
Antidepressant Effect of Crocin in Mice with Chronic Mild Stress

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Effect of Crocin and Imipramine on Period of Immobility
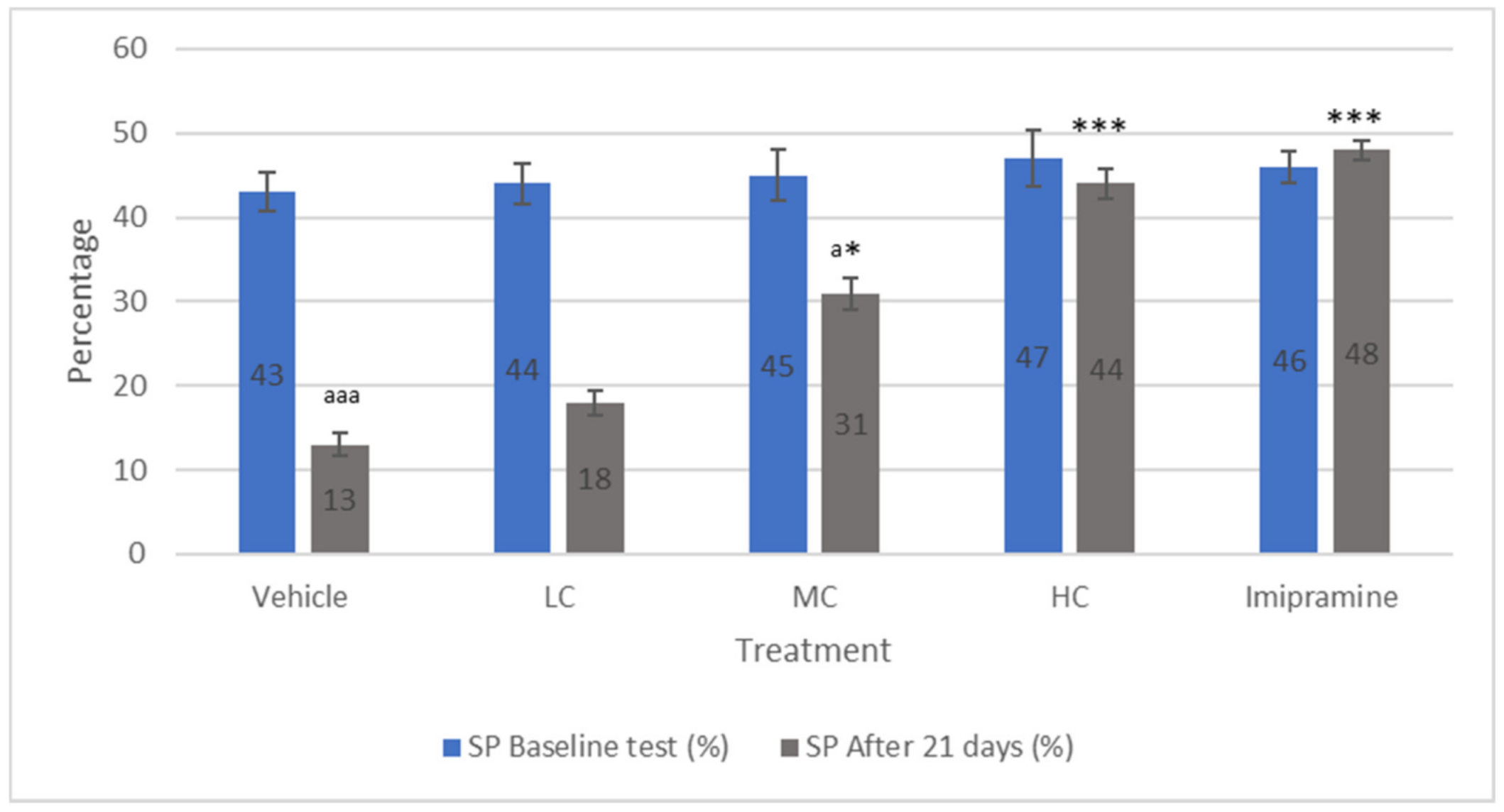
2.2. The Effect of Crocin and Imipramine on Sucrose Preference Model
2.3. The Effect of Crocin and Imipramine on Locomomotor Activity
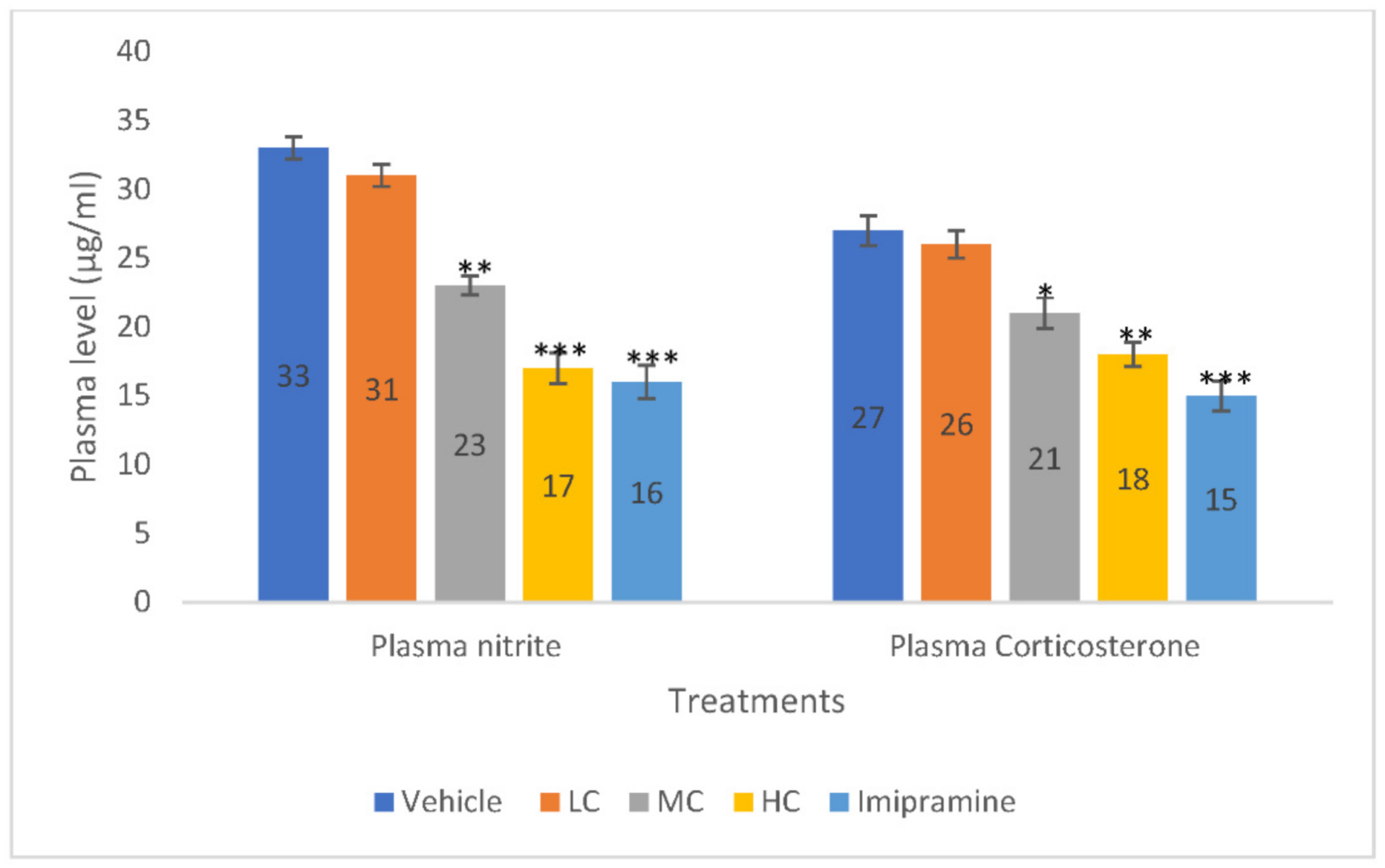
2.4. The Effect of Crocin and Imipramine on Plasma Nitrite and Corticosterone
2.5. The Effect of Crocin and Imipramine on Brain Malondialdehyde (MDA) Level
2.6. The Effect of Crocin and Imipramine on Brain Catalase Activity
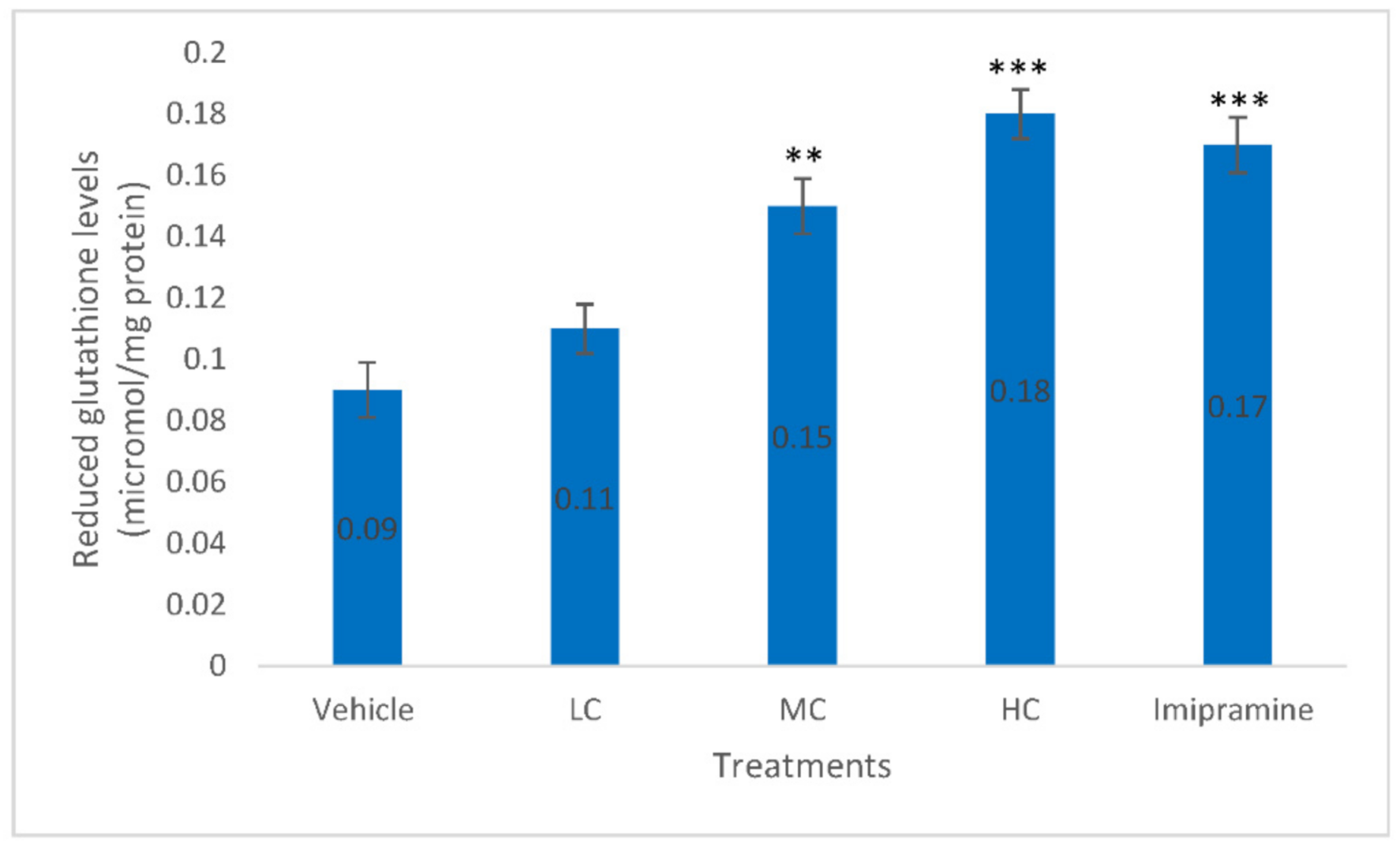
2.7. The Effect of Crocin and Imipramine on Brain Glutathione (GSH) Level
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Materials
4.3. Experimental Grouping
4.4. Chronic Mild Stress (CMS)
4.5. Tail Suspension Test (TST)
4.6. Measurement of Locomotor Activity
4.7. Sucrose Preference Test
4.8. Biochemical Estimations in Plasma
4.9. Biochemical Estimations in Brain Homogenate
4.10. Statistical Evaluation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Schechter, L.E.; Ring, R.H.; Beyer, C.E.; Hughes, Z.A.; Khawaja, X.; Malberg, J.E.; Rosenzweig-Lipson, S. Innovative approaches for the development of antidepressant drugs: Current and future strategies. J. Am. Soc. Exp. Neurother. 2005, 2, 590–611. [Google Scholar] [CrossRef] [PubMed]
- Manji, H.K.; Drevets, W.C.; Charney, D.S. The cellular neurobiology of depression. Nat. Med. 2001, 7, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Leonard, B.; Maes, M. Mechanistic explanations how cell-mediated immune activation, inflammation and oxidative and nitrosative stress pathways and their sequels and concimatants play a role in the pathophysiology of unipolar depression. Neurosci. Biobehav. Rev. 2012, 36, 764–785. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, A.; Nomura, S.; Rinsho, N. Pathophysiology of depression. Nihon Rinsho 2007, 65, 1585–1590. [Google Scholar] [PubMed]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (O & NS) pathways in major depression and their possible contribution to the (neuro) degenerative process in that illness. Prog. Neuro-Psychophar.-Macol. Biol. Psychiatr. 2011, 35, 676–692. [Google Scholar]
- Pariante, C.M.; Lightman, S.L. The HPA axis in major depression: Classical theories 486 and new developments. Trends Neurosci. 2008, 31, 464–468. [Google Scholar] [CrossRef]
- Young, E.A.; Haskett, R.F.; Murphy-Weihberg, V.; Watson, S.J.; Akil, H. Loss of gluco-corticoid fast feed back in depression. Arch. Gen. Psychiatr. 1991, 48, 693–699. [Google Scholar] [CrossRef]
- Nestler, E.J.; Barrot, M.; DiLeone, R.J.; Eisch, A.J.; Gold, S.J.; Monteggia, L.M. Neurobiology of depression. Neuron 2002, 34, 13–25. [Google Scholar] [CrossRef]
- Wróbel, A.; Serefko, A.; Wlaź, P.; Poleszak, E. The effect of imipramine, ketamine, and zinc in the mouse model of depression. Metab. Brain Dis. 2015, 30, 1379–1386. [Google Scholar] [CrossRef]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R. Reduction of sucrose preference by chronic unpredictable mild stress: And its restoration by a tricyclic antidepressant. Psychopharmacology 1987, 93, 358–364. [Google Scholar] [CrossRef]
- Willner, P. Validity, reliability and utility of the chronic mild stress model of depression: A 10-year review and evaluation. Psychopharmacology 1997, 134, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xu, Y.; Han, Q.; Yao, Y.; Xing, H.; Teng, X. Immunosuppression, oxidative stress, and glycometabolism disorder caused by cadmium in common carp (Cyprinus carpio L.): Application of transcriptome analysis in risk assessment of environmental contaminant cadmium. J. Hazard. Mater. 2019, 366, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Yu, M.; Cui, J.; Du, Y.; Teng, X.; Zhang, Z. Heat shock proteins took part in oxidative stress-mediated inflammatory injury via NF-κB pathway in excess manganese-treated chicken livers. Ecotoxicol. Environ. Saf. 2021, 226, 112833. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Miao, Z.; Teng, X.; Xu, S. Chlorpyrifos triggers epithelioma papulosum cyprini cell pyroptosis via miR-124-3p/CAPN1 axis. J. Hazard. Mater. 2022, 424, 127318. [Google Scholar] [CrossRef]
- Madrigal, J.L.; Olivenza, R.; Moro, M.A.; Lizasoain, I.; Lorenzo, P.; Rodrigo, J. Glutathione depletion, lipid peroxidation and mitochondrial dysfunction are induced by chronic stress in rat brain. Neuropsychopharmacology 2001, 24, 420–429. [Google Scholar] [CrossRef]
- Tsuboi, H.; Tatsumi, A.; Yamamoto, K.; Kobayashi, F.; Shimoi, K.; Kinae, N. Possible connections among job stress, depressive symptoms, lipid modulation and antioxidants. J. Affect Disord. 2006, 91, 63–70. [Google Scholar] [CrossRef]
- Goldman, L.S.; Nielson, N.H.; Champion, H.C. Awareness, diagnosis and treatment of depression. J. Gen. Intern. Med. 1999, 14, 569–580. [Google Scholar] [CrossRef]
- Abdullaev, F.I.; Espinosa-Aguirre, J.J. Biomedical properties of saffron and its potential use in cancer therapy and chemoprevention trials. Cancer Detect. Prev. 2004, 28, 426–432. [Google Scholar] [CrossRef]
- Rios, J.L.; Recio, M.C.; Giner, R.M.; Manez, S. An update review of saffron and its active constituents. Phytother. Res. 1996, 10, 189–193. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Khosravan, V. Anticonvulsant effect of Crocus sativus L. stigmas aqueous and ethanolic extracts in mice. Arch Iran Med. 2002, 5, 44–47. [Google Scholar]
- Hosseinzadeh, H.; Karimi, G.; Niapoor, M. Antidepressant effect of Crocus sativus L. stigma extracts and their constituents, crocin and safranal, in mice. Int. Symp. Saffron Biol. Biotechnol. 2003, 650, 435–445. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Younesi, H.M. Antinociceptive and anti-inflammatory effects of Crocus sativus L. stigma and petal extracts in mice. BMC Pharmacol. 2002, 2, 1–8. [Google Scholar]
- Abdullaev, F.I. Biological effects of saffron. BioFactors 1993, 4, 83–86. [Google Scholar] [PubMed]
- Duke, J.A. Handbook of Phytochemical Constituent Grass, Herbs and Other Economic Plants; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Soeda, S.; Ochiai, T.; Shimeno, H.; Saito, H.; Abe, K.; Tanaka, H.; Shoyama, Y. Pharmacological activities of crocin in saffron. J. Nat. Med. 2007, 61, 102–111. [Google Scholar] [CrossRef]
- Abe, K.; Saito, H. Effects of saffron extract and its constituent crocin on learning behaviour and long-term potentiation. Phytother. Res. 2000, 14, 149–152. [Google Scholar] [CrossRef]
- Fernandez, J.A. Anticancer properties of saffron, Crocus sativus Linn. In Advances in Phytomedicine Lead Molecules from Natural Products, 2nd ed.; Khan, M.T.H., Ather, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2006; pp. 313–330. [Google Scholar]
- Konoshima, T.; Tokuda, H.; Morimoto, S.; Tanaka, H.; Kawata, E.; Xuan, E.L.; Saito, H.; Sugiura, M.; Molnar, J.; Shoyama, Y. Crocin and crocetin derivatives inhibit skin tumour promotion in mice. Phytother. Res. 1998, 12, 400–404. [Google Scholar] [CrossRef]
- Kianbakht, S.; Mozaffari, K. Effects of saffron and its active constituents, crocin and safranal, on prevention of indomethacin induced gastric ulcers in diabetic and nondiabetic rats. J. Med. Plants 2009, 8, 30–38. [Google Scholar]
- Assimopoulou, A.N.; Sinakos, Z.; Papageorgiou, V.P. Radical scavenging activity of Crocus sativus L. extract and its bioactive constituents. Phytother. Res. 2005, 19, 997–1000. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Sadeghnia, H.R.; Ziaee, T.; Danaee, A. Protective effect of aqueous saffron extract (Crocus sativus L.) and crocin, its active constituent, on renal ischemia-reperfusion-induced oxidative damage in rats. J. Pharm. Pharm. Sci. 2005, 8, 387–393. [Google Scholar]
- Shen, X.C.; Qian, Z.Y. Effects of crocetin on antioxidant enzymatic activities in cardiac hypertrophy induced by norepinephrine in rats. Pharmazie 2006, 61, 348–352. [Google Scholar]
- Hosseinzadeh, H.; Karimi, G.; Niapoor, M. Antidepressant effects of Crocus sativus stigma extracts and its constituents, crocin and safranal, in mice. J. Med. Plants 2004, 3, 48–58. [Google Scholar]
- Hosseinzadeh, H.; Motamedshariaty, V.; Hadizadeh, F. Antidepressant effect of kaempferol, a constituent of saffron (Crocus sativus) petal, in mice and rats. Pharmacologyonline 2007, 2, 367–370. [Google Scholar]
- Karimi, G.R.; Hosseinzadeh, H.; Khaleghpanah, P. Study of antidepressant effect of aqueous and ethanolic extract of Crocus sativus in mice. Iran. J. Basic Med. Sci. 2001, 4, 11–15. [Google Scholar]
- Wang, Y.; Han, T.; Zhu, Y.; Zheng, C.J.; Ming, Q.L.; Rahman, K.; Qin, L.P. Antidepressant properties of bioactive fractions from the extract of Crocus sativus L. J. Nat. Med. 2010, 64, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Baloch, Z.; Mao, F. Natural Products as a Source for New Leads in Depression Treatment. Evid. -Based Complementary Altern. Med. 2022, 2022, 9791434. [Google Scholar] [CrossRef]
- Dai, W.; Feng, K.; Sun, X.; Xu, L.; Wu, S.; Rahmand, K.; Jia, D.; Han, T. Natural products for the treatment of stress-induced depression: Pharmacology, mechanism and traditional use. J. Ethnopharmacol. 2022, 285, 114692. [Google Scholar] [CrossRef]
- Steru, L.; Chermat, R.; Thierry, B.; Simon, P. The tail suspension test: A new method for screening antidepressants in mice. Psychopharmacology 1985, 85, 367–370. [Google Scholar] [CrossRef]
- Pan, Y.; Zhang, W.Y.; Xia, X.; Kong, L.D. Effects of icariin on hypothalamic-pituitary-adrenal axis action and cytokine levels in stressed Sprague-Dawley rats. Biol. Pharm. Bull. 2006, 29, 2399–2403. [Google Scholar] [CrossRef]
- Sousa, N.; Cerqueira, J.J.; Almeida, O.F. Corticosteroid receptors and neuroplasticity. Brain Res. Rev. 2008, 57, 561–570. [Google Scholar] [CrossRef]
- Swaab, D.F.; Bao, A.M.; Lucassen, P.J. The stress system in the human brain in depression and neurodegeneration. Ageing Res. Rev. 2005, 4, 141–194. [Google Scholar] [CrossRef]
- Bilici, M.; Efe, H.; Köroğlu, M.A.; Uydu, H.A.; Bekaroğlu, M.; Değer, O. Antioxidative enzyme activities and lipid peroxidation in major depression: Alterations by antidepressant treatments. J. Affect. Disord. 2001, 64, 43–51. [Google Scholar] [CrossRef]
- Dhingra, D.; Bansal, S. Antidepressant-like activity of plumbagin in unstressed and stressed mice. Pharmacol. Rep. PR 2015, 67, 1024–1032. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Zhou, Q.; Yu, M.; Liu, Y.; Teng, X.; Gu, X. 4-tert-butylphenol triggers common carp hepatocytes ferroptosis via oxidative stress, iron overload, SLC7A11/GSH/GPX4 axis, and ATF4/HSPA5/GPX4 axis. Ecotoxicol. Environ. Saf. 2022, 242, 113944. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Kuhad, A.; Chopra, K. Neuropsychopharmacological effect of sesamol in unpredictable chronic mild stress model of depression: Behavioral and biochemical evidences. Psychopharmacology 2011, 214, 819–828. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Shariaty, M.V.; Sameni, A.K.; Vahabzadeh, M. Acute and sub-acute toxicity of crocin, a constituent of Crocus sativus L.(saffron), in mice and rats. Pharmacologyonline 2010, 2, 943–951. [Google Scholar]
- Mohamadpour, A.H.; Ayati, Z.; Parizadeh, R.; Hosseinzadeh, H. Safety Evaluation of Crocin (a constituent of saffron) Tablets in Healthy Volunteers. Iran. J. Basic Med. Sci. 2013, 16, 39–46. [Google Scholar] [CrossRef]
- Suchareau, M.; Bordes, A.; Lemée, L. Improved quantification method of crocins in saffron extract using HPLC-DAD after qualification by HPLC-DAD-MS. Food Chem. 2021, 362, 130199. [Google Scholar] [CrossRef]
- Lech, K.; Witowska-Jarosz, J.; Jarosz, M. Saffron yellow: Characterization of carotenoids by high-performance liquid chromatography with electrospray mass spectrometric detection. J. Mass Spectrom. 2009, 44, 1661–1667. [Google Scholar] [CrossRef]
- Carmona, M.; Zalacain, A.; Sánchez, A.M.; Novella, J.L.; Alonso, G.L. Crocetin esters, picrocrocin and its related compounds present in Crocus sativus stigmas and Gardenia jasminoides fruits. Tentative identification of seven new compounds by LC-ESI-MS. J. Agric. Food Chem. 2006, 54, 973–979. [Google Scholar] [CrossRef]
- Xi, L.; Qian, Z.; Du, P.; Fu, J. Pharmacokinetic properties of Crocin (crocetin digentiobiose ester) following oral administration in rats. Phytomedicine 2007, 14, 633–636. [Google Scholar] [CrossRef]
- Lautenschlager, M.; Sendker, J.; Hüwel, S.; Galla, H.J.; Brandt, S.; Düfer, M.; Riehemann, K.; Hensel, A. Intestinal formation of trans-crocetin from saffron extract (Crocus sativus L.) and in vitro permeation through intestinal and blood brain barrier. Phytomedicine Int. J. Phytother. Phytopharm. 2015, 22, 36. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fei, F.; Zhen, L.; Zhu, X.; Wang, J.; Li, S.; Geng, J.; Sun, R.; Yu, X.; Chen, T.; et al. Sensitive analysis and simultaneous assessment of pharmacokinetic properties of Crocin and crocetin after oral administration in rats. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1044–1045, 1–7. [Google Scholar] [CrossRef]
- Karkoula, E.; Lemonakis, N.; Kokras, N.; Dalla, C.; Gikas, E.; Skaltsounis, A.L.; Tsarbopoulos, A. Trans-crocin 4 is not hydrolyzed to crocetin following i.p. administration in mice, while it shows penetration through the blood brain barrier. Fitoterapia 2018, 129, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.; de Lima Melchiades, G.; Michels, L.R.; Figueiró, F.; Bassani, V.L.; Teixeira, H.F.; Koester, L.S. Solid dispersion of kaempferol: Formulation development, characterization, and oral bioavailability assessment. AAPS PharmSciTech 2019, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Asdaq, S.M.B.; Inamdar, M.N. Potential of Crocus sativus (saffron) and its Constituent, Crocin, as Hypolipidemic and Antioxidant in Rats. Appl. Biochem. Biotechnol. 2010, 162, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.G.; Salles, L.A.; Stein, A.C.; Betti, A.H.; Sakamoto, S.; Cassel, E.; Vargas, R.F.; von Poser, G.L.; Rates, S.M. Antidepressant-like effect of Valeriana glechomifolia Meyer (Valerianaceae) in mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2012, 36, 101–109. [Google Scholar] [CrossRef]
- Dhingra, D.; Joshi, P.; Gupta, A.; Chhillar, R. Possible involvement of monoaminergic neurotransmission in antidepressant-like activity of Emblica officinalis fruits in mice. CNS Neurosci. Ther. 2012, 18, 419–425. [Google Scholar] [CrossRef]
- Green, L.C.; Wagner, D.A.; Glogowski, J.; Skipper, P.L.; Wishnok, J.S.; Tannenbaum, S.R. Analysis of nitrate, nitrite, and [15N] nitrate in biological fluids. Anal. Biochem. 1982, 126, 131–138. [Google Scholar] [CrossRef]
- Bartos, J.; Pesez, M. Colorimetric and fluorimetric determination of steroids. Int. Union Pure Appl. Chem. 1979, 51, 2157–2169. [Google Scholar]
- Wills, E.D. Mechanisms of lipid peroxide formation in tissues: Role of metals and haematin proteins in the catalysis of the oxidation of unsaturated fatty acids. Biochem. Biophys. Acta 1965, 98, 238–251. [Google Scholar] [CrossRef]
- Jollow, D.J.; Mitchell, J.R.; Zampaglione, N.; Gillette, J.R. Bromobenz induced liver necrosis: Protective role of glutathione and evidence for 3,4-bromobenzen- 564 oxide as the hepatotoxic metabolite. Pharmacology 1974, 11, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Claiborne, A. Catalase activity. In Handbook of Methods for Oxygen Radical Research; Greenwald, R.A., Ed.; CRC: Boca Raton, FI, USA, 1985; pp. 283–284. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsanie, W.F.; Alamri, A.S.; Abdulaziz, O.; Salih, M.M.; Alamri, A.; Asdaq, S.M.B.; Alhomrani, M.H.; Alhomrani, M. Antidepressant Effect of Crocin in Mice with Chronic Mild Stress. Molecules 2022, 27, 5462. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27175462
Alsanie WF, Alamri AS, Abdulaziz O, Salih MM, Alamri A, Asdaq SMB, Alhomrani MH, Alhomrani M. Antidepressant Effect of Crocin in Mice with Chronic Mild Stress. Molecules. 2022; 27(17):5462. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27175462
Chicago/Turabian StyleAlsanie, Walaa F., Abdulhakeem S. Alamri, Osama Abdulaziz, Magdi M. Salih, Abdulwahab Alamri, Syed Mohammed Basheeruddin Asdaq, Mohammed Hisham Alhomrani, and Majid Alhomrani. 2022. "Antidepressant Effect of Crocin in Mice with Chronic Mild Stress" Molecules 27, no. 17: 5462. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27175462