
Combination Therapy of Carnosic Acid and Methotrexate Effectively Suppressed the Inflammatory Markers and Oxidative Stress in Experimental Arthritis
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Change in Body Weight
2.2. Hind Paw Volume
2.3. Plasmatic Interleukin-17A
2.4. Matrix Metalloproteinase-9 in Plasma
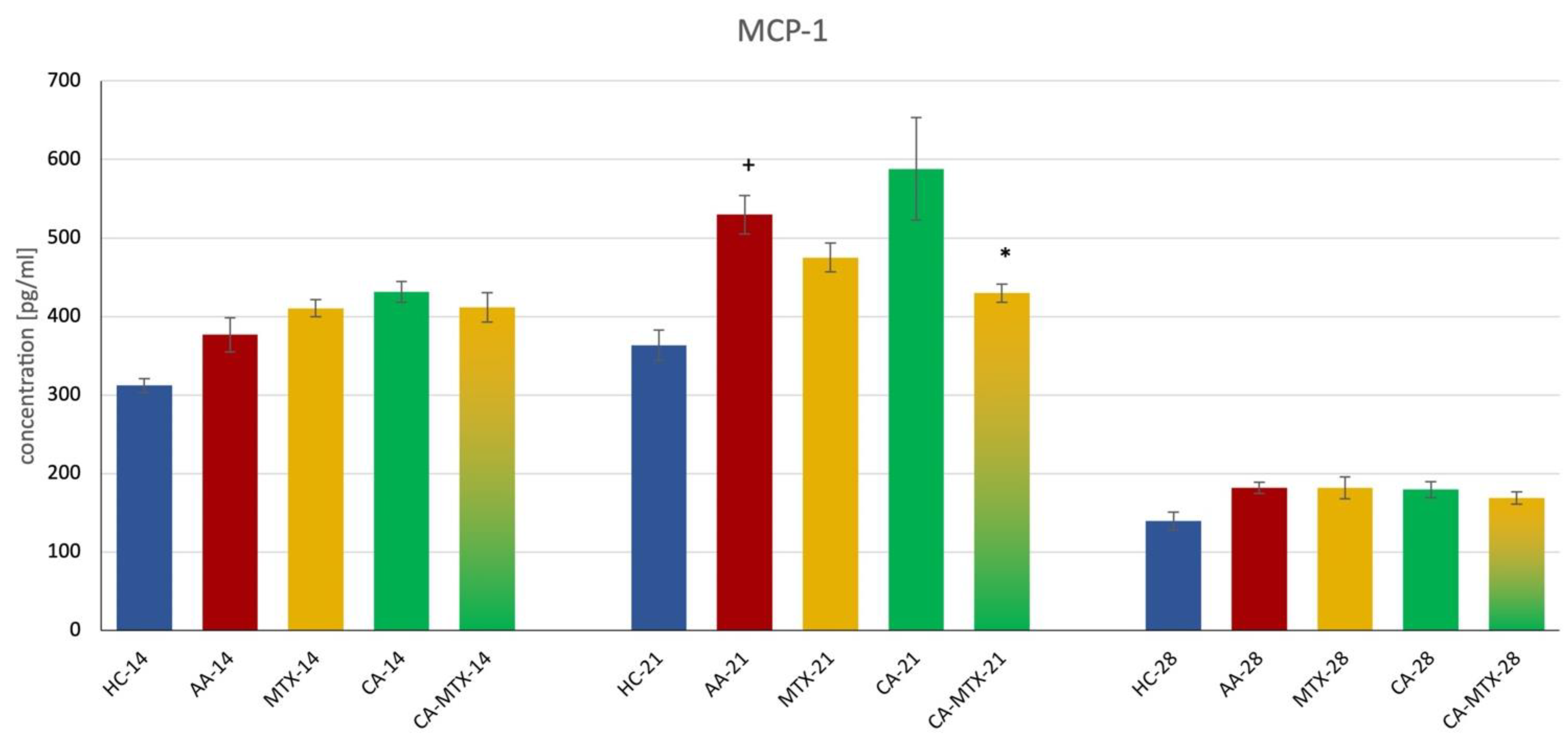
2.5. Monocyte Chemotactic Protein-1 in Plasma
2.6. Activity of Gamma-Glutamyl Transferase in Joint
2.7. Relative mRNA Expression of Interleukin-1β and Antioxidant Enzymes Heme Oxygenase (HO-1) and Catalase (CAT)
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Induction of Adjuvant Arthritis in Lewis Rats
4.3. Experimental Design and Treatment
4.4. Change in Body Weight
4.5. Hind Paws’ Volume Change
4.6. Markers of Inflammation
4.7. The Activity of the Gamma-Glutamyltransferase in Joint
4.8. RNA Extraction and Reverse Transcription
4.9. Quantitative Real-Time RT-PCR
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Almutairi, K.B.; Nossent, J.C.; Preen, D.B.; Keen, H.I.; Inderjeeth, C.A. The Prevalence of Rheumatoid Arthritis: A Systematic Review of Population-based Studies. J. Rheumatol. 2021, 48, 669–676. [Google Scholar] [CrossRef] [PubMed]
- Mateen, S.; Zafar, A.; Moin, S.; Khan, A.Q.; Zubair, S. Understanding the role of cytokines in the pathogenesis of rheumatoid arthritis. Clin. Chim. Acta 2016, 455, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Rahman, I.; Bahgi, D. Inflammation. Advancing Age and Nutrition, 1st ed.; Academic Press: Cambridge, MA, USA; Elsevier: London, UK, 2014; pp. 215–225. [Google Scholar]
- Van der Heide, A.; Jacobs, J.W.; Bijlsma, J.W.; Heurkens, A.H.; van Booma-Frankfort, C.; van der Veen, M.J.; Haanen, H.C.; Hofman, D.M.; van Albada-Kuipers, G.A.; ter Borg, E.J.; et al. The effectiveness of early treatment with “second-line” antirheumatic drugs. A randomized, controlled trial. Ann. Intern. Med. 1996, 124, 699–707. [Google Scholar] [CrossRef]
- Chetan, G.; Shinde, M.P.; Venkatesh, T.M.; Kumar, P.; Shivakumar, H.G. Methotrexate: A Gold Standard for Treatment of Rheumatoid Arthritis. J. Pain Palliat. Care Pharmacother. 2014, 28, 351–358. [Google Scholar] [CrossRef]
- Salliot, C.; van der Heijde, D. Long-term safety of methotrexate monotherapy in patients with rheumatoid arthritis: A systematic literature research. Ann. Rheum. Dis. 2009, 68, 1100–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cronstein, B.N.; Aune, T.M. Methotrexate and its mechanisms of action in inflammatory arthritis. Nat. Rev. Rheumatol. 2020, 16, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Smolen, J.S.; Landewé, R.; Breedveld, F.C.; Dougados, M.; Emery, P.; Gaujoux-Viala, C.; Gorter, S.; Knevel, R.; Nam, J.; Schoels, M.; et al. EULAR recommendations for the management of rheumatoid arthritis with synthetic and biological disease-modifying antirheumatic drugs. Ann. Rheum. Dis. 2010, 69, 964–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubbert-Roth, A.; Finckh, A. Treatment options in patients with rheumatoid arthritis failing initial TNF inhibitor therapy: A critical review. Arthritis Res. Ther. 2009, 11, S1. [Google Scholar] [CrossRef] [Green Version]
- Pope, J.; Sawant, R.; Tundia, N.; Du, E.X.; Qi, C.Z.; Song, Y.; Tang, P.; Betts, K.A. Comparative Efficacy of JAK Inhibitors for Moderate-To-Severe Rheumatoid Arthritis: A Network Meta-Analysis. Adv. Ther. 2020, 37, 2356–2372. [Google Scholar] [CrossRef] [Green Version]
- Smolen, J. Response to: “Concerns on Glucocorticoid Use for Japanese Patients with Established Rheumatoid Arthritis” by Oiwa. Ann. Rheum. Dis. 2018, 17, e36. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, N.; Bhatt, L.K.; Prabhavalkar, K.S. Experimental animal models for rheumatoid arthritis. Immunopharmacol. Immunotoxicol. 2018, 40, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Roy, T.; Ghosh, S. Animal models of rheumatoid arthritis: Correlation and usefulness with human rheumatoid arthritis. Indo Amer. J. Pharm. Res. 2013, 3, 6131–6142. [Google Scholar]
- Stolina, M.; Bolon, B.; Middleton, S.; Dwyer, D.; Brown, H.; Duryea, D.; Zhu, L.; Rohner, A.; Pretorius, J.; Kostenuik, P.; et al. The evolving systemic and local biomarker milieu at different stages of disease progression in rat adjuvant-induced arthritis. J. Clin. Immunol. 2009, 29, 158–174. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Yang, W.S.; Kim, C.H. Physiological Properties, Functions, and Trends in the Matrix Metalloproteinase Inhibitors in Inflammation-Mediated Human Diseases. Curr. Med. Chem. 2022. [Google Scholar] [CrossRef] [PubMed]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Ayala, J.M.; Goyal, S.; Liverton, N.J.; Claremon, D.A.; O’Keefe, S.J.; Hanlon, W.A. Serum-induced monocyte differentiation and monocyte chemotaxis are regulated by the p38 MAP kinase signal transduction pathway. J. Leukoc. Biol. 2000, 67, 869–875. [Google Scholar] [CrossRef]
- Boring, L.; Gosling, J.; Chensue, S.W.; Kunkel, S.L.; Farese, R.V.; Broxmeyer, H.E., Jr.; Charo, I.F. Impaired monocyte migration and reduced type 1 (Th1) cytokine responses in C-C chemokine receptor 2 knockout mice. J. Clin. Investig. 1997, 100, 2552–2561. [Google Scholar] [CrossRef]
- Volin, M.V.; Shah, M.R.; Tokuhira, M.; Haines, G.K.; Woods, J.M.; Koch, A.E. RANTES expression and contribution to monocyte chemotaxis in arthritis. Clin. Immunol. Immunopathol. 1998, 89, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Ulrich-Merzenich, G.S. Combination screening of synthetic drugs and plant derived natural products—Potential and challenges for drug development. Synergy 2014, 1, 59–69. [Google Scholar] [CrossRef]
- Birtić, S.; Dussort, P.; Pierre, F.-X.; Bily, A.C.; Roller, M. Carnosic acid. Phytochemistry 2015, 115, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Li, J.; Ning, F.; Wijaya, W.; Chen, Y.; Xiao, J.; Huang, Q. Improving in vitro bioaccessibility and bioactivity of carnosic acid using a lecithin-based nanoemulsion system. Food Funct. 2021, 12, 1558–1568. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Smuts, J.P.; Dodbiba, E.; Rangarajan, R.; Lang, J.C.; Armstrong, D.W. Degradation study of carnosic acid, carnosol, rosmarinic acid, and rosemary extract (Rosmarinus officinalis L.) assessed using HPLC. J. Agric. Food Chem. 2012, 60, 9305–9314. [Google Scholar] [CrossRef] [PubMed]
- Koutsoulas, A.; Čarnecká, M.; Slanina, J.; Tóth, J.; Slaninová, I. Characterization of Phenolic Compounds and Antiproliferative Effects of Salvia pomifera and Salvia fruticosa Extracts. Molecules 2019, 24, 2921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abadi, M.N.A.; Mortazavi, M.; Kalani, N.; Marzouni, H.Z.; Kooti, W.; Ali-Akbari, S. Effect of hydroalcoholic extract of Rosmarinus officinalis L. leaf on anxiety in mice. Evid. Based Complement. Altern. Med. 2016, 21, NP85–NP90. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Sedighi, R.; Wang, P.; Chen, H.; Zhu, Y.; Sang, S. Carnosic acid as a major bioactive component in rosemary extract ameliorates high-fat-diet-induced obesity and metabolic syndrome in mice. J. Agric. Food Chem. 2015, 63, 4843–4852. [Google Scholar] [CrossRef] [PubMed]
- Erkan, N.; Ayranci, G.; Ayranci, E. Antioxidant activities of rosemary (Rosmarinus Officinalis L.) extract, blackseed (Nigella sativa L.) essential oil, carnosic acid, rosmarinic acid and sesamol. Food Chem. 2008, 110, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Yanagitai, M.; Itoh, S.; Kitagawa, T.; Takenouchi, T.; Kitani, H.; Satoh, T. Carnosic acid, a pro-electrophilic compound, inhibits LPS-induced activation of microglia. Biochem. Biophys. Res. Commun. 2012, 418, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhao, L.; Han, J.J.; Zhang, F.; Liu, S.; Zhu, L.; Wang, Z.Z.; Zhang, G.X.; Zhang, Y. Carnosol Modulates Th17 Cell Differentiation and Microglial Switch in Experimental Autoimmune Encephalomyelitis. Front Immunol. 2018, 9, 1807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khella, K.F.; el Maksoud, A.I.A.; Hassan, A.; Abdel-Ghany, S.E.; Elsanhoty, R.M.; Aladhadh, M.A.; Abdel-Hakeem, M.A. Carnosic Acid Encapsulated in Albumin Nanoparticles Induces Apoptosis in Breast and Colorectal Cancer Cells. Molecules 2022, 27, 4102. [Google Scholar] [CrossRef] [PubMed]
- Den Hartogh, D.J.; Vlavcheski, F.; Giacca, A.; MacPherson, R.E.K.; Tsiani, E. Carnosic Acid Attenuates the Free Fatty Acid-Induced Insulin Resistance in Muscle Cells and Adipocytes. Cells 2022, 11, 167. [Google Scholar] [CrossRef] [PubMed]
- Growth Chart. Available online: https://www.criver.com/products-services/find-model/lewis-rat?region=3616#panel1-growth-chart (accessed on 12 October 2022).
- Burmester, G.; Feist, E.; Dörner, T. Emerging cell and cytokine targets in rheumatoid arthritis. Nat. Rev. Rheumatol. 2014, 10, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Chaubey, P.; Suvarna, V. Role of natural products in alleviation of rheumatoid arthritis—A review. J. Food Biochem. 2021, 45, e13673. [Google Scholar] [CrossRef] [PubMed]
- Kunsch, C.; Sikorski, J.A.; Sundell, C.L. Oxidative stress and the use of antioxidants for the treatment of rheumatoid arthritis. Curr. Med. Chem. Immunol. Endoc. Metab. Agents 2005, 5, 249–258. [Google Scholar] [CrossRef]
- Bauerová, K.; Bezek, A. Role of reactive oxygen and nitrogen species in etiopathogenesis of rheumatoid arthritis. Gen. Physiol. Biophys. 1999, 18, 15–20. [Google Scholar] [PubMed]
- Kundu, S.; Ghosh, P.; Datta, S.; Ghosh, A.; Chattopadhyay, S.; Chatterjee, M. Oxidative stress as a potential biomarker for determining disease activity in patients with rheumatoid arthritis. Free Radic. Res. 2012, 46, 1482–1489. [Google Scholar] [CrossRef] [PubMed]
- Stamp, L.K.; Khalilova, I.; Tarr, J.M.; Senthilmohan, R.; Turner, R.; Haigh, R.C.; Winyard, P.G.; Kettle, A.J. Myeloperoxidase and oxidative stress in rheumatoid arthritis. Rheumatology 2012, 51, 1796–1803. [Google Scholar] [CrossRef] [Green Version]
- Rovenský, J.; Stancíková, M.; Svík, K.; Utesený, J.; Bauerová, K.; Jurcovicová, J. Treatment of adjuvant-induced arthritis with the combination of methotrexate and probiotic bacteria Escherichia coli O83 (Colinfant). Folia Microbiol. 2009, 54, 359–363. [Google Scholar] [CrossRef] [PubMed]
- Refaat, R.; Salama, M.; Meguid, E.A.; el Sarha, A.; Gowayed, M. Evaluation of the effect of losartan and methotrexate combined therapy in adjuvant-induced arthritis in rats. Eur. J. Pharmacol. 2013, 698, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Van Eden, W.; Wage-naar-Hilbers, J.P.; Wauben, M.H. Adjuvant arthritis in the rat. Curr. Protoc. Immunol. 2001, 15, 15.4. [Google Scholar] [CrossRef]
- Tsiklauri, L.; Švík, K.; Chrastina, M.; Poništ, S.; Dráfi, F.; Slovák, L.; Alania, M.; Kemertelidze, E.; Bauerova, K. Bio-flavonoid Robinin from Astragalus falcatus Lam. Mildly Improves the Effect of Metothrexate in Rats with Adjuvant Arthritis. Nutrients 2021, 13, 1268. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.; Wang, X.; Sun, H.; Qin, Y.; Fu, M. Carnosic acid (CA) attenuates collagen-induced arthritis in db/db mice via inflammation suppression by regulating ROS-dependent p38 pathway. Free Radic. Biol. Med. 2017, 108, 418–432. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Zhou, X.; Zhou, L.; Liu, Z.; Yuan, J.; Cheng, J.; Zhao, J.; Wu, L.; Li, H.; Qiu, H.; et al. Carnosic acid inhibits inflammation response and joint destruction on osteoclasts, fibroblast-like synoviocytes, and collagen-induced arthritis rats. J. Cell. Physiol. 2018, 233, 6291–6303. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, A.; Cases, J.; Roller, M.; Chiralt-Boix, A.; Coussaert, A.; Ripoll, C. Carnosic acid-rich rosemary (Rosmarinus officinalis L.) leaf extract limits weight gain and improves cholesterol levels and glycaemia in mice on a high-fat diet. Br. J. Nutr. 2011, 106, 1182–1189. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Takikawa, Y.; Satoh, T.; Yoshioka, Y.; Kosaka, K.; Tatemichi, Y.; Suzuki, K. Carnosic acid prevents obesity and hepatic steatosis in ob/ob mice. Hepatol. Res. 2011, 41, 87–92. [Google Scholar] [CrossRef]
- Santo, R.C.E.; Fernandes, K.Z.; Lora, P.S.; Filippin, L.I.; Xavier, R.M. Prevalence of rheumatoid cachexia in rheumatoid arthritis: A systematic review and meta-analysis. J. Cachexia Sarcopenia Muscle 2018, 9, 816–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, C.C.; Chen, S.J.; Adamopoulos, I.E.; Davis, N.; Hong, K.; Vu, A.; Kwan, S.; Fayadat-Dilman, L.; Asio, A.; Bowman, E.P. Anti-IL-17A therapy protects against bone erosion in experimental models of rheumatoid arthritis. Autoimmunity 2011, 44, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.L.; Li, H.; Li, X.X.; Cui, C.Y.; Wang, R.; Yu, N.X.; Chen, L.X. Acute and 30-day oral toxicity studies of administered carnosic acid. Food Chem. Toxicol. 2012, 50, 4348–4355. [Google Scholar] [CrossRef] [PubMed]
- Chae, I.G.; Yu, M.H.; Im, N.K.; Jung, Y.T.; Lee, J.; Chun, K.S.; Lee, I.S. Effect of Rosemarinus Officinalis L. on MMP-9, MCP-1 levels, and cell migration in RAW 264.7 and smooth muscle cells. J. Med. Food 2012, 15, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.M.; Lin, H.C.; Chang, W.C. Carnosic acid prevents the migration of human aortic smooth muscle cells by inhibiting the activation and expression of matrix metalloproteinase-9. Br. J. Nutr. 2008, 100, 731–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiedorczyk, M.; Klimiuk, P.A.; Sierakowski, S.; Gindzienska-Sieskiewicz, E.; Chwiecko, J. Serum matrix metalloproteinases and tissue inhibitors of metalloproteinases in patients with early rheumatoid arthritis. J. Rheumatol. 2006, 33, 1523–1529. [Google Scholar] [PubMed]
- Hosaka, S.; Akahoshi, T.; Wada, C.; Kondo, H. Expression of the chemokine superfamily in rheumatoid arthritis. Clin. Exp. Immunol. 1994, 97, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Shahrara, S.; Proudfoot, A.E.; Park, C.C.; Volin, M.V.; Haines, G.K.; Woods, J.M.; Aikens, C.H.; Handel, T.M.; Pope, R.M. Inhibition of monocyte chemoattractant protein-1 ameliorates rat adjuvant-induced arthritis. J. Immunol. 2008, 180, 3447–3456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q.; Shen, Z.; Yu, H.; Lu, G.; Yu, Y.; Liu, X.; Zheng, P. Carnosic acid protects against acetaminophen-induced hepatotoxicity by potentiating Nrf2-mediated antioxidant capacity in mice. Korean J. Physiol. Pharmacol. 2016, 20, 15–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Takikawa, Y.; Tabuchi, T.; Satoh, T.; Kosaka, K.; Suzuki, K. Carnosic acid (CA) prevents lipid accumulation in hepatocytes through the EGFR/MAPK pathway. J. Gastroenterol. 2012, 47, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Jabbari, N.; Eftekhari, Z.; Roodbari, N.H.; Parivar, K. Evaluation of Encapsulated Eugenol by Chitosan Nanoparticles on the aggressive model of rheumatoid arthritis. Int. Immunopharmacol. 2020, 85, 106554. [Google Scholar] [CrossRef]
- Nielsen, M.A.; Lomholt, S.; Mellemkjaer, A.; Andersen, M.N.; Buckley, C.D.; Kragstrup, T.W. Responses to Cytokine Inhibitors Associated with Cellular Composition in Models of Immune-Mediated Inflammatory Arthritis. ACR Open Rheumatol. 2020, 2, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Kuncirova, V.; Ponist, S.; Mihalova, D.; Drafi, F.; Nosal, R.; Acquaviva, A.; Gardi, C.; Harmatha, J.; Hradkova, I.; Bauerova, K. N-feruloylserotonin in preventive combination therapy with methotrexate reduced inflammation in adjuvant arthritis. Fundam. Clin. Pharmacol. 2014, 28, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Tsiklauri, L.; Drafi, F.; Poništ, S.; Slovák, L.; Chrastina, M.; Švík, K.; Kemoklidze, Z.; Kemertelidze, E.; Bauerová, K. Study of anti-inflammatory activity of Fatsiphloginum™ (Fatsia japonica) and a new purified triterpene-rich extract of saponins (PS-551) in an experimental model of arthritis. Physiol. Res. 2019, 68, S75–S85. [Google Scholar] [CrossRef] [PubMed]
- Bauerova, K.; Paulovicova, E.; Mihalova, D.; Drafi, F.; Strosova, M.; Mascia, C.; Biasi, F.; Rovensky, J.; Kucharska, J.; Gvozdjakova, A.; et al. Combined methotrexate and coenzyme Q₁₀ therapy in adjuvant-induced arthritis evaluated using parameters of inflammation and oxidative stress. Acta Biochim. Pol. 2010, 57, 347–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauerová, K.; Ponist, S.; Ondrejicková, O.; Komendová, D.; Mihalová, D. Association between tissue gamma-glutamyl-transferase and clinical markers of adjuvant arthritis in Lewis rats. Neuro Endocrinol. Lett. 2006, 27, 172–175. [Google Scholar] [PubMed]
- Pašková, Ľ.; Kuncírová, V.; Poništ, S.; Mihálová, D.; Nosáľ, R.; Harmatha, J.; Hrádková, I.; Čavojský, T.; Bilka, F.; Šišková, K.; et al. Effect of N-Feruloylserotonin and Methotrexate on Severity of Experimental Arthritis and on Messenger RNA Expression of Key Proinflammatory Markers in Liver. J. Immunol. Res. 2016, 2016, 7509653. [Google Scholar] [CrossRef]
- Häupl, T.; Yahyawi, M.; Lübke, C.; Ringe, J.; Rohrlach, T.; Burmester, G.R.; Sittinger, M.; Kaps, C. Gene expression profiling of rheumatoid arthritis synovial cells treated with antirheumatic drugs. J. Biomol. Screen 2007, 12, 328–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateen, S.; Moin, S.; Khan, A.Q.; Zafar, A.; Fatima, N. Increased Reactive Oxygen Species Formation and Oxidative Stress in Rheumatoid Arthritis. PLoS ONE 2016, 11, e0152925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comar, J.F.; de Sá-Nakanishi, A.B.; de Oliveira, A.L.; Wendt, M.M.N.; Amado, C.A.B.; Iwamoto, E.L.I.; Peralta, R.M.; Bracht, A. Oxidative state of the liver of rats with adjuvant-induced arthritis. Free Radic. Biol. Med. 2013, 58, 144–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funes, S.C.; Rios, M.; Fernández-Fierro, A.; Covián, C.; Bueno, S.M.; Riedel, C.A.; Mackern-Oberti, J.P.; Kalergis, A.M. Naturally Derived Heme-Oxygenase 1 Inducers and Their Therapeutic Application to Immune-Mediated Diseases. Front. Immunol. 2020, 11, 1467. [Google Scholar] [CrossRef] [PubMed]
- Alam, J.; Cook, J.L. How many transcription factors does it take to turn on the heme oxygenase-1 gene? Am. J. Respir. Cell Mol. Biol. 2007, 36, 166–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahri, S.; Jameleddine, S.; Shlyonsky, V. Relevance of carnosic acid to the treatment of several health disorders: Molecular targets and mechanisms. Biomed. Pharmacother. 2016, 84, 569–582. [Google Scholar] [CrossRef]
- Ishitobi, H.; Sanada, Y.; Kato, Y.; Ikuta, Y.; Shibata, S.; Yamasaki, S.; Lotz, M.K.; Matsubara, K.; Miyaki, S.; Adachi, N. Carnosic acid attenuates cartilage degeneration through induction of heme oxygenase-1 in human articular chondrocytes. Eur. J. Pharmacol. 2018, 830, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bina, J.; Wilder, R.L. Animal models of rheumatoid arthritis. Mol. Med. Today 1999, 5, 367–369. [Google Scholar] [CrossRef]
- Ondrejickova, O.; Ziegelhoeffer, A.; Gabauer, I.; Sotnikova, R.; Styk, J.; Gibala, P.; Sedlak, J.; Horakova, L. Evaluation of ischemia-reperfusion injury by malondialdehyde, glutathione and gamma-glutamyltranspeptidase: Lack of specific localeffects in diverse parts of the dog heart following acute coronary occlusion. Cardioscience 1993, 4, 225–230. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Mean |
|---|---|
| HC | 11.46 ± 1.39 |
| AA | 20.94 ± 2.01 + |
| MTX | 12.48 ± 2.59 |
| CA | 17.88 ± 1.00 |
| CA-MTX | 10.95 ± 1.40 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chrastina, M.; Poništ, S.; Tóth, J.; Czigle, S.; Pašková, Ľ.; Vyletelová, V.; Švík, K.; Bauerová, K. Combination Therapy of Carnosic Acid and Methotrexate Effectively Suppressed the Inflammatory Markers and Oxidative Stress in Experimental Arthritis. Molecules 2022, 27, 7115. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27207115
Chrastina M, Poništ S, Tóth J, Czigle S, Pašková Ľ, Vyletelová V, Švík K, Bauerová K. Combination Therapy of Carnosic Acid and Methotrexate Effectively Suppressed the Inflammatory Markers and Oxidative Stress in Experimental Arthritis. Molecules. 2022; 27(20):7115. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27207115
Chicago/Turabian StyleChrastina, Martin, Silvester Poništ, Jaroslav Tóth, Szilvia Czigle, Ľudmila Pašková, Veronika Vyletelová, Karol Švík, and Katarína Bauerová. 2022. "Combination Therapy of Carnosic Acid and Methotrexate Effectively Suppressed the Inflammatory Markers and Oxidative Stress in Experimental Arthritis" Molecules 27, no. 20: 7115. https://0-doi-org.brum.beds.ac.uk/10.3390/molecules27207115