

Molecular Shield for Protection of Buckwheat Plants from UV-B Radiation

 , ,
, ,
Abstract
:1. Introduction
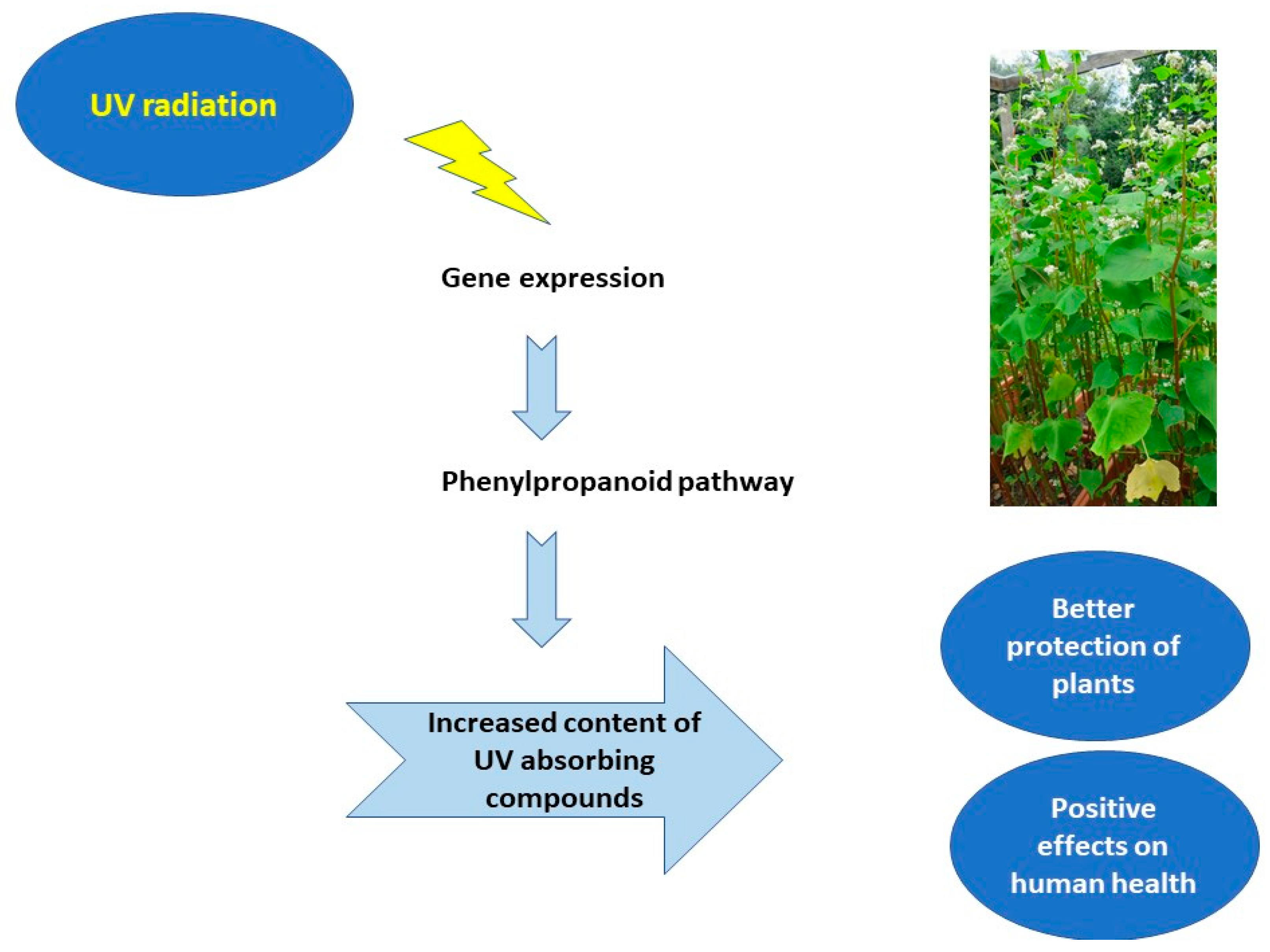
2. Impact of Solar Radiation
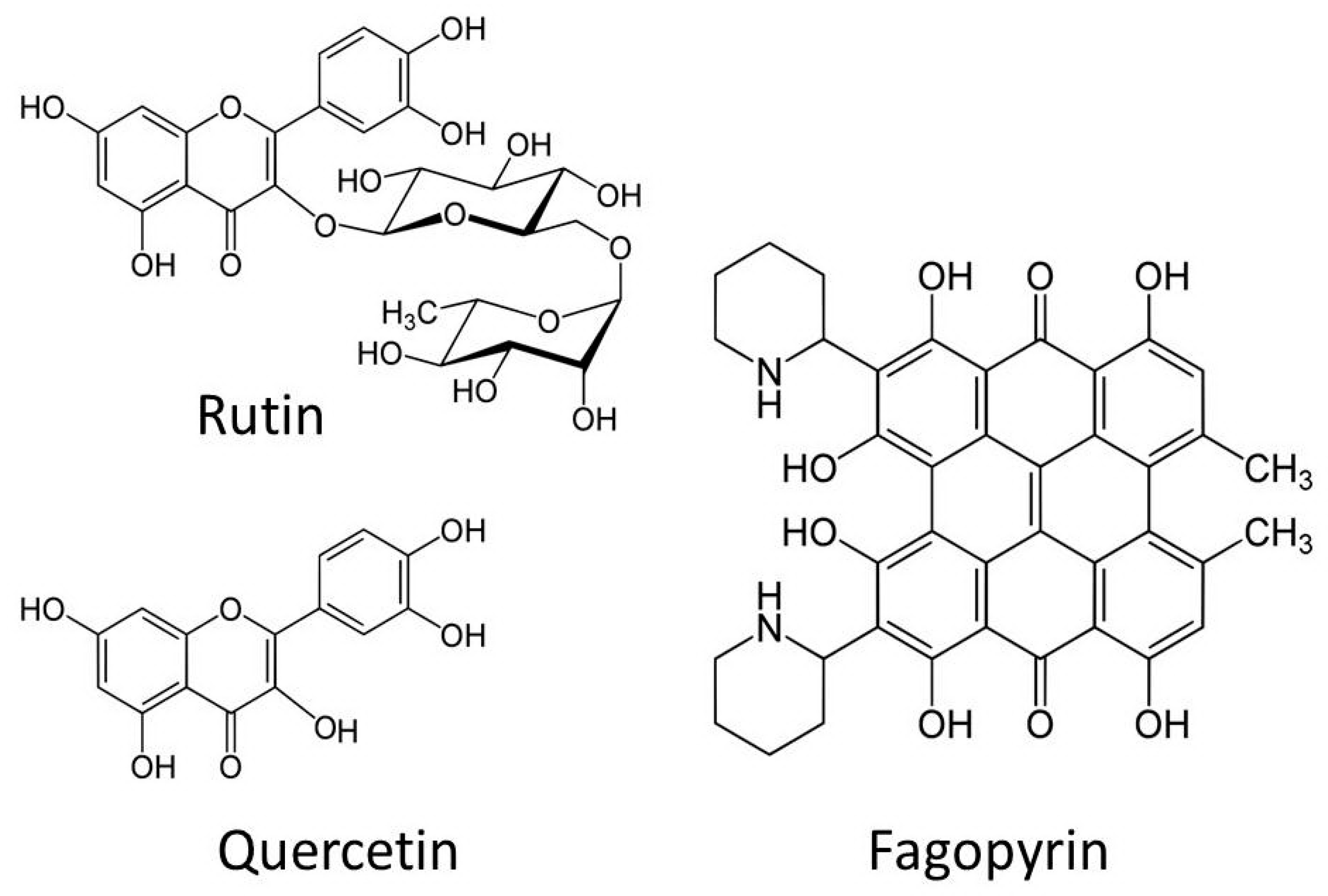
3. Human-Health-Related Effects of Buckwheat
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ohnishi, O. Search for the wild ancestor of buckwheat—III. The wild ancestor of cultivated common buckwheat, and of tatary buckwheat. Econ. Bot. 1998, 52, 123–133. [Google Scholar] [CrossRef]
- Ohnishi, O. Buckwheat in the Himalayan Hills. In Ethnobotany of Buckwheat; Kreft, I., Chang, K.J., Choi, Y.S., Park, C.H., Eds.; Jinsol Publishing Co.: Seoul, Korea, 2003; pp. 21–33. [Google Scholar]
- Ohnishi, O. The origin of cultivated buckwheat in Mankang district of the Sanjiang area of Eastern Tibet and its diffusion to India and the Himalayan hills. Folia Bio. Geol. 2020, 61, 7–15. [Google Scholar] [CrossRef]
- Kreft, I.; Germ, M.; Golob, A.; Vombergar, B.; Bonafaccia, F.; Luthar, Z. Impact of rutin and other phenolic substances on the digestibility of buckwheat grain metabolites. Int. J. Mol. Sci. 2022, 23, 3923. [Google Scholar] [CrossRef]
- Zhang, C.N.; Zhang, R.; Li, Y.M.; Liang, N.; Zhao, Y.M.; Zhu, H.Y.; He, Z.Y.; Liu, J.H.; Hao, W.J.; Jiao, R.; et al. Cholesterol-lowering activity of Tartary buckwheat protein. J. Agr. Food Chem. 2017, 65, 1900–1906. [Google Scholar] [CrossRef]
- Dohm, J.C.; Minoche, A.E.; Holtgräwe, D.; Capella-Gutiérrez, S.; Zakrzewski, F.; Tafer, H.; Rupp, O.; Sörensen, T.R.; Stracke, R.; Reinhardt, R.; et al. The genome of the recently domesticated crop plant sugar beet (Beta vulgaris). Nature 2014, 505, 546–549. [Google Scholar] [CrossRef]
- Zhou, M.; Sun, Z.; Ding, M.; Logacheva, M.D.; Kreft, I.; Wang, D.; Yan, M.; Shao, J.; Tang, Y.; Wu, Y.; et al. FtSAD2 and FtJAZ1 regulate activity of the FtMYB11 transcription repressor of the phenylpropanoid pathway in Fagopyrum tataricum. New Phytol. 2017, 216, 814–828. [Google Scholar] [CrossRef]
- Virant, I.; Kajfež-Bogataj, L. Vesicular–arbuscular mycorrhiza in buckwheat. Fagopyrum 1988, 8, 10–14. [Google Scholar]
- Regvar, M.; Bukovnik, U.; Likar, M.; Kreft, I. UV-B radiation affects flavonoids and fungal colonisation in Fagopyrum esculentum and F. tataricum. Open Life Sci. 2012, 7, 275–283. [Google Scholar] [CrossRef]
- Amann, C.; Amberger, A. Phosphorus Efficiency of Buckwheat (Fagopyrum esculentum). J. Plant Nutr. Soil Sci. 1989, 152, 181–189. [Google Scholar] [CrossRef]
- Possinger, A.R.; Byrne, L.B.; Breen, N.E. Effect of buckwheat (Fagopyrum esculentum) on soil-phosphorus availability and organic acids. J. Plant Nutr. Soil Sci. 2013, 176, 16–18. [Google Scholar] [CrossRef]
- Suzuki, T.; Morishita, T.; Kim, S.-J.; Park, S.U.; Woo, S.H.; Noda, T.; Takigawa, S. Physiological roles of rutin in the buckwheat plant. Jpn. Agric. Res. 2015, 49, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Stojilkovski, K.; Kočevar Glavač, N.; Kreft, S.; Kreft, I. Fagopyrin and flavonoid contents in common, Tartary, and cymosum buckwheat. J. Food Compost. Anal. 2013, 32, 126–130. [Google Scholar] [CrossRef]
- Vollmannová, A.; Musilová, J.; Lidiková, J.; Árvay, J.; Šnirc, M.; Tóth, T.; Bojňanská, T.; Čičová, I.; Kreft, I.; Germ, M. Concentrations of phenolic acids are differently genetically determined in leaves, flowers, and grain of common buckwheat (Fagopyrum esculentum Moench). Plants 2021, 10, 1142. [Google Scholar] [CrossRef]
- Matsushita, K.; Taki, H.; Yamane, F.; Asano, K. Shadow value of ecosystem resilience in complex natural land as a wild pollinator habitat. Am. J. Agric. Econ. 2017, 100, 829–843. [Google Scholar] [CrossRef]
- Germ, M.; Gaberščik, A. The effect of environmental factors on buckwheat. In Molecular Breeding and Nutritional Aspects of Buckwheat; Zhou, M., Kreft, I., Woo, S.H., Chrungoo, N., Wieslander, G., Eds.; Academic Press: London, UK, 2016; pp. 273–282. [Google Scholar] [CrossRef]
- Gaberščik, A.; Grašič, M.; Vogel-Mikuš, K.; Germ, M.; Golob, A. Water shortage strongly alters formation of calcium oxalate druse crystals and leaf traits in Fagopyrum esculentum. Plants 2020, 9, 917. [Google Scholar] [CrossRef]
- Fabjan, N.; Rode, J.; Košir, I.J.; Zhang, Z.; Kreft, I. Tartary buckwheat (Fagopyrum tataricum Gaertn.) as a source of dietary rutin and quercetin. J. Agric. Food Chem. 2003, 51, 6452–6455. [Google Scholar] [CrossRef]
- Vombergar, B.; Luthar, Z. The concentration of flavonoids, tannins and crude proteins in grain fractions of common buckwheat (Fagopyrum esculentum Moench) and Tartary buckwheat (Fagopyrum tataricum Gaertn.). Folia Biol. Geol. 2018, 59, 101–157. [Google Scholar] [CrossRef]
- Tavčar Benković, E.; Kreft, S. Fagopyrins and protofagopyrins: Detection, analysis and potential phototoxicity in buckwheat. J. Agric. Food Chem. 2015, 63, 5715–5724. [Google Scholar] [CrossRef]
- Sytar, O.; Brestic, M.; Rai, M. Possible ways of fagopyrin biosynthesis and production in buckwheat plants. Fitoterapia 2013, 84, 72–79. [Google Scholar] [CrossRef]
- Kočevar Glavač, N.; Stojilkovski, K.; Kreft, S.; Park, C.H.; Kreft, I. Determination of fagopyrins, rutin, and quercetin in Tartary buckwheat products. LWT—Food Sci. Technol. 2017, 79, 423–427. [Google Scholar] [CrossRef]
- Lukšič, L.; Bonafaccia, G.; Timoracka, M.; Vollmannova, A.; Trček, J.; Nyambe, T.K.; Melini, V.; Acquistucci, R.; Germ, M.; Kreft, I. Rutin and quercetin transformation during preparation of buckwheat sourdough bread. J. Cereal Sci. 2016, 69, 71–76. [Google Scholar] [CrossRef]
- Lukšič, L.; Árvay, J.; Vollmannová, A.; Tóth, T.; Škrabanja, V.; Trček, J.; Germ, M.; Kreft, I. Hydrothermal treatment of Tartary buckwheat grain hinders the transformation of rutin to quercetin. J. Cereal Sci. 2016, 72, 131–134. [Google Scholar] [CrossRef]
- Kreft, I.; Zhou, M.; Golob, A.; Germ, M.; Likar, M.; Dziedzic, K.; Luthar, Z. Breeding buckwheat for nutritional quality. Breed. Sci. 2020, 70, 67–73. [Google Scholar] [CrossRef]
- Golob, A.; Luzar, N.; Kreft, I.; Germ, M. Adaptative Responses of Common and Tartary Buckwheat to Different Altitudes. Plants 2022, 11, 1439. [Google Scholar] [CrossRef]
- Zielińska, D.; Turemko, M.; Kwiatkowski, J.; Zieliński, H. Evaluation of flavonoid contents and antioxidant capacity of the aerial parts of common and Tartary buckwheat plants. Molecules 2012, 17, 9668–9682. [Google Scholar] [CrossRef]
- Kim, J.; Hwang, K.T. Fagopyrins in different parts of common buckwheat (Fagopyrum esculentum) and Tartary buckwheat (F. tataricum) during growth. J. Food Compos. Anal. 2020, 86, 103354. [Google Scholar] [CrossRef]
- Kreft, S.; Štrukelj, B.; Gaberščik, A.; Kreft, I. Rutin in buckwheat herbs grown at diferent UV-B radiation levels: Comparison of two UV spectrophotometric and an HPLC method. J. Exp. Bot. 2002, 53, 1801–1804. [Google Scholar] [CrossRef]
- Gaberščik, A.; Vončina, M.; Trošt Sedej, T.; Germ, M.; Björn, L.O. Growth and production of buckwheat (Fagopyrum esculentum) treated with reduced, ambient, and enhanced UV-B radiation. J. Photochem. Photobio. B Biol. 2002, 66, 30–36. [Google Scholar] [CrossRef]
- Golob, A.; Stibilj, V.; Kreft, I.; Vogel-Mikuš, K.; Gaberscik, A.; Germ, M. Selenium treatment alters the effects of UV radiation on chemical and production parameters in hybrid buckwheat. Acta Agric. Scand. Sect. B Plant Soil Sci. 2018, 68, 5–15. [Google Scholar] [CrossRef]
- Debski, H.; Szwed, M.; Wiczkowski, W.; Szawara-Nowak, D.; Baczek, N.; Horbowicz, M. UV-B radiation increases anthocyanin levels in cotyledons and inhibits the growth of common buckwheat seedlings. Acta Biol. Hung. 2016, 67, 403–411. [Google Scholar] [CrossRef] [Green Version]
- Tsurunaga, Y.; Takahashi, T.; Katsube, T.; Kudo, A.; Kuranitsu, O.; Ishiwata, M.; Matsumoto, S. Effects of UV-B irradiation on the levels of anthocyanin, rutin and radical scavenging activity of buckwheat sprouts. Food Chem. 2013, 141, 552–556. [Google Scholar] [CrossRef]
- Chipperfield, M.P.; Dhomse, S.S.; Feng, W.; McKenzie, R.L.; Velders, G.J.M.; Pyle, J.A. Quantifying the ozone and ultraviolet benefits already achieved by the Montreal. Protocol. Nat. Commun. 2015, 6, 7233. [Google Scholar] [CrossRef]
- Hollósy, F. Effects of ultraviolet radiation on plant cells. Micron 2002, 33, 179–197. [Google Scholar] [CrossRef]
- Germ, M.; Breznik, B.; Dolinar, N.; Kreft, I.; Gaberščik, A. The combined effect of water limitation and UV-B radiation on common and Tartary buckwheat. Cereal Res. Commun. 2013, 41, 97–105. [Google Scholar] [CrossRef]
- Morishita, T.; Yamaguchi, H.; Degi, K. The Contribution of Polyphenols to Antioxidative Activity in Common Buckwheat and Tartary Buckwheat Grain. Plant Prod. Sci. 2007, 10, 99–104. [Google Scholar] [CrossRef]
- Sood, P.; Rana, S.S. Integrated effects of genotypes and fertility levels on leaf weevil infestation in buckwheat, Fagopyrum sp. under Western Himalayan conditions. Fagopyrum 2004, 21, 114–116. [Google Scholar]
- Stec, K.; Kordan, B.; Gabryś, B. Quercetin and Rutin as Modifiers of Aphid Probing Behavior. Molecules 2021, 26, 3622. [Google Scholar] [CrossRef]
- Kreft, I. Grenko Seme Tatarske Ajde; SAZU: Ljubljana, Slovenia, 2020; p. 79. [Google Scholar]
- Sytar, O.; Kotta, K.; Valasiadis, D.; Kosyan, A.; Brestic, M.; Koidou, V.; Papadopoulou, E.; Kroustalaki, M.; Emmanouilidou, C.; Pashalidis, A.; et al. The Effects of Photosensitizing Dyes Fagopyrin and Hypericin on Planktonic Growth and Multicellular Life in Budding Yeast. Molecules 2021, 26, 4708. [Google Scholar] [CrossRef]
- Nobili, C.; de Acutis, A.; Reverberi, M.; Bello, C.; Leone, G.P.; Palumbo, D.; Natella, F.; Procacci, S.; Zjalic, S.; Brunori, A. Buckwheat hull extracts inhibit Aspergillus flavus growth and AFB1 biosynthesis. Front. Microbiol. 2019, 10, 1997. [Google Scholar] [CrossRef]
- Chitarrini, G.; Nobili, C.; Pinzari, F.; Antonini, A.; de Rossi, P.; del Fiore, A.; Procacci, S.; Tolaini, V.; Scala, V.; Scarpari, M.; et al. Buckwheat achenes antioxidant profile modulates Aspergillus flavus growth and aflatoxin production. Int. J. Food Microbiol. 2014, 189, 1–10. [Google Scholar] [CrossRef]
- Klug, B.; Horst, W.J. Oxalate exudation into the root-tip water free space confers protection from Al toxicity and allows Al accumulation in the symplast in buckwheat (Fagopyrum esculentum). New Phytol. 2010, 187, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Yu, L.; Yang, C.; Liu, Y. Genotypic difference in aluminum resistance and oxalate exudation of buckwheat. J. Plant Nutr. 2002, 26, 1767–1777. [Google Scholar] [CrossRef]
- Golob, A.; Stibilj, V.; Nečemer, M.; Kump, P.; Kreft, I.; Hočevar, A.; Gaberščik, A.; Germ, M. Calcium oxalate druses affect leaf optical properties in selenium-treated Fagopyrum tataricum. J. Photochem. Photobiol. B Biol. 2018, 180, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.-H.; Tanaka, S.; Goto, E. Growth and Biosynthesis of Phenolic Compounds of Canola (Brassica napus L.) to Different Ultraviolet (UV)-B Wavelengths in a Plant Factory with Artificial Light. Plants 2022, 11, 1732. [Google Scholar] [CrossRef]
- Santin, M.; Sciampagna, M.C.; Mannucci, A.; Puccinelli, M.; Angelini, L.G.; Tavarini, S.; Accorsi, M.; Incrocci, L.; Ranieri, A.; Castagna, A. Supplemental UV-B Exposure Influences the Biomass and the Content of Bioactive Compounds in Linum usitatissimum L. Sprouts and Microgreens. Horticulturae 2022, 8, 213. [Google Scholar] [CrossRef]
- Yin, Y.; Tian, X.; Yang, J.; Yang, Z.; Tao, J.; Fang, W. Melatonin Mediates Isoflavone Accumulation in Germinated Soybeans (Glycine max L.) Under Ultraviolet-B Stress. Plant Physiol. Biochem. 2022, 175, 23–32. [Google Scholar] [CrossRef]
- Breznik, B.; Germ, M.; Gaberšcik, A.; Kreft, I. Combined effects of elevated UV-B radiation and the addition of selenium on common (Fagopyrum esculentum Moench) and tartary (Fagopyrum tataricum (L.) Gaertn.) buckwheat. Photosyntetica 2005, 43, 583–589. [Google Scholar] [CrossRef]
- Tadina, N.; Germ, M.; Kreft, I.; Breznik, B.; Gaberščik, A. Effects of water deficit and selenium on common buckwheat (Fagopyrum esculentum Moench.) plants. Photosynthetica 2007, 45, 472–476. [Google Scholar] [CrossRef]
- Golob, A.; Gadžo, D.; Stibilj, V.; Djikić, M.; Gavrić, T.; Kreft, I.; Germ, M. Sulphur interferes with selenium accumulation in Tartary buckwheat plants. Plant Physiol. Biochem. 2016, 108, 32–36. [Google Scholar] [CrossRef]
- Luthar, Z.; Golob, A.; Germ, M.; Vombergar, B.; Kreft, I. Tartary Buckwheat in Human Nutrition. Plants 2021, 10, 700. [Google Scholar] [CrossRef]
- Bae, M.; Kim, H. The role of vitamin C, vitamin D, and selenium in immune system against COVID-19. Molecules 2020, 25, 5346. [Google Scholar] [CrossRef] [PubMed]
- Luthar, Z.; Germ, M.; Likar, M.; Golob, A.; Vogel-Mikuš, K.; Pongrac, P.; Kušar, A.; Pravst, I.; Kreft, I. Breeding buckwheat for increased levels of rutin, quercetin and other bioactive compounds with potential antiviral effects. Plants 2020, 9, 1638. [Google Scholar] [CrossRef] [PubMed]
- Bonafaccia, G.; Marocchini, M.; Kreft, I. Composition and technological properties of the flour and bran from common and Tartary buckwheat. Food Chem. 2003, 80, 9–15. [Google Scholar] [CrossRef]
- Sytar, O.; Kosyan, A.; Taran, N.; Smetanska, I. Anthocyanin’s as marker for selection of buckwheat plants with high rutin content. Gesunde Pflanz. 2014, 66, 165–169. [Google Scholar] [CrossRef]
- Kuwabara, T.; Han, K.H.; Hashimoto, N.; Yamauchi, H.; Shimada, K.I.; Sekikawa, M.; Fukushima, M. Tartary buckwheat sprout powder lowers plasma cholesterol level in rats. J. Nutr. Sci. Vitaminol. 2007, 53, 501–507. [Google Scholar] [CrossRef]
- Suzuki, T.; Morishita, T.; Mukasa, Y.; Takigawa, S.; Yokota, S.; Ishiguro, K.; Noda, T. Breeding of “Manten-Kirari”, a non-bitter and trace-rutinosidase variety of Tartary buckwheat (Fagopyrum tataricum Gaertn.). Breed. Sci. 2014, 64, 344–350. [Google Scholar] [CrossRef]
- Nishimura, M.; Ohkawara, T.; Sato, Y.; Satoh, H.; Suzuki, T.; Ishiguro, K.; Noda, T.; Morishita, T.; Nishihira, J. Effectiveness of rutin-rich Tartary buckwheat (Fagopyrum tataricum Gaertn) ‘Manten-Kirari’ in body weight reduction related to its antioxidant properties: A randomised, double-blind, placebo-controlled study. J. Funct. Foods 2016, 26, 460–469. [Google Scholar] [CrossRef]
- Yu, J.H.; Kwon, S.J.; Choi, J.Y.; Ju, Y.H.; Roy, S.K.; Lee, D.G.; Park, C.H.; Woo, S.H. Variation of rutin and quercetin contents in Tartary buckwheat germplasm. Fagopyrum 2019, 36, 51–65. [Google Scholar] [CrossRef]
- Li, H.; Lv, Q.; Liu, A.; Wang, J.; Sun, X.; Deng, J.; Chen, Q.; Wu, Q. Comparative metabolomics study of Tartary (Fagopyrum tataricum (L.) Gaertn) and common (Fagopyrum esculentum Moench) buckwheat seeds. Food Chem. 2022, 371, 131125. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, K.; Guo, W.; Zhang, C.; Chen, H.; Xu, T.; Lu, Y.; Wu, Q.; Li, Y.; Chen, Y. Aspergillus niger fermented Tartary buckwheat ameliorates obesity and gut microbiota dysbiosis through the NLRP3/Caspase-1 signaling pathway in high-fat diet mice. J. Funct. Foods 2022, 95, 105171. [Google Scholar] [CrossRef]
- Suzuki, T.; Morishita, T.; Takigawa, S.; Noda, T.; Ishiguro, K. Characterization of rutin-rich bread made with ‘Manten-Kirari’, a trace-rutinosidase variety of Tartary buckwheat (Fagopyrum tataricum Gaertn.). Food Sci. Technol. Res. 2015, 21, 733–738. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Morishita, T.; Noda, T.; Ishiguro, K. Acute and subacute toxicity studies on rutin-rich Tartary buckwheat dough in experimental animals. J. Nutr. Sci. Vitaminol. 2015, 61, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Morishita, T.; Noda, T.; Ishiguro, K.; Otsuka, S.; Katsu, K. Breeding of buckwheat to reduce bitterness and rutin hydrolysis. Plants 2021, 10, 791. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Morishita, T.; Takigawa, S.; Noda, T.; Ishiguro, K.; Otsuka, S. Development of Novel Detection Method for Rutinosidase in Tartary Buckwheat (Fagopyrum tataricum Gaertn.). Plants 2022, 11, 320. [Google Scholar] [CrossRef]
- Germ, M.; Árvay, J.; Vollmannová, A.; Tóth, T.; Golob, A.; Luthar, Z.; Kreft, I. The temperature threshold for the transformation of rutin to quercetin in Tartary buckwheat dough. Food Chem. 2019, 283, 28–31. [Google Scholar] [CrossRef]
- Fujita, K.; Yoshihashi, T. Heat-treatment of Tartary buckwheat (Fagopyrum tataricum Gaertn.) provides dehulled and gelatinized product with denatured rutinosidase. Food Sci. Technol. Res. 2019, 25, 613–618. [Google Scholar] [CrossRef]
- Kawabata, K.; Mukai, R.; Ishisaka, A. Quercetin and related polyphenols: New insights and implications for their bioactivity and bioavailability. Food Funct. 2015, 6, 1399–1417. [Google Scholar] [CrossRef]
- Rusu, M.E.; Simedrea, R.; Gheldiu, A.M.; Mocan, A.; Vlase, L.; Popa, D.S.; Ferreira, I.C.F.R. Benefits of Tree Nut Consumption on Aging and Age-Related Diseases: Mechanisms of Actions. Trends Food Sci. Technol. 2019, 88, 104–120. [Google Scholar] [CrossRef]
- Mateș, L.; Popa, D.S.; Rusu, M.E.; Fizeșan, I.; Leucuța, D. Walnut intake interventions targeting biomarkers of metabolic syndrome and inflammation in middle-aged and older adults: A systematic review and meta-analysis of randomized controlled trials. Antioxidants 2022, 11, 1412. [Google Scholar] [CrossRef]
- Hirudkar, J.R.; Parmar, K.M.; Prasad, R.S.; Sinha, S.K.; Lomte, A.D.; Itankar, P.R.; Prasad, S.K. The antidiarrhoeal evaluation of Psidium guajava L. against enteropathogenic Escherichia coli induced infectious diarrhoea. J. Ethnopharmacol. 2020, 251, 112561. [Google Scholar] [CrossRef]
- Paul, A.K. Potential role of flavonoids against SARS-CoV-2 induced diarrhea. Trop. Biomed. 2021, 38, 360–365. [Google Scholar] [CrossRef]
- Paul, A.K.; Hossain, K.; Mahboob, T.; Nissapatorn, V.; Wilairatana, P.; Jahan, R.; Jannat, K.; Bondhon, T.A.; Hasan, A.; Pereira, M.D.L.; et al. Does oxidative stress management help alleviation of COVID-19 symptoms in patients experiencing diabetes? Nutrients 2022, 14, 321. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Cai, C.; Kreft, I.; Lah Turnšek, T.; Zu, M.; Hu, Y.; Zhou, M.; Liao, Z. Tartary Buckwheat Flavonoids Improve Colon Lesions and Modulate Gut Microbiota Composition in Diabetic Mice. Evid Based Complement Alternat Med. 2022, 2022, 1–14. [Google Scholar] [CrossRef]
- Li, Z.; Li, Z.; Huang, Y.; Jiang, Y.; Liu, Y.; Wen, W.; Li, H.; Shao, J.; Wang, C.; Zhu, X. Antioxidant capacity, metal contents, and their health risk assessment of Tartary buckwheat teas. ACS Omega 2020, 5, 9724–9732. [Google Scholar] [CrossRef]
- Peng, L.; Zhang, Q.; Zhang, Y.; Yao, Z.; Song, P.; Wei, L.; Zhao, G.; Yan, Z. Effect of tartary buckwheat, rutin, and quercetin on lipid metabolism in rats during high dietary fat intake. Food Sci. Nutr. 2019, 8, 199–213. [Google Scholar] [CrossRef]
- Škrabanja, V.; Liljeberg Elmståhl, H.G.M.; Kreft, I.; Björck, I.M.E. Nutritional properties of starch in buckwheat products: Studies in vitro and in vivo. J. Agric. Food Chem. 2001, 49, 490–496. [Google Scholar] [CrossRef]
- Ikeda, K.; Ishida, Y.; Ikeda, S.; Asami, Y.; Lin, R. Tartary, but not common, buckwheat inhibits α-glucosidase activity: Its nutritional implications. Fagopyrum 2017, 34, 13–18. [Google Scholar] [CrossRef]
- Luo, K.; Zhou, X.; Zhang, G. The impact of Tartary buckwheat extract on the nutritional property of starch in a whole grain context. J. Cereal Sci. 2019, 89, 102798. [Google Scholar] [CrossRef]
- Suzuki, T.; Morishita, T.; Takigawa, S.; Noda, T.; Ishiguro, K. Evaluation of the Mutagenicity Potential of Trace-Rutinosidase Variety of Tartary Buckwheat (Fagopyrum tataricum Gaertn.) Using the Ames Test. J. Agric. Chem. Environ. 2016, 5, 100–105. [Google Scholar] [CrossRef]
- Vogrinčič, M.; Kreft, I.; Filipič, M.; Žegura, B. Antigenotoxic effect of tartary (Fagopyrum tataricum) and common (Fagopyrum esculentum) buckwheat flour. J. Med. Food 2013, 16, 944–952. [Google Scholar] [CrossRef]
- Eguchi, K.; Anase, T.; Osuga, H. Development of a high-performance liquid chromatography method to determine the fagopyrin content of Tartary buckwheat (Fagopyrum tartaricum Gaertn.) and common buckwheat (F. esculentum Moench). Plant Prod. Sci. 2009, 12, 475–480. [Google Scholar] [CrossRef]
- Benković, E.T.; Žigon, D.; Friedrich, M.; Plavec, J.; Kreft, S. Isolation, analysis and structures of phototoxic fagopyrins from buckwheat. Food Chem. 2014, 143, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Szymański, S.; Majerz, I. Theoretical studies on the structure and intramolecular interactions of fagopyrins—Natural photosensitizers of fagopyrum. Molecules 2022, 27, 3689. [Google Scholar] [CrossRef] [PubMed]
- Zambounis, A.; Sytar, O.; Valasiadis, D.; Hilioti, Z. Effect of photosensitisers on growth and morphology of Phytophthora citrophthora coupled with leaf bioassays in pear seedlings. Plant Protect. Sci. 2020, 56, 74–82. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Part of Plant | Common Buckwheat | Tartary Buckwheat | ||
|---|---|---|---|---|
| Flavonoids (mg RE/g DW) | Fagopyrins (mg HE/g DW) | Flavonoids (mg RE/g DW) | Fagopyrins (mg HE/g DW) | |
| Stems | / | 1.20 | / | 2.00 |
| Leaves | 82 | 1.03 | 78 | 0.56 |
| Flowers | 204 | 2.25 | 145 | 8.06 |
| Ripe seeds | 6 | / | 20 | / |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kreft, I.; Vollmannová, A.; Lidiková, J.; Musilová, J.; Germ, M.; Golob, A.; Vombergar, B.; Kocjan Ačko, D.; Luthar, Z. Molecular Shield for Protection of Buckwheat Plants from UV-B Radiation. Molecules 2022, 27, 5577. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27175577
Kreft I, Vollmannová A, Lidiková J, Musilová J, Germ M, Golob A, Vombergar B, Kocjan Ačko D, Luthar Z. Molecular Shield for Protection of Buckwheat Plants from UV-B Radiation. Molecules. 2022; 27(17):5577. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27175577
Chicago/Turabian StyleKreft, Ivan, Alena Vollmannová, Judita Lidiková, Janette Musilová, Mateja Germ, Aleksandra Golob, Blanka Vombergar, Darja Kocjan Ačko, and Zlata Luthar. 2022. "Molecular Shield for Protection of Buckwheat Plants from UV-B Radiation" Molecules 27, no. 17: 5577. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27175577