

Switching the N-Capping Region from all-L to all-D Amino Acids in a VEGF Mimetic Helical Peptide
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
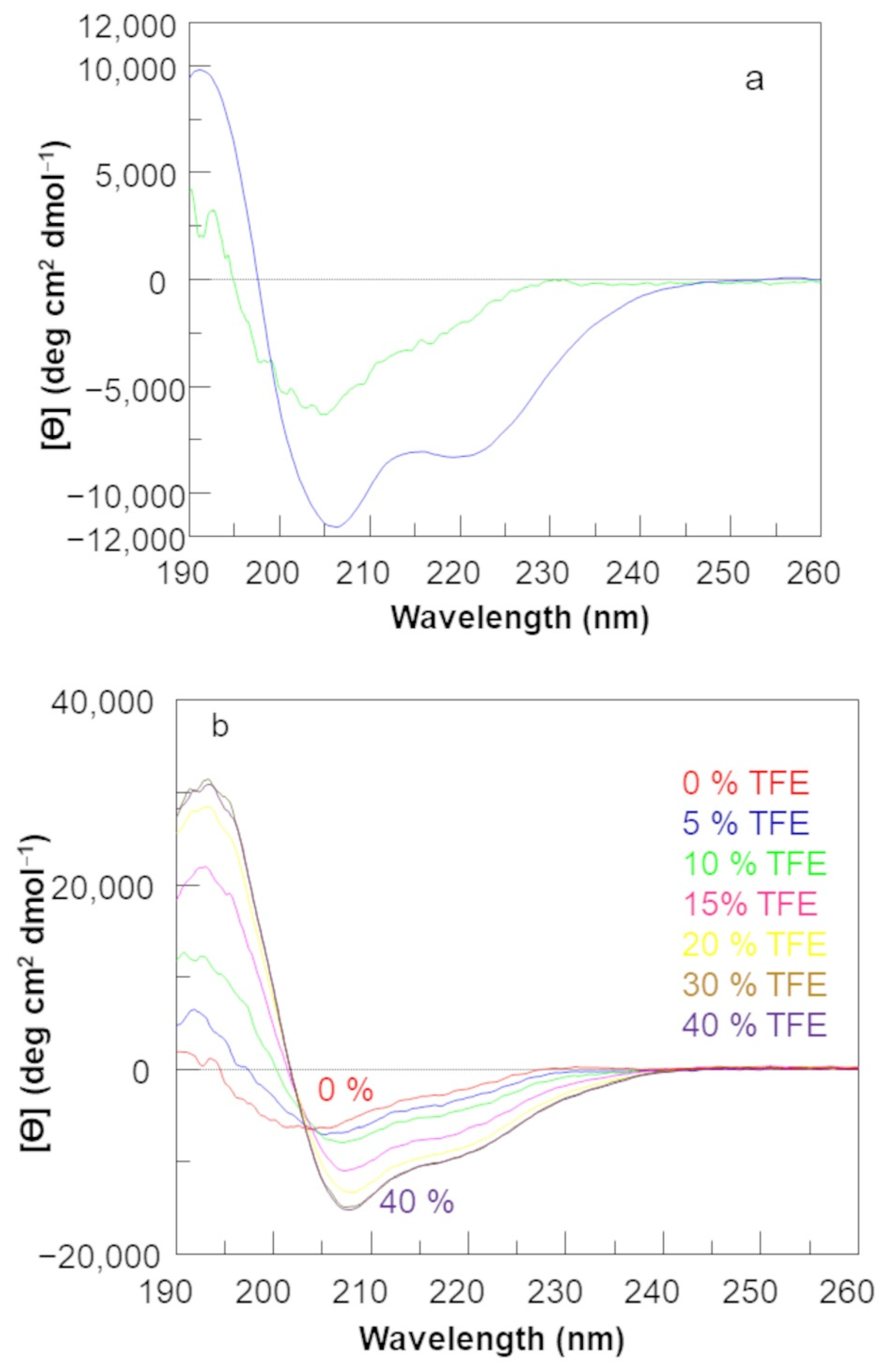
2.1. Circular Dichroism Analysis
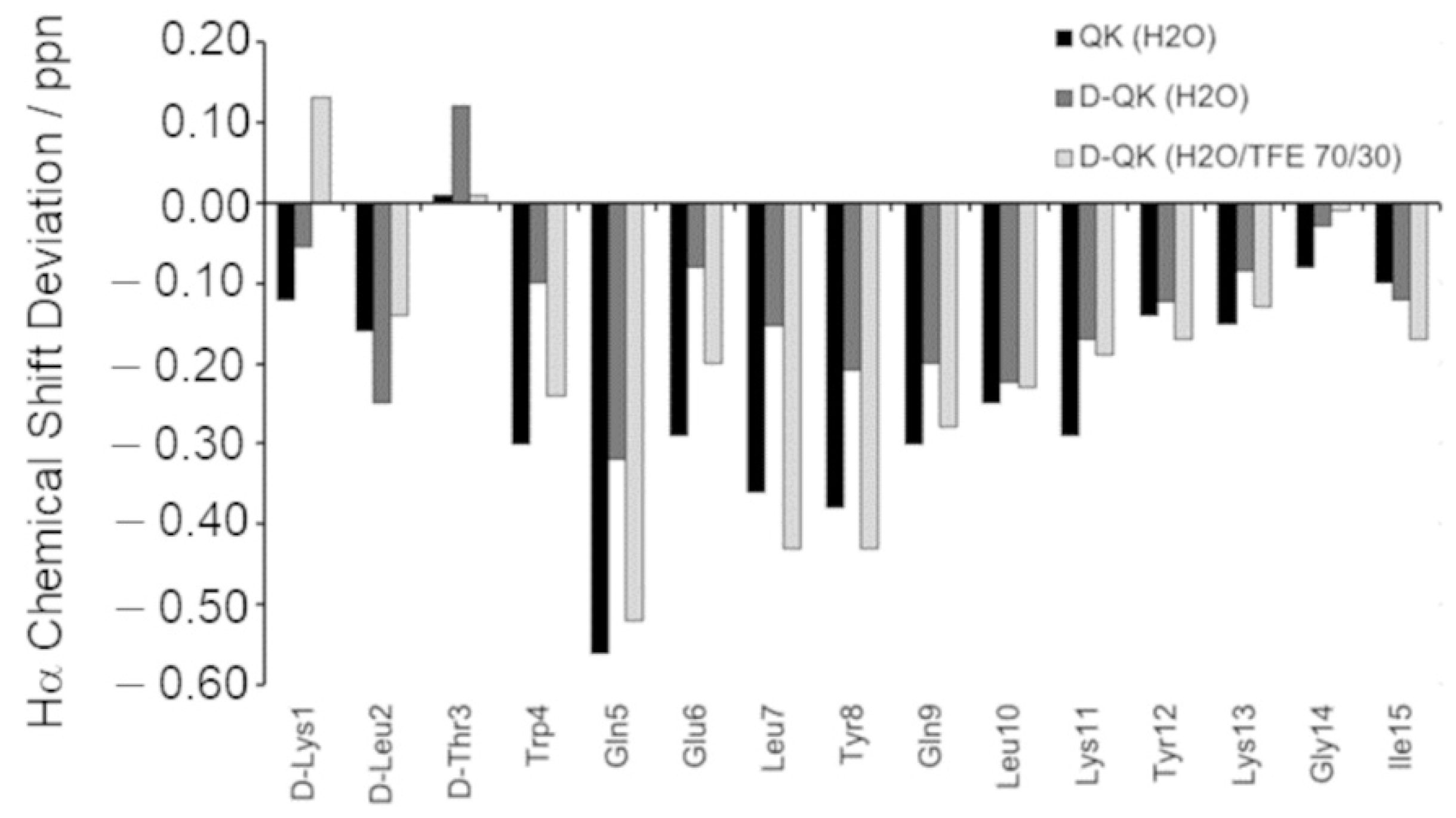
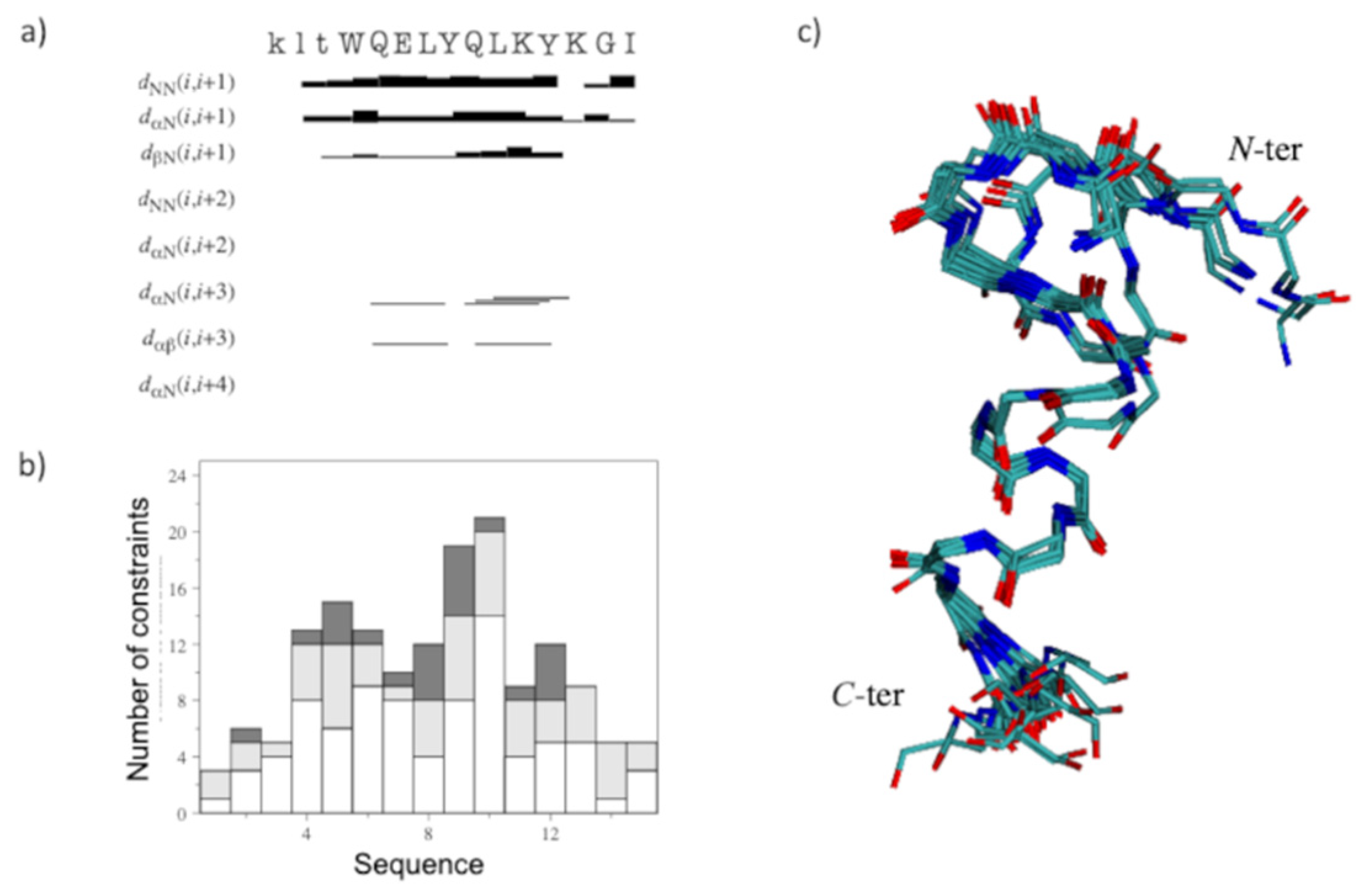
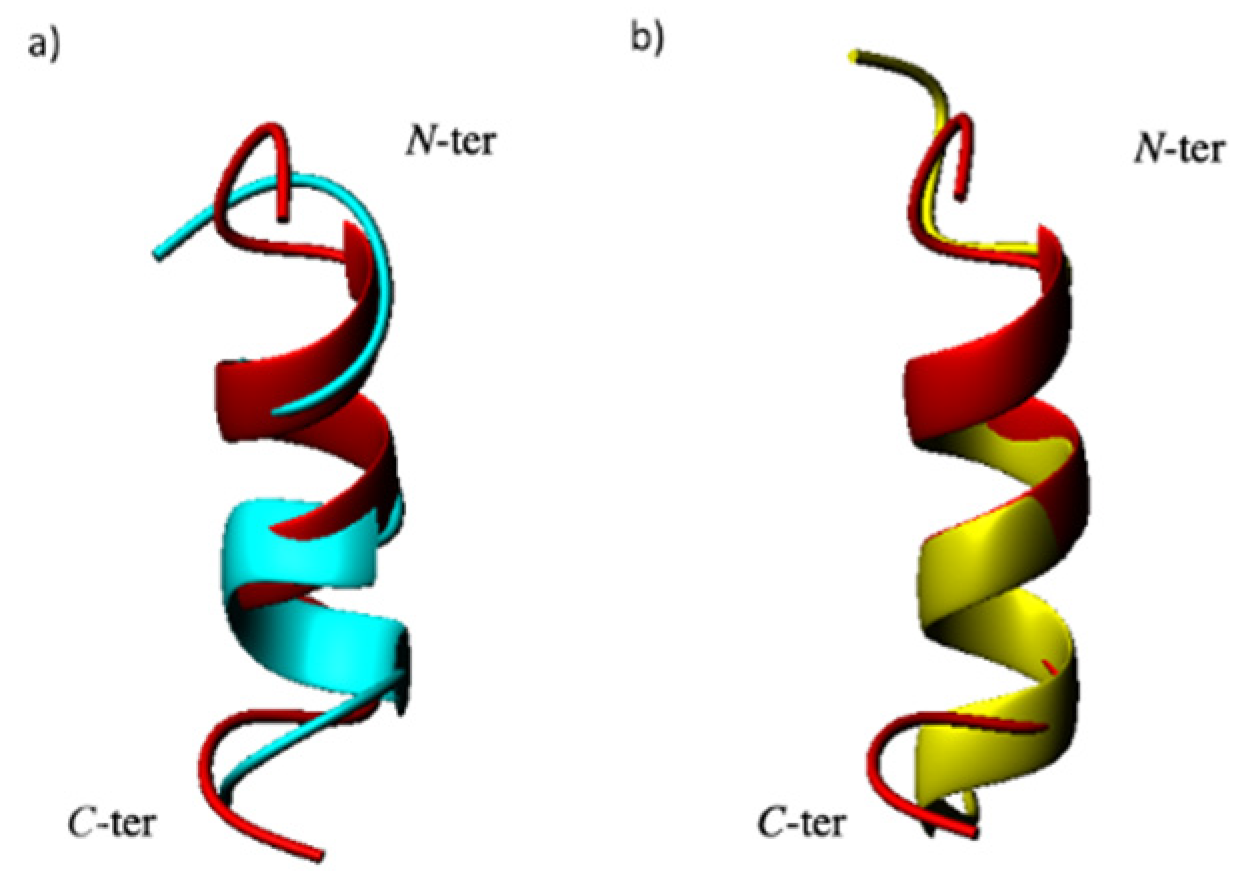
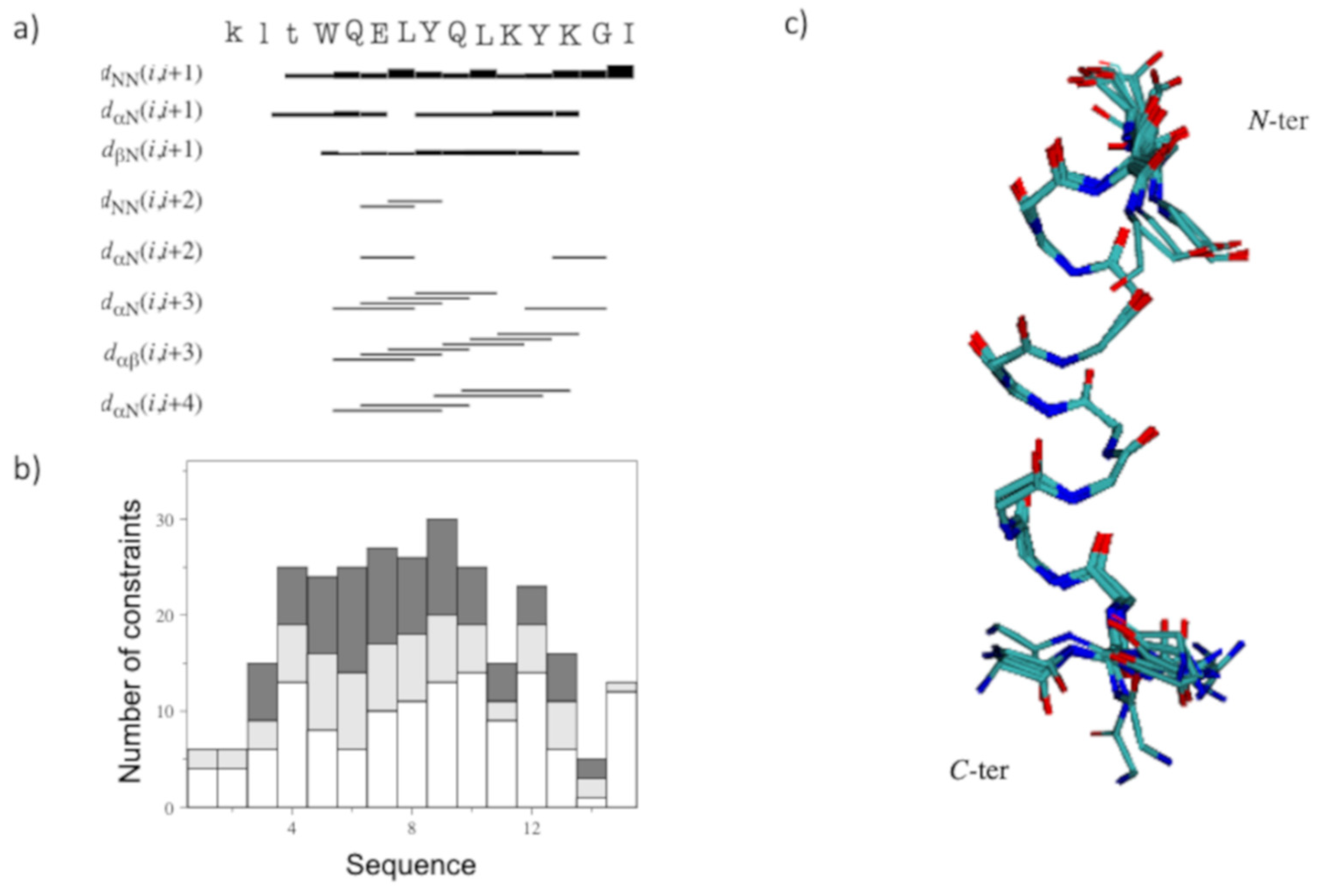
2.2. Structural Investigation by NMR Spectroscopy
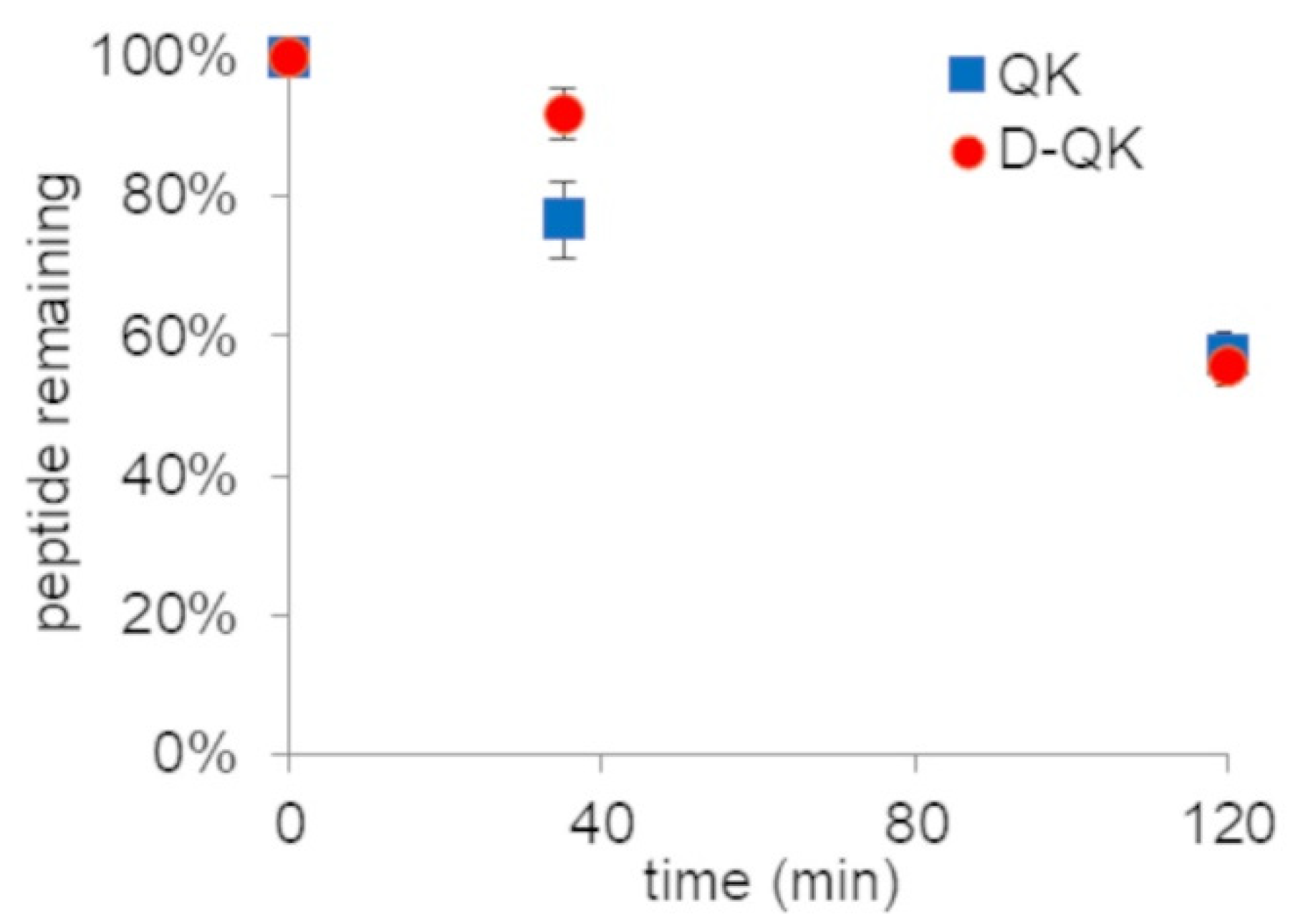
2.3. Serum Stability Analysis
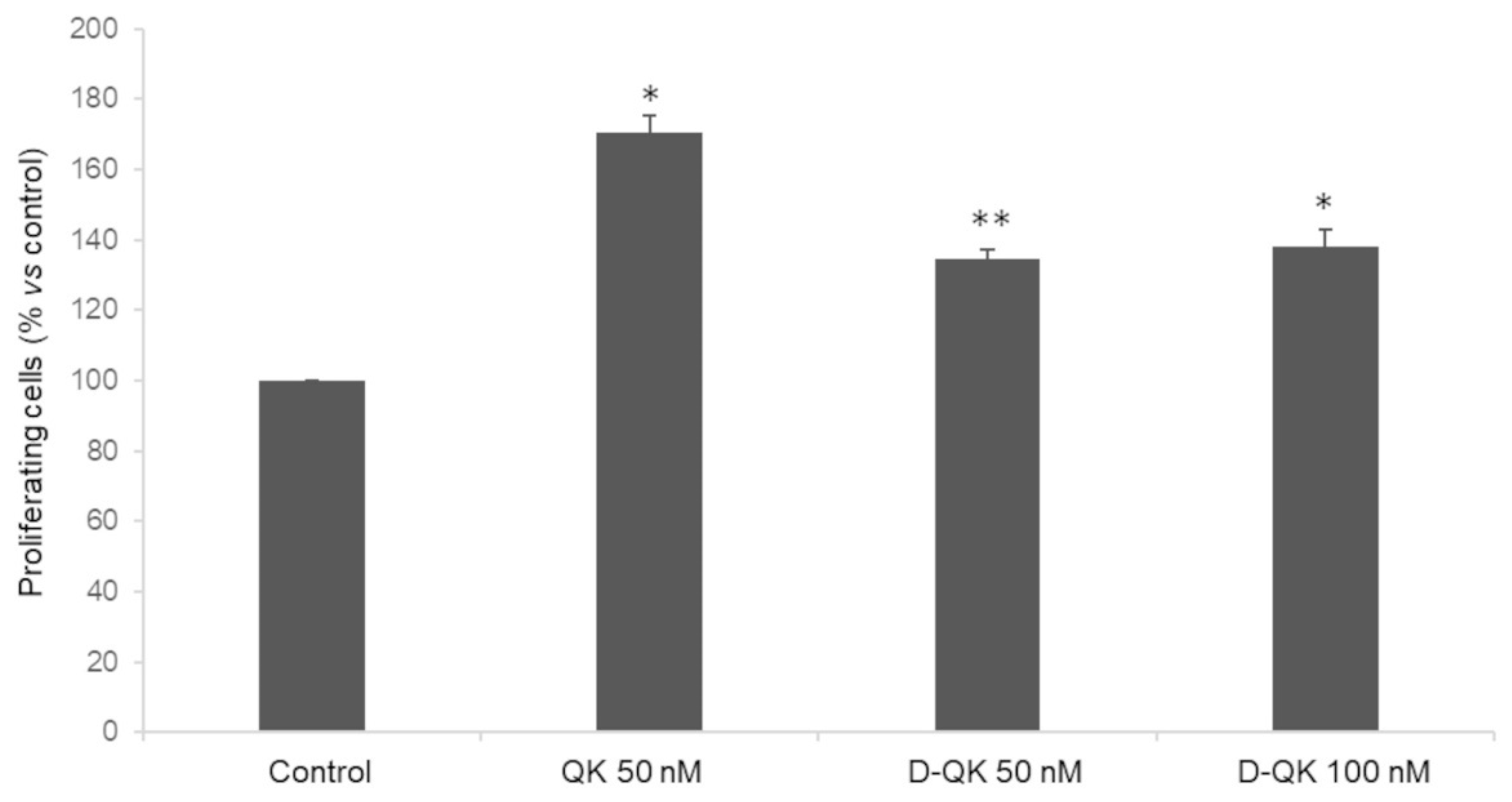
2.4. Cell Proliferation Analysis
3. Discussion
4. Materials and Methods
4.1. Peptide Synthesis
4.2. Circular Dichroism Spectroscopy
4.3. Nuclear Magnetic Resonance Spectroscopy
4.4. Serum Stability Analysis
4.5. Cell Culture and Conditions
4.6. Cell Proliferation Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rohl, C.A.; Baldwin, R.L. Deciphering rules of helix stability in peptides. Methods Enzymol. 1998, 295, 1–26. [Google Scholar] [PubMed]
- Chakrabartty, A.; Baldwin, R.L. Stability of Alpha-Helices. Adv. Protein Chem. 1995, 46, 141–176. [Google Scholar] [PubMed]
- DeGrado, W.F.; Summa, C.M.; Pavone, V.; Nastri, F.; Lombardi, A. De novo design and structural characterization of proteins and metalloproteins. Annu. Rev. Biochem. 1999, 68, 779–819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, M.J.I.; Tabor, A.B. Forming stable helical peptides using natural and artificial amino acids. Tetrahedron 1999, 55, 11711–11743. [Google Scholar] [CrossRef]
- Venkatraman, J.; Shankaramma, S.C.; Balaram, P. Design of folded peptides. Chem. Rev. 2001, 101, 3131–3152. [Google Scholar] [CrossRef] [PubMed]
- Makhatadze, G.I. Thermodynamics Of alpha-Helix Formation. Adv. Protein Chem. 2005, 72, 199–226. [Google Scholar] [PubMed]
- Aurora, R.; Rose, G.D. Helix capping. Protein Sci. 1998, 7, 21–38. [Google Scholar] [CrossRef] [PubMed]
- Serrano, L.; Fersht, A.R. Capping and alpha-helix stability. Nature 1989, 342, 296–299. [Google Scholar] [CrossRef] [PubMed]
- D’Andrea, L.D.; Iaccarino, G.; Fattorusso, R.; Sorriento, D.; Carannante, C.; Capasso, D.; Trimarco, B.; Pedone, C. Targeting angiogenesis: Structural characterization and biological properties of a de novo engineered VEGF mimicking peptide. Proc. Natl. Acad. Sci. USA 2005, 102, 14215–14220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finetti, F.; Basile, A.; Capasso, D.; Di Gaetano, S.; Di Stasi, R.; Pascale, M.; Turco, C.M.; Ziche, M.; Morbidelli, L.; D’Andrea, L.D. Functional and pharmacological characterization of a VEGF mimetic peptide on reparative angiogenesis. Biochem. Pharmacol. 2012, 84, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Dudar, G.K.; D’Andrea, L.D.; Di Stasi, R.; Pedone, C.; Wallace, J.L. A vascular endothelial growth factor mimetic accelerates gastric ulcer healing in an iNOS-dependent manner. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G374–G381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pignataro, G.; Ziaco, B.; Tortiglione, A.; Gala, R.; Cuomo, O.; Vinciguerra, A.; Lapi, D.; Mastantuono, T.; Anzilotti, S.; D’Andrea, L.D.; et al. Neuroprotective Effect of VEGF-Mimetic Peptide QK in Experimental Brain Ischemia Induced in Rat by Middle Cerebral Artery Occlusion. ACS Chem. Neurosci. 2015, 6, 1517–1525. [Google Scholar] [CrossRef] [PubMed]
- Santulli, G.; Ciccarelli, M.; Palumbo, G.; Campanile, A.; Galasso, G.; Ziaco, B.; Altobelli, G.G.; Cimini, V.; Piscione, F.; D’Andrea, L.D.; et al. In vivo properties of the proangiogenic peptide QK. J. Transl. Med. 2009, 7, 41. [Google Scholar] [CrossRef] [Green Version]
- Diana, D.; Ziaco, B.; Scarabelli, G.; Pedone, C.; Colombo, G.; D’Andrea, L.D.; Fattorusso, R. Structural Analysis of a Helical Peptide Unfolding Pathway. Chem. Eur. J. 2010, 16, 5400–5407. [Google Scholar] [CrossRef] [PubMed]
- Diana, D.; Ziaco, B.; Colombo, G.; Scarabelli, G.; Romanelli, A.; Fedone, C.; Fattorusso, R.; D’Andrea, L.D. Structural determinants of the unusual helix stability of a De Novo engineered vascular endothelial growth factor (VEGF) mimicking peptide. Chem. Eur. J. 2008, 14, 4164–4166. [Google Scholar] [CrossRef] [PubMed]
- Ziaco, B.; Diana, D.; Capasso, D.; Palumbo, R.; Celentano, V.; Di Stasi, R.; Fattorusso, R.; D’Andrea, L.D. C-terminal truncation of Vascular Endothelial Growth Factor mimetic helical peptide preserves structural and receptor binding properties. Biochem. Biophys. Res. Commun. 2012, 424, 290–294. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, L.; Diana, D.; Di Stasi, R.; Romanelli, A.; Sciacca, M.F.M.; Milardi, D.; Isernia, C.; Fattorusso, R.; D’Andrea, L.D. Probing the helical stability in a VEGF-mimetic peptide. Bioorganic Chem. 2021, 116, 105379. [Google Scholar] [CrossRef]
- Di Stasi, R.; Diana, D.; Capasso, D.; Di Gaetano, S.; De Rosa, L.; Celentano, V.; Isernia, C.; Fattorusso, R.; D’Andrea, L.D. VEGFR Recognition Interface of a Proangiogenic VEGF-Mimetic Peptide Determined In Vitro and in the Presence of Endothelial Cells by NMR Spectroscopy. Chemistry 2018, 24, 11461–11466. [Google Scholar] [CrossRef]
- D’Andrea, L.D.; Del Gatto, A.; De Rosa, L.; Romanelli, A.; Pedone, C. Peptides targeting angiogenesis related growth factor receptors. Curr. Pharm. Des. 2009, 15, 2414–2429. [Google Scholar] [CrossRef]
- De Rosa, L.; Di Stasi, R.; D’Andrea, L.D. Pro-angiogenic peptides in biomedicine. Arch. Biochem. Biophys. 2018, 660, 72–86. [Google Scholar] [CrossRef]
- D’Andrea, L.D.; De Rosa, L.; Vigliotti, C.; Cataldi, M. VEGF mimic peptides: Potential applications in central nervous system therapeutics. New Horiz. Transl. Med. 2017, 3, 233–251. [Google Scholar]
- Basile, A.; Del Gatto, A.; Diana, D.; Di Stasi, R.; Falco, A.; Festa, M.; Rosati, A.; Barbieri, A.; Franco, R.; Arra, C.; et al. Characterization of a Designed Vascular Endothelial Growth Factor Receptor Antagonist Helical Peptide with Antiangiogenic Activity in Vivo. J. Med. Chem. 2011, 54, 1391–1400. [Google Scholar] [CrossRef] [PubMed]
- Diana, D.; Basile, A.; De Rosa, L.; Di Stasi, R.; Auriemma, S.; Arra, C.; Pedone, C.; Turco, M.C.; Fattorusso, R.; D’Andrea, L.D. beta-Hairpin Peptide That Targets Vascular Endothelial Growth Factor (Vegf) Receptors Design, Nmr Characterization, and Biological Activity. J. Biol. Chem. 2011, 286, 41680–41691. [Google Scholar] [CrossRef] [Green Version]
- De Rosa, L.; Diana, D.; Basile, A.; Russomanno, A.; Isernia, C.; Turco, M.C.; Fattorusso, R.; D’Andrea, L.D. Design, structural and biological characterization of a VEGF inhibitor beta-hairpin-constrained peptide. Eur. J. Med. Chem. 2014, 73, 210–216. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, L.; Finetti, F.; Diana, D.; Di Stasi, R.; Auriemma, S.; Romanelli, A.; Fattorusso, R.; Ziche, M.; Morbidelli, L.; D’Andrea, L.D. Miniaturizing VEGF: Peptides mimicking the discontinuous VEGF receptor-binding site modulate the angiogenic response. Sci. Rep. 2016, 6, 31295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capasso, D.; Di Gaetano, S.; Celentano, V.; Diana, D.; Festa, L.; Di Stasi, R.; De Rosa, L.; Fattorusso, R.; D’Andrea, L.D. Unveiling a VEGF-mimetic peptide sequence in the IQGAP1 protein. Mol. Biosyst. 2017, 13, 1619–1629. [Google Scholar] [CrossRef]
- De Rosa, L.; Capasso, D.; Diana, D.; Stefania, R.; Di Stasi, R.; Fattorusso, R.; D’Andrea, L.D. Metabolic and conformational stabilization of a VEGF-mimetic beta-hairpin peptide by click-chemistry. Eur. J. Med. Chem. 2021, 222, 113575. [Google Scholar] [CrossRef]
- Diana, D.; Russomanno, A.; De Rosa, L.; Di Stasi, R.; Capasso, D.; Di Gaetano, S.; Romanelli, A.; Russo, L.; D’Andrea, L.D.; Fattorusso, R. Functional binding surface of a beta-hairpin VEGF receptor targeting peptide determined by NMR spectroscopy in living cells. Chemistry 2015, 21, 91–95. [Google Scholar] [CrossRef]
- Wiesmann, C.; Fuh, G.; Christinger, H.W.; Eigenbrot, C.; Wells, J.A.; de Vos, A.M. Crystal structure at 1.7 A resolution of VEGF in complex with domain 2 of the Flt-1 receptor. Cell 1997, 91, 695–704. [Google Scholar] [CrossRef] [Green Version]
- Greenfield, N.J. Using circular dichroism spectra to estimate protein secondary structure. Nat. Protoc. 2006, 1, 2876–2890. [Google Scholar] [CrossRef]
- Luo, P.Z.; Baldwin, R.L. Mechanism of helix induction by trifluoroethanol: A framework for extrapolating the helix-forming properties of peptides from trifluoroethanol/water mixtures back to water. Biochemistry 1997, 36, 8413–8421. [Google Scholar] [CrossRef] [PubMed]
- Rohl, C.A.; Chakrabartty, A.; Baldwin, R.L. Helix propagation and N-cap propensities of the amino acids measured in alanine-based peptides in 40 volume percent trifluoroethanol. Protein Sci. 1996, 5, 2623–2637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wishart, D.S.; Nip, A.M. Protein chemical shift analysis: A practical guide. Biochem. Cell. Biol. 1998, 76, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Pimenta, J.; Viegas, A.; Sardinha, J.; Martins, I.C.; Cabrita, E.J.; Fontes, C.M.; Prates, J.A.; Pereira, R.M. NMR solution structure and SRP54M predicted interaction of the N-terminal sequence (1-30) of the ovine Doppel protein. Peptides 2013, 49, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Doig, A.J.; Baldwin, R.L. N- and C-Capping Preferences for All 20 Amino-Acids in Alpha-Helical Peptides. Protein Sci. 1995, 4, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Petukhov, M.; Yumoto, N.; Murase, S.; Onmura, R.; Yoshikawa, S. Factors that affect the stabilization of alpha-helices in short peptides by a capping box. Biochemistry 1996, 35, 387–397. [Google Scholar] [CrossRef] [PubMed]
- Durani, S. Protein Design with L- and D-alpha-Amino Acid Structures as the Alphabet. Acc. Chem. Res. 2008, 41, 1301–1308. [Google Scholar] [CrossRef]
- Simon, M.D.; Maki, Y.; Vinogradov, A.A.; Zhang, C.; Yu, H.T.; Lin, Y.S.; Kajihara, Y.; Pentelute, B.L. D-Amino Acid Scan of Two Small Proteins. J. Am. Chem. Soc. 2016, 138, 12099–12111. [Google Scholar] [CrossRef]
- Scarborough, R.M.; Mcenroe, G.A.; Arfsten, A.; Kang, L.L.; Schwartz, K.; Lewicki, J.A. D-Amino Acid-Substituted Atrial Natriuretic Peptide Analogs Reveal Novel Receptor Recognition Requirements. J. Biol. Chem. 1988, 263, 16818–16822. [Google Scholar] [CrossRef]
- Hong, S.Y.; Oh, J.E.; Lee, K.H. Effect of D-amino acid substitution on the stability, the secondary structure, and the activity of membrane-active peptide. Biochem. Pharmacol. 1999, 58, 1775–1780. [Google Scholar] [CrossRef]
- Bang, D.; Gribenko, A.V.; Tereshko, V.; Kossiakoff, A.A.; Kent, S.B.; Makhatadze, G.I. Dissecting the energetics of protein alpha-helix C-cap termination through chemical protein synthesis. Nat. Chem. Biol. 2006, 2, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Harper, E.T.; Rose, G.D. Helix Stop Signals in Proteins and Peptides—The Capping Box. Biochemistry 1993, 32, 7605–7609. [Google Scholar] [CrossRef] [PubMed]
- Seale, J.W.; Srinivasan, R.; Rose, G.D. Sequence Determinants of the Capping Box, a Stabilizing Motif at the N-Termini of Alpha-Helices. Protein Sci. 1994, 3, 1741–1745. [Google Scholar] [CrossRef] [PubMed]
- Towse, C.L.; Hopping, G.; Vulovic, I.; Daggett, V. Nature versus design: The conformational propensities of d-amino acids and the importance of side chain chirality. Protein Eng. Des. Sel. 2014, 27, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griesinger, C.; Otting, G.; Wuthrich, K.; Ernst, R.R. Clean Tocsy for H-1 Spin System-Identification in Macromolecules. J. Am. Chem. Soc. 1988, 110, 7870–7872. [Google Scholar] [CrossRef]
- Kumar, A.; Ernst, R.R.; Wuthrich, K. A two-dimensional nuclear Overhauser enhancement (2D NOE) experiment for the elucidation of complete proton-proton cross-relaxation networks in biological macromolecules. Biochem. Biophys. Res. Commun. 1980, 95, 1–6. [Google Scholar] [CrossRef]
- Piantini, U.; Sorensen, O.W.; Ernst, R.R. Multiple quantum filters for elucidating NMR coupling networks. J. Am. Chem. Soc. 1982, 104, 6800–6801. [Google Scholar] [CrossRef]
- Hwang, T.L.; Shaka, A.J. Water Suppression That Works—Excitation Sculpting Using Arbitrary Wave-Forms and Pulsed-Field Gradients. J. Magn. Reason. Ser. A 1995, 112, 275–279. [Google Scholar] [CrossRef]
- Wüthrich, K. NMR of Proteins and Nucleic Acids; Wiley: New York, NY, USA, 1986. [Google Scholar]
- Goddard, T.D.; Kneller, D.G. SPARKY 3; University of California: San Francisco, CA, USA, 2004. [Google Scholar]
- Bartels, C.; Xia, T.H.; Billeter, M.; Guntert, P.; Wuthrich, K. The program XEASY for computer-supported NMR spectral analysis of biological macromolecules. J. Biomol. NMR 1995, 6, 1–10. [Google Scholar] [CrossRef]
- Kjaergaard, M.; Brander, S.; Poulsen, F.M. Random coil chemical shift for intrinsically disordered proteins: Effects of temperature and pH. J. Biomol. NMR 2011, 49, 139–149. [Google Scholar] [CrossRef]
- Wishart, D.S.; Sykes, B.D. Chemical shifts as a tool for structure determination. Methods Enzymol. 1994, 239, 363–392. [Google Scholar] [PubMed]
- Partida-Hanon, A.; Trevino, M.A.; Mompean, M.; Jimenez, M.A.; Bruix, M. Structural insight into the XTACC3/XMAP215 interaction from CD and NMR studies on model peptides. Biopolymers 2017, 107, e23039. [Google Scholar] [CrossRef] [PubMed]
- Guntert, P. Automated NMR structure calculation with CYANA. Methods Mol. Biol. 2004, 278, 353–378. [Google Scholar] [PubMed]
- Koradi, R.; Billeter, M.; Wuthrich, K. MOLMOL: A program for display and analysis of macromolecular structures. J. Mol. Graph. 1996, 14, 51–55. [Google Scholar] [CrossRef]
- Schrödinger, L.; DeLano, W. PyMOL. Available online: http://www.pymol.org/pymol (accessed on 28 September 2022).







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Rosa, L.; Diana, D.; Capasso, D.; Stefania, R.; Di Stasi, R.; Fattorusso, R.; D’Andrea, L.D. Switching the N-Capping Region from all-L to all-D Amino Acids in a VEGF Mimetic Helical Peptide. Molecules 2022, 27, 6982. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27206982
De Rosa L, Diana D, Capasso D, Stefania R, Di Stasi R, Fattorusso R, D’Andrea LD. Switching the N-Capping Region from all-L to all-D Amino Acids in a VEGF Mimetic Helical Peptide. Molecules. 2022; 27(20):6982. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27206982
Chicago/Turabian StyleDe Rosa, Lucia, Donatella Diana, Domenica Capasso, Rachele Stefania, Rossella Di Stasi, Roberto Fattorusso, and Luca Domenico D’Andrea. 2022. "Switching the N-Capping Region from all-L to all-D Amino Acids in a VEGF Mimetic Helical Peptide" Molecules 27, no. 20: 6982. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27206982