma, S.; Shukla, A.; Khan, F.; Srivastava, S.K.; Pal, A. Membrane stabilisation: A possible anti-inflammatory mechanism for the extracts and compounds from Spathodea campanulata. Nat. Prod. Res. 2014, 28, 2203–2207. [Google Scholar] [CrossRef]
- Niyonzima, G.; Laekeman, G.; Witvrouw, M.; Van Poel, B.; Pieters, L.; Paper, D.; De Clercq, E.; Franz, G.; Vlietinck, A. Hypoglycemic, anticomplement and anti-HIV activities of Spathodea campanulata stem bark. Phytomedicine 1999, 6, 45–49. [Google Scholar] [CrossRef]
- Begum, A.; Biswas, P.; Shahed-Al-Mahmud, M. Methanol extract of Spathodea campanulata P.(Beauv.) leaves demonstrate sedative and anxiolytic like actions on swiss albino mice. Clin. Phytosci. 2020, 6, 41. [Google Scholar] [CrossRef]
- Adesiyan, G.O.; Talabi, T.O.; Kale, O.E.; Ogundare, T.F. Ethyl acetate fraction of Spathodea campanulata (Bignoniaceae) attenuates lead acetate induced testicular toxicity in male Wistar rats. J. Complement. Integr. Med. 2022, 19, 19–26. [Google Scholar] [CrossRef]
- Sy, G.Y.; Nongonierma, R.B.; Ngewou, P.W.; Mengata, D.E.; Dieye, A.M.; Cisse, A.; Faye, B. Healing activity of methanolic extract of the barks of Spathodea campanulata Beauv. (Bignoniaceae) in rat experimental burn model. Dakar Med. 2005, 50, 77–81. [Google Scholar] [PubMed]
- Ofori-Kwakye, K.; Kwapong, A.A.; Bayor, M.T. Wound healing potential of methanol extract of Spathodea campanulata stem bark forumlated into a topical preparation. Afr. J. Tradit. Complement. Altern. Med. 2011, 8, 218–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ilodigwe, E.E.; Akah, P.A.; Nworu, C.S. Anticonvulsant activity of ethanol leaf extract of Spathodea campanulata P. Beauv. (Bignoniaceae). J. Med. Food 2010, 13, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Sangeetha, S.; Meenakshi, S.; Akshaya, S.; Vadivel, V.; Brindha, P. Evaluation of total phenolic content and antioxidant activity of different solvent extracts of leaf material of Spathodea campanulata P. Beauv. and investigation of their proliferation inhibition potential against EAC cell line. J. Appl. Pharm. Sci. 2016, 6, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Diane, K.; Oumar, Y.; Idrissa, D.; Felix, Y.; David, N.; Joseph, D. Evaluation of the Antioxidant Activity and Dosage of Polyphenols in Aqueous, Hydroethanolic and Hexane Extracts of the Bark of Spathodea campanulata P. Beauv. (Bignoniaceae). Int. J. Curr. Microbiol. App. Sci. 2021, 10, 154–163. [Google Scholar]
- Bibi Sadeer, N.; Montesano, D.; Albrizio, S.; Zengin, G.; Mahomoodally, M.F. The versatility of antioxidant assays in food science and safety—Chemistry, applications, strengths, and limitations. Antioxidants 2020, 9, 709. [Google Scholar] [CrossRef]
- Hossain, M.B.; Rai, D.K.; Brunton, N.P.; Martin-Diana, A.B.; Barry-Ryan, C. Characterization of phenolic composition in Lamiaceae spices by LC-ESI-MS/MS. J. Agric. Food Chem. 2010, 58, 10576–10581. [Google Scholar] [CrossRef]
- Łaska, G.; Maciejewska-Turska, M.; Sieniawska, E.; Świątek, Ł.; Pasco, D.S.; Balachandran, P. Extracts from Pulsatilla patens target cancer-related signaling pathways in HeLa cells. Sci. Rep. 2021, 11, 10654. [Google Scholar] [CrossRef]
- Padhy, G.K. Spathodea campanulata P. Beauv.—A review of its ethnomedicinal, phytochemical, and pharmacological profile. J. Appl. Pharm. Sci. 2021, 11, 17–44. [Google Scholar]
- Gouda, Y.G. Iridoids from Spathodea campanulata P. Beauvais leaves. Nat. Prod. Commun. 2009, 4, 1934578X0900400602. [Google Scholar] [CrossRef] [Green Version]
- Li, X.-N.; Sun, J.; Shi, H.; Yu, L.L.; Ridge, C.D.; Mazzola, E.P.; Okunji, C.; Iwu, M.M.; Michel, T.K.; Chen, P. Profiling hydroxycinnamic acid glycosides, iridoid glycosides, and phenylethanoid glycosides in baobab fruit pulp (Adansonia digitata). Food Res. Int. 2017, 99, 755–761. [Google Scholar] [CrossRef]
- Li, C.; Liu, Y.; Abdulla, R.; Aisa, H.A.; Suo, Y. Characterization and identification of chemical components in Neopicrorhiza scrphulariiflora roots by liquid chromatography-electrospray ionization quadrupole time-of-flight tandem mass spectrometry. Anal. Methods 2014, 6, 3634–3643. [Google Scholar] [CrossRef]
- Sahakitpichan, P.; Chimnoi, N.; Srinroch, C.; Petchthong, C.; Ruchirawat, S.; Kanchanapoom, T. A New Iridoid Glycoside from Santisukia pageth. Heterocycles 2022, 104, 548–554. [Google Scholar]
- Wagh, A.S.; Butle, S.R. Plant profile, phytochemistry and pharmacology of Spathodea campanulata P. Beauvais (African Tulip Tree): A review. Int. J. Pharm. Pharm. Sci. 2018, 10, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Grabarics, M.; Lettow, M.; Kirschbaum, C.; Greis, K.; Manz, C.; Pagel, K. Mass spectrometry-based techniques to elucidate the sugar code. Chem. Rev. 2021, 122, 7840–7908. [Google Scholar] [CrossRef]
- Wang, M.; Cao, A.; Ouyang, C.; Li, Y.; Wei, Y. Rapid screening and identification of non-target flavonoid components in invasive weeds by LC/MS-IT-TOF. Anal. Methods 2015, 7, 10207–10216. [Google Scholar] [CrossRef]
- Maciejewska-Turska, M.; Zgórka, G. In-depth phytochemical and biological studies on potential AChE inhibitors in red and zigzag clover dry extracts using reversed–phase liquid chromatography (RP-LC) coupled with photodiode array (PDA) and electron spray ionization-quadrupole/time of flight-mass spectrometric (ESI-QToF/MS-MS) detection and thin-layer chromatography-bioautography. Food Chem. 2022, 375, 131846. [Google Scholar]
- Trifan, A.; Zengin, G.; Sinan, K.I.; Sieniawska, E.; Sawicki, R.; Maciejewska-Turska, M.; Skalikca-Woźniak, K.; Luca, S.V. Unveiling the Phytochemical Profile and Biological Potential of Five Artemisia Species. Antioxidants 2022, 11, 1017. [Google Scholar] [CrossRef]
- Wang, C.R.; Zhou, R.; Ng, T.B.; Wong, J.H.; Qiao, W.T.; Liu, F. First report on isolation of methyl gallate with antioxidant, anti-HIV-1 and HIV-1 enzyme inhibitory activities from a mushroom (Pholiota adiposa). Environ. Toxicol. Pharmacol. 2014, 37, 626–637. [Google Scholar] [CrossRef]
- Zengin, G.; Sieniawska, E.; Senkardes, I.; Picot-Allain, M.C.N.; Sinan, K.I.; Mahomoodally, M.F. Antioxidant abilities, key enzyme inhibitory potential and phytochemical profile of Tanacetum poteriifolium Grierson. Ind. Crops Prod. 2019, 140, 111629. [Google Scholar] [CrossRef]
- Heim, S.; Guarnier, F.; Ferreira, D.; Braz-Filho, R.; Cecchini, R.; Cecchini, A. Antioxidant activity of Spathodea campanulata (Bignoneaceae) extracts. Rev. Bras. Plantas Med. 2012, 14, 287–292. [Google Scholar] [CrossRef] [Green Version]
- Masike, K.; Mhlongo, M.I.; Mudau, S.P.; Nobela, O.; Ncube, E.N.; Tugizimana, F.; George, M.J.; Madala, N.E. Highlighting mass spectrometric fragmentation differences and similarities between hydroxycinnamoyl-quinic acids and hydroxycinnamoyl-isocitric acids. Chem. Cent. J. 2017, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Abidi, J.; Ammar, S.; Brahim, S.B.; Skalicka-Woźniak, K.; Ghrabi-Gammar, Z.; Bouaziz, M. Use of ultra-high-performance liquid chromatography coupled with quadrupole-time-of-flight mass spectrometry system as valuable tool for an untargeted metabolomic profiling of Rumex tunetanus flowers and stems and contribution to the antioxidant activity. J. Pharm. Biomed. Anal. 2019, 162, 66–81. [Google Scholar] [CrossRef] [PubMed]
- Pubchem. Available online: https://pubchem.ncbi.nlm.nih.gov/ (accessed on 30 June 2022).
- Sieniawska, E.; Świątek, Ł.; Sinan, K.I.; Zengin, G.; Boguszewska, A.; Polz-Dacewicz, M.; Bibi Sadeer, N.; Etienne, O.K.; Mahomoodally, M.F. Phytochemical Insights into Ficus sur Extracts and Their Biological Activity. Molecules 2022, 27, 1863. [Google Scholar] [CrossRef] [PubMed]
- Świątek, Ł.; Sieniawska, E.; Sinan, K.I.; Maciejewska-Turska, M.; Boguszewska, A.; Polz-Dacewicz, M.; Senkardes, I.; Guler, G.O.; Bibi Sadeer, N.; Mahomoodally, M.F. LC-ESI-QTOF-MS/MS Analysis, Cytotoxic, Antiviral, Antioxidant, and Enzyme Inhibitory Properties of Four Extracts of Geranium pyrenaicum Burm. f.: A Good Gift from the Natural Treasure. Int. J. Mol. Sci. 2021, 22, 7621. [Google Scholar] [CrossRef] [PubMed]
- Obied, H.K.; Song, Y.; Foley, S.; Loughlin, M.; Rehman, A.-U.; Mailer, R.; Masud, T.; Agboola, S. Biophenols and antioxidant properties of Australian canola meal. J. Agric. Food Chem. 2013, 61, 9176–9184. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Kyotani, Y.; Iinuma, M.; Tanaka, T.; Kojima, H.; Iwatsuki, K. Kaempferol glycosides in Asplenium scolopendvium Newm. Z. Nat. C 1990, 45, 143–146. [Google Scholar] [CrossRef]
- Lachowicz-Wiśniewska, S.; Kapusta, I.; Stinco, C.M.; Meléndez-Martínez, A.J.; Bieniek, A.; Ochmian, I.; Gil, Z. Distribution of Polyphenolic and Isoprenoid Compounds and Biological Activity Differences between in the Fruit Skin+ Pulp, Seeds, and Leaves of New Biotypes of Elaeagnus multiflora Thunb. Antioxidants 2021, 10, 849. [Google Scholar] [CrossRef]
- Mwangi, R.W.; Macharia, J.M.; Wagara, I.N.; Bence, R.L. The antioxidant potential of different edible and medicinal mushrooms. Biomed. Pharmacother. 2022, 147, 112621. [Google Scholar] [CrossRef]
- Alkhaldi, M.M.; Aldisi, D.; Elshafie, M.M.; Alghamdi, M.N.; Sabico, S.; Al-Daghri, N.M. Antioxidant status and dietary pattern of Arab adults with and without metabolic syndrome. J. King Saud Univ.-Sci. 2021, 33, 101561. [Google Scholar] [CrossRef]
- Psaltopoulou, T.; Panagiotakos, D.B.; Pitsavos, C.; Chrysochoou, C.; Detopoulou, P.; Skoumas, J.; Stefanadis, C. Dietary antioxidant capacity is inversely associated with diabetes biomarkers: The ATTICA study. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 561–567. [Google Scholar] [CrossRef] [PubMed]
- Mucaca, C.; Tavares Filho, J.H.; Arruda, L.L.; Galvão, A.M.; Bernadete de Sousa Maia, M.; Maria dos Santos Alves, E.; de Souza Franco, E.; do Nascimento, E. Antioxidant, hypolipidemic and hypoglycemic effect of red wine in wistar rats fed with obesogenic diet. Food Biosci. 2022, 47, 101671. [Google Scholar] [CrossRef]
- Arias, A.; Feijoo, G.; Moreira, M.T. Exploring the potential of antioxidants from fruits and vegetables and strategies for their recovery. Innov. Food Sci. Emerg. Technol. 2022, 77, 102974. [Google Scholar] [CrossRef]
- Jung, S.H.; Kim, B.J.; Lee, E.H.; Osborne, N.N. Isoquercitrin is the most effective antioxidant in the plant Thuja orientalis and able to counteract oxidative-induced damage to a transformed cell line (RGC-5 cells). Neurochem. Int. 2010, 57, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-J.; Kim, H.-N.; Kim, C.Y.; Seo, M.-D.; Baek, S.-H. Synergistic Protection by Isoquercitrin and Quercetin against Glutamate-Induced Oxidative Cell Death in HT22 Cells via Activating Nrf2 and HO-1 Signaling Pathway: Neuroprotective Principles and Mechanisms of Dendropanax morbifera Leaves. Antioxidants 2021, 10, 554. [Google Scholar] [CrossRef]
- Vásquez-Espinal, A.; Yañez, O.; Osorio, E.; Areche, C.; García-Beltrán, O.; Ruiz, L.M.; Cassels, B.K.; Tiznado, W. Theoretical Study of the Antioxidant Activity of Quercetin Oxidation Products. Front. Chem. 2019, 7, 818. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Orčić, D.; Mimica-Dukić, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. J. Funct. Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Rauf, A.; Jehan, N. Natural products as a potential enzyme inhibitors from medicinal plants. Enzym. Inhib. Act. 2017, 165, 177. [Google Scholar]
- Papoutsis, K.; Zhang, J.; Bowyer, M.C.; Brunton, N.; Gibney, E.R.; Lyng, J. Fruit, vegetables, and mushrooms for the preparation of extracts with α-amylase and α-glucosidase inhibition properties: A review. Food Chem. 2021, 338, 128119. [Google Scholar] [CrossRef]
- Liu, T.-T.; Liu, X.-T.; Chen, Q.-X.; Shi, Y. Lipase Inhibitors for Obesity: A Review. Biomed. Pharmacother. 2020, 128, 110314. [Google Scholar] [CrossRef]
- Rajan, L.; Palaniswamy, D.; Mohankumar, S.K. Targeting obesity with plant-derived pancreatic lipase inhibitors: A comprehensive review. Pharmacol. Res. 2020, 155, 104681. [Google Scholar] [CrossRef] [PubMed]
- García-Molina, P.; García-Molina, F.; Teruel-Puche, J.A.; Rodríguez-López, J.N.; García-Cánovas, F.; Muñoz-Muñoz, J.L. Considerations about the kinetic mechanism of tyrosinase in its action on monophenols: A review. Mol. Catal. 2022, 518, 112072. [Google Scholar] [CrossRef]
- Ademosun, A.O.; Oboh, G.; Bello, F.; Ayeni, P.O. Antioxidative Properties and Effect of Quercetin and Its Glycosylated Form (Rutin) on Acetylcholinesterase and Butyrylcholinesterase Activities. J. Evid. Based Complement. Altern. Med. 2016, 21, NP11–NP17. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Gonzalez, A.I.; Díaz-Sánchez, Á.G.; de la Rosa, L.A.; Bustos-Jaimes, I.; Alvarez-Parrilla, E. Inhibition of α-amylase by flavonoids: Structure activity relationship (SAR). Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2019, 206, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Zhang, G.; Hu, X.; Xu, X.; Gong, D. Quercetin as a tyrosinase inhibitor: Inhibitory activity, conformational change and mechanism. Food Res. Int. 2017, 100, 226–233. [Google Scholar] [CrossRef]
- Zhang, A.L.; Ye, Q.; Li, B.G.; Qi, H.Y.; Zhang, G.L. Phenolic and triterpene glycosides from the stems of Ilex litseaefolia. J. Nat. Prod. 2005, 68, 1531–1535. [Google Scholar] [CrossRef]
- Ramsay, K.S.T.; Wafo, P.; Ali, Z.; Khan, A.; Oluyemisi, O.O.; Marasini, B.P.; Khan, I.A.; Bonaventure, N.T.; Choudhary, M.I.; ur-Rahmanb, A. Chemical constituents of Stereospermum acuminatissimum and their urease and alpha-chymotrypsin inhibitions. Fitoterapia 2012, 83, 204–208. [Google Scholar] [CrossRef]
- Mukungu, N.; Abuga, K.; Okalebo, F.; Ingwela, R.; Mwangi, J. Medicinal plants used for management of malaria among the Luhya community of Kakamega East sub-County, Kenya. J. Ethnopharmacol. 2016, 194, 98–107. [Google Scholar] [CrossRef] [Green Version]
- Ochwang’i, D.O.; Kimwele, C.N.; Oduma, J.A.; Gathumbi, P.K.; Mbaria, J.M.; Kiama, S.G. Medicinal plants used in treatment and management of cancer in Kakamega County, Kenya. J. Ethnopharmacol. 2014, 151, 1040–1055. [Google Scholar] [CrossRef]
- Eid, H.H.; Shehab, N.G.; El Zalabani, S.M. GC-MS profile and cytotoxicity of the hydrodistilled and extracted volatiles of the buds and flowers of Spathodea campanulata P. Beauv. J. Biol. Act. Prod. Nat. 2014, 4, 196–208. [Google Scholar]
- Anani, K.; Hudson, J.; De Souza, C.; Akpagana, K.; Tower, G.; Arnason, J.; Gbeassor, M. Investigation of medicinal plants of Togo for antiviral and antimicrobial activities. Pharm. Biol. 2000, 38, 40–45. [Google Scholar] [CrossRef]
- Kane, C.J.; Menna, J.H.; Sung, C.-C.; Yeh, Y.-C. Methyl gallate, methyl-3, 4, 5-trihydroxybenzoate, is a potent and highly specific inhibitor of herpes simplex virusin vitro. II. Antiviral activity of methyl gallate and its derivatives. Biosci. Rep. 1988, 8, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Narayanan, S.; Chang, K.-O. Inhibition of influenza virus replication by plant-derived isoquercetin. Antivir. Res. 2010, 88, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Kim, Y.S.; Hwang, Y.-H.; Yang, H.J.; Li, W.; Kwon, E.-B.; Kim, T.I.; Go, Y.; Choi, J.-G. Quercus acuta Thunb.(Fagaceae) and Its Component, Isoquercitrin, Inhibit HSV-1 Replication by Suppressing Virus-Induced ROS Production and NF-κB Activation. Antioxidants 2021, 10, 1638. [Google Scholar] [CrossRef]
- Mbikay, M.; Chrétien, M. Isoquercetin as an Anti-Covid-19 Medication: A Potential to Realize. Front. Pharmacol. 2022, 13, 830205. [Google Scholar] [CrossRef]
- Qiu, X.; Kroeker, A.; He, S.; Kozak, R.; Audet, J.; Mbikay, M.; Chrétien, M. Prophylactic efficacy of quercetin 3-β-O-D-glucoside against Ebola virus infection. Antimicrob. Agents Chemother. 2016, 60, 5182–5188. [Google Scholar] [CrossRef] [Green Version]
- Wong, G.; He, S.; Siragam, V.; Bi, Y.; Mbikay, M.; Chretien, M.; Qiu, X. Antiviral activity of quercetin-3-β-OD-glucoside against Zika virus infection. Virol. Sin. 2017, 32, 545–547. [Google Scholar] [CrossRef]
- Zengin, G.; Aktumsek, A. Investigation of antioxidant potentials of solvent extracts from different anatomical parts of Asphodeline anatolica E. Tuzlaci: An endemic plant to Turkey. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Uysal, S.; Zengin, G.; Locatelli, M.; Bahadori, M.B.; Mocan, A.; Bellagamba, G.; De Luca, E.; Mollica, A.; Aktumsek, A. Cytotoxic and enzyme inhibitory potential of two Potentilla species (P. speciosa L. and P. reptans Willd.) and their chemical composition. Front. Pharmacol. 2017, 8, 290. [Google Scholar] [CrossRef]
- Grochowski, D.M.; Uysal, S.; Aktumsek, A.; Granica, S.; Zengin, G.; Ceylan, R.; Locatelli, M.; Tomczyk, M. In vitro enzyme inhibitory properties, antioxidant activities, and phytochemical profile of Potentilla thuringiaca. Phytochem. Lett. 2017, 20, 365–372. [Google Scholar] [CrossRef]
- Liu, S.; Knafels, J.D.; Chang, J.S.; Waszak, G.A.; Baldwin, E.T.; Deibel, M.R.; Thomsen, D.R.; Homa, F.L.; Wells, P.A.; Tory, M.C.; et al. Crystal Structure of the Herpes Simplex Virus 1 DNA Polymerase. J. Biol. Chem. 2006, 281, 18193–18200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoog, S.S.; Smith, W.W.; Qiu, X.; Janson, C.A.; Hellmig, B.; McQueney, M.S.; O’Donnell, K.; O’Shannessy, D.; DiLella, A.G.; Debouck, C.; et al. Active Site Cavity of Herpesvirus Proteases Revealed by the Crystal Structure of Herpes Simplex Virus Protease/Inhibitor Complex. Biochemistry 1997, 36, 14023–14029. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]










| Assays | Leaves-MeOH | Leaves-Infusion | Stem Bark-MeOH | Stem Bark-Infusion |
|---|---|---|---|---|
| Extraction Yields (%) Total bioactive compounds | 9.03 | 7.15 | 4.65 | 4.44 |
| Total phenolic content (mg GAE/g) | 89.39 ± 1.69 a | 50.31 ± 0.38 a | 23.58 ± 0.64 d | 31.72 ± 0.41 c |
| Total flavonoid content (mg RE/g) | 6.32 ± 0.01 b | 26.96 ± 0.09 a | 2.38 ± 0.15 d | 3.70 ± 0.21 c |
| Antioxidant assays | ||||
| DPPH radical scavenging (mg TE/g) | 177.48 ± 1.77 a | 42.90 ± 0.03 b | 31.90 ± 0.19 c | 33.37 ± 1.81 c |
| ABTS radical scavenging (mg TE/g) | 186.22 ± 3.52 a | 80.02 ± 1.11 b | 49.33 ± 0.48 d | 60.24 ± 0.89 c |
| CUPRAC (mg TE/g) | 329.69 ± 7.10 a | 131.72 ± 1.78 b | 63.07 ± 2.66 d | 76.04 ± 1.41 c |
| FRAP (mg TE/g) | 220.23 ± 2.50 a | 76.91 ± 1.70 b | 43.23 ± 1.01 d | 47.97 ± 0.95 c |
| Metal chelating (mg EDTAE/g) | 25.30 ± 0.98 b | 33.95 ± 0.28 a | 3.15 ± 0.30 d | 18.61 ± 0.48 c |
| Phosphomolybdenum (mmol TE/g) | 2.58 ± 0.15 a | 1.32 ± 0.04 b | 0.97 ± 0.10 c | 1.14 ± 0.03 bc |
| Enzyme inhibitory assays | ||||
| AChE inhibition (mg GALAE/g) | 1.88 ± 0.22 | na | 1.85 ± 0.19 | na |
| BChE inhibition (mg GALAE/g) | na | na | 6.98 ± 1.04 a | 1.20 ± 0.06 b |
| Tyrosinase inhibition (mg KAE/g) | 59.72 ± 1.53 b | 10.94 ± 0.97 c | 64.41 ± 0.22 a | 5.10 ± 0.66 d |
| Amylase inhibition (mmol ACAE/g) | 0.53 ± 0.01 a | 0.13 ± 0.01 c | 0.43 ± 0.03 b | 0.12 ± 0.01 c |
| Glucosidase inhibition (mmol ACAE/g) | 2.77 ± 0.02 b | 0.85 ± 0.03 c | 3.82 ± 0.07 a | 2.74 ± 0.17 b |
| Spathodea campanulata | Solvent | Sample | CC50 ± SD (µg/mL) | |||
|---|---|---|---|---|---|---|
| VERO | FaDu | HeLa | RKO | |||
| Leaves | MeOH | ScLM | >1000 | 358 ± 11.88 | 914.07 ± 69.81 | 382.07 ± 35.86 |
| Water | ScLI | >1000 | 122.65 ± 19.45 | >1000 | >1000 | |
| Stem bark | MeOH | ScSbM | >1000 | 162.55 ± 15.49 | 119.03 ± 20.72 | 222.07 ± 17.27 |
| Water | ScSbI | >1000 | 196.5 ± 25.74 | 137.97 ± 18.41 | >1000 | |
| S. campanulata | Solvent | Sample | Concentration (µg/mL) | Reduction in HHV-1 Infectious Titre (Δlog) * | Reduction in HSV-1 Viral Load (Δlog’) ** |
|---|---|---|---|---|---|
| Leaves | MeOH | ScLM | 500 | 5.11 ± 0.93 | 1.45 ± 0.13 |
| 250 | 4.22 ± 1.06 | 0.38 ± 0.29 | |||
| Water | ScLI | 125 | 0.16 ± 0.21 | 0.31 ± 0.03 | |
| 62.5 | 0.08 ± 0.07 | 0.28 ± 0.05 | |||
| Stem bark | MeOH | ScSbM | 500 | 1.19 ± 0.46 | 0.28 ± 0.1 |
| 250 | 0.41 ± 0.33 | 0.16 ± 0.15 | |||
| Water | ScSbI | 125 | 0.71 ± 0.19 | 0.17 ± 0.24 | |
| 62.5 | 0.28 ± 0.06 | 0.18 ± 0.1 |
| Binding Energy (Kcal/mol) | |||
|---|---|---|---|
| Compounds | HSV-1 DNA Polymerase | HSV-1 Protease | HSV-1 Thymidine Kinase |
| Caffeic acid | −3.39 | −5.60 | −8.21 |
| Kaempferol 3-O-(2-O-β-d-xylopyranosyl)-β-d-galactopyranoside | −7.37 | −10.10 | - |
| 6-O-trans-caffeoyl-asystasioside E (Spatheoside B) | −6.86 | −10.77 | - |
| Quercetin-3-O-(2-O-β-d-xylopyranosyl)-β-d-galactopyranoside | −7.20 | −10.44 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Świątek, Ł.; Sieniawska, E.; Sinan, K.I.; Zengin, G.; Uba, A.I.; Bene, K.; Maciejewska-Turska, M.; Rajtar, B.; Polz-Dacewicz, M.; Aktumsek, A. Bridging the Chemical Profiles and Biological Effects of Spathodea campanulata Extracts: A New Contribution on the Road from Natural Treasure to Pharmacy Shelves. Molecules 2022, 27, 4694. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27154694
Świątek Ł, Sieniawska E, Sinan KI, Zengin G, Uba AI, Bene K, Maciejewska-Turska M, Rajtar B, Polz-Dacewicz M, Aktumsek A. Bridging the Chemical Profiles and Biological Effects of Spathodea campanulata Extracts: A New Contribution on the Road from Natural Treasure to Pharmacy Shelves. Molecules. 2022; 27(15):4694. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27154694
Chicago/Turabian StyleŚwiątek, Łukasz, Elwira Sieniawska, Kouadio Ibrahime Sinan, Gokhan Zengin, Abdullahi Ibrahim Uba, Kouadio Bene, Magdalena Maciejewska-Turska, Barbara Rajtar, Małgorzata Polz-Dacewicz, and Abdurrahman Aktumsek. 2022. "Bridging the Chemical Profiles and Biological Effects of Spathodea campanulata Extracts: A New Contribution on the Road from Natural Treasure to Pharmacy Shelves" Molecules 27, no. 15: 4694. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27154694